Central [CNS] and Peripheral [Gastric Tissue] Selective Monitoring of Somatostatin (SRIF) with Micro-Sensor and Voltammetry in Rats: Influence of Growth Factors (GH, EGF)

Abstract
:1. Introduction
2. Methods & Results
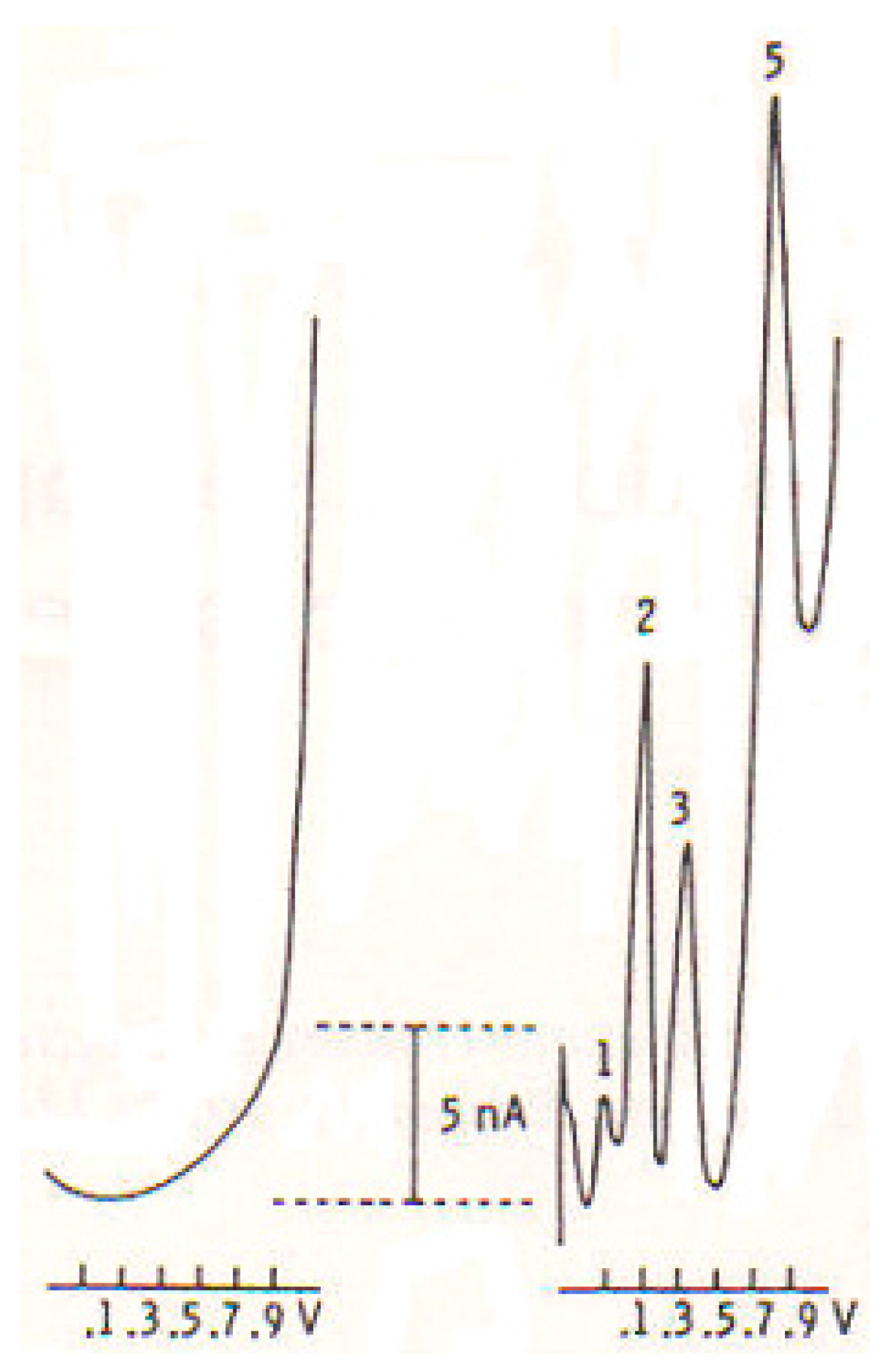
2.1. DPV and Micro-Sensors
- The undersized dimensions of the probe allow measurements with minimal damage to the nervous tissue and disturbance to the animal.
- The area sampled is approximately 10–6 mm3: this means high anatomical resolution of the location of measurement within discrete brain regions.
- Fast, continuous measurements in vivo, in situ in real time, without requiring perfusion or sample preparation or chromatographic separation or radiolabelled transmitter supplies.
- Feasibility of performing DPV in freely moving rodents; this solves the problems associated with anesthetics permitting correlations within neuronal activities.
- Wireless DPV measurements allow electrochemical studies in completely free-moving situations [21].
2.2. In Vitro Studies
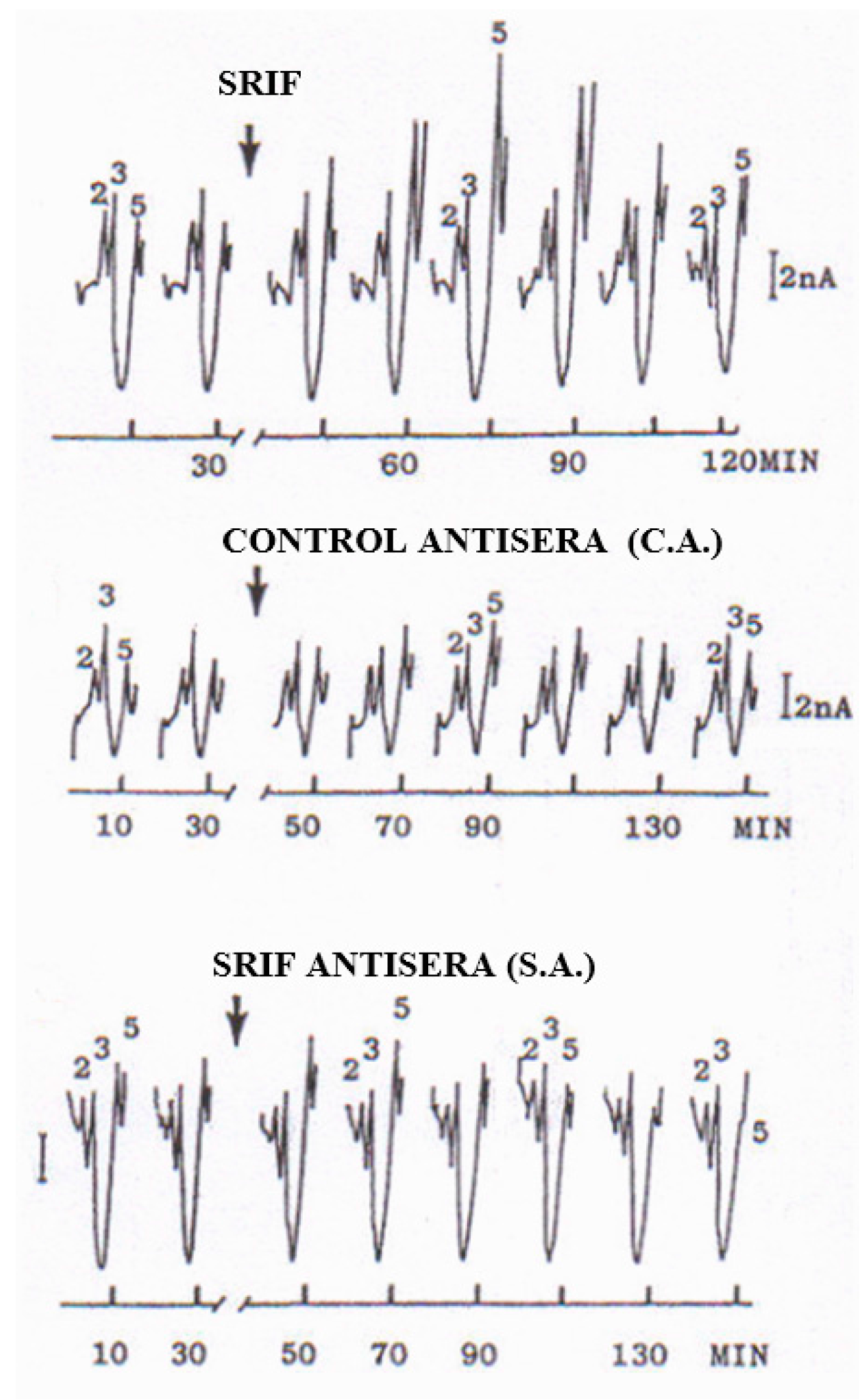
2.3. In Vivo CNS Studies
- -
- Bacitracin, which strongly inhibits peptidase activity, as described in [23], resulted in a large increase of Peak 5, therefore supporting it as a peptidergic signal.
- -
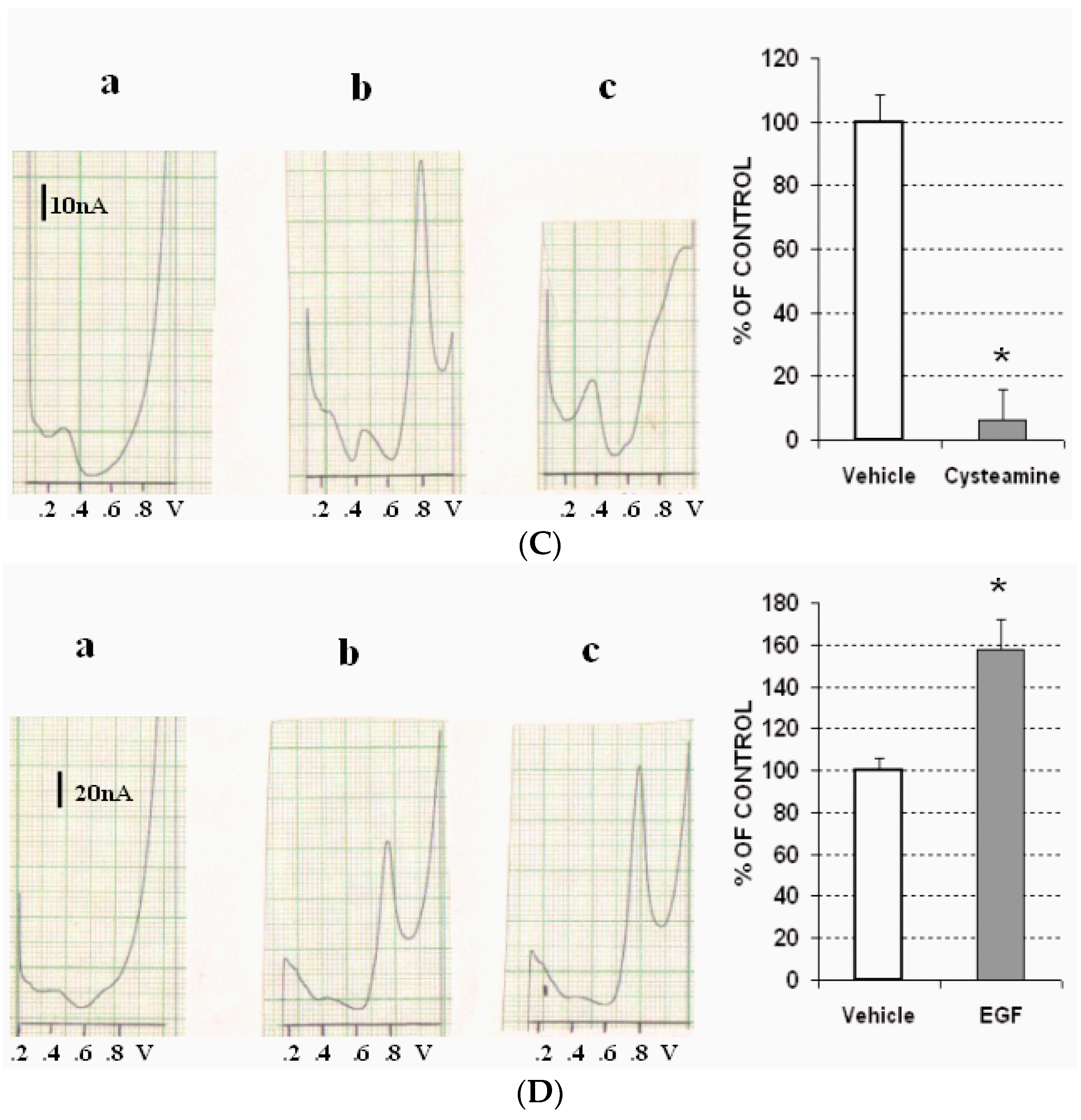
- Cysteamine, which is a selective depletory of cerebral SRIF [24], was followed by a rapid decrease until disappearance of Peak 5.
- -
- SRIF antisera, i.e., antibodies for SRIF (rabbit polyclonal, IgG antiSRIF AB5494); Millipore (MERCK), but not control antisera, i.e., non-specific antibodies, as described by Funato, et al. [25], resulted in a rapid decrease until disappearance of striatal Peak 5.
- -
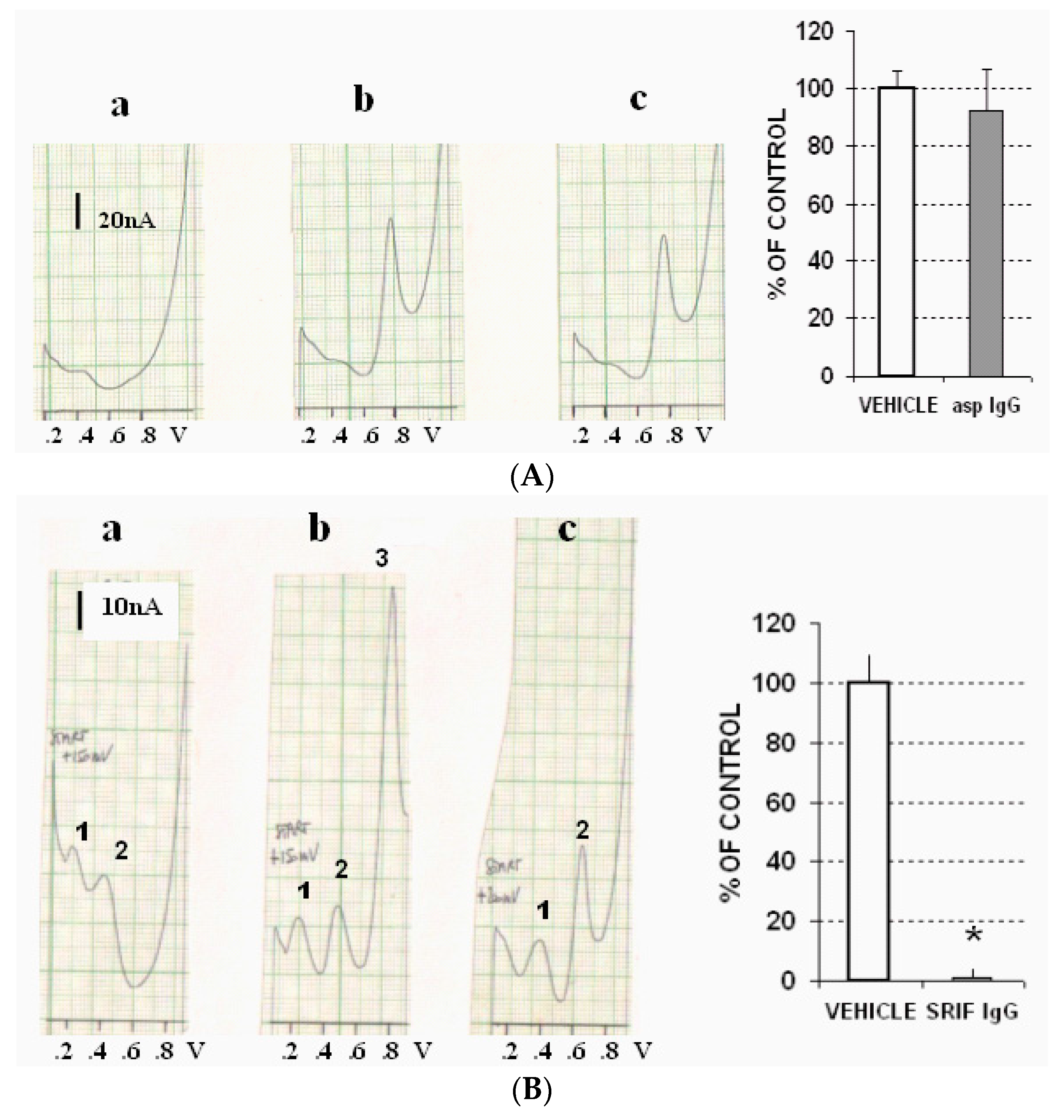
2.4. Ex Vivo Gastric Tissue Studies
- (1)
- Vehicle (PBS), or
- (2)
- Antibodies for SRIF (rabbit polyclonal, IgG antiSRIF AB5494); Millipore (MERCK S.p.A., Vimodrone, Milan, Italy) or with non-specific antibodies as described by Funato, et al. [25], or
- (3)
- Cysteamine 1 mM, or
- (4)
- Epidermal growth factor (EGF) 1 mM.
3. Statistical Analysis
4. Discussion and Conclusions
Conflicts of Interest
References
- Epelbaum, J. Somatostatin in the central nervous system: Physiology and pathological modifications. Prog. Neurobiol. 1986, 27, 63–100. [Google Scholar] [CrossRef]
- Somatostatin. Encyclopædia Britannica. Encyclopædia Britannica Inc., 2016. Available online: http://www.britannica.com/science/somatostatin (accessed on 4 May 2016).
- Reichlin, S.N. Somatostatin (two parts). N. Engl. Med. J. 1983, 309, 1494–1501 and 1556–1563. [Google Scholar] [CrossRef] [PubMed]
- Weckbecker, G.; Lewis, I.; Albert, R.; Schmid, H.A.; Hoyer, D.; Bruns, C. Opportunities in somatostatin research: Biological, chemical and therapeutic aspects. Nat. Rev. Drug Discov. 2003, 2, 999–1017. [Google Scholar] [CrossRef] [PubMed]
- Schally, A. Oncological Applications of Somatostatin Analogues. Cancer Res. 1988, 48, 6977–6985. [Google Scholar] [PubMed]
- Massa, A.; Barbieri, F.; Aiello, C.; Arena, S.; Pattarozzi, A.; Pirani, P.; Corsaro, A.; Iuliano, R.; Fusco, A.; Zona, G.; et al. The expression of the phosphotyrosine phosphatase DEP-1/PTPeta dictates the responsivity of glioma cells to somatostatin inhibition of cell proliferation. J. Biol. Chem. 2004, 279, 29004–29012. [Google Scholar] [CrossRef] [PubMed]
- Dockray, G.J. Topical Review. J. Physiol. 1999, 518, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Lehy, T.; Dubrasquet, M.; Bonfils, S. Effect of Somatostatin on Normal and Gastric-Stimulated Cell Proliferation in the Gastric and Intestinal Mucosae of the Rat. Digestion 1979, 19, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Rao, J.N.; Wang, J.Y. Morgan & Claypool Life Sciences Book; University of Maryland: San Rafael, CA, USA, 2010. [Google Scholar]
- Takagi, K.; Okabe, S. The effects of drugs on the production and recovery processes of the stress ulcer. Jpn. J. Pharmacol. 1968, 18, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Li, R.; Xu, S.; Liu, Z.; Ma, X. Hypothalamic Astrocytes Respond to Gastric Mucosal Damage Induced by Restraint Water-Immersion Stress in Rat. Front. Behav. Neurosci. 2016, 10, 210. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Swierczek, J.S.; Ogden, W.D.; Thompson, J.C. Cytoprotective effect of pentagastrin and epidermal growth factor on stress ulcer formation. Possible role of somatostatin. Ann. Surg. 1985, 201, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, I.; Yusof, K.; Ismail, N.; Fahami, N. Protective effect of palm vitamin E and α-tocopherol against gastric lesions induced by water immersion restraint stress in Sprague-Dawley rats. Indian J. Pharmacol. 2008, 40, 73–77. [Google Scholar] [PubMed]
- Szabo, S.; Reichlin, S. Somatostatin in rat tissues is depleted by cysteamine administration1. Endocrinology 1981, 109, 2255–2257. [Google Scholar] [CrossRef] [PubMed]
- Brendan, J.; Grabau, B.J.; Zavros, Y.; Hardt, K.J.; Shulkes, A. Developmental Regulation of Gastric Somatostatin Secretion in the Sheep. Endocrinology 1999, 140, 603–608. [Google Scholar]
- Pallis, E.; Vasilaki, A.; Fehlmann, D.; Kastellakis, A.; Hoyer, D.; Spyraki, C.; Thermos, K. Antidepressants Influence Somatostatin Levels and Receptor Pharmacology in Brain. Neuropsychopharmacology 2009, 34, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Crespi, F.; Sharp, T.; Maidment, N.; Marsden, C.A. Differential pulse voltammetry: Simultaneous in vivo measurement of ascorbic acid, catechols and 5-hydroxyindoles in the rat striatum. Brain Res. 1984, 322, 135–138. [Google Scholar] [CrossRef]
- Crespi, F. In vivo voltammetric detection of neuropeptides with micro carbon fiber biosensors: Possible selective detection of somatostatin. Anal. Biochem. 1991, 194, 1–8. [Google Scholar] [CrossRef]
- Crespi, F.J. In vivo voltammetry with micro-biosensors for analysis of neurotransmitter release and metabolism. Neurosci. Methods 1990, 34, 53–65. [Google Scholar] [CrossRef]
- Crespi, F. Serotonin, how to find it. J. Malta Chamb. Sci. 2013, 14–22. [Google Scholar] [CrossRef]
- Crespi, F. Wireless in vivo voltammetric measurements of neurotransmitters in freely behaving rats. Biosens. Bioelectron. 2010, 25, 2425–2430. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.W.; Brazell, M.P.; Marsden, C.A. Electrochemistry of neuropeptides: A possible method for assay and in vivo detection. Life Sci. 1981, 29, 1001–1007. [Google Scholar] [CrossRef]
- Hokfelt, T.; Lundberg, J.M.; Schultzberg, M.; Johansson, O.; Skirboll, L.; Anggard, A.; Fredholm, B.; Hamberger, B.; Pernow, B.; Rehfeld, J.; et al. Cellular Localization of Peptides in Neural Structures [and Discussion]. Proc. R. Soc. Lond. B 1980, 210, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Chattha, G.K.; Beal, M.F. Effect of cysteamine on somatostatin and neuropeptide Y in rat striatum and cortical synaptosomes. Brain Res. 1987, 401, 359–364. [Google Scholar] [CrossRef]
- Funato, H.; Sato, M.; Sinton, C.M.; Gautronc, L.; Williamsa, S.C.; Skacha, A.; Elmquistc, J.K.; Skoultchid, A.I.; Yanagisawa, M. Loss of Goosecoid-like and DiGeorge syndrome critical region 14 in interpeduncular nucleus results in altered regulation of rapid eye movement sleep. Proc. Natl. Acad. Sci. USA 2010, 107, 18155–18160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chihara, K.; Minamitani, N.; Kaji, H.; Arimura, A.; Fujita, T. Intraventricularly injected growth hormone stimulates somatostatin release into RAT hypophysial portal blood. Endocrinology 1981, 109, 2279–2281. [Google Scholar] [CrossRef] [PubMed]
- Stachura, E.; Tyler, J.; Farmer, P. Combined Effects of Human Growth Hormone (GH)-Releasing Factor-44 (GRF) and Somatostatin (SRIF) on Post-SRIF Rebound Release of GH and Prolactin: A Model for GRF-SRIF Modulation of Secretion. Endocrinology 1988, 123, 1476–1482. [Google Scholar] [CrossRef] [PubMed]
- Tannenbaum, G.S.; Epelbaum, J. Somatostatin. Compr. Physiol. 2011, 221–265. [Google Scholar] [CrossRef]
- Finley, J.C.W.; Madernt, J.L.; Rogers, L.J.; Petrusz, P. The immunocytochemical localization of somatostatin-containing neurons in the rat central nervous system. Neuroscience 1981, 6, 2173–2192. [Google Scholar] [CrossRef]
- Beal, M.F.; Domesick, V.B.; Martin, J.B. Regional somatostatin distribution in the rat striatum. Brain Res. 1983, 278, 103–108. [Google Scholar] [CrossRef]
- Allen, J.P.; Hathway, G.J.; Clarke, N.J.; Jowett, M.I.; Topps, S.; Kendrick, K.M.; Humphrey, P.; Wilkinson, L.S.; Emson, P.C. Somatostatin receptor 2 knockout/lacZ knockin mice show impaired motor coordination and reveal sites of somatostatin action within the striatum. Eur. J. Neurosci. 2003, 17, 1881–1895. [Google Scholar] [CrossRef] [PubMed]
- Palkovits, M.; Brownstein, M.J.; Eiden, L.E.; Beinfeld, M.C.; Russell, J.; Arimura, A.; Szabo, S. Selective depletion of somatostatin in rat brain by cysteamine. Brain Res. 1982, 240, 178–180. [Google Scholar] [CrossRef]
- Bakhit, C.; Benoit, R.; Bloom, F. Effects of cysteamine on pro-somatostatin related peptides. Regul. Pept. 1983, 6, 169–177. [Google Scholar] [CrossRef]
- Sagar, S.M.; Landry, D.; Millard, W.J.; Badger, T.M.; Arnold, M.A.; Martin, J.B. Depletion of somatostatin-like immunoreactivity in the rat central nervous system by cysteamine. J. Neurosci. 1982, 2, 225–231. [Google Scholar] [PubMed]
- Beal, M.; Martin, J. Depletion of striatal somatostatin by local cysteamine injection. Brain Res. 1984, 308, 319–324. [Google Scholar] [CrossRef]
- Kwok, R.P.; Cameron, J.L.; Faller, D.V.; Fernstrom, J.D. Effects of cysteamine administration on somatostatin biosynthesis and levels in rat hypothalamus. Endocrinology 1992, 131, 2999–3009. [Google Scholar] [CrossRef] [PubMed]
- Kraicer, J.; Sheppard, M.S.; Luke, J.; Lussier, B.; Moor, B.C.; Cowan, J.S. Effect of Withdrawal of Somatostatin and Growth Hormone (GH)-Releasing Factor on GH Release in Vitro. Endocrinology 1988, 122, 1810–1815. [Google Scholar] [CrossRef] [PubMed]
- Sam, S.; Frohman, L.A. Normal Physiology of Hypothalamic Pituitary Regulation. Endocrinol. Metab. Clin. N. Am. 2008, 37, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Mogi, K.; Yonezawa, T.; Chen, D.S.; Li, J.Y.; Suzuki, M.; Yamanouchi, K.; Sawasaki, T.; Nishihara, M. Relationship between Growth Hormone (GH) Pulses in the Peripheral Circulation and GH-Releasing Hormone and Somatostatin Profiles in the Cerebrospinal Fluid of Goats. J. Vet. Med. Sci. 2004, 66, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.D.; Kim, K.; Ren, S.G.; Chelly, M.; Umehara, Y.; Melmed, S. Central and peripheral actions of somatostatin on the growth hormone-IGF-I axis. J. Clin. Investig. 2004, 114, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Berelowitz, M.; Frohman, L.A. Effect of catecholamines on somatostatin secretion by isolated perfused rat stomach. Am. J. Physiol. 1981, 240, E274–E278. [Google Scholar] [PubMed]
- Kitajima, N.; Chihara, K.; Abe, H.; Okimura, Y.; Fujii, Y.; Sato, M.; Shakutsui, S.; Watanabe, M.; Fujita, T. Effects of Dopamine on Immunoreactive Growth Hormone-Releasing Factor and Somatostatin Secretion from Rat Hypothalamic Slices Perifused in Vitro. Endocrinology 1989, 124, 69–76. [Google Scholar] [CrossRef] [PubMed]
- West, C.R.; Lookingland, H.J.; Tucker, H.A. Regulation of growth hormone-releasing hormone and somatostatin from perifused, bovine hypothalamic slices. II. Dopamine receptor regulation. Domest. Anim. Endocrinol. 1997, 14, 349–357. [Google Scholar] [CrossRef]
- Crespi, F. Functional in vivo interaction between growth hormone and dopamine systems are correlated to changes in striatal somatostatin levels as detected by voltammetry. Exp. Brain Res. 1993, 94, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Drago, F.; Montoneri, C. Influence of growth hormone on cysteamine-induced gastro-duodenal lesions in rats: The involvement of somatostatin. Life Sci. 1997, 61, 21–28. [Google Scholar] [CrossRef]
- Abe, H.; Molitch, M.E.; Van Wyk, J.J.; Underwood, L.E. Human Growth Hormone and Somatomedin C Suppress the Spontaneous Release of Growth Hormone in Unanesthetized Rats. Endocrinology 1983, 113, 1319–1324. [Google Scholar] [CrossRef] [PubMed]
- Tsigos, C.; Chrousos, G. Hypothalamic–pituitary–adrenal axis, neuroendocrine factors and stress. J. Psychosom. Res. 2002, 53, 865–871. [Google Scholar] [CrossRef]
- Sanghavi, B.J.; Wolfbeis, O.S.; Hirsch, T.; Swami, N.S. Nanomaterial-based electrochemical sensing of neurological drugs and neurotransmitters. Mikrochim. Acta 2015, 182, 1–41. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance Potential (mV) | |
|---|---|
| Tyrosin | 720 |
| Tryptophan | 860 |
| Cysteine | 870 |
| Neurotensin | 670 |
| Oxytocin | 585 |
| Vasopressin | 610 |
| Caerulein | 670 |
| Leu-enkephalin | 605 |
| Met-enkephalin | 570 |
| ACTH 1-24 | 650 |
| ACTH 17-39 | 700 |
| ß-endorphin | 800 |
| Somatostatin | 805 |
| Cholecystokinin (CCK-4) | 730 |
| Cholecystokinin (CCK-8) | 810 |
| LH-RH | 700 |
| Alpha-MSH | 795 |
| LOCAL | TIME (min) | |||
| TREATMENT | 0 | 40 | 80 | 120 |
| NaCl 0.9% | 100 | 102 | 95 | 93 |
| 2 µL, n = 9 | ±5 | ±8 | ±11 | ±16 |
| SRIF | 100 | 475 * | 155 * | 89 |
| 2 µg, n = 7 | ±9 | ±98 | ±41 | ±22 |
| (C.A.) | 100 | 122 | 136 | 114 |
| 2 µL, n = 7 | ±8 | ±18 | ±23 | ±11 |
| (S.A.) | 100 | 125 | 65 * | 13 * |
| 2 µL, n = 7 | ±5 | ±13 | ±11 | ±6 |
| Bacitracin | 100 | 130 | 159 * | 153 * |
| 10 ng, n = 5 | ±4 | ±13 | ±16 | ±14 |
| GH | 100 | 141 * | 114 | 106 |
| 2 µg, n = 5 | ±6 | ±8 | ±7 | ±12 |
| SYSTEMIC | TIME (min) | |||
| TREATMENT | 0 | 10 | 20 | 30 |
| Cysteamine | 100 | 65 * | 22 * | 8 * |
| 100 mg/kg, n = 7 | ±10 | ±13 | ±11 | |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crespi, F. Central [CNS] and Peripheral [Gastric Tissue] Selective Monitoring of Somatostatin (SRIF) with Micro-Sensor and Voltammetry in Rats: Influence of Growth Factors (GH, EGF). Biosensors 2017, 7, 53. https://doi.org/10.3390/bios7040053
Crespi F. Central [CNS] and Peripheral [Gastric Tissue] Selective Monitoring of Somatostatin (SRIF) with Micro-Sensor and Voltammetry in Rats: Influence of Growth Factors (GH, EGF). Biosensors. 2017; 7(4):53. https://doi.org/10.3390/bios7040053
Chicago/Turabian StyleCrespi, Francesco. 2017. "Central [CNS] and Peripheral [Gastric Tissue] Selective Monitoring of Somatostatin (SRIF) with Micro-Sensor and Voltammetry in Rats: Influence of Growth Factors (GH, EGF)" Biosensors 7, no. 4: 53. https://doi.org/10.3390/bios7040053