
Enhanced Plant Performance in Cicer arietinum L. Due to the Addition of a Combination of Plant Growth-Promoting Bacteria


1
Department of Physical, Chemical and Natural Systems, University Pablo de Olavide, Seville 41013, Spain
2
Botany and Zoology Department, University of Stellenbosch, Private Bag X1, Matieland 7602, South Africa
*
Author to whom correspondence should be addressed.
Agriculture 2017, 7(5), 40; https://doi.org/10.3390/agriculture7050040
Submission received: 27 February 2017
/
Revised: 24 April 2017
/
Accepted: 27 April 2017
/
Published: 2 May 2017
(This article belongs to the Special Issue Dynamics of Root–Soil–Microbial Interactions)
Abstract
:Current agriculture is based on external fertilizers that jeopardize soil fertility. Alternative fertilization systems might come from the use of soil-borne bacteria with plant growth enhancing ability. Here, six bacterial strains that produce varying concentrations of indole acetic acid (IAA) were tested individually and in consortia for plant growth promotion and fitness-related traits of Cicer arietinum. The nitrogen fixer Mesorhizobium ciceri consistently increased biomass production and N content. In the absence of this strain, IAA Psedomonas putida and Bacillus megaterium hindered plant growth and fitness-related traits. The application of mixes of the three strains always resulted in better plant performance when M. ciceri was present. Mixes that included a P. putida strain that produced low levels of IAA appeared more likely to promote plant growth than mixes that included P. putida strains that produced high levels of IAA or mixes that included B. megaterium. The low levels of IAA produced by the selected strains, compared to the high IAA-producing strains had a significantly greater positive effect on plant biomass accumulation, flower, pod, and seed production, and total plant nitrogen and nitrogen concentrations in seeds.
1. Introduction
Cicer arietinum L., commonly known as chickpeas, is one of the most important legume crops worldwide and represents an important source of protein for humans and fodder for livestock in temperate and semiarid climates [1,2]. It is in these semiarid areas where chickpea’s production is jeopardized due to impoverished soils and hindered geochemical cycles, due to the high temperature and low rainfall. The traditional way of improving soil fertility and the only way to obtain reasonable crops has been the addition of fertilizers. However, chemical fertilizers are costly and have shown negative effects on the soil fertility in long and intermediate terms [3]. Traditional nutrient management includes external fertilizer inputs for the maintenance of high crop productivity [4]; however, in the last several decades, crop yield has not increased proportionally as fertilizer input has increased, leading to low nutrient use efficiency and strong environmental imbalances [4]. Overcoming the lack of fertilization without soil pollution implies exploring alternative routes of nutrient use and input, possible through biological rhizosphere processes.
Soil microbes are paramount for soil health and maintaining nutrient cycling [5]. They convert forms of nitrogen and phosphorus in soil into soluble forms readily assimilated by plants [6]. In addition, rhizobial strains establish a positive symbiosis with plants in the legume family that result in biological nitrogen fixation and enhanced plant growth [7]. Rhizobia and many other microorganisms, concentrated in the rhizosphere of plants, are described as plant growth promoting rizhobacteria (PGPR) [8], which benefit not only the growth of plants [9] but the entire community, including bacteria, plants, and soil fauna [10]; such benefits include an increase in growth and N- and P-uptake by plants, increased photosynthesis, and decreased carbon construction costs, through the inoculation of elite strains in pot experiments [11] and under field conditions [7,12,13]. The mechanisms by which PGPR promote plant growth are still unclear, but among them are (i) the production of phyto-hormones such as indole acetic acid (IAA), gibberellic acid, cytokinins, and ethylene; (ii) asymbiotic N2 fixation; (iii) antagonism against phytopathogenic microorganisms through the production of siderophores or cyanide; and (iv) the solubilization of mineral phosphates [14,15]. Plant growth promotion can be direct, like the symbiotically fixed nitrogen in the nodules of legumes, or can be indirect through the modification of nodulation, competition at the rhizosphere level, or the production of HCN or NH3, which have been reported to be either beneficial or detrimental for plant growth [15]. Therefore, the inoculation of plants by selected microorganisms at different concentrations can have beneficial effects on plant performance. Moreover, as different microbial strains produce different levels of the same phyto-hormone, results might vary from strain to strain and from positive to negative. Mittal [16] reported that effective plant growth promotion may require combinations of bacteria or fungus. Non-indigenous strains would not compete with native microorganisms or could not colonize the rhizosphere properly in new soil environments. Therefore, we suggest that the answer of plants to inoculation should vary according to the strain used and that it is likely that a consortium of PGPR would be preferred for soil inoculations to enhance plant crop.
The present study investigates the effects of six strains of Pseudomonas putida, Bacillus megaterium, and Mesorhizobium ciceri alone and in combination, on plant performance of Cicer arietinum. We hypothesize that chickpea plants inoculated with indole acetic-producing bacteria and Rhizobium are likely to produce more biomass, flowers, and viable seeds than those inoculated with only one of the bacterial strains. We used low and high IAA-producing strains for the three microorganisms. The study was conducted in a greenhouse during the spring season (March–June 2015).
2. Materials and Methods
2.1. Biological Material
The strains of P. putida, B. megaterium, and M. ciceri were from our own collection, obtained from a rhizosphere of diverse plant species in an Iberian bush. The chosen strains were initially selected from several hundred root-colonizing bacteria isolated from soil attached to mat-roots of field-grown plants in the southwest of the Iberian Peninsula. Mat-roots are complex entanglements of roots of annuals growing in the lower layer of bushes; their complexity makes it impossible to ascertain what portion of soil belongs to each root. Potential PGPR were selected for their ability to produce IAA (Table 1), aminocyclopropane-1-carboxilate (ACC deaminase), and siderophores. All strains were identified by sequencing of the 16rRNA section using 1387r and 63f primers for amplification and sequences were identified through NCBI Blast [17]. Identified strains were submitted to the DDJB database for accession numbers (Table 1). IAA production was detected by the Salkowsky reaction [18]; ACC deaminase activity was qualitatively tested in peptone-protease water with hydrolyzed casein, anhydrous glycerol, using aminocyclopropano-1-carboxilic acid as the only source of N [19]; siderophores were assayed on the Chrome azurol S agar medium [20].
The strains were maintained on yeast extract mannitol (YEM) agar and Louria Bertani media [21] at 4 °C. For the inoculation of seedlings, cultures of P. putida and B. megaterium were grown for 36 h in LB broth at 23 °C in an orbital shaker at 100 rpm before dilution to the required concentration of cells. Culture of M. ciceri was grown for six days in the appropriate YMA broth at 27 °C as described before.
Seeds of C. arietinum were surface-sterilized in 70% ethanol for 5 min and 1% sodium hypochlorite for 3 min and then washed six times in sterile distilled water. Once germinated, 15 seedlings were transplanted to the appropriate treatment.
2.2. Experimental Setting
Experiments were conducted in a greenhouse in bleached plastic pots (20 × 18 cm), using sterile loamy soil (pH 6.2, organic matter 1.77%, available-N 12 mg kg−1, available-P 2 mg kg−1, K 112 mg kg−1, Ca 0.417 mEq 100 g−1 soil, Mg 0.084 mEq 100 g−1 soil, 1.5 kg soil per pot). Soil was thoroughly mixed and passed through a 2 mm sieve to remove large particulate matter. Soil was autoclaved at 121 °C, 25 atmosphere for 1 h on three alternate days. Counts of agar plates after autoclaving proved the absence of cultivable microorganisms in the used soil. Surface of the filled pots was covered with sterile polyurethane beds to prevent airborne contamination, and watering was conducted weekly through a capped watering pipe. All pots were randomly arranged in the greenhouse at the University Pablo de Olavide (Seville, Spain) and rotated each week to avoid environmental positional effects within the greenhouse. The pots of each inoculation treatment were maintained on independent benches 2 m apart from each other.
The seeds were inoculated in 15 different sets of pots with 15 replications each. These sets included the following: (1) sterile soil; (2) soil + B. megaterium, low IAA producer (BmL); (3) soil + P. putida Low IAA (PpL); (4) soil + M. ciceri Low IAA (McL); (5) soil + B. megaterium, high IAA producer (BmH); (6) soil + P. putida High IAA (PpH); (7) soil + M. ciceri, high IAA (McH); (8) soil + BmL + PpL; (9) soil + BmL + McL; (10) soil + PpL + McL; (11) soil + BmH + PpH; (12) soil + BmH + McH; (13) soil + PpH + McH; (14) soil + BmL + PpL + McL; (15) soil + BmH + PpH + McH. The control treatment consisted of sterile un-inoculated soil watered with 50% strength of a Hoaglands solution [22]. Inoculation treatments consisted of growth phase broth cultured inoculant at 1 × 106 cells mL−1. The three strains, P. putida (Pp), B. megaterium (Bm), and M. ciceri (Mc) were first individually applied to pots. For each bacteria, we had high and low IAA producers (Table 1).
One germinated seed was transferred to each pot in all treatments. At the emergence of the cotyledons, 100 mL of the appropriate inoculum was added according to treatment and covered with polyurethane beads to prevent contamination from airborne microorganisms. After 4 h, each pot was watered with distilled sterile water. Water was added as necessary to maintain soil moisture through the length of the experiment. All pots were initially supplemented with 50% strength of a Hoaglans solution to avoid plant mortality due to the lack of nutrients. The average number of flowers and pods were calculated in each treatment. Plants were harvested after maturity, 17 weeks after planting. Their shoot and root dry weights, seed number, and weight was measured and recorded. The nitrogen accumulated in shoots was calculated by multiplying the weight of dry shoots by the nitrogen content as measured by the semi micro-Kjedahl method [23].
2.3. Statistical Analysis
Pot experiments were arranged in a completely randomized block design. Statistical analysis was conducted using a one- or two-way ANOVA with the Statistical Package for Social Sciences (SPSS) software, version 11.5 (IBM Corp., Armonk, NY, USA). Comparisons of means were performed by the LSD test at p = 0.05.
3. Results
Three bacterial strains, B. megaterium, P. putida, and M. ciceri, each with differential ability to produce IAA, chosen for their capacity to induce growth promotion in plants, were tested for their ability to promote the growth of C. arietinum in sterile soils. Simultaneously, we tested the ability of the same strains to exert a positive effect in other plant traits, such as flower, pod, and seed production and the amount of nitrogen accumulated by both shoots and seeds. The individual and combined positive effect of the strains on the growth and seed production of chickpea plants was observed by comparing the plant performance in pots containing only sterile soil with that containing low-nutrient soil supplemented with 50% strength of a Hoaglans solution and that containing the same solution plus the inoculants.
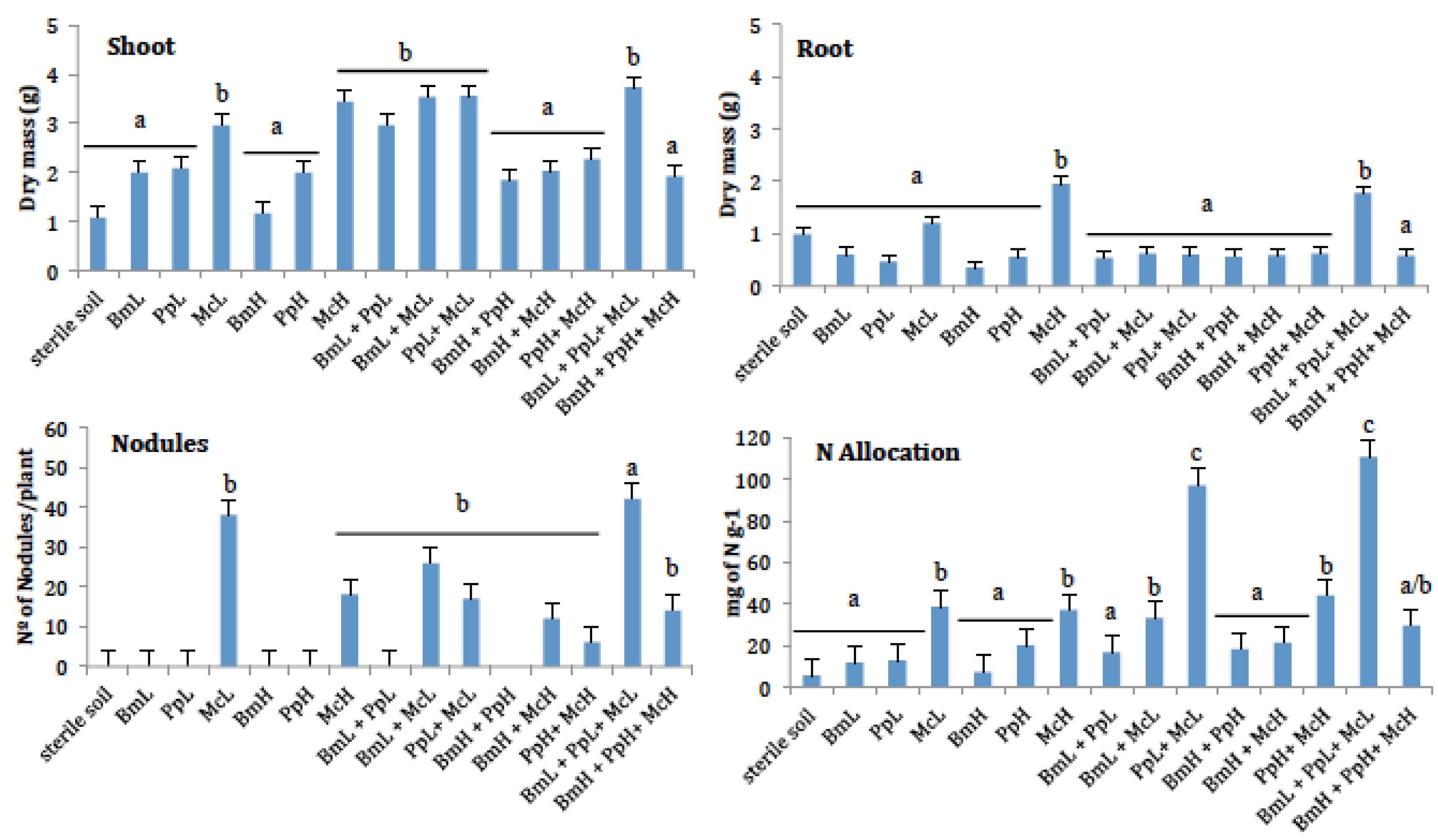
The inoculation of McL and McH IAA-producing strains had the most significant (p = 0.0021) stimulatory effect in terms of shoot production (Figure 1). Shoot biomass production ranked between 1.085 mg in the sterile soil and 3.562 mg in the PpL + McL treatment. Nodules were only produced in plants inoculated with M. ciceri (McL and McH). The higher number of nodules was consistently and significantly (p > 0.0001) obtained in the presence of the low IAA-producing strain (Figure 1). Similarly, plants in all treatments with McL alone or in combination with PpL attained the most significant (p = 0.0403) total N content per gram of dry biomass (Figure 1). Nitrogen concentrations were in the range of 2.7445 mg of N/g of biomass in plants grown in sterile soil, and 20.096 mg of N/g of biomass in plants under the BmL + PpL + McL treatment.
In pots containing only soil, the number of seeds, pods, and seeds per plant attained values of 36.0, 24.6, and 26.03, respectively (Table 2). These values significantly (p = 0.0334) differed from those obtained in plants under the PpL + McL treatment, whose values for the same variables were 252.0, 213.2, and 266.79. High values of flower, pod, and seed production were also attained in plants treated only with PpL, followed by those treated with PpH + McH and BmL + PpL + McL. Moreover, significantly high values were achieved by treatments BmH + PpH, MmL, and McL (Table 2).
Significant differences were observed in seed nitrogen accumulation with the greatest value of 12.67 mg of N per g of dry biomass in plants treated with PpL + McL, followed by the combination of BmL + PpL + McL, PpH + McH, BmH + PpH + McH, and McL (Table 2). The lowest value of 1.49 mg of N per g of dry biomass was reached again in plants grown in sterile soil. Significantly low values (p > 0.0001) were observed in plants grown inoculated with BmL (2.4621 mg N. g biomass), BmH (2.56 mg N. g biomass), BmL + PpL (2.62 mh N. g biomass), and PpL (2.72 mh N. g biomass) (Table 2).
4. Discussion
Agriculture could take advantage of symbiotic relationships between plants and microbes to enhance plant productivity and crops by manipulating the composition of soil microbial communities [24,25]. Different bacterial strains have the ability to induce growth promotion on diverse crop production [25,26,27,28] through either the induction of nitrogen fixation or the production of varying plant hormones. Results from the present study reinforce this conceit and emphasize the fact that the individual effect of a single strain is modified by the presence of one or more other strains, both in the same and in different species.
We report that chickpea plants inoculated with indole acetic-producing bacteria and Rhizobium rendered more plant biomass, flowers, and viable seed than those inoculated with only one of the bacterial strains and that the level of IAA produced by the selected strains might have an effect on the total plant performance. The seedlings treated with M. ciceri attained high values of shoot biomass. Any time this strain was added both biomass and total nitrogen concentration in plants was enhanced. Simultaneously, plant growth-promoting effect was observed in plants inoculated with the low IAA-producing B. megaterium and P. putida bacteria. Interestingly, any of the consortia that included McL, BmL, and PpL resulted in a great increase in all studied plant parameters, while the use of any of the H strains resulted in the reverse. Nevertheless, when the strain tested was the one producing high levels of IAA, the enhancing effect was hindered. This is not the first time we describe the need for relatively low levels of IAA to enhance plant performance. In a previous experiment [29], we described how an intermediate production of IAA resulted in both enhanced root cluster production and plant biomass crop in Leucadendron salicifolium, Viminaria juncea, and Lupinus albus. This experiment supports the previous ones in terms of the low IAA needed for the increase in plant performance in C. arietinum. The ability of microorganisms to solubilize phosphate has rendered contradictory results [19]. Here, there were only two strains with P-solubilization ability, BmH and PpH. As these two strains were not among those inducing the greatest biomass production, neither helped in plant reproduction, and we cannot relate them to the PGPR properties. Bacillus amyloliquefaciens has been reported to produce IAA and to have ACC deaminase activity [19,30]. The production of the two compounds induces the production of ethylene, which is a root elongation inhibitor and consequently might reduce the total plant biomass production [19,30]. We described three strains with ACC deaminase activity in our study, BmH, PpH, and McL. This fact explains the reduced growth induced by this bacteria; the high IAA in combination with AAC deaminase activity could be responsible for the ethylene production that inhibits plant growth.
It was also interesting to observe that flower, pod, and seed production were not enhanced at their maximum by McL or McH, but by PpL alone or in combination with Mc. Ulzen [31] reported no increase in shoot biomass after inoculation with rhizobia on soybean and cowpea, and similar results were reported by Albareda [32]. Simultaneously, there was a consistent negative correlation among the four reproduction values in all but one (BmH + PpH + McH) treatment. Our work reports reduced the production of reproduction-related traits. These traits are highly N demanding, and we propose that the amount of nitrogen fixed by M. ciceri was inadequate due to the competition of this strain with the other ones and that P. putida can fix nitrogen asymbiotically in a greater proportion than that of M. ciceri. Although we did not measure biological nitrogen fixation by any of the six strains, this ability has been reported in P. agglomerans [33] and in Pseudomonas sp. [34]. The very high number of seeds in treatment BmH + PpH + McH demanded high levels of N that were initially met, but that was not enough to translate in high levels of it in the seeds, as N only accumulates when it is in excess [29,31]. In fact, it was the presence of PpL in all Mc treatments (H and L) that had the greatest effect on plant reproduction traits. We think this effect is due to the IAA added to the plant, which has a synergistic effect in combination with M. ciceri. B. megaterium also produces IAA and its presence at both H or L values did not give the same results as those observed in the presence of P. putida. This fact led us to think that (i) not only IAA but other compounds produced by the bacteria enter into the plant growth-promoting equation [3,6]; (ii) there is competition among the strains that results in improved expression of one or another depending on the superior competitor, a fact that has been described in several soil types [35], under differing levels of nutrients [36], or under changing plant community composition [37]; (iii) there is a plant sanction against particular strains, as has been observed in the case of legumes inoculated with particular rhizobial strains that result in the infection of only one strain [7,38,39].
Here, we described the PGPR effect of low IAA-producing bacterial strains on C. arietinum. These strains not only enhanced plant biomass production but also had a beneficial effect on reproduction-related traits. This study goes one step beyond the in vitro tests and shows that the selected isolates benefit chickpea plants growing in soil. This reveals the potential of these strains for biofertilizer applications and commercial use as biocontrol agents in the field. However, from the estimation of a PGPR-potential to a biofertilizer application, further greenhouse experiments in more complex systems than the gnotobiotic one used here are required. Pots need to be filled with different types of soils that harbor their native microbiota, and how plants and the inoculated strains behave needs to be observed. Field experiments will then need to be conducted to find out the right inoculums that work under real crop conditions.
5. Conclusions
Our study has proven that low IAA production enhanced C. arietinun growth by both individually applied plant growth-promoting bacteria and by elite consortiums of such bacteria. The plant hormone IAA is crucial in determining both plant biomass production and N accumulation in shoots and seeds. Low amounts of IAA, compared to high concentrations, induce greater biomass and N accumulation. IAA has a more pronounced effect on the performance of chickpea plants than P-solubilizing agents of ACC deaminase. The right amount of IAA needs to be balanced with other bacterial derived compounds, such as P-solubilizing agents of ACC deaminase activity.
Acknowledgments
The present study has been self-funded. E. Velazquez from the University of Salamanca provided us with strains of M. ciceri.
Author Contributions
Both authors have equally contributed to the experimental design, the greenhouse experiment performance, and the writing and revising of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Siddiqui, A.Z.; Mahmood, I. Effects of rhizobacteria and root symbionts on the reproduction of Meloidogyne javanica and growth of chickpea. Bioresour. Technol. 2001, 79, 41–45. [Google Scholar] [CrossRef]
- Jukanti, A.K.; Gaur, P.M.; Gowda, C.L.; Chibbar, R.N. Nutritional quality and health benefits of chickpea (Cicer arietinum L.): A review. Br. J. Nutr. 2012, 108, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, N.; Vassilev, M. Biotechnological solubilization of rock phosphate on media containing agro-industrial wastes. Appl. Microbiol. Biotechnol. 2003, 61, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, Y.; Xie, X.; Kim, M.S.; Dowd, S.E.; Paré, P.W. A soil bacterium regulates plant acquisition of iron via deficiency inducible mechanisms. Plant J. 2009, 58, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Ingham, R.E.; Trofymow, J.A.; Ingham, E.R.; Coleman, D.V. Interactions of bacteria, fungi, and their nematode grazers: Effects on nutrient cycling and plant growth. Ecol. Monogr. 1985, 55, 119–140. [Google Scholar] [CrossRef]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Pérez-Fernández, M.A.; Hill, Y.J.; Calvo-Magro, E.; Valentine, A. Competing Bradyrhizobia strains determine niche occupancy by two native legumes in the Iberian Peninsula. Plant Ecol. 2015, 216, 1537–1549. [Google Scholar] [CrossRef]
- Vesquez, P.; Holguin, G.; Puente, M.E.; López-Cortes, A.; Bashan, Y. Phosphate-solubilizing microorganisms associated with the rhizosphere of mangroves in a semiarid coastal lagoon. Biol. Fertil. Soils 2000, 30, 460–468. [Google Scholar] [CrossRef]
- Pii, Y.; Mimmo, T.; Tomasi, N.; Terzano, R. Cesco and Crecchio C Microbial interactions in the rhizosphere: Beneficial influences of plant growth-promoting rhizobacteria on nutrient acquisition process. A Rev. Biol. Fertil. Soils 2015, 51, 403–415. [Google Scholar] [CrossRef]
- Pérez-Fernánde, M.A.; Calvo-Magro, E.; Valentine, A. Benefits of the symbiotic association of shrubby legumes to re-vegetate heavily damaged soils. Land Degrad. Dev. 2016, 27, 395–405. [Google Scholar]
- Omar, S.A. The role of rock phosphate solubilizing fungi and vesicular arbuscular mycorrhiza (VAM) in growth of wheat plants fertilized with rock phosphate. World J. Microbiol. Biotechnol. 1998, 14, 211–219. [Google Scholar] [CrossRef]
- Santi, C.; Bogusz, D.; Franche, C. Nitrogen fixation in non-legumes. Ann. Bot. 2013, 111, 743–767. [Google Scholar] [CrossRef] [PubMed]
- Palacios, O.; Bashan, Y.; de-Bashan, L. Proven and potential involvement of vitamins in interactions of plants with plant growth-promoting bacteria—An overview. Biol. Fertil. Soils 2014, 50, 415–432. [Google Scholar] [CrossRef]
- Mehnaz, S.; Lazarovits, G. Inoculation effects of Pseudomonas putida, Gluconacetobacter azotocaptans, and Azospirillum lipoferum on corn plant growth under greenhouse conditions. Microb. Ecol. 2006, 51, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Ahmad, I.; Kjan, M.S. Screening of free-living rhizospheric bacteria for their multiple plant growth promotion activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Mittal, V.; Singh, O.; Nayyar, H.; Kaur, J.; Tewari, R. Simulatory effect of phosphate-solubilizing fungal strains (Aspergillus awamori and Penicillium citrinum) on the yield of chickpea (Cicer arietinun L. cv GPF2). Soil Biol. Bichem. 2008, 40, 718–727. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI). Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information 1988. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 6 April 2017).
- Cappuccino, J.C.; Sherman, N. Microbiology: A Laboratory Manual, 3rd ed; Benjamin Cummings Pub Co.: New York, NY, USA, 1992; pp. 125–179. [Google Scholar]
- Angulo, V.C.; Sanfuentes, E.A.; Rodríguez, F.; Sossa, K.E. Caracterización de rhizobacterias promotoras de crecimiento en plántulas de Eucalyptus nitens. Revista Argentina de Microbiologia 2014, 46, 338–347. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Vincent, J.M. A Manual for the Practical Study of Root-Nodule Bacteria; Blackwell Scientific Publications Ltd.: Oxford, UK, 1970. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method of Growing Plants without Soil; California Agricultural Experimental Station: Berkeley, CA, USA, 1950; p. 347. [Google Scholar]
- Sarruge, J.R.; Haag, H.P. Análises Químicas em Plantas; ESALQ/USP: Piracicaba, Brazil, 1979; p. 27. [Google Scholar]
- Woomer, P.L.; Huising, J.; Giller, K.E. N2 Africa Final Report of the First Phase 2009–2013. Available online: http://www. 2014, N2Africa.org 138 (accessed on 6 April 2017).
- Asei, R.; Ewusi-Mensah, N.; Abaidoo, R.C. Response of Soybean (Glycine max L.) to rhizobia inoculation and molybdenum application in the Northern savannah zones of Ghana. J. Plant Sci. 2015, 3, 64–70. [Google Scholar]
- Mayak, S.; Tirosh, T.; Glick, B. Plant growth-promoting bacteria that confer resistance to water stress in tomatoes and peppers. Plant Sci. 2004, 166, 525–530. [Google Scholar] [CrossRef]
- Mathu, S.; Herrmann, L.; Pypers, P.; Matiru, V.; Mwirichia, R.; Lesueur, D. Potential of indigenous bradyrhizobia versus commercial inoculants to improve cowpea (Vigna unguiculata L. walp) and green gram (Vigna radiate L. wilczek.) yields in Kenya. Soil Sci. Plant Nutr. 2012, 58, 750–763. [Google Scholar] [CrossRef]
- Nieto-Jacobo, M.F.; Steyaert, J.M.; Salazar-Badillo, F.B.; Nguyen, D.V.; Rostás, M.; Braithwaite, M.; De Souza, J.T.; Jimenez-Bremont, J.F.; Ohkura, M.; Stweart, A.; et al. Environmental growth conditions of Trichoderma spp affects indole acetic adic derivates, volatile organic compounds, and plant growth promotion. Front. Plant Sci. 2017, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Lamont, B.; Pérez-Fernández, M.A. Soil bacteria control plant growth and root-cluster production by species from three continents. Ann. Bot. 2016, 118, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Read, T.D.; Salzberg, S.L.; Pop, M.; Shumway, M.; Umayam, L.; Jiang, L.; Holtzapple, E.; Busch, J.D.; Smith, K.L.; Schupp, J.M.; et al. Comparative genome sequencing for discovery of novel polymorphisms in Bacillus anthracis. Science 2002, 296, 2028–2033. [Google Scholar] [CrossRef] [PubMed]
- Ulzen, J.; Abaidoo, R.C.; Mensah, N.A.; Masso, C.; AbdelGadir, A.H. Bradyrhizobium inoculants enhance grain yields of soybean and cowpea in Northern Ghana. Front. Plant Sci. 2016, 7, 1770. [Google Scholar] [CrossRef] [PubMed]
- Albareda, M.; Rodríguez, D.N.; Temprano, F.J. Soybean incoulation: Dose, N fertilizer supplementation and rhizobia persistence in soils. Field Crops Res. 2009, 113, 352–356. [Google Scholar] [CrossRef]
- Wright, S.; Zumoff, C.; Scheider, L.; Beer, S. Pantoea agglomerans EH318 producer two antibiotics that inhibit Erwinia amylovora in vitro. Appl. Environ. Microbiol. 2001, 67, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.; Valencia, S.; Bernal, J.; Martínez, P. Isolation of Enterobacteria, Azotobacter sp. and Pseudomonas spp. producers of 71 Indole-3-Acetic Acid and Siderophores, from Colombian Rice Rhizosphere. Revista Latinoamericana de Microbiología 2000, 42, 171–176. [Google Scholar]
- Bever, J.D. Soil commynity feedback and the coexistence of competitors: Conceptual framework and empirical tests. New Phytol. 2003, 157, 465–473. [Google Scholar] [CrossRef]
- He, W.M.; Zhang, H.; Dong, M. Plasticity in fitness and fitness-related traits at ramet and genet levels in a tillering grass Panicum miiaceum under patchy soil nutrients. Plant Ecol. 2004, 172, 1–10. [Google Scholar] [CrossRef]
- Hawkes, C.V.; Belnap, J.; D’Antonio, C.; Firestone, M.K. Arbuscular mycorrhizal assemblages in native plant roots change in the presence of exotic grasses. Plant Soil 2006, 281, 369–380. [Google Scholar] [CrossRef]
- Hodge, A.; Stewart, J.; Robinson, D.; Griffiths, B.S.; Fitter, A.H. Competition between roots and soil-microorganisms for nutrients from nitrogen-rich patches of varying complexity. J. Ecol. 2000, 88, 150–164. [Google Scholar] [CrossRef]
- Gubry-Rangin, C.; Garcia, M.; Béna, G. Partner choice in Medicago truncatula—Sinorhizobium symbiosis. Proc. R. Soc. 2010, 277, 1947–1951. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Plant biomass (shoots and roots), the number of nodules, and the nitrogen per gram of plant biomass produced by plants of Cicer arietinum inoculated with 15 combinations of low and high IAA-producing bacteria. Different letters on top of the columns indicate significant differences among treatments after significant ANOVA.
Figure 1.
Plant biomass (shoots and roots), the number of nodules, and the nitrogen per gram of plant biomass produced by plants of Cicer arietinum inoculated with 15 combinations of low and high IAA-producing bacteria. Different letters on top of the columns indicate significant differences among treatments after significant ANOVA.

{kind=link}
Table 1.
Amounts of indole acetic acid (IAA) (µg m−1) produced by six bacterial strains and their aminocyclopropane-1-carboxilate (deaminase ACC) activity and siderophores production ability qualitatively determined. Different letters close to IAA numbers indicate significant differences in the production of this hormone, after a one-way ANOVA. (DDBJ are registration numbers and those strains are awaiting for accession number).
Table 1.
Amounts of indole acetic acid (IAA) (µg m−1) produced by six bacterial strains and their aminocyclopropane-1-carboxilate (deaminase ACC) activity and siderophores production ability qualitatively determined. Different letters close to IAA numbers indicate significant differences in the production of this hormone, after a one-way ANOVA. (DDBJ are registration numbers and those strains are awaiting for accession number).
| Strain | Accession Numbers | IAA (µg m−1) | Deaminase ACC Activity | Siderophores |
|---|---|---|---|---|
| BmL | DDBJ_58fe2926b6e6bc04ce0040f4 | 132 b | − | − |
| PpL | DDBJ_58fe25e1b6e6bcfeb1005c86 | 176 a | − | − |
| McL | USDA 3383T | 121 b | + | − |
| BmH | JSB 31 | 62 b | + | + |
| PpH | DDBJ_58fe02d7b6e6bcfeb1005be4 | 99 b | + | + |
| McH | WSM1271 | 86 b | − | − |
Table 2.
Effect of plant growth promoting bacteria, B. megaterium, P. putida, and M. ciceri (L = low IAA producers and H = high IAA producers) on the plant performance of chickpea plants grown in sterile soil. Different letters next to the numbers indicate significant differences among treatments after significant ANOVA.
Table 2.
Effect of plant growth promoting bacteria, B. megaterium, P. putida, and M. ciceri (L = low IAA producers and H = high IAA producers) on the plant performance of chickpea plants grown in sterile soil. Different letters next to the numbers indicate significant differences among treatments after significant ANOVA.
| Treatment | N° Flowers | N° Pods | N° Seed | N in Seed (mg N. g plant−1) |
|---|---|---|---|---|
| sterile soil | 36 a | 25 a | 2 a | 1.49 a |
| BmL | 132 b | 90 c | 90 b | 2.46 a |
| PpL | 176 b | 148 d | 162 c | 2.72 a |
| McL | 121 b | 86 c | 100 b | 5.06 a,b |
| BmH | 62 b | 49 b | 46 b | 2.57 a |
| PpH | 99 b | 78 c | 82 b | 4.28 a,b |
| McH | 86 b | 66 b | 62 b | 3.72 a |
| BmL + PpL | 49 a | 37 a | 39 b | 2.62 a |
| BmL + McL | 98 b | 74 c | 78 b | 4.33 a,b |
| PpL + McL | 252 c | 213 d | 267 c | 12.67 c |
| BmH + PpH | 139 b | 109 d | 104 c | 4.14 a,b |
| BmH + McH | 77 b | 70 c | 77 b | 4.43 a,b |
| PpH + McH | 163 b | 135 d | 143 c | 8.32c |
| BmL + PpL+ McL | 149 c | 119 d | 115 | 10.93 c |
| BmH + PpH+ McH | 73 b | 53 c | 56 | 6.40 a,b |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pérez-Fernández, M.; Alexander, V. Enhanced Plant Performance in Cicer arietinum L. Due to the Addition of a Combination of Plant Growth-Promoting Bacteria. Agriculture 2017, 7, 40. https://doi.org/10.3390/agriculture7050040
AMA Style
Pérez-Fernández M, Alexander V. Enhanced Plant Performance in Cicer arietinum L. Due to the Addition of a Combination of Plant Growth-Promoting Bacteria. Agriculture. 2017; 7(5):40. https://doi.org/10.3390/agriculture7050040
Chicago/Turabian StylePérez-Fernández, María, and Valentine Alexander. 2017. "Enhanced Plant Performance in Cicer arietinum L. Due to the Addition of a Combination of Plant Growth-Promoting Bacteria" Agriculture 7, no. 5: 40. https://doi.org/10.3390/agriculture7050040
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.