Possible Criterion to Estimate the Juvenile Reference Length of Common Sardine (Strangomera bentincki) off Central-Southern Chile



Abstract
:1. Introduction
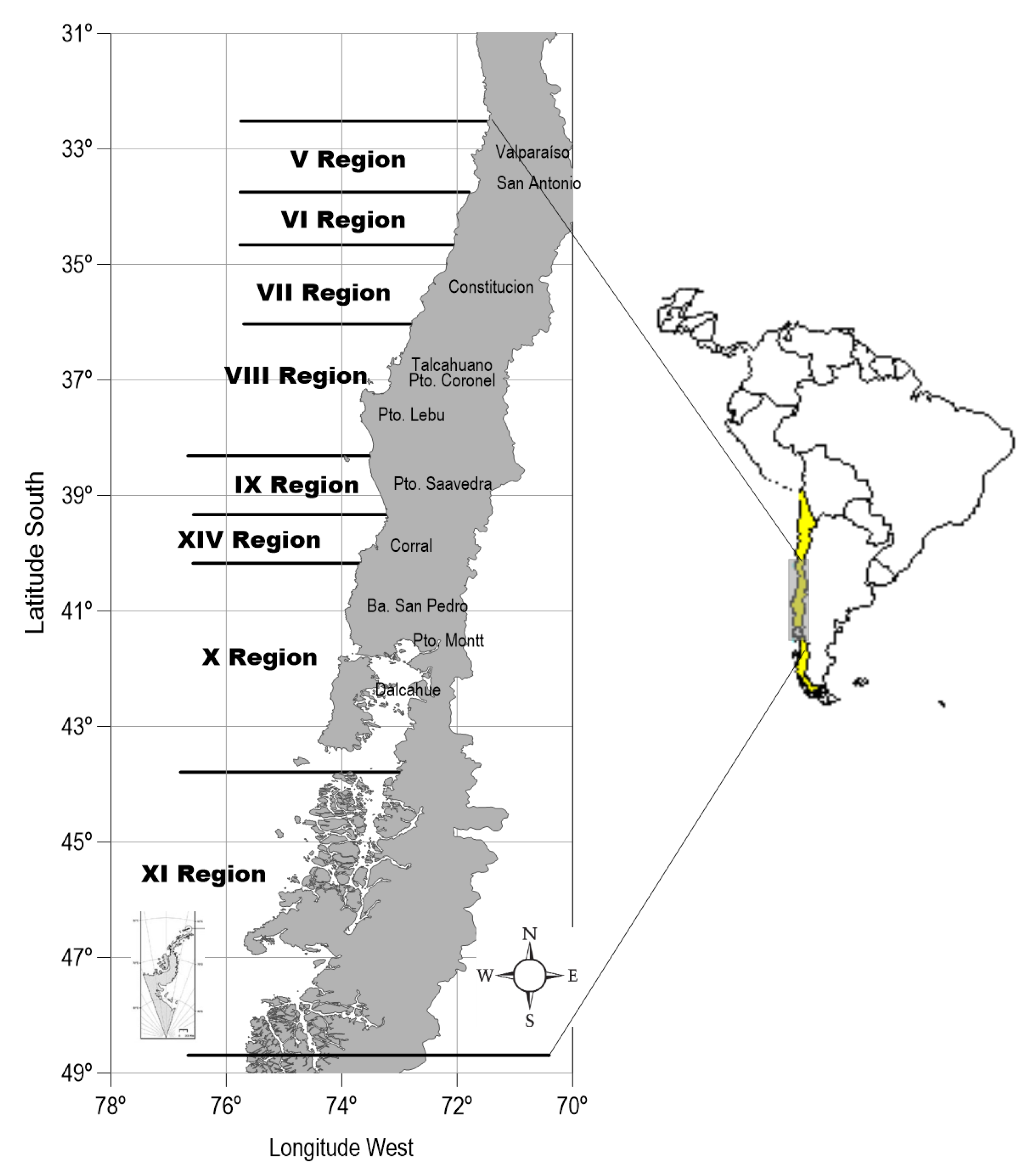
2. Materials and Methods
2.1. Data
2.2. Change Point Criterion
2.3. Model Selection Criterion
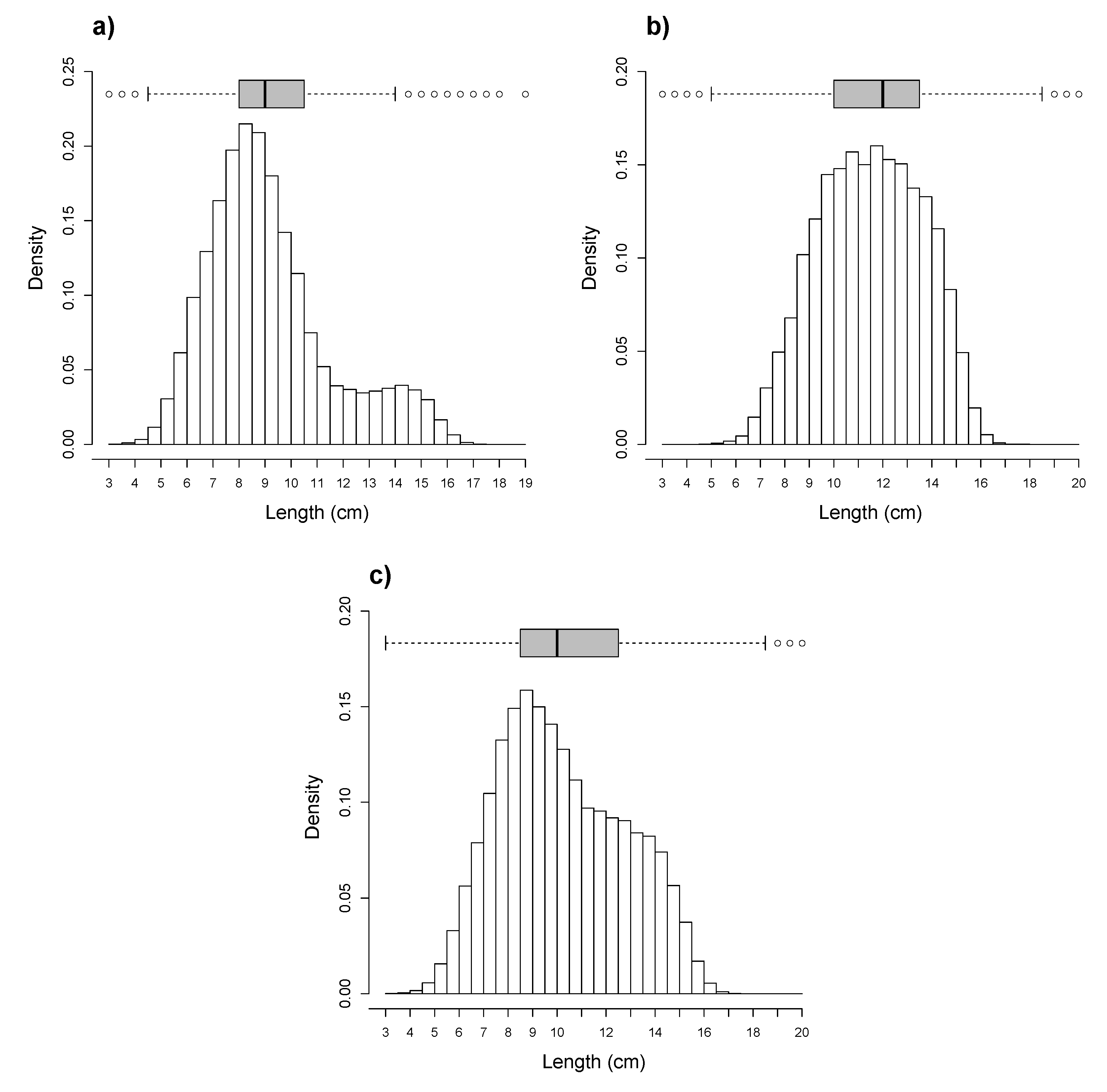
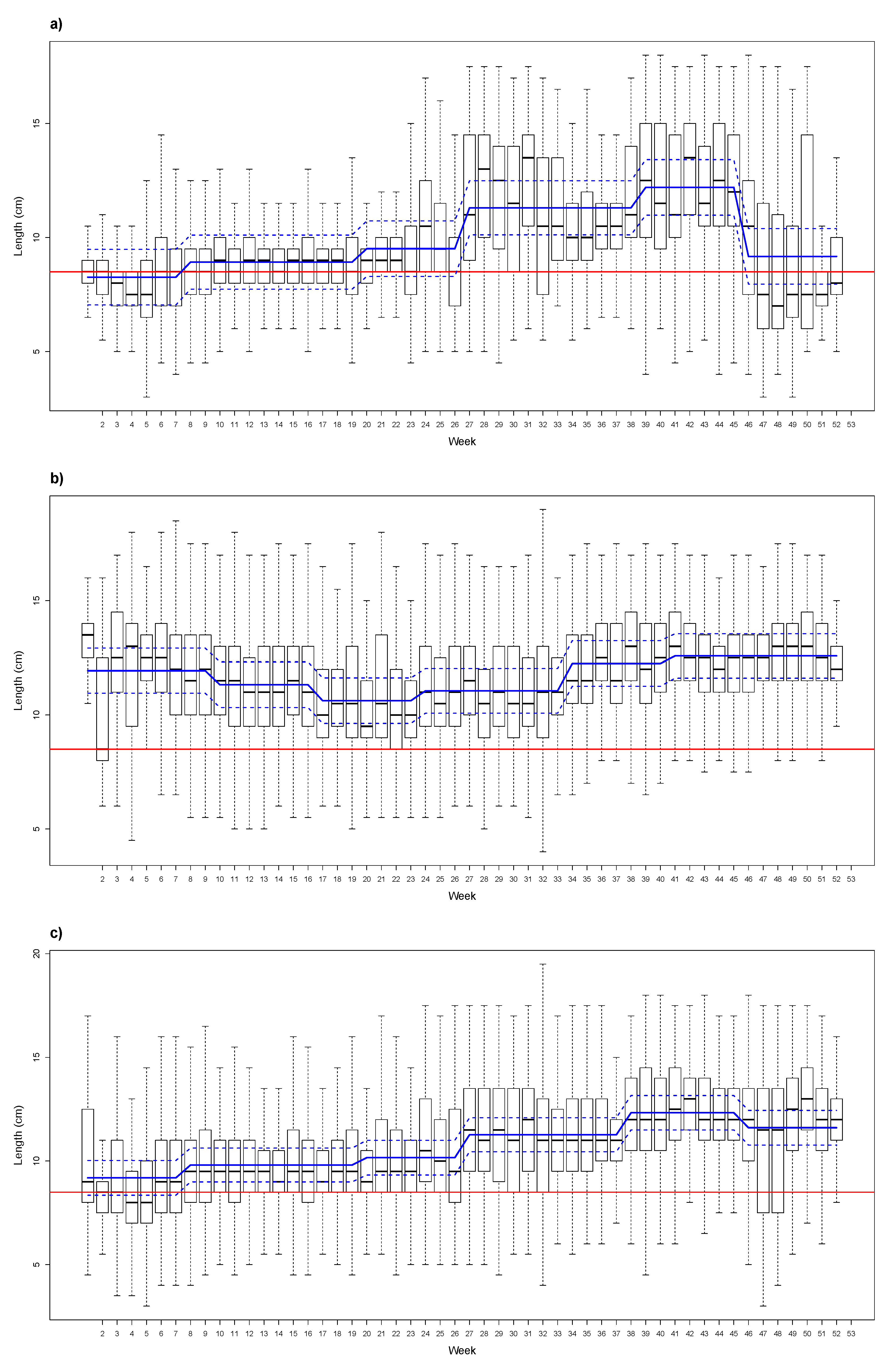
3. Results
4. Discussion and Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Cochrane, K.L. (Ed.) Guía del Administrador Pesquero. Medidas de Ordenación y su Aplicación; FAO Documento Técnico de Pesca 424; División de Recursos Pesqueros, Departamento de pesca, FAO: Rome, Italy, 2005. [Google Scholar]
- Cubillos, L.A.; Bucarey, D.A.; Canales, M. Monthly abundance estimation for common sardine Strangomera bentincki and anchovy Engraulis ringens in the central-southern area off Chile (34-40 S). Fish. Res. 2002, 57, 117–130. [Google Scholar] [CrossRef]
- Aranis, A.; Gómez, A.; Caballero, L.; Walker, K.; Ramírez, M.; Muñoz, G.; Eisele, G.; Cerna, F.; Valero, C.; López, A.; et al. Informe Final, Programa de Seguimiento de Pesquerías Pelágicas de la Zona Centro-Sur de Chile, 2016; Subsecretaría de Economía y EMT, Instituto de Fomento Pesquero: Valparaíso, Chile, 2015; 337p. [Google Scholar]
- Cubillos, L. Diseño y Evaluación Integral de Políticas de Escape de la Fracción Recluta, Pelágicos Pequeños, Zona Centro sur de Chile; Informe de Avance, Universidad de Concepción: Concepción, Chile, 2013; 131p. [Google Scholar]
- Csirke, J. Introducción a la Dinámica de Poblaciones de Peces; FAO Documento Técnico de Pesca; FAO: Rome, Italy, 1989; Volume 192, 82p. [Google Scholar]
- Mace, P.M. Relationships between common biological reference points used as thresholds and targets of fisheries management strategies. Can. J. Fish. Aquat. Sci. 1994, 51, 110–122. [Google Scholar] [CrossRef]
- Cubillos, L.A.; Arcos, D.F.; Bucarey, D.A.; Canales, M.T. Seasonal growth of small pelagic fish off Talcahuano, Chile (37∘ S, 73∘ W): A consequence of their reproductive strategy to seasonal upwelling? Aquat. Liv. Res. 2001, 14, 115–124. [Google Scholar] [CrossRef]
- Contreras-Reyes, J.E.; Arellano-Valle, R.B.; Canales, T.M. Comparing growth curves with asymmetric heavy-tailed errors: Application to southern blue whiting (Micromesistius australis). Fish. Res. 2014, 159, 88–94. [Google Scholar] [CrossRef]
- Comité Científico Pelágicos Pequeños. Análisis de Indicadores del Reclutamiento de Peces Pelágicos en la Zona Centro-Sur. In Proceedings of the GT Historia de Vida y Ecología Reproductiva, 20p, Viña del Mar, Chile, 4–5 October 2012. [Google Scholar]
- Andrews, D.W. Tests for parameter instability and structural change with unknown change point. Econometrica 1993, 61, 821–856. [Google Scholar] [CrossRef]
- Balke, N.S. Detecting level shifts in time series. J. Bus. Econ. Stat. 1993, 11, 81–92. [Google Scholar]
- Valderrama, L.; Contreras-Reyes, J.E.; Carrasco, R. Ecological impact of forest fires and subsequent restoration in Chile. Resources 2018, 7, 26. [Google Scholar] [CrossRef]
- Contreras-Reyes, J.E. Analyzing fish condition factor index through skew-gaussian information theory quantifiers. Fluct. Noise Lett. 2016, 15, 1650013. [Google Scholar] [CrossRef]
- Contreras-Reyes, J.E.; Canales, T.M.; Rojas, P.M. Influence of climate variability on anchovy reproductive timing off northern Chile. J. Mar. Syst. 2016, 164, 67–75. [Google Scholar] [CrossRef]
- Arellano-Valle, R.B.; Contreras-Reyes, J.E.; Stehlík, M. Generalized skew-normal negentropy and its application to fish condition factor time series. Entropy 2017, 19, 528. [Google Scholar] [CrossRef]
- Perron, P.-M.; Bai, J. Computation and Analysis of Multiple Structural Change Models. J. Appl. Economet. 2003, 18, 1–22. [Google Scholar]
- Zeileis, A.; Kleiber, C.; Krämer, W.; Hornik, K. Testing and dating of structural changes in practice. Comput. Stat. Data Anal. 2003, 44, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Zeileis, A.; Leisch, F.; Hornik, K.; Kleiber, C. strucchange: An R Package for Testing for Structural Change in Linear Regression Models. J. Stat. Softw. 2002, 7, 1–38. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2017; ISBN 3-900051-07-0. [Google Scholar]
- Saavedra, A.; Catasti, V.; Leiva, F.; Vargas, R.; Cifuentes, U.; Reyes, H.; Molina, E.; Cerna, F. Evaluación Hidroacústica de los Stocks de Anchoveta y Sardina Común entre la V y X Regiones, aÑo 2014; Informe Final FIP Nro 2013-05; Subsecretaría de Pesca, Gobierno de Chile: Valparaíso, Chile, 2014; 306p. [Google Scholar]
- Cubillos, L.; Arancibia, H. On the seasonal growth of common sardine (Strangomera bentincki) and anchovy (Engraulis ringens) off Talcahuano, Chile. Rev. Biol. Mar. 1993, 28, 39–43. [Google Scholar]
- Sepúlveda, A. Variabilidad Temporal del Ictioplancton en El área de Surgencia Costera de Chile Central: Procesos Ambientales y Biológicos Asociados. M.Sc. Thesis, Universidad de Concepción, Concepción, Chile, 1990; 81p. [Google Scholar]
- Galleguillos, R.; Troncoso, L.; Oyarzún, C. Population differentiation in the Chilean herring, Strangomera bentincki (Pisces: Clupeidae) I: Genetic analysis of protein variability. Rev. Chil. Hist. Nat. 1997, 70, 351–361. [Google Scholar]
- Nicholson, M.D.; Jennings, S. Testing candidate indicators to support ecosystem-based management: The power of monitoring surveys to detect temporal trends in fish community metrics. ICES J. Mar. Sci. 2004, 61, 35–42. [Google Scholar] [CrossRef]
- Barría, P.; Böhm, M.G.; Aranis, A.; Gili, R.; Donoso, M.; Rosales, S. Evaluación Indirecta y Análisis de la Variabilidad del Crecimiento de Sardina Común y Anchoveta en la Zona Centro-Sur; Informe Final. FIP Nro. 97-10; Subsecretaría de Pesca, Gobierno de Chile: Valparaíso, Chile, 1999; 115p. [Google Scholar]
- Gulland, J. Informe sobre la dinámica de la población de la anchoveta peruana. Boletín IMARPE 1968, 1, 305–346. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Area | Intercept | 95% Confidence Bands | k | AIC | ||
|---|---|---|---|---|---|---|
| V–VIII | 8.773 | 8.336–9.210 | −74.986 | 2 | 155.972 | 0.506 |
| 8.903 | 8.629–9.177 | −53.385 | 3 | 114.770 | 0.785 | |
| 8.318 | 7.862–8.774 | −48.646 | 4 | 107.292 | 0.821 | |
| 8.318 | 7.900–8.736 | −43.527 | 5 | 99.054 | 0.853 | |
| 8.263 | 7.832–8.693 | −40.994 | 6 | 95.989 | 0.866 | |
| 8.263 | 7.775–8.750 | −46.808 | 7 | 109.617 | 0.833 | |
| IX–XIV | 11.261 | 11.053–11.470 | −45.855 | 2 | 97.709 | 0.494 |
| 11.854 | 11.554–12.153 | −35.591 | 3 | 79.183 | 0.659 | |
| 11.938 | 11.615–12.260 | −33.694 | 4 | 77.388 | 0.683 | |
| 11.938 | 11.623–12.252 | −31.837 | 5 | 75.674 | 0.705 | |
| 11.938 | 11.627–12.249 | −30.616 | 6 | 75.231 | 0.718 | |
| 11.938 | 11.598–12.278 | −34.645 | 7 | 85.289 | 0.671 | |
| V–XIV | 9.737 | 9.516–9.958 | −42.629 | 2 | 91.257 | 0.759 |
| 9.640 | 9.433–9.846 | −35.490 | 3 | 78.979 | 0.817 | |
| 9.188 | 8.851–9.526 | −29.515 | 4 | 69.030 | 0.854 | |
| 9.188 | 8.884–9.493 | −23.506 | 5 | 59.012 | 0.884 | |
| 9.188 | 8.892–9.485 | −21.526 | 6 | 57.052 | 0.893 | |
| 9.188 | 8.878–9.499 | −23.337 | 7 | 62.674 | 0.885 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker, K.; Aranis, A.; Contreras-Reyes, J.E. Possible Criterion to Estimate the Juvenile Reference Length of Common Sardine (Strangomera bentincki) off Central-Southern Chile. J. Mar. Sci. Eng. 2018, 6, 82. https://doi.org/10.3390/jmse6030082
Walker K, Aranis A, Contreras-Reyes JE. Possible Criterion to Estimate the Juvenile Reference Length of Common Sardine (Strangomera bentincki) off Central-Southern Chile. Journal of Marine Science and Engineering. 2018; 6(3):82. https://doi.org/10.3390/jmse6030082
Chicago/Turabian StyleWalker, Karen, Antonio Aranis, and Javier E. Contreras-Reyes. 2018. "Possible Criterion to Estimate the Juvenile Reference Length of Common Sardine (Strangomera bentincki) off Central-Southern Chile" Journal of Marine Science and Engineering 6, no. 3: 82. https://doi.org/10.3390/jmse6030082