1. Introduction
Verticillium wilt of olive (VWO) is one of the most serious diseases affecting olive (
Olea europaea L. subsp.
europaea var.
europaea) cultivation worldwide.
Verticillium dahliae Kleb., the causal agent of VWO, is responsible for substantial production losses thereby constituting the main phytopathological issue in many olive-growing regions. Multiple factors explain the current spread and importance of the disease: the long-term endurance of microsclerotia (resistance structures of
V. dahliae) in soil, the broad host range of the pathogen, the vascular nature of the disease, inappropriate agronomical practices, etc. [
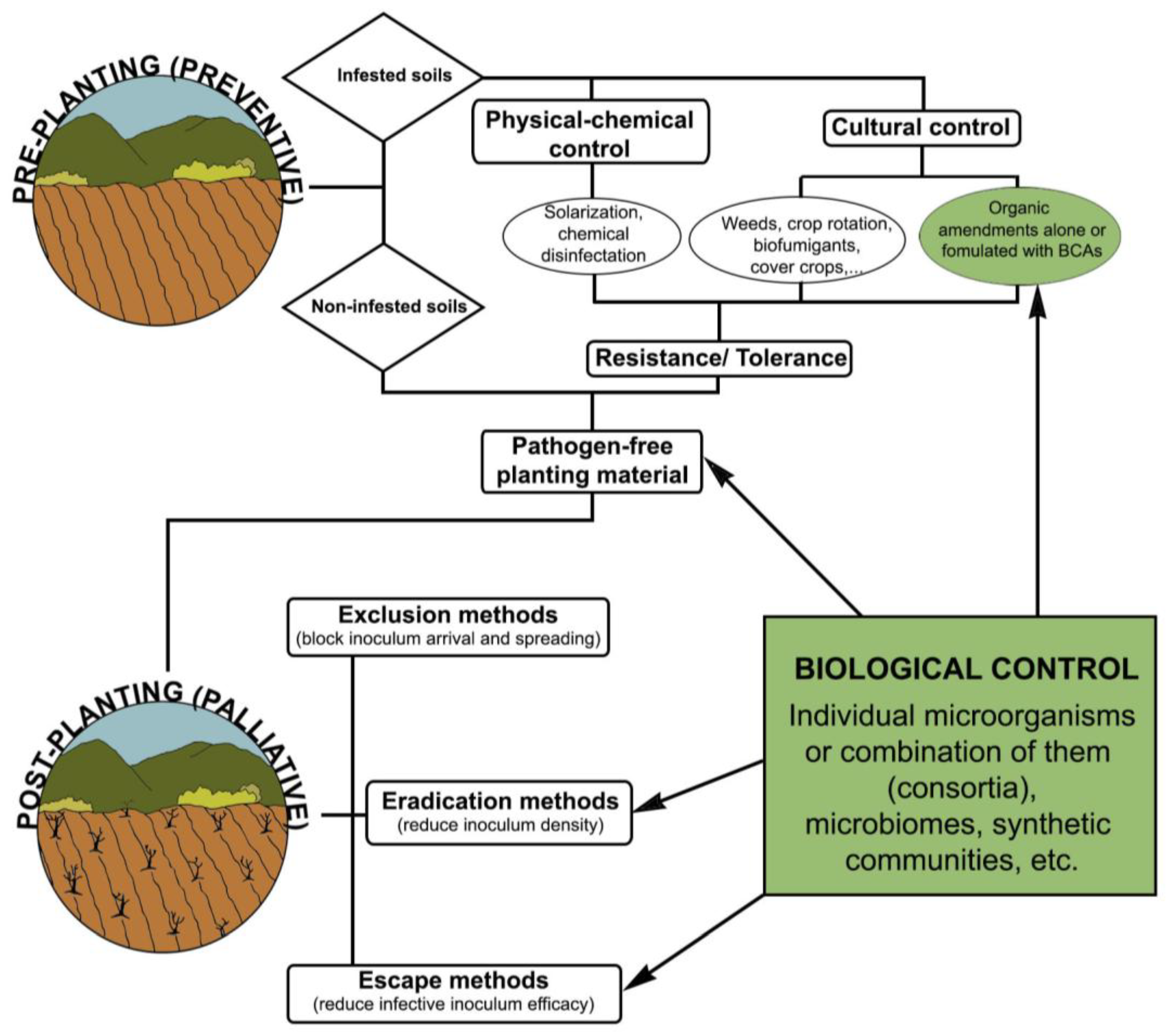
1]. All these factors make it difficult for an efficient control of VWO, and available measures have so far proven unsuccessful when implemented individually. Therefore, an integrated disease management strategy is recommended, highlighting pre-plating control tools since no resistant varieties and/or effective fungicides are available. Within this context, the development and use of biological control agents (BCAs) is an appealing, sustainable and environmentally friendly management approach to control VWO.
Biocontrol experiments are often carried out using sterile soils/substrates; therefore, major determinants influencing results are nutrient availability and/or soil physico-chemical conditions [
2]. Moreover, these bioassays are frequently performed using plants artificially inoculated via root dipping in (or soil drenching with) the pathogen’s conidia suspensions. However, this experimental scenario is quite different to the situation taking place under field conditions. For instance, in sterile soils there is no competence with other microorganisms and the BCA can easily be established. When several microorganisms are used at the same time under these very specific (and artificial) conditions, the final outcome is strongly determined by the inherent capability of each microorganism to multiply and prevail over the others. In contrast, under natural soil conditions, the introduced BCAs face a very complex situation in a harsh environment considered as one of the most microbiologically diverse on earth [
3]. This is particularly true when the introduced microorganism is not adapted to the target ecological niche.
The use of BCAs under greenhouse or nursery conditions is easier, cheaper, and less time-consuming than large-scale field treatments which, in addition, often suffer from inconsistency or lack of effectiveness. Hence, implementing biological control measures to protect woody plants (e.g., olive) at the propagation stage (i.e., greenhouses or nurseries) against soil-borne pathogens like
V. dahliae appears as a preventive and recommendable approach. This ‘pretreatment strategy’ can be performed before olive plants are transferred to the field, usually cultivated in large growing areas where biocontrol approaches are often unfeasible both from economical and agronomical perspectives [
4].
Our current knowledge on fungi, oomycetes and bacteria as effective BCAs against
V. dahliae infecting different crops is steadily increasing (reviewed by [
5]). Berg et al. [
6] already demonstrated that naturally occurring rhizobacteria are effective BCA against
V. dahliae. Zhengjun et al. [
7] reported on rhizosphere and endophytic bacteria able to control Verticillium wilt of cotton. Berg and Lottmann [
8] informed that Verticillium wilt of oilseed rape could be controlled by a
Stenotrophomonas maltophilia (Hugh) Palleroni and Bradbury. strain. Tjamos et al. [
9] selected two
Bacillales strains,
Bacillus sp. 5-27 and
Paenibacillus alvei K-165, from root tips of tomato plants grown in solarized soils able to inhibit
V. dahliae’s growth and to successfully control Verticillium wilt. The use of strain K-165 also reduced germination of
V. dahliae microsclerotia in root tips and in the root elongation zone of eggplants [
10]. Malandraki et al. [
11] selected two strains of the
Pseudomonas fluorescens Migula. complex and two isolates of the
Fusarium oxysporum Schltdl. complex, all capable to reduce disease severity in eggplants. Likewise, Veloso and Díaz [
12] demonstrated that inoculation of pepper plants with the nonpathogenic isolate
F. oxysporum 47 reduced Verticillium wilt blight symptoms. Regarding biocontrol of VWO, advances are still limited, although some examples have shown promising results. For instance, olive root-associated fluorescent pseudomonads have been demonstrated to control VWO caused by the highly virulent, defoliating (D) pathotype of
V. dahliae in young, nursery-produced plants [
13,
14,
15,
16].
Paenibacillus alvei K-165 has also been shown to suppress VWO under both greenhouse and field conditions [
17]. Finally,
Acetobacter aceti VIN02 reduced the percentage of mortality in
V. dahliae-inoculated plants to the level observed in non-inoculated plants [
18]. However, even though several microorganisms have been shown to be effective BCAs against Verticillium wilt, hardly any of them are available as biopesticides against
Verticillium in Europe [
19].
Disease suppressive soils, organic amendments, and healthy plants enduring in pathogen-infested fields are interesting reservoirs of potential BCAs [
5,
11,
16,
20]. Microbial communities associated with plant roots are complex and diverse, and they strongly influence the health and development of the host [
21]. Moreover, the rhizosphere microbiome plays a significant role in reprogramming defense responses of plants [
22]. Therefore, to look for beneficial indigenous microbial inhabitants adapted to the conditions found in the olive root/rhizosphere seems to be a good strategy to identify novel BCAs candidates against VWO. In a previous study, we designed a holistic approach to identify, characterize and evaluate new and effective BCAs originating from the roots of healthy nursery-propagated olive plants [
16]. This strategy was based on the isolation from the target niche, in vitro antagonism assays against relevant olive pathogens (
V. dahliae, Phytophthora cinnamomi Rands.,
Pseudomonas savastanoi (Janse) Gardan et al.), phenotypic and metabolic characterization in order to identify traits associated with biological control or plant growth promotion (PGP), in planta experiments under non-gnotobiotic conditions to assess their effectiveness against the D pathotype of
V. dahliae, genome sequencing and in silico analysis, taxonomic identification by multi-locus sequence analyses (MLSA), and evaluation of root colonization abilities [
16]. Eventually, we were able to identify and in-depth characterized three novel Gram-negative resident olive rhizobacteria belonging to the
Pseudomonas genus (namely, strains PIC25, PIC105 and PICF141) showing good biocontrol ability against VWO. Current research trends in biocontrol aim to investigate the role and effectiveness of microbial consortia, synthetic communities and even tailored microbiomes in promoting plant growth and/or defense against pathogens [
23,
24]. The use of bacterial consortia to control soil-borne pathogens may have the advantage over single microbe-based formulations to enhance biocontrol effectiveness. Indeed, the introduction of diverse beneficial microbes may facilitate the colonization of different microhabitats on/in roots thereby enhancing competition for colonization sites used by the pathogen. Furthermore, a range of biocontrol mechanisms can be deployed by each BCA in the consortium contributing in this manner to improved disease suppressiveness [
25].
This study aims to identify and characterize Gram-positive bacteria from the olive root/rhizosphere with potential as BCAs against olive pathogens, with emphasis on V. dahliae. We implemented the same comprehensive strategy that previously yielded successful results when screening and evaluating indigenous beneficial Pseudomonas spp. An additional aim was to in vitro assess whether combinations of these previously verified BCAs and the newly described antagonists here reported, all adapted to the olive rhizosphere, could be formulated as consortia to control VWO.
4. Discussion
The development and use of BCAs against
V. dahliae is an attractive measure within an integrated management strategy of VWO. The early introduction of the BCA, for instance at the nursery propagation stage, could ‘condition’ the plant to better confront subsequent infections by the pathogen’s propagules existing in the planting site. Moreover, selection of BCAs sharing the same ecological niche as
V. dahliae is a plausible approach since these microbial antagonists can compete with the pathogen for infection sites, space and nutrients [
5]. For instance, in the tripartite interaction
V. dahliae-olive-
P. fluorescens PICF7, niche overlap between the BCA and the pathogen
in planta was necessary for effective biocontrol [
52]. Among the bacteria thriving in the soil/rhizosphere, some spore-forming plant growth-promoting rhizobacteria (PGPR) and/or BCAs have gained attention due to their advantages over non-spore-formers in bioproduct formulation and stable maintenance in soil. Moreover, they are usually persistent to adverse conditions and can be preserved and distributed easily [
53].
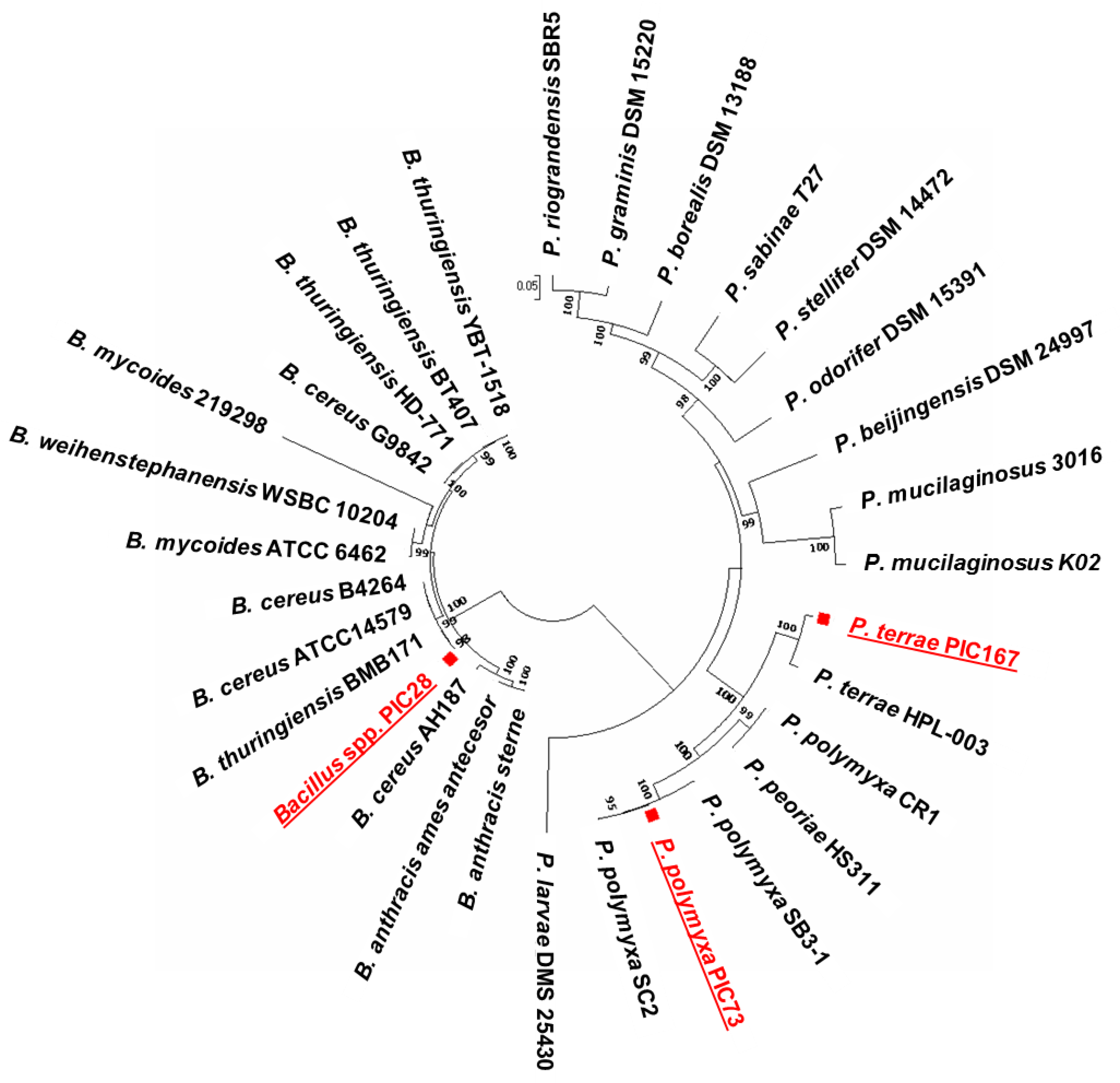
The holistic strategy previously designed to identify
Pseudomonas spp. strains from the olive root/rhizosphere as BCAs against VWO [
16], has also been successfully implemented to in-depth characterize Gram-positive bacteria effective against the D, highly virulent pathotype of
V. dahliae. Eventually, three
Bacillales members were selected (PIC28, PIC73 and PIC167) according to their ability to in vitro antagonize
V. dahliae isolates and other olive pathogens. The MLSA performed enabled to identify strain PIC73 as
P. polymyxa and strain PIC167 as
P. terrae. However, strain PIC28 could only be assigned to the genus
Bacillus and further analysis will be needed to determine the species level. Overall, results obtained from the phenotypic and metabolic analyses (e.g., Biolog Gen III) performed in our study confirmed the similarities between the two
Paenibacillus strains.
The use of spores of
Bacillus spp. as natural ‘biopesticides’ has previously been described as a safe and environmentally friendly alternative to traditional control methods.
Bacillus cereus and
B. subtilis (Ehrenberg) Cohn. [
54] strains showed fungicidal and PGP ability when they were inoculated directly into soils as seed pretreatments. Moreover, the high survival rate of their spores in soils play an important role in its effectiveness ([
55], and references therein). Likewise, one advantage of treating fields with endospore-forming bacteria such as
Paenibacillus spp. is their capability to persist for long periods of time in the soil under harsh environmental conditions [
56]. Representatives of the genus
Bacillus has been demonstrated as BCAs of Verticillium wilts, highlighting
Bacillus amyloliquefaciens Priest et al. and
B. subtilis strains. For instance,
B. amyloliquefaciens 5–127 reduced the percentage of diseased leaves in eggplants challenged with
Verticillium dahliae in the greenhouse, and also decreased the DI in potato under field conditions [
9].
Bacillus strains have been also demonstrated to be effective against Verticillium wilt in oilseed rape, cotton and strawberry [
5]. Only a few studies have been focused on biological control of Verticillium wilt by
Bacillales members in woody plants. Thus, some
B. subtilis strains isolated from healthy maple trees were checked in a greenhouse against
V. dahliae in this tree species, and disease reduction was reported [
57]. The genus
Paenibacillus is currently gaining interest as BCA of a range of plant diseases [
58,
59]. For instance,
P. alvei K-165 obtained from tomato plants cultivated in solarized soil [
9] showed effective biocontrol of
V. dahliae in eggplant under greenhouse conditions [
4,
9,
10,
60]. This strain was also able to control
V. dahliae in potato plants under field conditions and in olive trees under both field and greenhouse conditions [
9,
17]. In cotton, treatment with
Paenibacillus xylanilyticus YUPP-1 and
P. polymyxa YUPP-8 reduced DI and disease severity [
61].
Results from in vitro antagonism assays showed differences depending on the culture medium used (PDA or NA). The influence of the culture media on the results of in vitro antagonism tests has been previously reported [
16,
62], a likely consequence of the effects on antagonistic bacteria of specific nutritional factors present in the used media. According to our in vitro antagonism tests, the most promising olive rhizobacteria to be used as BCAs against VWO would be the
Paenibacillus spp. strains, particularly strain PIC73. Yet, these results are not enough to qualify them as effective BCAs since colonization of the target niche and biocontrol performance by a selected antagonist will be influenced by environmental factors (moisture, nutrients, pH, temperature, etc.) and its ability to compete with preexisting microbial communities present in the site where it will be deployed. It is well known that the capacity of a beneficial bacterium to act as a BCA could vary between laboratory and field conditions, and that the best in vitro antagonist is not always the best BCA [
63,
64,
65,
66]. Therefore, in planta bioassays are important to qualify an in vitro antagonist as a true and effective BCA. Our independent VWO biocontrol experiments (performed under non-gnotobiotic conditions) showed that the three
Bacillales members indigenous from olive roots were able to significantly reduce VWO development, and that their disease control ability was comparable to that observed for
Pseudomonas spp. strains well characterized as VWO BCAs [
15,
16]. Furthermore, all of the tested BCAs are well adapted to the olive root/rhizosphere since they originated from this niche. However, that biocontrol performance slightly decreased when the disease pressure was high, particularly in the case of
Bacillus sp. PIC28. Therefore, it is highly recommended that different experiments are conducted to obtain an accurate picture of the biocontrol performance of any BCA candidate.
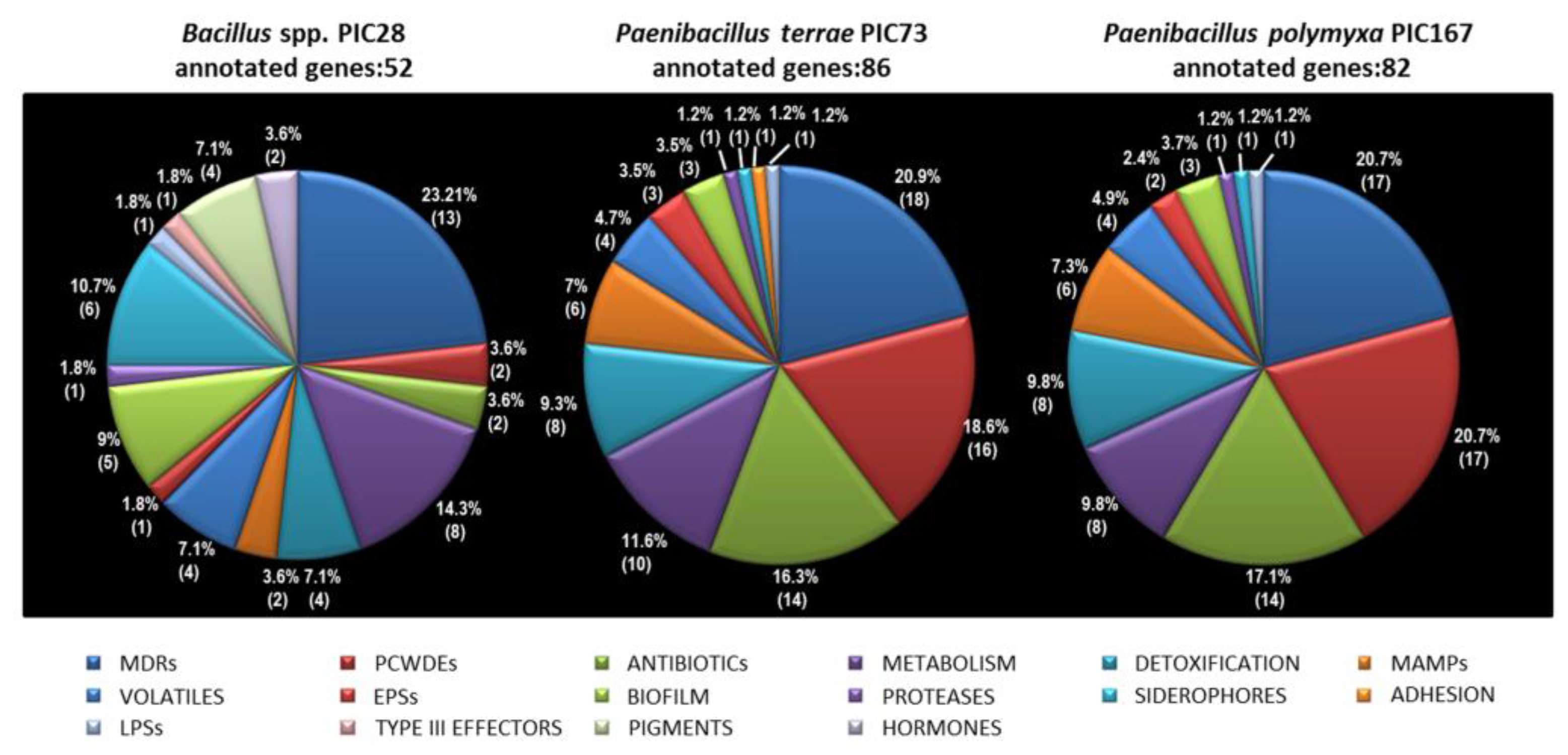
Biocontrol activity of PGPR relies, among other mechanisms, on the production of different cell wall-degrading enzymes (e.g., chitinase, protease/elastase, and β-1,3 glucanase, etc.) that play an important role in degrading the cell walls of phytopathogens [
67,
68]. Some of these enzymes also degrade plant cell walls and some of their components can be utilized by BCAs, thereby enhancing their competitiveness during the root colonization process. The three
Bacillales strains here studied were positive for protease, cellulase and β-glucosidase. Proteases play a significant role in cell wall lysis of phytopathogenic fungi, since proteolytic activity is a prerequisite to lyse whole fungal cells [
69]. Microorganisms displaying cellulase and protease activities do not only play a role in organic matter decomposition and PGP, but also contribute to disease suppression by inhibiting fungal growth [
70]. Only
Paenibacillus spp. strains PIC73 and PIC167 strains showed chitinase, amylase and xylanase activities. Chitinases lyse fungal cell wall through degradation of the chitin polymer present in fungal cell walls [
71]. Strain PIC73 was the only one to show phosphatase and glucanase activities, although at very low levels. Phosphatases mediate the release of inorganic P from organically bound P returned to soil [
72], increasing the root system and thus contributing to the growth of the plant. Glucanases participate in the degradation of cell walls in fungi, yeasts and plants [
73]. Complementing this phenotypic characterization, in silico analysis revealed that the genome of
Bacillus sp. PIC28 harbored a considerably lower number of genes potentially coding for cell wall-degrading enzymes (as well as antibiotics) than the two
Paenibacillus strains. The ability to produce fungal cell wall-degrading enzymes and a broad spectrum of antibiotics seem to be common to aerobic spore-forming bacteria [
74].
Paenibacillus terrae PIC167 was the only strain displaying phytase activity. Phytases catalyze the hydrolysis of phytic acid, releasing a utilizable form of inorganic phosphorus (P) to the plants [
75]. Rhizosphere bacteria with phytase activity have been isolated and proposed to promote plant growth in soils with high content of organic P [
76]. Another trait traditionally associated with PGPR is the production of the phytohormone IAA. In silico analysis revealed the presence of two genes related to IAA biosynthesis in the genome of
Bacillus sp. PIC28. However, production of IAA by this strain could not be demonstrated. Some PGPR elicit induced systemic resistance by emission of volatile organic compounds including short chain alcohols, acetoin, and 2,3-butanediol [
77]. In silico analysis predicted the ability to produce several volatile compounds, including 2,3-butanediol, for the three
Bacillales strains. Indeed, production of this volatile was confirmed for all of them; 2,3-butanediol produced by
Bacillus spp. is suggested to be involved in plant protection [
78]. Different types of siderophores promote the growth of several plant species and increase their yield by enhancing iron uptake in plants [
79]. Additionally, siderophore-producing microorganisms can suppress some soil-borne fungal pathogens through a siderophore-mediated iron competition mechanism. Genes putatively involved in siderophore(s) biosynthesis were detected by in silico analysis of the genomes of the three
Bacillales strains. Moreover, siderophore activity was confirmed by the CAS (Chrome Azurol S) for all of them (
Table 3),
Bacillus sp. PIC28 always showing much bigger orange halos than those observed for the
Paenibacillus strains.
Biofilm formation by beneficial strains of
Bacillus spp. and
Paenibacillus spp. is known to improve their biocontrol abilities [
80,
81,
82]. Biofilms facilitate the development of a microhabitat that protects bacteria against a/biotic stresses [
83,
84], enhancing root competitiveness of bacteria as well. The presence of genetic factors related to biofilm development has been predicted for strains PIC28, PIC73 and PIC167, although the number of genes related to this trait was higher in strain PIC28. The presence of TSSs in the newly identified
Bacillales members of the olive roots was also investigated by in silico analysis. Secretion systems in biocontrol bacteria have been related to plant root colonization, rhizosphere competence, environmental competition or defense against pathogens [
85,
86,
87]. According to ‘T346hunter’ predictions, only the flagellar-T3SS was detected in the three genomes analyzed. It is worth mentioning that ‘T346hunter’ and ‘PIFAR’ web-based tools only detect positive matches for (nearly) complete gene clusters, and that factors defined by several genes are reported only if at least 90% of such genes are identified within a given genome [
47,
48]. Many Gram-positive bacteria also produce a specialized protein secretion machinery termed the type VII secretion system (T7SS) that is not detected by the ‘T346Hunter’ tool. Genes related to T7SS have been detected during the examination of the annotated genomes. T7SS are widespread in representatives of the phlya
Actinobacteria and
Firmicutes, and affect a range of bacterial processes including virulence, sporulation, conjugation, and cell wall stability [
88]. Bioinformatics analyses have also predicted that this TSS is present in non-pathogenic bacteria [
89]. WXG-100-encoding genes have also been identified in different species of the genus
Bacillus, both in the non-pathogenic species
B. subtilis and in pathogenic representatives of
B. cereus,
B. thuringiensis and
Bacillus anthracis Cohn. [
90,
91]. Although some T7SSs can contribute to virulence, not all of these systems function in virulence or even in secretion of substrates. However, the biological functions of several type VII-secreted substrates and effector molecules are still being investigated. Finally, when the annotated genomes here reported were manually and thoroughly checked, genes putatively coding for pathogenicity/virulence factors were found,
Bacillus sp. PIC28 being the strain harboring the largest number of genes potentially related with deleterious, non-desirable effects including three genes involved in enterotoxin synthesis. This fact must be carefully examined regarding the use of this strain in biocontrol frameworks.
Recent trends in biocontrol of plant diseases have focused on the development and use of consortia of beneficial microorganisms [
25,
92]. This approach may improve the efficacy, reliability and consistency of individual microbe-based biocontrol tools [
93]. For instance, the mixture of two
Bacillus strains increased protection in tomato and pepper against different pathogens [
94]. Also, the application of a consortium of three rhizobacteria,
B. cereus AR156,
B. subtilis SM21 and
Serratia sp. XY21, resulted in higher biocontrol efficacy against Verticillium wilt in cotton compared to the outcomes observed with individual treatments [
95]. To be formulated in a consortium, microorganisms in the mixture must not show antagonism among them [
96,
97,
98]. Olive roots are a powerful reservoir of BCAs against VWO, mainly
Pseudomonas spp. [
13,
16] and
Bacillales (this study) strains. In order to check whether these BCAs could be formulated as consortia, the absence of antagonism among them was assessed. Results showed that, for instance, a consortium consisting of
Pseudomonas spp. strains was feasible since no incompatibilities were detected among them. Moreover, a consortium based on the two most effective
Pseudomonas strains (PICF7 and PICF141) [
15,
16] and
P. polymyxa strain PIC73 would also be interesting to be tested in future biocontrol assays. These combinations would provide an array of BCAs colonizing different microhabitats or sites of the olive root/rhizosphere, thereby enhancing competition for the colonization sites also used by
V. dahliae. Moreover, different biocontrol mechanisms (antagonism, competition, induction of host defense responses) [
52,
53,
99,
100] could be deployed by the olive indigenous BCAs in the consortia, improving biocontrol effectiveness.

 ,
, 




{kind=link}
{kind=link}
{kind=link}