Isolation of Mercury-Resistant Fungi from Mercury-Contaminated Agricultural Soil
by

Reginawanti Hindersah
1,2,*, 
Khainur Rasyid Asda
1,
Diyan Herdiyantoro
1 and
Nadia Nuraniya Kamaluddin
1 1
Department of Soil Science, Faculty of Agriculture, Universitas Padjadjaran Jatinangor Campus, Jalan Raya Bandung-Sumedang Km. 21, Sumedang, West Java 45363, Indonesia
2
Maluku Corner Center of Excellence Universitas Padjadjaran Bandung Campus, Graha Soeria Atmaja Lantai 2, Jalan Dipati Ukur No. 46 Bandung, West Java 40132, Indonesia
*
Author to whom correspondence should be addressed.
Agriculture 2018, 8(3), 33; https://doi.org/10.3390/agriculture8030033
Submission received: 15 December 2017
/
Revised: 16 February 2018
/
Accepted: 17 February 2018
/
Published: 27 February 2018
Abstract
:Illegal gold mining and the resulting gold mine tailing ponds on Buru Island in Maluku, Indonesia have increased Mercury (Hg) levels in agricultural soil and caused massive environmental damage. High levels of Hg in soil lowers plant productivity and threatens the equilibrium of the food web. One possible method of handling Hg-contaminated soils is through bioremediation, which could eliminate Hg from the rhizosphere (root zone). In this study, indigenous fungi isolated from Hg-contaminated soil exhibited Hg-resistance in vitro. Soil samples were collected from the rhizosphere of pioneer plants which grew naturally in areas contaminated with gold mine tailing. The fungi’s capacity for Hg-resistance was confirmed by their better growth in chloramphenicol-boosted potato dextrose agar media which contained various HgCl2 concentrations. Four isolates exhibited resistance of up to 25 mg kg−1 of Hg, and in an experiment with young Chinese cabbage (Brassica rapa L.) test plants, two fungi species (including Aspergillus) were demonstrated to increase the soil’s availability of Hg. The results suggest that Hg-resistant indigenous fungi can mobilize mercury in the soil and serve as potential bioremediation agents for contaminated agricultural land.
1. Introduction
Mercury (Hg) is an environmental pollutant that naturally presents in soil. Soil Hg concentration mainly depends on soil genesis and parent material. Hg enters the soil as a result of anthropogenic activities and through gold extraction [1]. On Buru Island in Maluku, Indonesia, illegal mining makes use of Hg for gold extraction, and has caused increased levels of Hg content in tailing disposal area. Tailing deposition has threatened nearby agricultural land and, subsequently, food production. Mercury can be absorbed by vegetation and may enter the human diet, posing a major threat to public health [2]. The increased possibility of Hg accumulation through plant uptake (along with other heavy metals such as cadmium and lead) is dangerous due to mercury’s status as a known health hazard for humans and other living organisms [3].
The soil Hg level on Maluku varies from 0.1 to 5.0 mg kg−1 [4]. In other locations around the world, the levels of Hg present in the soil are much lower—just 0.003–4.6 mg kg−1 [5]. The concentration of Hg in soil underneath tailing deposits reached up to 100 mg kg−1. Usually, Hg bioavailability is low due to its high affinity towards organic matter and absorption by clay minerals and soil oxide [4,6], as heavy metals can bind to soil particles or penetrate into soil profile through precipitation. This metal fraction is insoluble and largely unavailable for plant uptake [7]. However, increased Hg content in soil can enhance free Hg ions and soluble Hg complexes that are easily absorbed by plant roots [2].
Phytoremediation is a method of pollution extraction that utilizes plants to remove contaminants from the environment. It is considered as one of the most effective practices to address soil contaminated by heavy metals. The potential for accumulation via vegetation is limited by the low availability of heavy metals [7], especially Hg. Soil microorganisms—especially those that colonize the rhizosphere—possess several mechanisms that help change metal bioavailability for root uptake. In addition, the relationship between heavy metal availability and heavy metal toxicity towards soil microorganisms has not yet been proven [8]. Research [8] reported that both short- and long-term exposure of heavy metals results in microbial diversity and activity reduction in soil.
Fungi have been widely used to remediate soil contaminated by heavy metal through several mobilization mechanisms. The soil fungi Aspergillus flavus [9], Aspergillus niger, Trichoderma viride, Humanicola insolens [10], Fusarium oxysporum [11], and Rhizopus arrhizus [12] have been shown to be capable of carrying out heavy metal biotransformation.
Food production in former gold mining areas can be carried out if the Hg level in soil is below the maximum limit. The introduction of beneficial fungi capable of mobilizing heavy metals for plant uptake is a potential solution that could lower Hg levels in productive soil. This study was conducted in order to understand the role of indigenous fungi towards the promotion of Hg uptake. Indigenous fungi were isolated from the rhizosphere of plants grown in mercury-contaminated soil and their fungal resistance in various mercury levels was tested in vitro.
2. Materials and Methods
The study was carried out from February to June 2015 at Universitas Padjadjaran Bandung. Laboratory research was conducted in the Soil Biology Laboratory, while bioassays were performed at a greenhouse belonging to the Faculty of Agriculture in an experimental station located 752 m above sea level. The tailing for the bioassays was taken from Wamsait Village, Waeapo Sub district, Buru District, Maluku Province of Indonesia.
2.1. Isolation, Selection, and Identification of Hg-Resistant Indigenous Fungus
Soil fungus was isolated using the pour method [13] on potato dextrose agar (PDA) in plate (200 mL natural potato extract, 10 g dextrose, 15 g agar, 1 L distilled water). The fungal isolate was sourced from the rooting zone (rhizosphere) of several pioneer plants grown on Hg-contaminated tailings deposited in Wamsait village, Buru District (3°24′ S 126°40′ E). The soil sample was diluted to 10−3 via serial dilution. One milliliter of soil suspension was poured into the agar plate and spread manually prior to incubation at 30 °C for five to seven days. The purity of each fungal isolate grown on agar plate was determined by their morphological uniformity under a light microscope. Cultures were transferred into potato dextrose agar (PDA) slants and maintained at 30 °C for three days before being used for fungal selection.
Four fungal pure cultures were isolated and selected based on their growth characteristics on solid cultures both with and without HgCl2. Mercury chloride was added into 150 mL of PDA before sterilization to final concentrations of 5, 15, and 25 mg L−1 Hg. Sterilization was carried out in an autoclave at 121 °C for 20 min prior to the addition of chloramphenicol. Agar plates were made in 10-cm Petri dishes by pouring 20 mL of liquid PDA at 45 °C. The plates were kept at room temperature at least overnight before usage. Whatman paper disks was sterilized in an oven at 70 °C for 3 h, then dipped into the fungal suspension using sterilized curved forceps; this mixture contained approximately 107 propagul.mL−1. A single paper disk was placed in the middle of the plate agar and incubated for eight days at 30 °C; for each level of Hg, two agar plates were used. Fungal growth was determined by measuring the colonies’ diameters every 24 h for eight consecutive days.
Fungal identification was performed for two fungal isolates that showed the best growth in Hg-contaminated PDA. Fungal species determination was based on either macroscopic or microscopic morphology characteristics, as described by Samson et al. [14]. Macroscopic identification was based on colony color and morphology; microscopic identification was based on hyphae, generative mycelium, and spore type.
2.2. Bioassay on Chinese Cabbage (Brassica rapa L.)
The test plant of the bioassay was Chinese cabbage (Brassica rapa L.) of Brassicacea, a heavy metal accumulator [14]. The experiment was set up with three replications. Experimental factors were the combination of fungal inoculation (isolate A, isolate B, and isolate A + B), organic matter content (1.5%, 3%, and 4.5%), and non-inoculation (serves as a control).
The growth media was prepared in 15-cm diameter black polyethylene bags (polybags). Each individual polybag contained a 1.5-kg mixture of tailing and organic matter. The concentrations of organic matters were 1.5%, 3%, and 4.5% (w/w) of tailing weight. Fungi were inoculated by adding 10 mL liquid inoculant into potato dextrose broth for isolate A and B. The dose of mixed inoculation treatment was 5 mL for each isolate. After inoculation, growth media were incubated for three days in the greenhouse. Individual Chinese cabbage transplants were grown in individual polybags, inside soil cavities with depths of 2 cm. All transplants were maintained for 15 days prior to measuring plant height and dry weight, soil acidity, and available Hg, as well as total Hg content in the plant’s entire biomass.
At 15 days after planting, plants were taken from growth media and dried in a 70 °C oven for two days until they reached a constant weight. The growth media (soil) of the individual polybags was mixed thoroughly and 15 g composite samples collected from three different polybags with the same treatment. The available Hg was measured using Atomic Adsorption Spectrometry (AAS) after extraction via Morgan-Wolf solution (100 g Na-acetic acid (NaC2H3O2·3H2O), 30 mL glacial acetic acid, and 0.05 g diethylenetriaminepentaacetic acid (DTPA). Meanwhile, the total Hg content in plant biomass was measured by AAS after dry biomass destruction with mixed perchloric and nitric acid.
3. Results
3.1. Tailing Characterization
Tailing texture was sandy loam with 10.77 mg of total Hg. Chemical characteristics of tailing showed that the pH was 9.17 and that it was very low in C-organic, total N, total P, and total K; it was low in cation exchange capacity (CEC) while very high in base saturation. To increase organic matter content in the tailing, cow manure compost (C-organic 33.5%; C:N ratio 18:1; water content 22.34%, pH 6.21; P2O5 1.2%; K2O 3.71%) from the Faculty of Animal Husbandry, Universitas Padjadjaran, West Java, was mixed with the tailing.
3.2. Indigenous Fungal Isolates
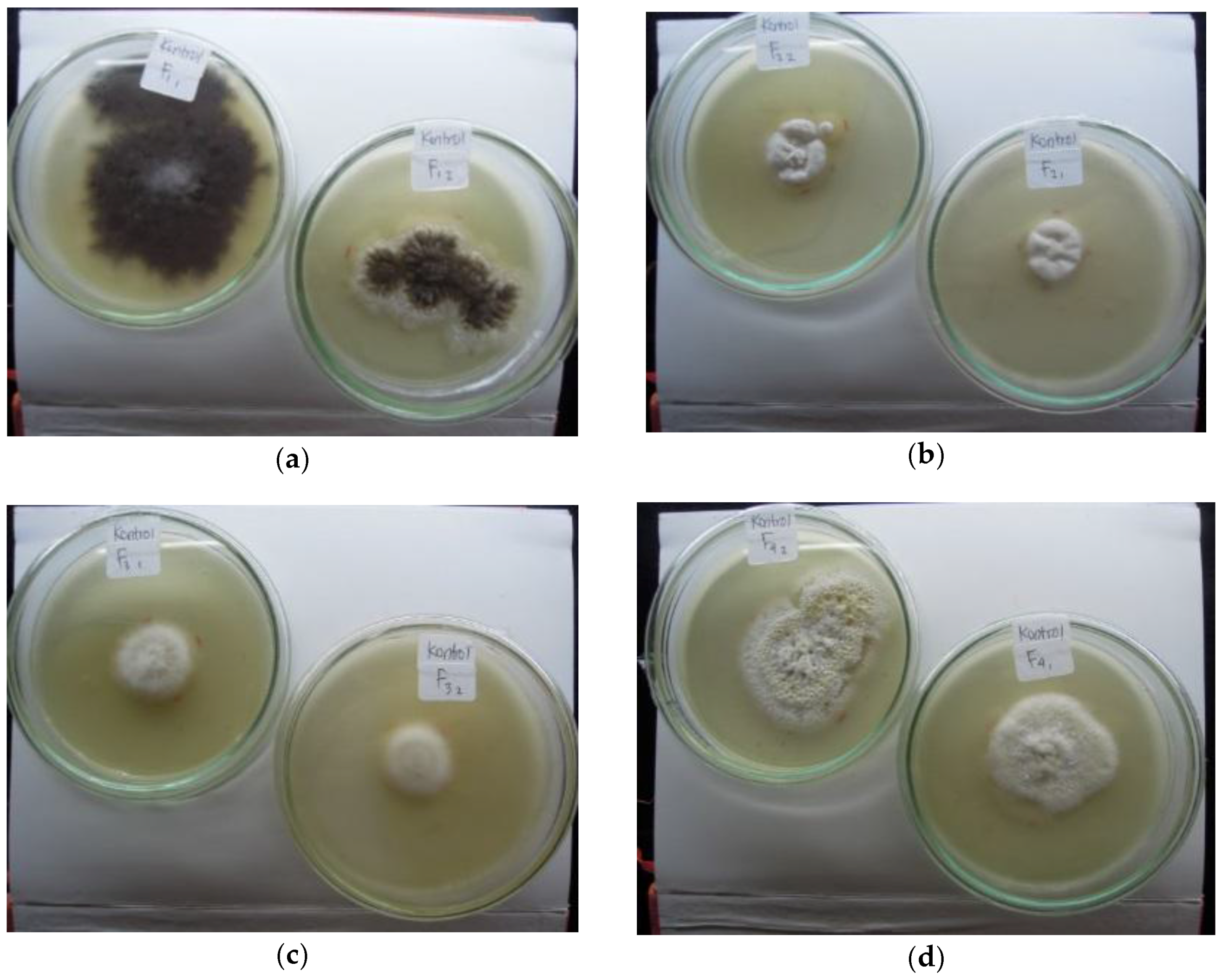
Four fungal isolates were isolated from the rhizosphere of pioneer plants grown in tailings deposited at Wamsait village, Buru Island. Isolates A and C were obtained from water spinach (Ipomoea reptans L.) rhizospheres, whereas isolates B and D were isolated from wiregrass (Eleusin indica (L.) Gaertn) rhizospheres. The macroscopic morphology of the four fungal colonies is shown in Figure 1.
Population density of all isolates in individual slants reached 109 propagules per mL of fungal suspension, based on the direct counting method using a counting chamber slide (hemocytometer). Before the study, the fungal density of each rhizosphere was counted using the dilution plate method. The results showed that the population reached up to 105 colony forming unit (CFU) g−1.
3.3. Mercury Resistant Fungus
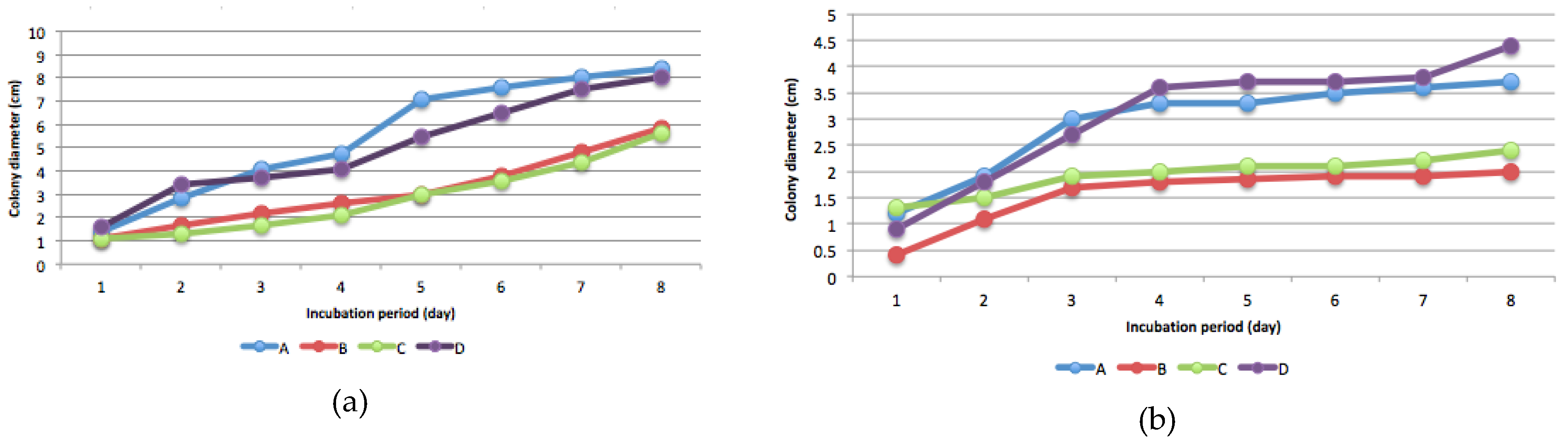
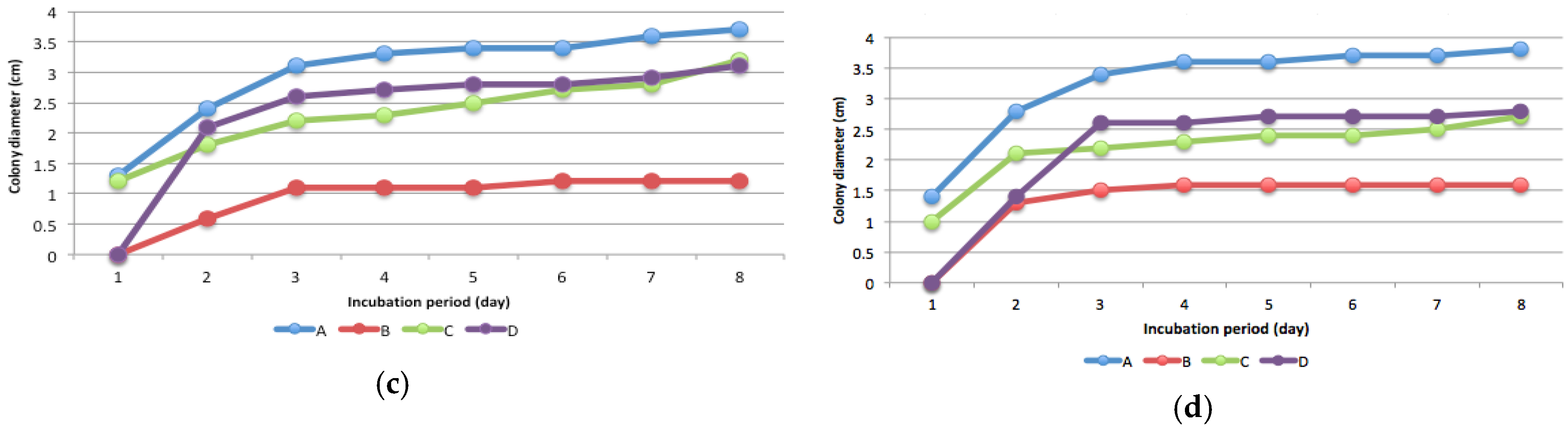
The presence of Hg-resistant fungus was determined by measuring colony diameter every 24 h for eight consecutive days after incubation. Isolates proliferated in potato dextrose plate agar containing 5, 15, and 25 mg L−1 Hg (Figure 2).
Isolate B was the most sensitive to high levels of Hg compared with the other isolates, whereas isolate C was the most resistant. The presence of Hg increased sporulation, as shown by the formation of a new colony (Figure 3). In media without Hg, the fungi grew in a single colony, but in media contaminated with Hg, multiple colonies formed (Figure 3), indicating increased spore production. In this study, sporulation depends on isolates and Hg levels. Generally, sporulation intensity increased at 15 mg kg−1 and 25 mg kg-1 Hg levels (Figure 3). Isolate D showed more intensive sporulation at every level of Hg compared to the other isolates. The ability of fungi to produce spores depended on the mercury level (Figure 3). Colony development of all four isolates was disturbed by every level of Hg, but in particular by 25 mg kg−1 Hg.
3.4. Fungal Species
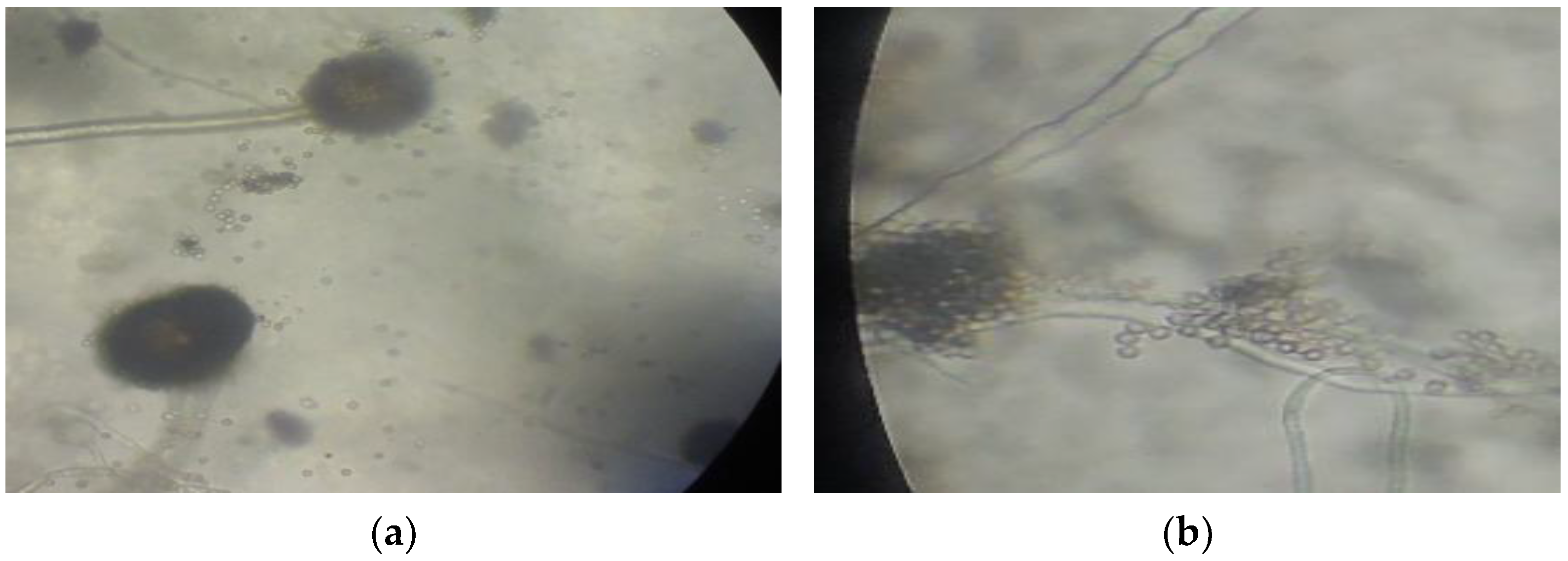
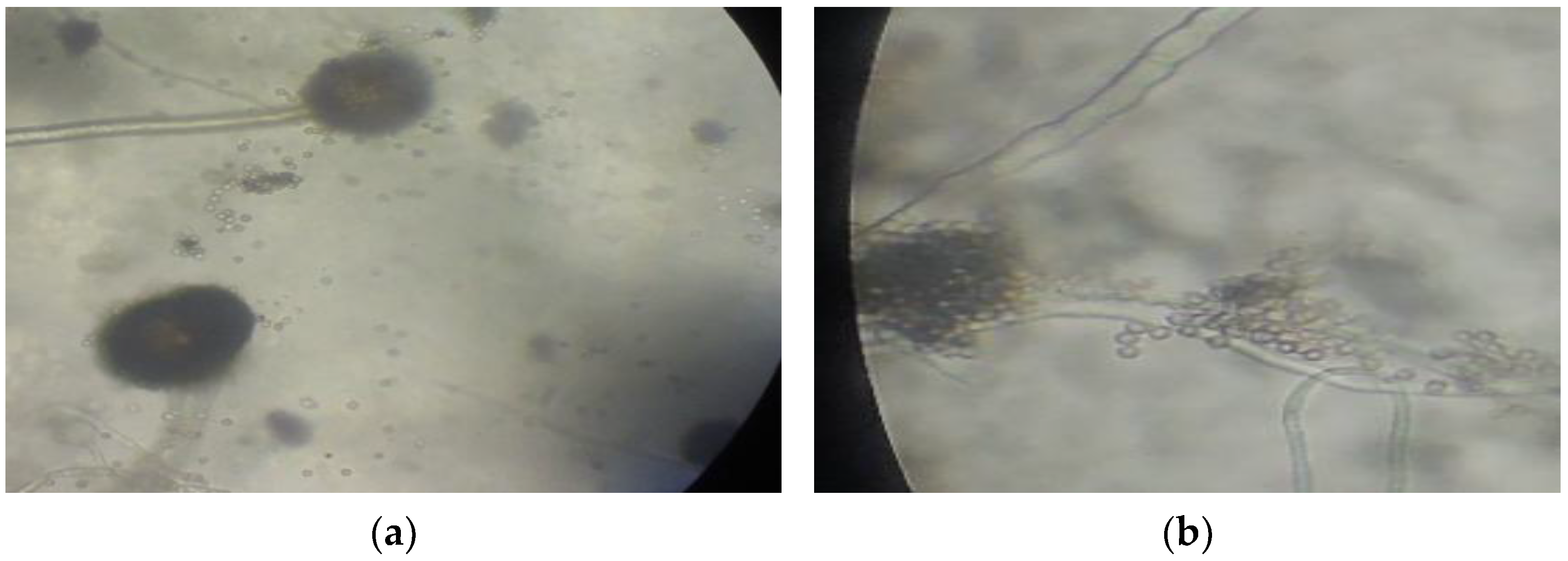
Due to their adaptability to the presence of mercury in the environment, fungi isolates A and D were characterized for species verification. Macroscopic and microscopic observation based on fungal characteristics described by Samson et al. [14] showed that isolate A was Aspergillus niger and isolate B was Aspergillus flavus (Figure 4). Both species belong to phylum Ascomycota, order Eurotiales, Family Trichocomaceae. Aspergillus niger and A. flavus are soil fungi that are often used as bioremediation agents for Hg-contaminated soil [9,10].
3.5. Effect of Fungal Inoculation on Hg in Soil and Biomass
At 15 days after planting, the height and dry weight of all treated plants were around 10–14 cm and 0.15–0.47 g, respectively (Table 1). Tailing has very low soil fertility [15] and the texture is not suitable for the nutrition requirements of Chinese cabbage. Organic matter and inorganic fertilizer in general could increase soil fertility. Therefore, in this bioassay, NPK fertilizer was applied during planting. During the experiment, the pH of growth media fluctuated; the 9.17 reading before the bioassay increased up to 9.39 after fungal inoculation, then decreased to 8.91 after transplanting. At the end of the bioassay, the recorded growth media pH was 7.72.
4. Discussion
The study of existing microorganisms in the Hg-contaminated area revealed that increased mercury concentrations in soil does not significantly correlate with the presence of indigenous fungi [16,17,18]. This group of microorganisms is involved in the change of Hg mobilization in soil. Fungi have the ability to retain Hg and decrease metal uptake through their roots; they can also secrete organic acid to increase heavy metal mobilization. The presence of fungal communities in Hg-contaminated soil also plays an important role in plant nutrition in particular, due to their role in the decomposition of organic matter and in the phosphorous cycle.
Fungal sensitivity towards heavy metals affects the formation of fungal spores [19]. Heavy metals react with sulfhydryl groups (-SH) of enzymes prior to the alteration of protein conformation, rendering them inactive [20] and restricting fungal growth.
Sporulation is a metal-avoidance strategy used by fungi in heavy metal-contaminated environments [21] as a natural response to nutrition depletion [22]. However, increasing Ni concentration induced sporulation in Aspergillus niger, Aspergillus giganteus, Penicillium vermiculatum, Gliocladium sp., Beauvaria sp., Trichoderma viride, and Rhizopus stolonifera [23]. In other experiments, the presences of Zn and Pb did not affect the fungal growth or sporulation at concentrations of 5 and 10 mg L−1 [24]. The increasing growth rate for Pb2+ and Ni3+ when compared to Cu2+ and Zn2+ concentrations and the lack of changes in mycelium growth and sporulation indicated that the fungi, several strains of T. harzianum, were resistant to those metals.
In our study, visible changes in appearance occurred in the mycelium or sporulation due to high levels of mercury (Figure 3). Fungi can metabolize heavy metals and reduce cytotoxicity by forming metallothionein polypeptides that detoxify Cd and Cu, since the polypeptide sequesters toxic metals in vacuoles [25]. Isolates A and D were the two most adaptable fungi in media containing high levels of Hg (Figure 3). One of the adaptation mechanisms towards heavy metals is changing the gene expression of enzymes and proteins [26]. Other mechanisms that microbes use to respond to heavy metals include utilizing it as an energy source by immobilizing it into the cell walls, producing a chelating agent such as metallothionein or phytochelatin, and changing microbial cell membrane permeability.
Increased acidity correlates with the presence of organic acids, released by the fungi. Aspergillus niger is acidophilic and known to produce organic acids such as gluconate, citrate, and oxalate [27]. The tailing pH was relatively high, preventing the Chinese cabbage from attaining good development and growth. The reduction of pH due to the release of organic acids by fungi created a more suitable environment for Chinese cabbage growth.
Soil fertility improvement to ensure fungal proliferation and activity is necessary so that the effects of either fungal inoculation or organic matter amendment on plant growth is more significant. In general, the amount of available Hg in soil increased following the fungal inoculation, especially in mixed inoculation treatment. Meanwhile, plants without fungal inoculation accumulated lower Hg in the tissue, indicating a fungal role in Hg mobilization. The fungi increased the amount of mobile Hg and cationic Hg species available for plant uptake. Further research is needed to verify certain biochemical mechanisms of mercury mobilization by soil fungi.
Acknowledgments
Authors thank the Head of Agricultural Department, Buru District, Maluku Province for the authorization to enter the gold mining site.
Author Contributions
Reginawanti Hindersah and Diyan Herdiyantoro conceived and designed the experiments; Reginawanti Hindersah and Khainur Rasyid Asda performed the experiments; Khainur Rasyid Asda and Diyan Herdiyantoro analyzed the data; Reginawanti Hindersah and Khainur Rasyid Asda contributed reagents/materials/analysis tools; Reginawanti Hindersah and Nadia Nuraniya Kamaluddin wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Porcella, D.B.; Ramel, C.; Jernelov, A. Global mercury pollution and the role of gold mining. Water Air Soil Pollut. 1997, 97, 205–207. [Google Scholar] [CrossRef]
- Wang, Y. Phytoremediation of Mercury by Terrestrial Plants. Ph.D. Thesis, Stockholm University, Stockholm, Sweden, 2004. [Google Scholar]
- Singh, R.; Gautam, N.; Mishra, A.; Gupta, R. Heavy metals and living systems: An overview. Indian J. Pharmacol. 2011, 43, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Steiness, E. Heavy Metals in Soils; Alloway, B.J., Ed.; Blackie Academic & Professional: London, UK, 1995. [Google Scholar]
- Steinnes, E.; Rühling, Å.; Lippo, H.; Mäkinen, A. Reference materials for large-scale metal deposition surveys. Accredit. Qual. Assur. 1997, 2, 243–249. [Google Scholar] [CrossRef]
- Bradl, H.B. Heavy Metal in Environment; Elsevier Academic Press: London, UK, 2005. [Google Scholar]
- Lasat, M.M. Phytoextraction of Toxic Metals. J. Environ. Qual. 2002, 31, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-P.; Shi, J.-Y.; Lin, Q.; Chen, X.-C.; Chen, Y.-X. Heavy metal availability and impact on activity of soil microorganisms along a Cu/Zn contamination gradient. J. Environ. Sci. 2007, 19, 848–853. [Google Scholar] [CrossRef]
- Kurniati, E.; Arfarita, N.; Imai, T.; Higuchi, T.; Kanno, A.; Yamamoto, K.; Sekine, M. Potential bioremediation of mercury-contaminated substrate using filamentous fungi isolated from forest soil. J. Environ. Sci. 2014, 26, 1223–1231. [Google Scholar] [CrossRef]
- Ul-Haq, I.; Javed, M.; Khan, T. Sugar cane bagasse pretreatment: An attempt to enhance the production potential of cellulases by Humicola insolensTAS-13. Biokemistri 2006, 18, 83–88. [Google Scholar] [CrossRef]
- Akinpelu, E.A. Bioremediation of Gold Mine Wastewater Using Fusarium oxysporum. Master’s Thesis, Cape Paninsula University of Technology, Cape Town, South Africa, 2014. [Google Scholar]
- Dave, S.R. Microbial Interactions with Inorganic Pollutants: Acid Mine Drainage, Microbial Accumulation of Heavy Metals and Radionuclides. Applied Microbiology, E-Book Chapter. 2008. Available online: http://nsdl.res.in/handle/123456789/646 (accessed on 27 March 2017).
- Awasthi, A.K.; Pandey, A.K.; Khan, J. A preliminary report of indigenous fungal isolates from contaminated municipal solid waste site in India. Environ. Sci. Pollut. Res. 2017, 24, 8880–8888. [Google Scholar] [CrossRef] [PubMed]
- Samson, A.R.; van Reenen Hoekstra, E.S. Introduction to Food Borne Fungi; Centralbureau Voor Schimmelcultures: Delft, The Netherlands, 1988. [Google Scholar]
- Alloway, B.J. The Origin of heavy metals in soils. In Heavy Metals in Soils; Alloway, B.J., Ed.; Blackie Academic & Professional: Glasgow, Scotland, 1995. [Google Scholar]
- Baldrian, P.; Wiesche, C.I.D.; Gabriel, J.; Nerud, F.; Zadrazil, F. Influence of Cadmium and Mercury on Activities of Ligninolytic Enzymes and Degradation of Polycyclic Aromatic Hydrocarbons by Pleurotus ostreatus in Soil. Appl. Environ. Microbiol. 2000, 66, 2471–2478. [Google Scholar] [CrossRef] [PubMed]
- Jean-Philippe, S.R.; Franklin, J.A.; Buckley, D.S.; Hughes, K. The Effect of Mercury on Trees and Their Mycorrhizal Fungi. Ph.D. Thesis, University of Tennessee, Knoxville, TN, USA, 2010. [Google Scholar]
- Dixit, R.; Wasiullah, E.; Malaviya, D.; Pandiyan, K.; Singh, U.; Sahu, A.; Shukla, R.; Singh, B.; Rai, J.; Sharma, P.; et al. Bioremediation of Heavy Metals from Soil and Aquatic Environment: An Overview of Principles and Criteria of Fundamental Processes. Sustainability 2015, 7, 2189–2212. [Google Scholar] [CrossRef]
- Hepper, C.M.; Smith, G.A. Observation’s on the germination of Endogone spore. Trans. Br. Mycol. Soc. 1976, 66, 189–194. [Google Scholar] [CrossRef]
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: Activity, action, and resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [PubMed]
- Pawlowska, T.E.; Charvat, I. Heavy-Metal Stress and Developmental Patterns of Arbuscular Mycorrhizal Fungi. Appl. Environ. Microbiol. 2004, 70, 6643–6649. [Google Scholar] [CrossRef] [PubMed]
- Moat, A.G.; Foster, J.W.; Spector, M.P. Microbial Physiology, 4th ed.; Wiley-Liss Inc.: New York, NY, USA, 2002. [Google Scholar]
- Raspanti, E.; Cacciola, S.O.; Gotor, C.; Romero, L.C.; García, I. Implications of cysteine metabolism in the heavy metal response in Trichoderma harzianum and in three Fusarium species. Chemosphere 2009, 76, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Babich, H.; Gamba-Vitalo, C.; Stotzky, G. Comparative toxicity of nickel to mycelial proliferation and spore formation of selected fungi. Arch. Environ. Contam. Toxicol. 1982, 11, 465–468. [Google Scholar] [CrossRef]
- Lanfranco, L. Differential Expression of a Metallothionein Gene during the Presymbiotic versus the Symbiotic Phase of an Arbuscular Mycorrhizal Fungus. Plant Physiol. 2002, 130, 58–67. [Google Scholar] [CrossRef] [PubMed]
- De Almeida Paula Figueira, E.M.; Lima, A.I.G.; Pereira, S.I.A. Cadmium tolerance plasticity in Rhizobium leguminosarum bv. viciae: Glutathione as a detoxifying agent. Can. J. Microbiol. 2005, 51, 7–14. [Google Scholar]
- Magnuson, J.K.; Lasure, L.L. Advances in Fungal Biotechnology for Industry, Agriculture, and Medicine; Tkacz, J.S., Lange, L., Eds.; Springer: Boston, MA, USA, 2004. [Google Scholar]
Figure 1.
Four isolates of indigenous fungi colonies: (a) isolate A (two replications: F11 and F12); (b) isolate B (two replications: F21 and F22); (c) isolate C (two replications: F31 and F32); (d) isolate D (two replications: F41 and F42) from the rhizospheres of pioneer plants grown on tailings.
Figure 1.
Four isolates of indigenous fungi colonies: (a) isolate A (two replications: F11 and F12); (b) isolate B (two replications: F21 and F22); (c) isolate C (two replications: F31 and F32); (d) isolate D (two replications: F41 and F42) from the rhizospheres of pioneer plants grown on tailings.
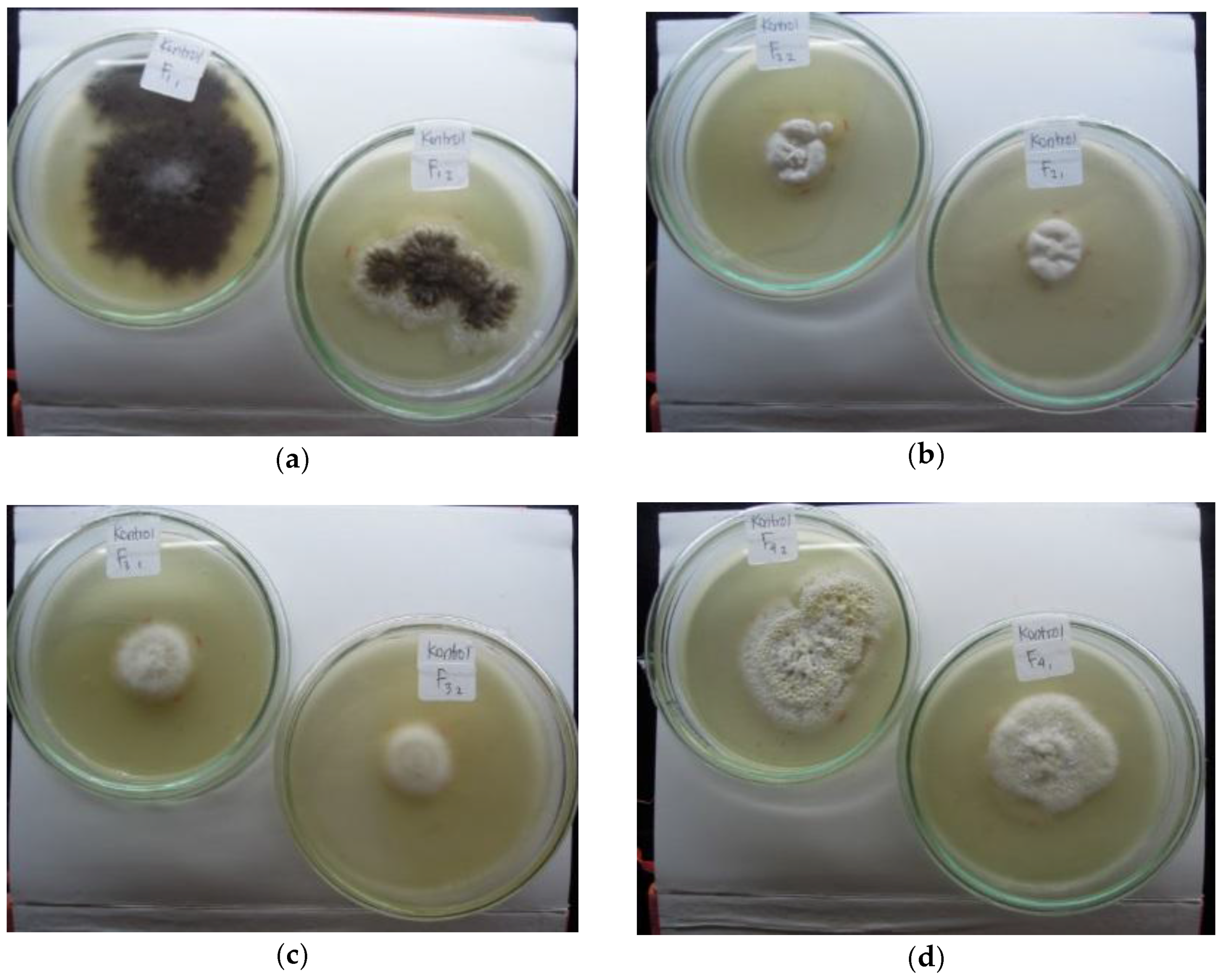
Figure 2.
Colony diameter of fungi isolates A, B, C, and D in potato dextrose plate agar: (a) 0 mg/kg mercury; (b) 5 mg/kg mercury; (c) 15 mg/kg mercury; (d) 25 mg/kg mercury.
Figure 2.
Colony diameter of fungi isolates A, B, C, and D in potato dextrose plate agar: (a) 0 mg/kg mercury; (b) 5 mg/kg mercury; (c) 15 mg/kg mercury; (d) 25 mg/kg mercury.
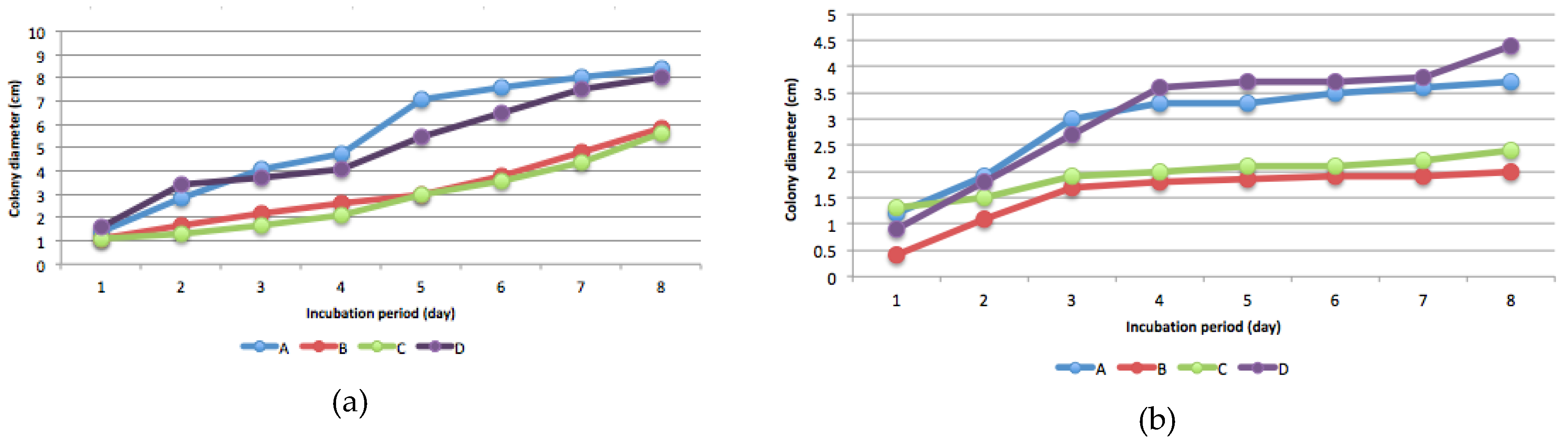
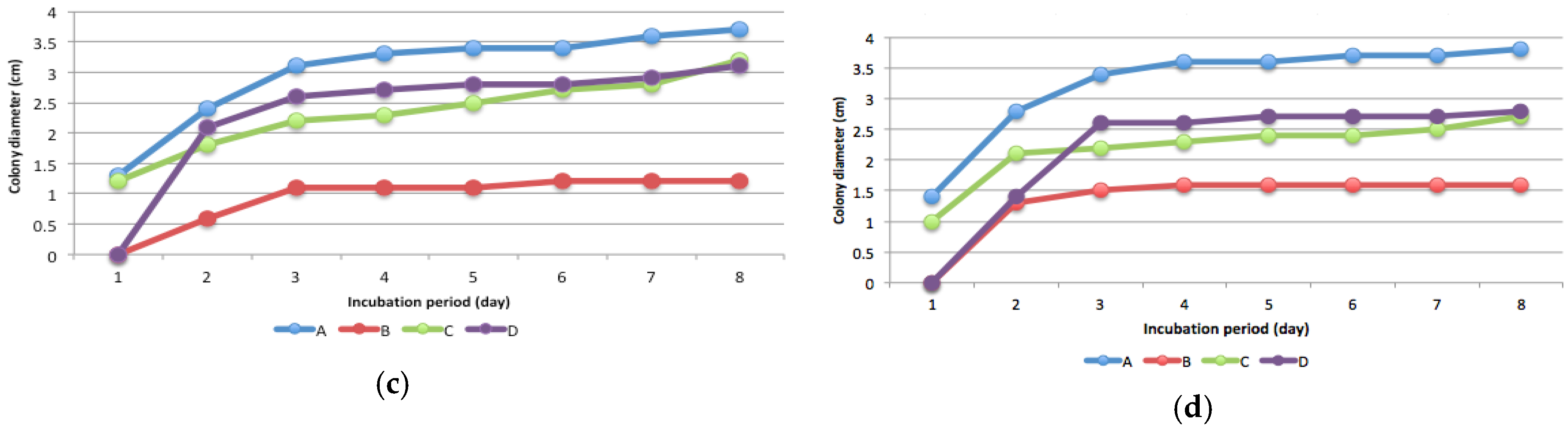
Figure 3.
Colony growth and sporulation of fungal isolates A, B, C, and D (two replications) at eight days after incubation in plate agar (a) without and with the addition of (b) 5; (c) 15; and (d) 25 mg kg−1 mercury.
Figure 3.
Colony growth and sporulation of fungal isolates A, B, C, and D (two replications) at eight days after incubation in plate agar (a) without and with the addition of (b) 5; (c) 15; and (d) 25 mg kg−1 mercury.

Figure 4.
Microscopic characteristics of (a) A. niger and (b) A. flavus at 400× magnification.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effect of fungal inoculation and organic matter amendment on plant growth, available Hg in soil, and Hg accumulation after 15 days of planting.
Table 1.
Effect of fungal inoculation and organic matter amendment on plant growth, available Hg in soil, and Hg accumulation after 15 days of planting.
| Treatments | Plant Height ** (cm) | Dry Weight ** (g) | Available Hg in Soil ** (µg kg−1) | Hg Content in Biomass ** (mg kg−1) |
|---|---|---|---|---|
| Without fungi and OM * | 10.50 | 0.23 | 125.86 | 1.35 |
| Isolate A; 1.5% OM | 11.07 | 0.30 | 115.22 | 3.63 |
| Isolate D, 1.5% OM | 11.43 | 0.23 | 137.13 | 7.48 |
| Isolate A + D; 1.5% OM | 13.43 | 0.33 | 162.81 | 6.20 |
| Without fungi; 3% OM | 11.53 | 0.26 | 121.48 | 1.93 |
| Isolate A; 3% OM | 12.07 | 0.44 | 120.23 | 3.63 |
| Isolate D, 3% OM | 12.07 | 0.33 | 14.,27 | 0.63 |
| Isolate A + D; 3% OM | 10.97 | 0.15 | 170.95 | 2.04 |
| Without fungi; 4.5% OM | 14.00 | 0.47 | 110.21 | 1.75 |
| Isolate A; 4.5% OM | 11.27 | 0.28 | 130.87 | 4.51 |
| Isolate D, 4.5% OM | 10.43 | 0.15 | 15717 | 8.85 |
| Isolate A + D; 3% OM | 10.57 | 0.27 | 181.56 | 0.40 |
* OM, organic matter; ** Mean of two replicates; isolate A, Aspergillus niger; isolate B, Aspergillus flavus.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hindersah, R.; Asda, K.R.; Herdiyantoro, D.; Kamaluddin, N.N. Isolation of Mercury-Resistant Fungi from Mercury-Contaminated Agricultural Soil. Agriculture 2018, 8, 33. https://doi.org/10.3390/agriculture8030033
AMA Style
Hindersah R, Asda KR, Herdiyantoro D, Kamaluddin NN. Isolation of Mercury-Resistant Fungi from Mercury-Contaminated Agricultural Soil. Agriculture. 2018; 8(3):33. https://doi.org/10.3390/agriculture8030033
Chicago/Turabian StyleHindersah, Reginawanti, Khainur Rasyid Asda, Diyan Herdiyantoro, and Nadia Nuraniya Kamaluddin. 2018. "Isolation of Mercury-Resistant Fungi from Mercury-Contaminated Agricultural Soil" Agriculture 8, no. 3: 33. https://doi.org/10.3390/agriculture8030033
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.