1. Introduction
In 2006, New Zealand (NZ) had the highest reported rates of Campylobacteriosis in the developed world at approximately 385 per 100,000 of population [
1]. Subsequent interventions in the poultry industry occurred in the years following [
2] and the current rate has declined to 159 per 100,000 of population [
3]. Two species,
Campylobacter jejuni and
Campylobacter coli, are identified in the majority of human cases of campylobacteriosis and have been isolated from environmental waters, faeces of dairy cows, cattle, sheep, chicken and their derived food products [
4,
5]. Well known routes of infection include the consumption of undercooked poultry, dairy products and polluted drinking water [
6,
7,
8]. It has been difficult, however, to establish direct links between human cases and an animal reservoir, or transmission routes such as chicken meat or water [
6,
9,
10]. A review of campylobacteriosis cases in England and Wales has noted an increase in cases particularly between 2004 and 2011 with marked seasonal trends associated with multiple sources of
Campylobacter highlighting the difficulty of source attribution [
11].
Difficulties in source attribution have supported the assertion that most cases of campylobacteriosis are sporadic [
8,
12,
13]. There is concern, however, that at least some of these sporadic infections are not recognised as linked cases because not all of the available subtypes have been identified from either the clinical specimens or the suspected source [
14]. For example, in an outbreak of campylobacteriosis linked to undercooked chicken liver pâte, there were multiple strains of
C. jejuni belonging to four distinct clades isolated from the stool samples of cases [
15]. Co-infection with multiple
Campylobacter strains was documented in several of the cases and highlights the requirement for recognition of multiple infecting strains in outbreak cases. This study provides a counterexample to the “dominant outbreak strain” recognised for many foodborne outbreaks [
15].
Co-infection of human cases with more than one
C. jejuni subtype is not common, but might occur in up to 10% of campylobacteriosis cases [
5,
16,
17,
18]. In a review addressing co-infections by multiple strains of a pathogenic species, Balmer and Tanner suggested that the infrequency of notifications of multiple infections in a single host may change as studies increasingly apply molecular tools to identify clonal diversity [
19]. Identification of minor subtypes from chicken carcasses may contribute to knowledge about the association of particular
C. jejuni subtypes with that food and whether all subtypes identified in chickens contribute to human disease. Several studies have identified multiple clones of
Campylobacter in chickens [
20,
21] and a high degree of genotypic relatedness has been noted between different subtypes isolated from individual flocks [
22].
Due to the diversity of
Campylobacter subtypes, the isolation and typing of one colony from individual environmental samples may only be valid when testing large numbers of samples over an extended time period. When investigating epidemiologically-linked cases, however, multiple colonies from a suspected source may need to be typed to provide an accurate assessment of the presence of pathogenic subtypes [
14]. The distribution of
C. jejuni subtypes in the environment may be more accurately represented if numerically dominant and minor subtypes from the same sample are identified.
Dopfer
et al. addressed the issue of how many isolates of a bacterial species need to be analysed when assessing the genetic heterogeneity within an individual sample [
14]. Using Bayesian statistical methods, they determined that from 2-20 isolates per sample need to be characterised to ensure detection of all strains in a sample with 95% certainty. Studies of
E. coli subtypes in human faeces discussed the importance of multiple isolations from a single stool sample to identify both dominant and minor subtypes [
23]. They caution that basing a study on the isolation of numerically dominant subtypes of a bacterial species may exclude significant information, especially where the study is examining the frequency of virulence factors in a bacterial population. Schlager
et al. [
23] defined a dominant subtype as a clone which is represented by >50% of typed isolates in a sample. A minor subtype was defined as a clone that represented <10% of typed isolates in a sample. Studies of
E. coli clonal diversity in individual samples have used a binomial formula to determine the number of randomly selected colonies required to achieve a 90% probability of identifying a minor clone: 1 − (1 −
p)
n, where
p is the frequency of the minor clone and
n is the number of colonies tested [
23].
This current study presents subtyping data for multiple
C. jejuni colonies isolated from individual chicken carcasses using an enrichment method. Isolates were subjected to pulsed-field gel electrophoresis (PFGE) typing with the restriction enzymes (RE)
SmaI and
KpnI. The binomial formula of Schlager
et al. [
23] was applied to establish a statistical basis for identifying minor clones from a single sample. Up to 25 colonies per sample were isolated to give a 93% probability of identifying numerically minor subtypes. The aim was to determine whether all subtypes isolated from chicken carcasses are pathogenic to humans by comparing them to NZ human clinical subtypes of
C. jejuni collected nationwide. The data set from human cases included a subset of human isolates collected from the same geographical area and over the same timeframe as the chicken samples [
4]. In addition, the recognition of clonally related subtypes from the same sample and/or human case was recognised as potentially significant for the epidemiological tracking of outbreak sources.
2. Results and Discussion
2.1. Identification of Multiple Subtypes of C. jejuni in Chicken Carcasses
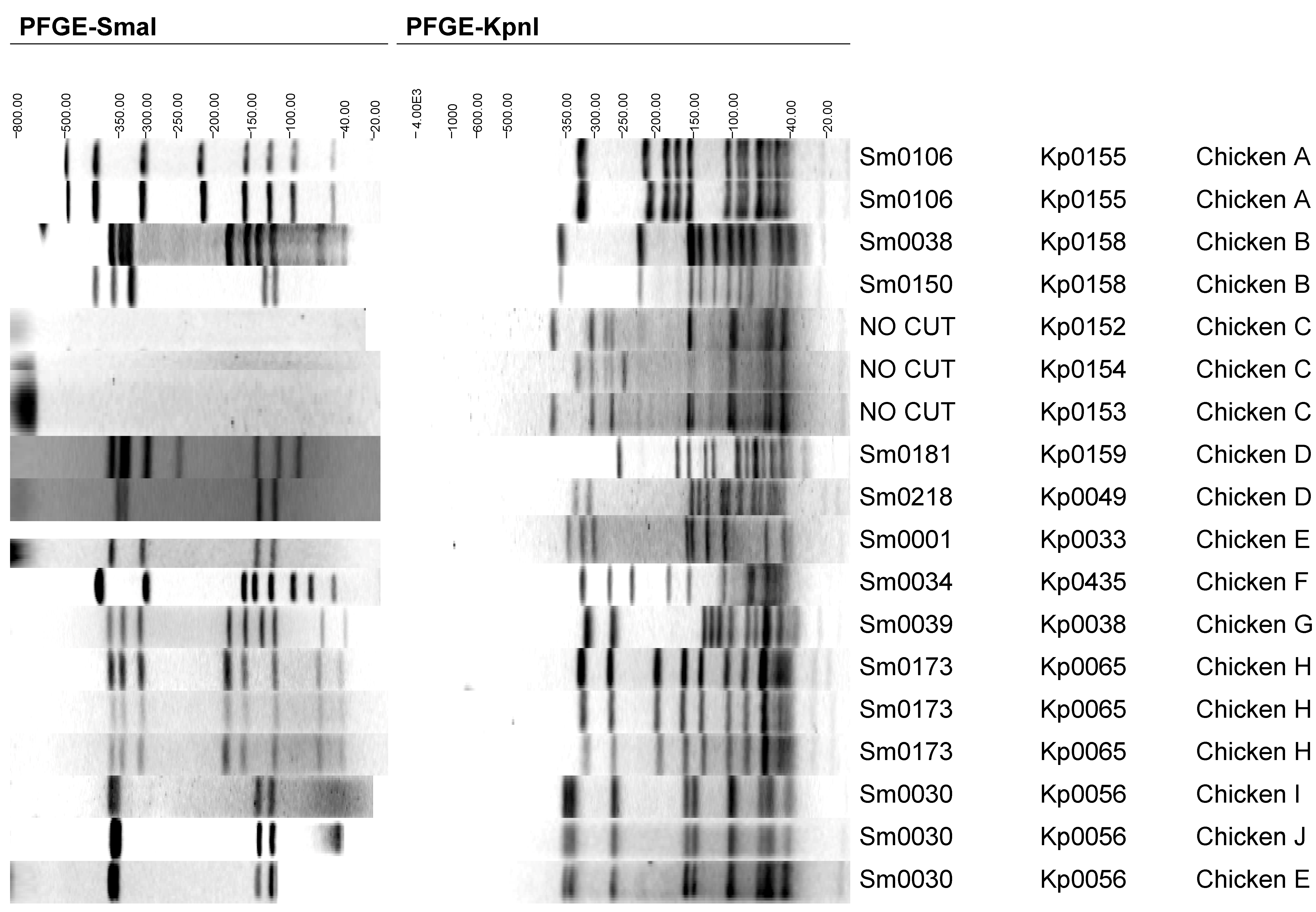
To address whether identification of minor subtypes in poultry increases the likelihood of identifying C. jejuni subtypes relevant to the human etiology of campylobacteriosis, we determined the diversity of subtypes on fresh chicken carcasses. Twenty-five isolates per carcass were collected; during storage however, several isolates were not recovered, reducing the number of isolates for some samples to less than 25 (range 21–25 isolates per sample). Purified isolates were Gram negative, oxidase, catalase and hippurate positive, grew microaerophilically at 42 °C but not aerobically at 25 °C, and were nalidixic acid susceptible and resistant to cephalothin, confirming their identification as C. jejuni. In total, 227 isolates were characterized by SmaI PFGE to identify subtypes of C. jejuni. To differentiate isolates further, 15-16 isolates per chicken carcass sample (total isolates n = 157) were subjected to KpnI digestion and electrophoresis.
Nine of the chicken carcasses carried one dominant subtype, represented by >50% of the typed isolates as defined by Schlager
et al. [
23]. One of the chickens (E) carried two subtypes with similar prevalences of approximately 50%, which were referred to as co-dominant. Thirteen distinguishable subtypes were identified from the ten carcasses based on the two RE profiles; four (40%) carried more than one subtype of
C. jejuni (
Table 1,
Figure 1). In these four chicken carcasses (B, C, D and E) carrying multiple subtypes, two subtypes were present in three carcasses and three subtypes present in one carcass. The percentages of minor and dominant subtypes identified in three (B, D and E) were based on the total number of isolates per sample because the
KpnI digestion did not differentiate the
SmaI subtype (
Table 1). Chicken C carried three subtypes that could not be digested with
SmaI RE but were resolved by
KpnI digestion, and therefore the percentage calculations for this sample were based on the number of isolates subjected to
KpnI digestion only (
n = 16).
Table 1.
Campylobacter jejuni subtypes isolated from chicken carcasses.
Table 1.
Campylobacter jejuni subtypes isolated from chicken carcasses.
| Chicken Sample | PFGE Subtype | Subtype Prevalence | Human Case Identity a | Identity to Other Sources a |
|---|
| A | Sm0106/Kp0155 | Single subtype | 1 | None |
| B | Sm0150/Kp0158 | Dominant (96%) | None | River water |
| | Sm0038/Kp0158 | Minor (4%) | None | None |
| C | NC b/Kp0152 | Dominant (81%) | None | None |
| | NC/Kp0153 | Intermediate (12%) | None | None |
| | NC/Kp0154 | Minor (6%) | None | None |
| D | Sm0218/Kp0049 | Dominant (77%) | 9 | Chicken meat, sheep faecal |
| | Sm0181/Kp0159 | Intermediate (23%) | None | Chicken meat |
| E | Sm0030/Kp0056 | Co-dominant (52%) | 3 c | Chicken meat |
| | Sm0001/Kp0033 | Co-dominant (48%) | 3 | Chick faeces |
| F | Sm0034/Kp0435 | Single subtype | 1 | None |
| G | Sm0039/Kp0038 | Single subtype | 32 | Dairy cow and chicken faeces, chicken meat, pig meat and sheep offal |
| H | Sm0173/Kp0065 | Single subtype | 1 | None |
| I | Sm0030/Kp0056 | Single subtype | 3 c | Chicken meat |
| J | Sm0030/Kp0056 | Single subtype | 3 c | Chicken meat |
Subtype Sm0030/Kp0056 was identified in three chicken samples (E, I, J), collected at least six weeks apart. A band of approximately 25 kb was intermittently observed in the DNA fragments from subtype Sm0030/Kp0056 isolated from chicken sample J, suggesting it may represent a plasmid [
24] (
Figure 1 and
Figure 2). In designation of a
SmaI banding profile, only bands between 700 and 50 kb were included, therefore the intermittent nature of this band did not affect the interpretation of members of this subtype.
Figure 1.
Pulsed-field gel electrophoresis (PFGE) profiles of Campylobacter jejuni representing all chicken carcass isolates.
Figure 1.
Pulsed-field gel electrophoresis (PFGE) profiles of Campylobacter jejuni representing all chicken carcass isolates.
The literature reports differences in the numbers of subtypes identified in individual samples when tested by various methods. A study using enrichment methods for isolation of campylobacters, identified only one subtype per chicken carcass when up to five isolates per sample were typed [
25]. In comparison, studies using direct plating or a combination of enrichment or direct plating noted higher numbers of subtypes (two-five) in
Campylobacter positive carcasses when, on average, less than seven colonies per carcass were tested [
26,
27,
28].
It is probable, given the selective nature of enrichment broths such as modified Exeter (m-Exeter), that growth characteristics of each environmental
Campylobacter subtype in the broth will vary. The degree of this variation is unknown and consequently, if the initial sample contained equal numbers of two subtypes, it is possible that one may grow faster and be identified by subtyping as being dominant due to its growth characteristics. The same premise will hold for direct plating where injured/stressed cells have a lower probability of recovery in comparison to enrichment [
26]. It is recognized that the problem of selection of dominant strains may be overcome by employing more than one procedure for each sample to enable identification of individual strains that vary under different growth conditions [
5].
None of these factors undermine the use of the enrichment-PCR method as a tool for the long term isolation of campylobacters from a wide range of environments. It does suggest, however, that if studies are time-limited and sample numbers are small then enrichment should be combined with direct plating or enrichment should employ two incubation temperatures, for example, 37 °C and 42 °C [
29]. This may achieve a higher diversity of subtypes from individual samples with less input of labour, as fewer colonies per method may need to be typed. A comparative study of the diversity of subtypes isolated by enrichment
versus direct plating may be worthwhile to establish the benefits of either method and to determine if it is better to use both methods and subtype only one or two colonies isolated from each.
2.2. Comparison of C. jejuni Isolates from Chicken Carcasses and Human Clinical Samples
To increase understanding of the impact of chicken consumption on the number of campylobacteriosis cases, it is important to determine what proportion of
C. jejuni subtypes identified on chicken carcasses are indistinguishable from those in human faecal specimens. Assumptions drawn from case studies and experimental data suggest up to 80% of human cases can be attributed directly to chicken preparation and consumption [
11,
30]. Multiple subtypes of
C. jejuni are present in chickens [
31], and it has been suggested that some of the subtypes causing disease in humans may be minor strains in the chicken microflora and therefore, not identified by conventional bacteriological techniques where only a single colony is typed [
32].
This study addressed whether the numerically minor subtypes identified on chicken carcasses were associated with human cases. Confirmation of this finding would imply that numerically non-dominant subtypes in chicken meat are important in the aetiology of campylobacteriosis, and suggest that isolation techniques need to be directed toward identifying both numerically dominant and non-dominant subtypes in a sample.
Results of the comparison between
C. jejuni isolates from humans and chicken carcasses are presented in
Table 1. Additional information on the identification of the
SmaI/
KpnI profiles in other matrices and other chicken meat samples from previous studies is also shown after comparison with
Campylobacter isolates in the PulseNet Aotearoa NZ database, 6 of the 13
SmaI/
KpnI subtypes identified in this study (
Table 1), had not previously been observed from cases of campylobacteriosis (
n = 1248) in NZ. Furthermore, five of these six subtypes had not been observed from any other matrix besides chicken meat. Three chicken samples (A, F and H) carried single subtypes that were each observed in one clinical case, but not from any other matrix, including chicken samples collected outside of this study. The seven subtypes which were identified in clinical samples were isolated from eight of the ten chicken carcasses analysed in this study. In addition, 2 of these subtypes (dominant and a sole subtype) were identified in 9 and 32 human clinical samples each, suggesting that these subtypes are important in the epidemiology of campylobacteriosis in NZ. In total, 50 clinical isolates in the database were indistinguishable from chicken isolates from this current study; however, none of the human isolates were collected in the same geographical location or timeframe as the chicken isolates.
In the three chickens that carried minor and/or intermediate subtypes, the numerically non-dominant subtypes did not correlate with any isolates from human cases. The fourth chicken (E), as previously mentioned, carried two co-dominant C. jejuni subtypes; and each of these subtypes was identified in three human cases. The subtype (Sm0030/Kp0056) was also identified in two other chicken carcasses (I and J), which were independent samples, having been collected at different times. The results from this study imply that the non-identification of minor/intermediate subtypes using the enrichment-PCR method from chicken carcasses may not be significantly limiting information relevant to tracking the source of an infection to chicken carcasses. Confirmation of this suggestion regarding numerically non-dominant subtypes would require a larger dataset of multiple subtypes collected from a large number of chickens and compared with human isolates from the same geographical area and timeframe.
2.3. Relevance of Clonally Related Campylobacter Subtypes
During this investigation, the question of the relevance of clonally related subtypes occurring in the same sample was identified as warranting further research in regards to outbreak investigations. An additional point, relevant to outbreak studies, is the identification of multiple subtypes (albeit infrequently) in individual human cases of campylobacteriosis [
16,
17].
The benefits of utilizing a double RE digestion were shown with Chickens B and E. For Chicken B, one isolate (labeled minor) had the same
KpnI profile as the other isolates in the same sample, but a different
SmaI pattern (Sm0038) (
Figure 1). In Chicken E, the
SmaI fingerprint for the two subtypes suggested that they were visually similar and could be clonally related (
Figure 1). Further discrimination revealed by the
KpnI digest, however, suggested no clonal relationship.
Chicken C carried potentially clonally related subtypes (
Figure 1). In Chicken C, the three
C. jejuni strains which produced
KpnI profiles could not be digested by
SmaI (subtypes: Non-cutting/Kp0152–Kp0154). The
KpnI profiles of two of these
C. jejuni strains were clonally related by the criteria of Tenover
et al. in that Kp0152 had a band at approximately 270 kb which was absent in Kp0153 [
33]. Profile Kp0154, although looking superficially similar to Kp0152 and Kp0153, had too many different bands from either subtype (five and four band differences, respectively) to be considered clonally related. One of the clonally related subtypes was intermediate in prevalence (12%) suggesting it would have a low probability of being identified. If the numerically dominant subtype was the only subtype isolated from the chicken, and the minor or intermediate chicken subtypes were present in the human case, then the link between the source and the case would have been missed. This highlights the need to examine both indistinguishable and closely related banding patterns when correlating human and chicken isolates. It should be noted, however, that in the current study, none of the potential clonal subtypes from the same carcass were identified in human cases. In addition, the definition of dominant and minor subtypes in a sample may be dependent on the isolation method employed. Future comparisons of multiple subtypes in a sample could, therefore, include quantitative studies such as quantitative PCR to clarify which
C. jejuni subtypes are numerically dominant in a sample [
34].
There were two examples of subtypes identified in human clinical samples that were potentially related to chicken subtypes raising the question whether the human and chicken subtypes would be recognised as potentially clonally related in an outbreak investigation (
Figure 2 and
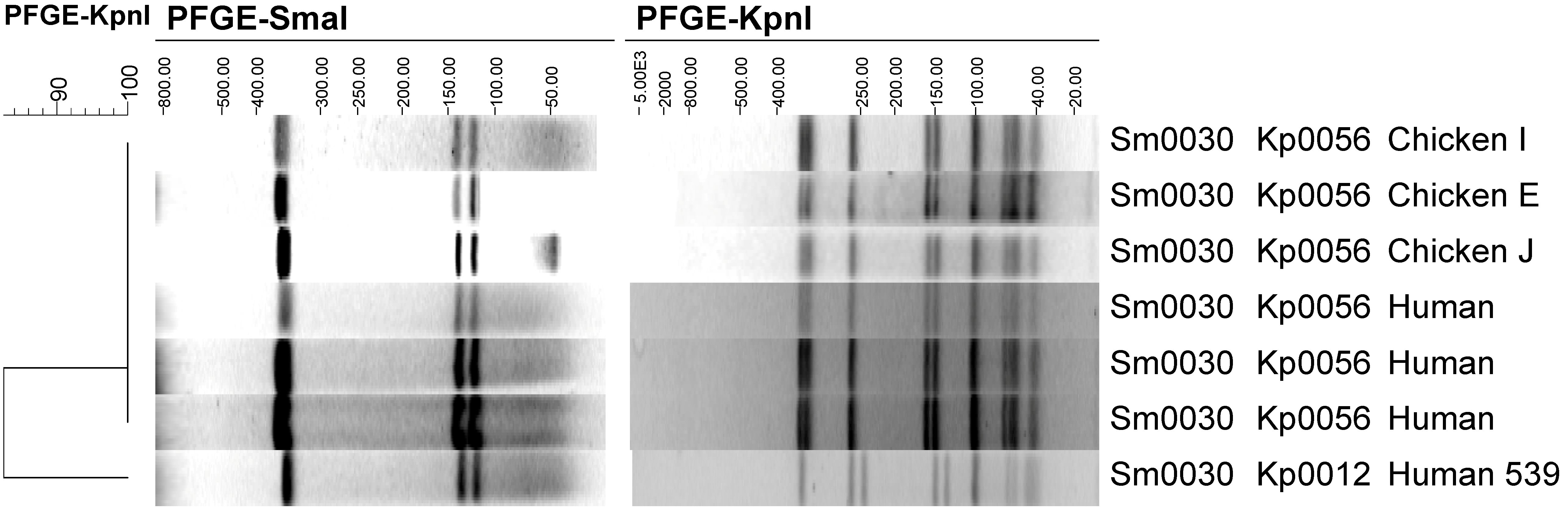
Figure 3). Both of the co-dominant subtypes from Chicken E were clonally related to human subtypes. The
KpnI profile of chicken isolate Sm0030/Kp0056 is presented in
Figure 2 along with the three human faecal isolates that had an indistinguishable profile by
SmaI and
KpnI digestion. In addition, the human faecal isolate 539 (Sm0030/Kp0012) is shown in
Figure 2. Clinical isolate 539 has the same
SmaI profile as the chicken subtype Sm0030/Kp0056 but a different
KpnI profile. This
KpnI profile (Kp0012) is, by the criteria of Tenover
et al. “possibly related” to the chicken isolate, because it has four band differences to Kp0056 [
33]. These band differences are consistent with the occurrence of two genetic events, probable deletions in the bands occurring at approximately 350 and 175 kb in Kp0056 and appearing as bands at approximately 250 and 130 kb (respectively) in Kp0012.
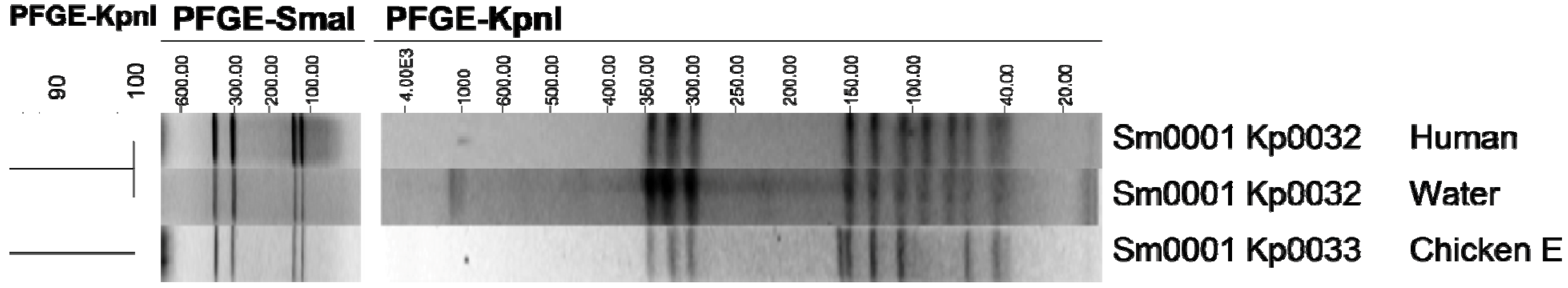
Subtype Sm0001/Kp0033, which was identified in 48% of the isolates from chicken carcass E, was identified in three human cases of campylobacteriosis. In addition, this subtype is potentially clonally related to Sm0001/Kp0032 which was isolated from a human case of campylobacteriosis and from a sample of roof water (
Figure 3). The band at approximately 160 kb in Kp0033 does not appear in Kp0032, but two bands appear at approximately 70 and 90 kb in Kp0032 (
Figure 3). This is suggestive of a single genetic event possibly resulting in a new RE site in the 160 kb fragment (Kp0033) which is cleaved by
KpnI into two fragments of 70 and 90 kb (Kp0032). Further genotypic and phenotypic characterisation of the potentially clonally related subtypes would be required to determine their degree of relatedness. The genotypic plasticity inferred by these isolates and their putative relatedness must be kept in context as Sm0030/Kp0056 was isolated from two other chicken samples in this study that were collected at different times, the closest interval between the three chicken samples being six weeks. Other studies have recognized that some strains of
Campylobacter appear to remain genetically stable under both environmental and laboratory conditions [
35,
36].
Figure 2.
Clonal relationships between Campylobacter jejuni human isolates and subtype Sm0030/Kp0056 from chicken meat.
Figure 2.
Clonal relationships between Campylobacter jejuni human isolates and subtype Sm0030/Kp0056 from chicken meat.
Figure 3.
Clonal relationships between Campylobacter jejuni human and environmental isolates and subtype Sm0001/Kp0032 from chicken meat.
Figure 3.
Clonal relationships between Campylobacter jejuni human and environmental isolates and subtype Sm0001/Kp0032 from chicken meat.
2.4. Chicken as a Vehicle for Transmission of Campylobacter to Humans
Eight of the ten chicken carcasses carried seven pathogenic subtypes, but none of the minor/intermediate subtypes were implicated in campylobacteriosis cases. Identification of non-dominant chicken subtypes such as those identified in this study; therefore, do not support the hypothesis that minor subtypes carried by retail chicken contribute to the transmission of
Campylobacter to humans. The proportion of subtypes from chickens (6 of 13) that were not identified in human cases raises the question as to whether those subtypes are adapted to colonise chickens or the processing environment, as 5 of these were not identified in any matrix other than chicken meat. Further investigation of multiple isolates from a larger sample size of chicken carcasses would be required to confirm this proposal, and could lead to an investigation of virulence factors in pathogenic subtypes. A time-course study of the co-colonisation of chickens with multiple strains of
C. jejuni previously isolated from chickens, reported that two of the seven
C. jejuni strains were dominant in the
Campylobacter flora [
31]. In addition, there was a shift in the dominant strain during the investigation, which the researchers ascribed to influences from the chicken host immune response rather than differences in background flora. Studies of the co-infection dynamics of campylobacters in poultry are important for understanding
Campylobacter epidemiology, in particular, whether pathogenic strains dominate the infection cycle in chickens or can be outcompeted by campylobacters of less significance to the human population.
The lack of correlation between subtypes in chicken carcasses and the 61 human cases from the same region and timeframe was unexpected. A factor that may have affected this result was the rural nature of the region where the study was conducted. The fact that eight of the ten chickens were contaminated with subtypes implicated in human campylobacteriosis, but not from the human cases within the study region, may suggest other routes of transmission of campylobacters in this rural area. We have previously shown that cases mainly occurred in abattoir workers, farmers, their families and farm workers, and from the largest town in the area, which had a population of 14,000 [
4,
9]. Subsequent analysis of human cases and subtypes isolated from the environment revealed the highest similarity to human cases was with subtypes isolated from ruminant animals (sheep, dairy cows and cattle). The greater similarity between human and ruminant isolates may suggest that close contact between farm/abattoir workers and the animals they work with is a more likely route of
Campylobacter infection in comparison with chicken consumption for this rural community. A study in Finland typed human isolates (
n = 176) from two geographically distinct rural and urban areas and their findings also supported a potential distinction between routes of infection in rural and urban environments [
37].
3. Experimental Setup
Fresh chicken carcasses were collected from retail stores in a rural township and transported to the laboratory under refrigeration to be processed by a m-Exeter enrichment-Polymerase Chain Reaction (PCR) method within 24 h [
4,
38]. In brief, whole chicken carcasses were aseptically transferred into a sterile plastic bag with 250 mL of sterile buffered peptone water (pH 7.2, BPW) (1.07228, Merck, Darmstedt, Germany). After massaging the carcass with the BPW for 2 min, 10 mL of the chicken rinse was transferred to a sterile Whirl Pak bag. 90 mL of enrichment broth was added and the sample mixed in a Colworth Stomacher 400 (A.J. Seward, London, UK) for 15 s. The primary enrichment was incubated for 4 h at 37 °C and then incubated for a further 44 h at 42 °C. All incubations were carried out in a 10% CO
2 incubator [
39]. After 48 h, 100 µL of primary enrichment was transferred to 10 mL of m-Exeter broth, and incubated at 42 °C for 24 h under the same conditions. Preparation of
Campylobacter cells for DNA extraction from enrichment broth and analysis by multiplex PCR is described by Wong
et al. [
38]. In brief, 1 mL of enrichment broth was removed, centrifuged (4000
g, 20 min) and the pellet washed three times in 1 mL of sterile phosphate buffered saline (PBS; BR14a Oxoid, Hampshire, UK) by vortexing and centrifugation (4000
g, 10 min). Final re-suspension of washed pellet was in 400 µL of PBS prior to heat lysis preparation for DNA amplification [
38].
3.1. Characterisation of C. jejuni from Chicken Carcasses
Ten chicken carcass enrichments positive for
C. jejuni by PCR were spread onto m-Exeter plates and 25 individual colonies per sample purified by streaking onto Columbia blood agar (CBA, Merck, Darmstedt, Germany) and incubated at 42 °C under 10% CO
2 [
39]. Isolated colonies were stored in cryovials containing glass beads (3 mm, Ajax Finechem, Taren Point, Australia) and 20% glycerol (BDH Laboratory supplies, Poole, England) in Brain Heart Infusion broth (Difco, Detroit, MI, USA) in a −80 °C freezer. The ten chicken carcasses were collected over seven months.
3.2. Colony Identification by PCR
Identification of bacterial colonies by PCR directly from plate cultures, was achieved by removal of a portion of a single isolated colony and resuspension in 27.0 µL of sterile distilled deionized water in a 0.5 mL thin-walled PCR tube. Within 15 min from the time of colony resuspension, the tube was heated for 3 min at 100 °C and then cooled to 4 °C. Prepared premix was added to the PCR tube to obtain a final volume of 50 µL. Details of PCR premix preparation and PCR analysis is outlined in Wong
et al. [
38]. Initially, isolates from the enrichment were confirmed as
C. jejuni by PCR analysis of a single colony per enrichment sample. Subsequently, PCR confirmation was performed on up to five isolates per PFGE subtype identified in each chicken carcass [
26].
3.3. Colony Identification by Biochemical Analysis
Phenotypic characterisations of up to four isolates per PFGE subtype identified in each chicken carcass were subjected to confirmatory identification by the following biochemical tests; Gram stain, hippurate hydrolysis, catalase and oxidase reactions, nitrate reduction, susceptibility to nalixidic acid and resistance to cephalothin [
40,
41]. Lack of growth at 25 °C and under aerobic conditions was also verified for the isolates. Positive controls of
C. jejuni (Enteric Reference Laboratory (ERL) 96 3376) and
C. coli (ERL 97/454) were included for all tests.
3.4. Subtyping of C. jejuni by Pulsed-Field Gel Electrophoresis (PFGE)
Pulsed-field gel electrophoresis was performed using the 24 h PulseNet protocol for the analysis of Gram negative microbes [
42].
Salmonella Branderup (H9812) [
43] was used as the molecular size standard and Megabase agarose (Bio-Rad Laboratories, Hercules, CA, USA) was used to make the plugs. All
Campylobacter isolates were subjected to restriction enzyme digestion using
SmaI enzyme (Roche, Applied Science, Penzberg, Germany). A subset of the 25 isolates from each chicken sample was restricted with a second enzyme
KpnI (New England Biolabs (NEB), Hitchin, UK). Due to resource constraints not all of the 25 isolates from each chicken sample were analysed by
KpnI digestion and electrophoresis. A statistical method termed the Sampling Attribute Plan was employed to calculate the sample size required to determine 95% confidence levels for the results [
44]. If the sample number was 15 from the batch of 25 indistinguishable
SmaI subtypes, and none of the 15 isolates produced a different
KpnI profile then there would be 95% confidence that the batch of 25 will have no more than five isolates (0%–21.8%) with a different
KpnI profile.
Gel electrophoresis was carried out on a CHEF DRIII system (Bio-Rad) with the cooling module set at 14 °C. The image was captured with a Gel Doc 2000 gel documentation system (Bio-Rad), and band patterns analyzed and compared using BioNumerics software (Applied Maths, Kortrijk, Belgium). Only fragments in the range 700 to 50 kb were analyzed for
SmaI, and 700 to 80 kb fragments were analyzed for
KpnI [
10]. Smaller fragments were not consistently resolved. The analytical parameters were the band-based Dice similarity coefficient and the unweighted pairs geometric matched analysis (UPGMA) dendrogram type with a position tolerance setting of 1.5% for optimization and position tolerance of 1.5% band comparison. All test isolates were normalized to the known molecular size bands of the
Salmonella serotype Braenderup H9812 standard subtype [
43]. In addition, all gels were run with a well-characterised subtype of
C. jejuni, CPH011453, which acted as a control for the digestion reaction for both restriction enzymes.
3.5. Correlation of Chicken Carcass Isolates and Subtypes Identified in Human Clinical Samples
The PFGE profiles of isolates identified from chicken carcasses were compared with profiles in the PulseNet Aotearoa NZ Campylobacter database. At the time of comparison, the database contained data for 3301 isolates collected nationwide, including 1248 C. jejuni human clinical isolates, 581 chicken meat, 199 bovine meat, 171 river water, and 221 bovine, 93 ovine, 295 chicken and 66 duck composite/individual faecal sample isolates. The 1248 C. jejuni isolates from human sources included 61 human clinical isolates obtained from the same geographical area and within a one year period of the chicken carcass isolates. Human clinical isolates of C. jejuni in the database which had been analysed for SmaI but not KpnI and were of interest to this study were subjected to PFGE KpnI digestion.


{kind=link}
{kind=link}
{kind=link}