Amperometric Self-Referencing Ceramic Based Microelectrode Arrays for D-Serine Detection


1
Institute of Experimental and Clinical Pharmacology and Toxicology, University of Lübeck, Ratzeburger Allee 160, 23562 Lübeck, Germany
2
The Department of Pharmaceutical Biosciences, Uppsala University, 75124 Uppsala, Sweden
3
The Department of Physiology and Pharmacology, Karolinska Institutet, 17177 Stockholm, Sweden
4
CenMeT, University of Kentucky, Lexington, 40506 KY, USA
5
Quanteon LLC, Nicholasville, 40356 KY, USA
*
Author to whom correspondence should be addressed.
Biosensors 2018, 8(1), 20; https://doi.org/10.3390/bios8010020
Submission received: 1 February 2018
/
Revised: 15 February 2018
/
Accepted: 2 March 2018
/
Published: 6 March 2018
(This article belongs to the Special Issue Implantable Biosensors for in vivo Detection and Measurement)
Abstract
:D-serine is the major D-amino acid in the mammalian central nervous system. As the dominant co-agonist of the endogenous synaptic NMDA receptor, D-serine plays a role in synaptic plasticity, learning, and memory. Alterations in D-serine are linked to neuropsychiatric disorders including schizophrenia. Thus, it is of increasing interest to monitor the concentration of D-serine in vivo as a relevant player in dynamic neuron-glia network activity. Here we present a procedure for amperometric detection of D-serine with self-referencing ceramic-based microelectrode arrays (MEAs) coated with D-amino acid oxidase from the yeast Rhodotorula gracilis (RgDAAO). We demonstrate in vitro D-serine recordings with a mean sensitivity of 8.61 ± 0.83 pA/µM to D-serine, a limit of detection (LOD) of 0.17 ± 0.01 µM, and a selectivity ratio of 80:1 or greater for D-serine over ascorbic acid (mean ± SEM; n = 12) that can be used for freely moving studies.
1. Introduction
In the 1990’s, the occurrence of D-serine in the mammalian brain and the similarity in its distribution pattern to that of the N-methyl D-aspartate receptor (NMDAR) were first reported [1,2]. D-serine has since been confirmed as the main endogenous co-agonist of the glycine modulatory site of the NMDAR in corticolimbic areas of the brain [3,4,5]. As such, it is presumed to be crucially involved in neuroplasticity and cognitive functions, in particular, for learning and memory [5,6,7,8,9,10,11,12,13,14,15,16,17,18]. Furthermore, D-serine has been attributed a role in the hyper- and/or hypofunction of the NMDAR in neuropsychiatric disorders [19,20,21]. For example, a major component in the pathophysiology of schizophrenia, in particular, the cognitive deficits, is suggested to be NMDAR hypofunction associated with reduced D-serine levels [12,20]. In the brain, D-serine is synthesized in neurons and/or astrocytes from L-serine by the enzyme serine racemase [22]. Endogenous degradation after cellular uptake can occur via the enzyme D-amino-acid oxidase (DAAO), located in peroxisomes, although for the brain other mechanisms are also discussed [23].
Thus, independent of the ongoing debate on the neuronal and/or glial sources of D-serine [22,24,25,26,27], its level and dynamics are pressing issues for understanding cognitive impairment and neuropsychiatric diseases, as well as for basic research in neuroplasticity.
While several types of biosensors have already been employed for amperometric D-serine detection [18,28,29,30,31,32,33,34], the extracellular levels of D-serine in the brain of freely moving rodents have so far only been measured by microdialysis [32,35,36,37,38]. A main advantage of amperometric recordings is the ability to detect neurochemical changes in the range of seconds [39,40,41]. Such high temporal resolution is relevant not only to detect putative fast transients as found for glutamate [41], but also for monitoring neurochemical changes in relation to both state-dependent [42], as well as fast changes in brain electric activity. Here, we provide a protocol for amperometric D-serine measurement with D-amino acid oxidase (DAAO) coated ceramic-based microelectrode arrays (MEAs), whose configuration allows for offline self-referencing. The principle of self-referencing employs 2 types of channels, here with each consisting of a pair of platinum recording sites: D-serine detecting channels and sentinel channels. Both of the channel types are able to measure background noise and interferent activity, but only the enzyme coated sites (D-serine detecting channels) are able to detect D-serine [43,44,45]. Background noise and neurochemical interferent activity can thus be easily subtracted from the D-serine detecting channels [43,44,45]. Sentinel and D-serine detecting channels are spaced only tens of micrometers apart on the MEA [44]. In fact, dual-sided MEAs with eight recording sites have been used to record simultaneously in multiple regions of the prefrontal cortex [46]. With specific configurations, recordings of different analytes with one MEA [47,48] and of local field potentials in parallel using a high data acquisition rate were conducted [48,49,50]. MEAs possess a high spatial resolution, and are made of a biocompatible material having been shown to produce only minimal tissue damage [44,51], a feature that is required for chronic recordings.
2. Materials and Methods
2.1. Chemicals
L-ascorbic acid (AA), D-serine, D-alanine, L-glutamate, dopamine hydrochloride (DA), bovine serum albumin (BSA), sodium chloride (NaCl), sodium phosphate monobasic monohydrate (NaH2PO4), sodium phosphate dibasic anhydrous (Na2HPO4), meta-phenylenediamine dihydrochloride (mPD), and glutaraldehyde were purchased from Sigma-Aldrich (St. Louis, MO, USA). Hydrogen peroxide (H2O2) 3% (Paul W. Beyvers GmbH, Berlin, Germany) was purchased at a local pharmacy. The recombinant D-amino acid oxidase from the yeast Rhodotorula gracilis (RgDAAO, EC 1.4.3.3) was purified, as stated in Fantinato et al., [52], and was purchased with an approximate activity of D-serine given as 20 U/mg dry protein (and 100 U/mg dry protein on D-alanine) from “The Protein Factory research center” (Milano, Italy). Porcine kidney D-amino acid oxidase (pkDAAO, EC 1.4.3.3) with a specific activity of D-alanine given as 2.53 U/mg dry weight protein (data on D-serine was not provided) was obtained from Worthington Biochemical Corporation (Lakewood, NJ, USA). Stock solutions were prepared using deionized distilled water, except in the case of dopamine that was mixed with 1% perchloric acid (PCA) in addition, for a long shelf life. The 0.05 M phosphate-buffered saline (PBS) solution used for in vitro calibration had a pH close to 7.4, mimicking the physiological levels. A 5 mM mPD solution was made using nitrogen saturated PBS to reduce the oxidation of the compound.
2.2. MEA Preparation
Ceramic-based S2 MEAs were obtained from CenMeT (Lexington, KT, USA; a comprehensive fabrication review can be found elsewhere [44]). The S2 style MEA is arranged as two pairs of platinum recording sites of 333 × 15 µm, separated by 30 µm from the neighboring recording site and by 100 µm from the other pair (cp. Figure 1) [44]. After arrival, MEAs were cleaned and calibrated in vitro using H2O2 to test performance (data not shown). Subsequently, MEAs were enzyme coated (see “Enzyme preparation and immobilization” section) and stored at −20 °C until use, to maintain enzymatic stability. An exclusion layer was applied (see “Electropolymerization of meta-phenylenediamine (mPD)”) one day prior to calibration (see “In vitro calibration and recording parameters”). In-depth background on the relevance of these steps has been reported elsewhere in regard to other enzyme-based MEAs [43,44,53].
For the freely moving recording the printed circuit board of the MEA was soldered onto a miniature Omnetics connector (Omnetics Connector Corporation, Minneapolis, MN, USA) and a chloride coated silver (Ag/AgCl) wire was used as reference electrode (Teflon-coated silver wire, 125 µm bare diameter, A-M Systems, Inc., Sequim, WA, USA). This MEA configuration enables the use of a smaller preamplifier and thus, reduces weight of the headstage on the rat.
2.3. Enzyme Preparation and Immobilization
The MEA configuration allows for offline self-referencing: two enzyme-coated platinum sites were used for detecting D-serine (in Figure 1B), while two sentinel channels (in Figure 1A) detected other potential electroactive compounds that could likewise be oxidized at +0.7 V, e.g., endogenous H2O2. Measurements recorded with the sentinel channels are subtracted offline from the D-serine detecting channels to obtain an interference-free D-serine signal. For coating, one droplet from an aliquot of each 1% BSA, 0.125% glutaraldehyde, and 0.1 U/µL RgDAAO solution was applied to the two D-serine recording channels under a stereomicroscope (0.8×–4×; Olympus Optical Company, Hamburg, Germany) using a Hamilton microsyringe (Hamilton Company, Reno, NV, USA). Similarly, a chemically inactive protein matrix composed of only BSA and glutaraldehyde was cross-linked onto the sentinel channels. Following the same procedures as described above, some MEAs (n = 4) were coated with mammalian DAAO purified from porcine kidney.
2.4. Electropolymerization of Meta-Phenylenediamine (mPD)
Meta-phenylenediamine (mPD) served as an exclusion layer to increase the selectivity for D-serine. The layer of mPD functions as a size exclusion barrier, prohibiting electroactive substances, such as AA or DA, from reaching the platinum sites of the MEA [44]. mPD was electropolymerized onto all platinum sites (D-serine recording and sentinel channels) after coating using a cycled potential between +0.25 and +0.75 V vs. Ag/AgCl at a frequency of 0.05 V/s in a 5 mM mPD solution for 22 min (FAST electroplating tool, Quanteon, LLC, Lexington, KT, USA). Note, mPD electropolymerization occurs onto the surface of the platinum sites of the MEA, below the enzyme layer [44].
2.5. In Vitro Calibration and Recording Parameters
Enzyme coated MEAs were calibrated in vitro to determine their sensitivity to D-Serine (pA/µM D-serine), selectivity against ascorbic acid or glutamate, limit of detection (LOD), and linearity (R2) under an amperometric fixed potential of +0.7 V versus a glass Ag/AgCl reference electrode (RE-5B Ag/AgCl, ProSense, Oosterhout, The Netherlands). Signals were first preamplified using a microamplifier (2 pA/mV 500×; Quanteon, LLC, Lexington, KT, USA) and then amplified and digitized at 10 Hz with the FAST-16mkIII data acquisition system (Quanteon, LLC, Lexington, KT, USA). Standard calibrations were performed by immersing the MEAs in a calibration media, which consisted of phosphate buffered-saline (PBS, 40 mL). The solution was stirred using a magnetic stirrer device (Stuart, Bibby Scientific Limited, Staffordshire, UK) and kept at body temperature in a water bath chamber (Pronexus, Analytical AB, Stockholm, Sweden) at 37 °C controlled by a water heater system (Micro-Temp LT, Cincinnati Sub Zero, Cincinnati, OH, USA).
According to a standard protocol [43,44,45,53,54,55], after a stabilization period of at least 30 min, aliquots of AA, D-serine, DA, and H2O2 were given. For the D-serine calibration three consecutive 40 µL additions of a 20 mM D-serine solution produced three consecutive total concentrations in the beaker of 20 µM, 40 µM, and 60 µM. For AA the resulting concentration was 250 µM (500 µL of 20 mM), for DA 2 µM (40 µL of 2 mM), and for H2O2 8.8 µM (40 µL of 8.8 mM); n = 12 for RgDAAO coated and n = 4 for pkDAAO coated MEAs.
We selected these concentrations as they belong to an established in vitro calibration methodology using enzyme-based MEAs for the detection of other molecules like L-glutamate [43,44,45,53,54,55,56]. Thus, our calibration results are directly comparable to previous calibration measurements, although a much lower basal D-serine concentration in the prefrontal cortex of the anesthetized rat was reported (~3 µM) [28]. Moreover, calibrations using only 1 mM D-serine (reaching a final concentration in the calibration media of 1 µM) met the same threshold criteria (n = 5) as the calibrations using a 20 mM D-serine solution (Figure S1 and Table S1). Figure 2 depicts a typical calibration of a MEA from which the following parameters were derived: selectivity for D-serine against AA (concentration ratio of D-serine over AA), sensitivity to D-serine (pA/µM D-serine), limit of detection (LOD) for D-serine (in µM), and linearity (R2; FAST analysis 6.1 software; Jason Burmeister Consulting, LLC, KT, USA). Hereby, the responsivity of the MEA to changes in concentrations of D-serine is indicated by the sensitivity; the LOD (i.e., the lowest change in D-serine detected by the DAAO coated MEAs that cannot be attributed to noise, calculated as three times the standard deviation of the noise of the corresponding MEA channel during calibration), and the linear regression shows the linear response to the accumulating concentrations of D-serine that were added during the calibration [44,45]. Hydrogen peroxide was added to check for equal response performance to the reporter molecule of all the channels (data not shown) [42].
Two other calibrations of the RgDAAO coated MEAs were performed to test selectivity against L-glutamate and sensitivity to D-alanine. For the former, a final concentration of 20 µM of L-glutamate instead of AA were added to the calibration media (n = 10). D-alanine is a substrate for DAAO as well, thus the sensitivity to D-alanine was tested adding three aliquots of D-alanine to create a final concentration of 20 µM, 40 µM, and 60 µM in the calibration media (n = 12). All data are presented using mean ± standard error of the mean (SEM).
2.6. Surgery and Freely Moving Test of the D-Serine MEAs
As proof of principle, a RgDAAO coated MEA was implanted in vivo in a male Long Evans rat (15 weeks at time of surgery; Janvier, Le Genest-Saint-Isle, France). All of the experimental procedures were performed in accordance with the European animal protection laws and policies (directive 86/609, 1986, European Community) and were approved by the Schleswig-Holstein state authority. The animal was housed individually with ad libitum access to food and water under a 12 h/12 h light-dark cycle (lights on at 06:00 A.M.), and was handled seven days prior to surgery.
Stereotaxic surgery took place under isoflurane anesthesia. The MEA was implanted in the prefrontal cortex (AP: +2.5 mm, L: −0.5 mm, DV: 2.5 mm) using an anterior Ag/AgCl electrode as reference (AP: +5.5 mm, L: +1.0 mm, DV: 2.5 mm) [57]. The prefrontal cortex was selected due to its reported high levels of D-serine [3,35].
After seven days of recovery, the freely moving recording was performed in a dark PVC recording box with the same amperometric system as described for calibrations using cabling attached to a low torque slip-ring commutator (Dragonfly Research and Development, Inc., Ridgeley, WV, USA). D-serine signals were obtained after a one hour baseline recording during which channels stabilized.
Analyses were conducted with the FAST analysis 6.1 software as mentioned above. Only transients measured in the D-serine channels and not in the sentinel channels were included in the analyses.
3. Results and Discussion
3.1. In Vitro Calibration Results
Mean values for the calibration parameters are given in Table 1, together with the threshold criteria for sufficient MEA properties [44,45]. Our RgDAAO coated MEAs demonstrated a sensitivity of 8.61 ± 0.83 pA/µM to D-serine, a linearity of 0.9986 ± 0.0005 and an LOD of 0.17 ± 0.01 µM (mean ± SEM; n = 12, using a data acquisition rate of 10 Hz. In addition, all of the channels on a given MEA revealed a similar response to H2O2; meaning that D-serine recording and sentinel channels only differed in their response to D-serine (data not shown) [42].
3.2. Selectivity
In addition to the use of enzymes, selective detection of non-electroactive molecules, such as D-serine, is achieved by using size exclusion layers which form a barrier for larger interfering electroactive molecules (e.g., ascorbic acid). While in the past Nafion® was utilized as an exclusion layer [44,56], Meta-phenylenediamine (mPD) has proven to be a better barrier against interfering molecules as monoamines are attracted by Nafion® [45,46,58,59]. After electropolymerization of mPD onto the MEA only small molecules like H2O2 can readily cross this layer and reach the platinum sites. We tested our D-serine MEAs against AA and L-glutamate (Table 1) since both of them are found at high concentrations in the same brain regions as D-serine, and glutamate is involved in the release of both AA and D-serine [8,60,61,62,63]. Glutamate should not be detected by the D-serine-selective MEAs as a different enzyme (glutamate oxidase) is required to transform glutamate into a measurable compound, but AA is electroactive and therefore a possible interferent. Selectivity is the ratio of the sensitivity for D-serine over the interferent and has to fulfill our criteria of being higher than 80:1. This means an increase in interferent concentration above 80 µM would be necessary to produce a signal corresponding to a 1 µM increase of D-serine [44]. Our RgDAAO coated MEAs effectively blocked both AA and glutamate, with a mean selectivity for D-serine against AA and L-glutamate of 191.75 ± 19.55 and 521.3 ± 112.23, respectively (n = 12 and n = 10; mean ± SEM; Table 1).
3.3. Other D-Amino Acids
D-amino acids aside from D-serine can also be a substrate of D-amino acid oxidase. D-aspartate, which is the second most abundant D-amino acid, is not detected by D-serine biosensors because it is not a substrate of DAAO, but D-alanine, the third most abundant D-isomer found in the mammalian brain, can be detected (e.g., RgDAAO MEAs had a sensitivity of 18.56 ± 1.6 pA/µM to D-alanine; n = 12). Yet, the endogenous concentration of D-alanine in the brain is only 3% of the D-serine concentration and can be thus considered negligible for in vivo measurements [28,64].
3.4. RgDAAO vs. pkDAAO
The choice of DAAO from a yeast, Rhodotorula gracilis (RgDAAO), over the mammalian DAAO from porcine kidney (pkDAAO) was made given the previously reported superior enzymatic properties of RgDAAO [65,66]. Other studies have demonstrated a tolerable sensitivity using pkDAAO [28], but the parameters that were obtained from our first calibrations using pkDAAO did not fulfill our criteria: the sensitivity of the pkDAAO coated MEAs to D-serine was 0.45 ± 0.2 pA/µM and the LOD was 2.93 ± 0.7 µM (n = 4). Since the enzyme layers, BSA and glutaraldehyde layers were inspected under a stereoscope (0.8×–4×; Olympus Deutschland GmbH. Hamburg, Germany), as for all calibrated MEAs, the poor parameters obtained with pkDAAO cannot be attributed to peeling, cracking, or swelling of the coatings, the insulation layer or the platinum sites per se. Consequently, we employed D-amino acid oxidase from the yeast Rhodotorula gracilis for all further measurements.
3.5. Freely Moving Measurement of D-Serine
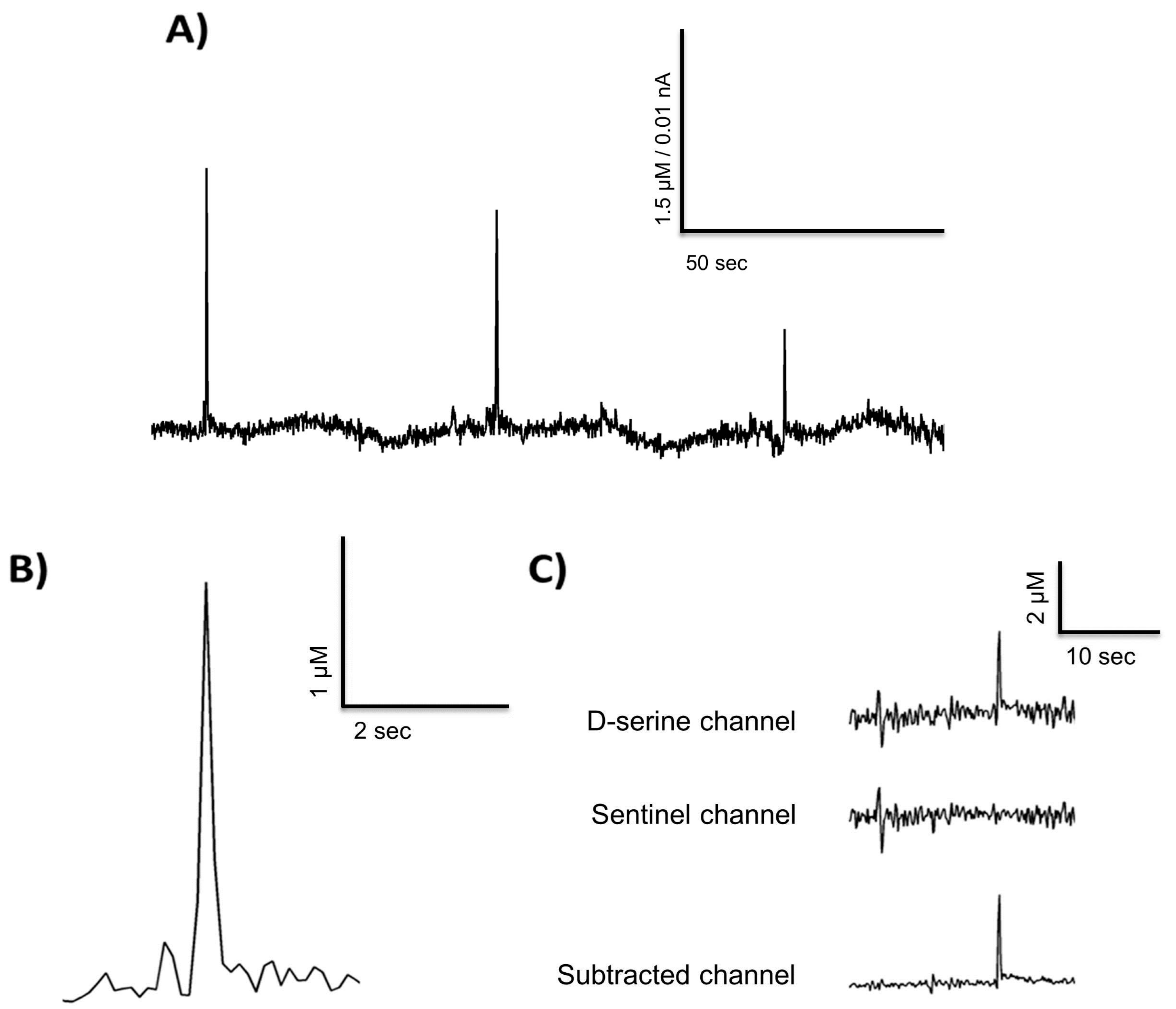
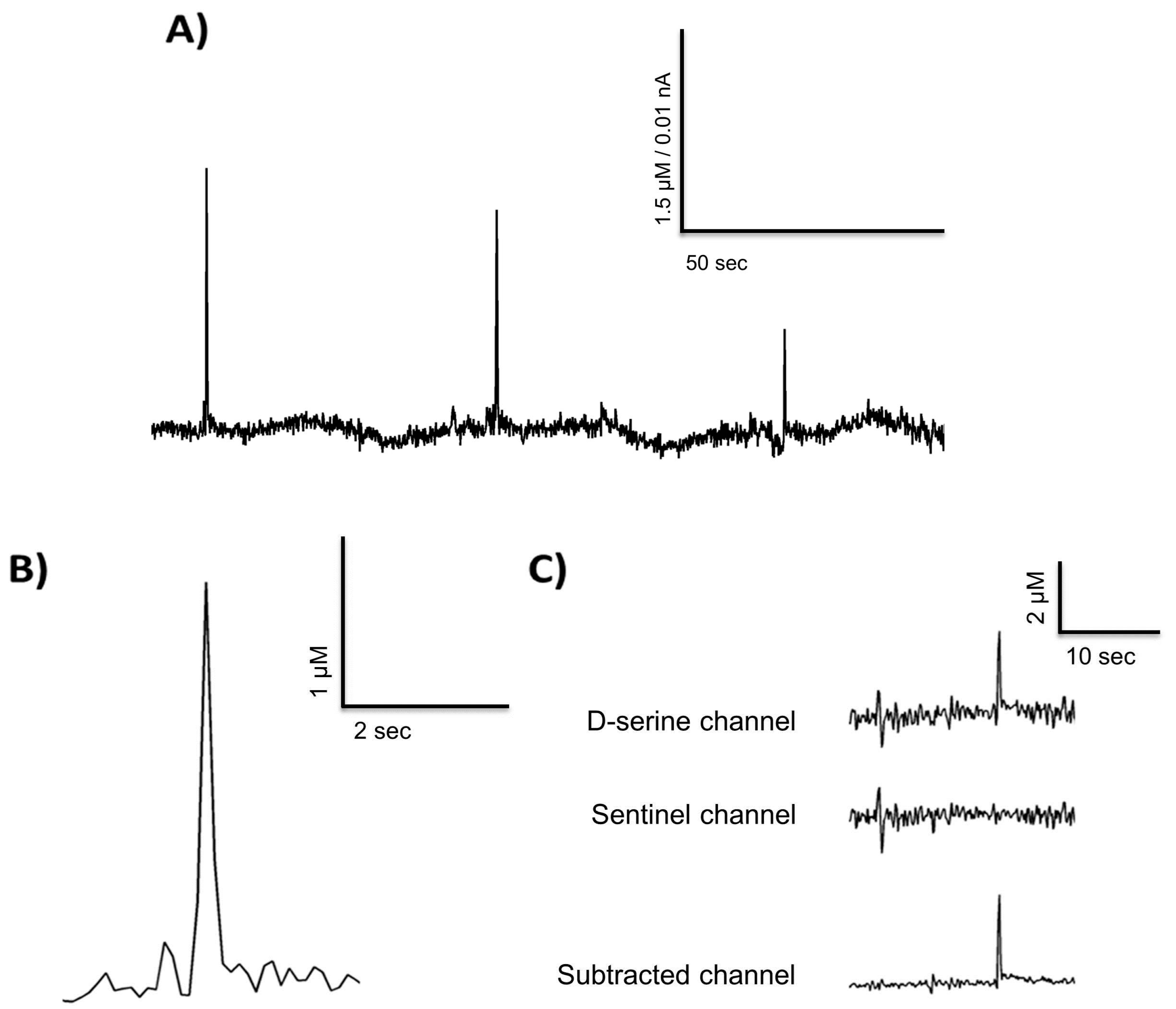
Figure 3A depicts spontaneous D-serine transients recorded from the prefrontal cortex in a freely moving animal. A close up of one transient is depicted in Figure 3B. These exemplary measurements in a freely moving animal suggest that D-serine could be released into the extracellular space in fast transients as has been reported for glutamate [41,43,44,45,56]. A number of 89 D-serine transients were found in a 10 h recording, with a mean frequency of five per hour. Thus, we demonstrate that RgDAAO coated MEAs can be used to record in vivo, but further measurements have to be performed on a larger sample of animals to characterize any changes in basal D-serine levels and in the parameters of the transients (e.g., amplitude, frequency, temporal occurrence) and relate them to behavioral states [32,42] of freely moving animals.
4. Conclusions
We demonstrate here a reliable in vitro protocol for preparing RgDAAO coated multisite MEAs for fast and selective amperometric measurements of D-serine. Our preliminary results in the freely moving recording support the need for a recording technique with a high temporal resolution to detect fast D-serine dynamics in the mammalian brain. Further in vivo studies using freely moving animals need to be conducted given the recent findings on the functions of D-serine within the central nervous system.
Supplementary Materials
The following are available online at https://www.mdpi.com/2079-6374/8/1/20/s1, Figure S1: In vitro calibration of one D-serine detecting Microelectrode Array (MEA) using 1 mM D-serine challenges, Figure S2: Linearity (R2) curve obtained from the calibration of the RgDAAO coated D-serine detecting MEA depicted in Figure 2, Table S1: In vitro calibration measurements for the RgDAAO coated D-serine detecting MEAs using final concentrations of 1 µM D-serine in the solution media (n = 5; mean ± SEM), Table S2: In vitro calibration measurements obtained from the calibration of the RgDAAO coated D-serine detecting MEA depicted in Figure 2.
Acknowledgments
We thank Sonja Binder, Katia Monsorno, Ines Stölting, Horst Koller, Andreas Moser and Jose Ostos Muraday for technical assistance and advice. This work was supported by Deutsche Forschungsgemeinschaft SFBTR654 Plasticity and Sleep (TPA06) and Deutsche Forschungsgemeinschaft SPP1665 (MA2053/4-2).
Author Contributions
D.C.B. conducted the experimental procedures and analyzed the data. D.C.B. and L.M. planned and Å.K.-G. and J.E.Q. assisted with the experimental procedures. The manuscript was written through contributions of all authors. All authors have given approval to the final version of the manuscript.
Conflicts of Interest
J.E.Q. serves as a consultant to Quanteon LLC which manufactures the FAST system. D.C.B., Å.K.-G. and L.M. declare no conflict of interest.
References
- Hashimoto, A.; Nishikawa, T.; Hayashi, T.; Fujii, N.; Harada, K.; Oka, T.; Takahashi, K. The presence of free d-serine in rat brain. FEBS Lett. 1992, 296, 33–36. [Google Scholar] [CrossRef]
- Hashimoto, A.; Nishikawa, T.; Oka, T.; Takahashi, K. Endogenous d-serine in rat brain: N-methyl-d-aspartate receptor-related distribution and aging. J. Neurochem. 1993, 60, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Schell, M.J.; Molliver, M.E.; Snyder, S.H. d-D-serine, an endogenous synaptic modulator: Localization to astrocytes and glutamate-stimulated release. Proc. Natl. Acad. Sci. USA 1995, 92, 3948–3952. [Google Scholar] [CrossRef] [PubMed]
- Schell, M.J.; Brady, R.O.; Molliver, M.E.; Snyder, S.H. d-serine as a neuromodulator: Regional and developmental localizations in rat brain glia resemble nmda receptors. J. Neurosci. 1997, 17, 1604–1615. [Google Scholar] [PubMed]
- Balu, D.T.; Presti, K.T.; Huang, C.C.Y.; Muszynski, K.; Radzishevsky, I.; Wolosker, H.; Guffanti, G.; Ressler, K.J.; Coyle, J.T. Serine racemase and d-serine in the amygdala are dynamically involved in fear learning. Biol. Psychiatry 2018, 83, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Mothet, J.P.; Parent, A.T.; Wolosker, H.; Brady, R.O., Jr.; Linden, D.J.; Ferris, C.D.; Rogawski, M.A.; Snyder, S.H. d-serine is an endogenous ligand for the glycine site of the n-methyl-d-aspartate receptor. Proc. Natl. Acad. Sci. USA 2000, 97, 4926–4931. [Google Scholar] [CrossRef] [PubMed]
- Panatier, A.; Theodosis, D.T.; Mothet, J.P.; Touquet, B.; Pollegioni, L.; Poulain, D.A.; Oliet, S.H. Glia-derived d-serine controls nmda receptor activity and synaptic memory. Cell 2006, 125, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Martineau, M.; Baux, G.; Mothet, J.P. d-serine signalling in the brain: Friend and foe. Trends Neurosci. 2006, 29, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Wolosker, H.; Dumin, E.; Balan, L.; Foltyn, V.N. d-amino acids in the brain: d-serine in neurotransmission and neurodegeneration. FEBS J. 2008, 275, 3514–3526. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.C.; Tsai, G.E.; Ma, C.L.; Ehmsen, J.T.; Mustafa, A.K.; Han, L.; Jiang, Z.I.; Benneyworth, M.A.; Froimowitz, M.P.; Lange, N.; et al. Targeted disruption of serine racemase affects glutamatergic neurotransmission and behavior. Mol. Psychiatry 2009, 14, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Henneberger, C.; Papouin, T.; Oliet, S.H.; Rusakov, D.A. Long-term potentiation depends on release of d-serine from astrocytes. Nature 2010, 463, 232–236. [Google Scholar] [CrossRef] [PubMed]
- DeVito, L.M.; Balu, D.T.; Kanter, B.R.; Lykken, C.; Basu, A.C.; Coyle, J.T.; Eichenbaum, H. Serine racemase deletion disrupts memory for order and alters cortical dendritic morphology. Genes Brain Behav. 2011, 10, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Fossat, P.; Turpin, F.R.; Sacchi, S.; Dulong, J.; Shi, T.; Rivet, J.M.; Sweedler, J.V.; Pollegioni, L.; Millan, M.J.; Oliet, S.H.; et al. Glial d-serine gates nmda receptors at excitatory synapses in prefrontal cortex. Cereb. Cortex 2012, 22, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Henneberger, C.; Bard, L.; Rusakov, D.A. d-serine: A key to synaptic plasticity? Int. J. Biochem. Cell Biol. 2012, 44, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Curcio, L.; Podda, M.V.; Leone, L.; Piacentini, R.; Mastrodonato, A.; Cappelletti, P.; Sacchi, S.; Pollegioni, L.; Grassi, C.; D’Ascenzo, M. Reduced d-serine levels in the nucleus accumbens of cocaine-treated rats hinder the induction of nmda receptor-dependent synaptic plasticity. Brain 2013, 136, 1216–1230. [Google Scholar] [CrossRef] [PubMed]
- Balu, D.T.; Takagi, S.; Puhl, M.D.; Benneyworth, M.A.; Coyle, J.T. d-serine and serine racemase are localized to neurons in the adult mouse and human forebrain. Cell. Mol. Neurobiol. 2014, 34, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.; Li, L.; Moss, J.; Petrelli, F.; Casse, F.; Gebara, E.; Lopatar, J.; Pfrieger, F.W.; Bezzi, P.; Bischofberger, J.; et al. Synaptic integration of adult-born hippocampal neurons is locally controlled by astrocytes. Neuron 2015, 88, 957–972. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Crux, S.; Marinesco, S.; Montagna, E.; Sgobio, C.; Shi, Y.; Shi, S.; Zhu, K.; Dorostkar, M.M.; Muller, U.C.; et al. Amyloid precursor protein maintains constitutive and adaptive plasticity of dendritic spines in adult brain by regulating d-serine homeostasis. EMBO J. 2016, 35, 2213–2222. [Google Scholar] [CrossRef] [PubMed]
- Durrant, A.R.; Heresco-Levy, U. d-serine in neuropsychiatric disorders: New advances. Adv. Psychiatry 2014, 2014, 16. [Google Scholar] [CrossRef]
- Balu, D.T.; Coyle, J.T. The nmda receptor ‘glycine modulatory site’ in schizophrenia: d-serine, glycine, and beyond. Curr. Opin. Pharmacol. 2015, 20, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Madeira, C.; Lourenco, M.V.; Vargas-Lopes, C.; Suemoto, C.K.; Brandao, C.O.; Reis, T.; Leite, R.E.; Laks, J.; Jacob-Filho, W.; Pasqualucci, C.A.; et al. d-serine levels in alzheimer’s disease: Implications for novel biomarker development. Transl. Psychiatry 2015, 5, e561. [Google Scholar] [CrossRef] [PubMed]
- Wolosker, H.; Balu, D.T.; Coyle, J.T. The rise and fall of the d-serine-mediated gliotransmission hypothesis. Trends Neurosci. 2016, 39, 712–721. [Google Scholar] [CrossRef] [PubMed]
- Pollegioni, L.; Sacchi, S. Metabolism of the neuromodulator d-serine. Cell. Mol. Life Sci. 2010, 67, 2387–2404. [Google Scholar] [CrossRef] [PubMed]
- Papouin, T.; Henneberger, C.; Rusakov, D.A.; Oliet, S.H.R. Astroglial versus neuronal d-serine: Fact checking. Trends Neurosci. 2017, 40, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Wolosker, H.; Balu, D.T.; Coyle, J.T. Astroglial versus neuronal d-serine: Check your controls! Trends Neurosci. 2017, 40, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Fiacco, T.A.; McCarthy, K.D. Multiple lines of evidence indicate that gliotransmission does not occur under physiological conditions. J. Neurosci. 2018, 38, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Savtchouk, I.; Volterra, A. Gliotransmission: Beyond black-and-white. J. Neurosci. 2018, 38, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Pernot, P.; Mothet, J.P.; Schuvailo, O.; Soldatkin, A.; Pollegioni, L.; Pilone, M.; Adeline, M.T.; Cespuglio, R.; Marinesco, S. Characterization of a yeast d-amino acid oxidase microbiosensor for d-serine detection in the central nervous system. Anal. Chem. 2008, 80, 1589–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zain, Z.M.; O’Neill, R.D.; Lowry, J.P.; Pierce, K.W.; Tricklebank, M.; Dewa, A.; Ab Ghani, S. Development of an implantable d-serine biosensor for in vivo monitoring using mammalian d-amino acid oxidase on a poly (O-phenylenediamine) and nafion-modified platinum-iridium disk electrode. Biosens. Bioelectron. 2010, 25, 1454–1459. [Google Scholar] [CrossRef] [PubMed]
- Pernot, P.; Maucler, C.; Tholance, Y.; Vasylieva, N.; Debilly, G.; Pollegioni, L.; Cespuglio, R.; Marinesco, S. d-serine diffusion through the blood-brain barrier: Effect on d-serine compartmentalization and storage. Neurochem. Int. 2012, 60, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Shigetomi, E.; Jackson-Weaver, O.; Huckstepp, R.T.; O’Dell, T.J.; Khakh, B.S. Trpa1 channels are regulators of astrocyte basal calcium levels and long-term potentiation via constitutive d-serine release. J. Neurosci. 2013, 33, 10143–10153. [Google Scholar] [CrossRef] [PubMed]
- Papouin, T.; Dunphy, J.M.; Tolman, M.; Dineley, K.T.; Haydon, P.G. Septal cholinergic neuromodulation tunes the astrocyte-dependent gating of hippocampal nmda receptors to wakefulness. Neuron 2017, 94, 840–854. [Google Scholar] [CrossRef] [PubMed]
- Polcari, D.; Kwan, A.; Van Horn, M.R.; Danis, L.; Pollegioni, L.; Ruthazer, E.S.; Mauzeroll, J. Disk-shaped amperometric enzymatic biosensor for in vivo detection of d-serine. Anal. Chem. 2014, 86, 3501–3507. [Google Scholar] [CrossRef] [PubMed]
- Polcari, D.; Perry, S.C.; Pollegioni, L.; Geissler, M.; Mauzeroll, J. Localized detection of d-serine by using an enzymatic amperometric biosensor and scanning electrochemical microscopy. ChemElectroChem 2017, 4, 920–926. [Google Scholar] [CrossRef]
- Hashimoto, A.; Oka, T.; Nishikawa, T. Extracellular concentration of endogenous free d-serine in the rat brain as revealed by in vivo microdialysis. Neuroscience 1995, 66, 635–643. [Google Scholar] [CrossRef]
- Fukushima, T.; Kawai, J.; Imai, K.; Toyo’oka, T. Simultaneous determination of d- and l-serine in rat brain microdialysis sample using a column-switching HPLC with fluorimetric detection. Biomed. Chromatogr. 2004, 18, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Onozato, M.; Nakazawa, H.; Ishimaru, K.; Nagashima, C.; Fukumoto, M.; Hakariya, H.; Sakamoto, T.; Ichiba, H.; Fukushima, T. Alteration in plasma and striatal levels of d-serine after d-serine administration with or without nicergoline: An in vivo microdialysis study. Heliyon 2017, 3, e00399. [Google Scholar] [CrossRef] [PubMed]
- Umino, A.; Ishiwata, S.; Iwama, H.; Nishikawa, T. Evidence for tonic control by the gabaa receptor of extracellular d-serine concentrations in the medial prefrontal cortex of rodents. Front. Mol. Neurosci. 2017, 10, 240. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.L.; Wightman, R.M. Rapid dopamine release in freely moving rats. In Electrochemical Methods for Neuroscience; Michael, A.C., Borland, L.M., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2007. [Google Scholar]
- Eschenko, O.; Sara, S.J. Learning-dependent, transient increase of activity in noradrenergic neurons of locus coeruleus during slow wave sleep in the rat: Brain stem-cortex interplay for memory consolidation? Cereb. Cortex 2008, 18, 2596–2603. [Google Scholar] [CrossRef] [PubMed]
- Hascup, K.N.; Hascup, E.R.; Stephens, M.L.; Glaser, P.E.; Yoshitake, T.; Mathe, A.A.; Gerhardt, G.A.; Kehr, J. Resting glutamate levels and rapid glutamate transients in the prefrontal cortex of the flinders sensitive line rat: A genetic rodent model of depression. Neuropsychopharmacology 2011, 36, 1769–1777. [Google Scholar] [CrossRef] [PubMed]
- Dash, M.B.; Douglas, C.L.; Vyazovskiy, V.V.; Cirelli, C.; Tononi, G. Long-term homeostasis of extracellular glutamate in the rat cerebral cortex across sleep and waking states. J. Neurosci. 2009, 29, 620–629. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, J.J.; Gerhardt, G.A. Self-referencing ceramic-based multisite microelectrodes for the detection and elimination of interferences from the measurement of l-glutamate and other analytes. Anal. Chem. 2001, 73, 1037–1042. [Google Scholar] [CrossRef] [PubMed]
- Hascup, K.N.; Rutherford, E.C.; Quintero, J.E.; Day, B.K.; Nickell, J.R.; Pomerleau, F.; Huettl, P.; Burmeister, J.J.; Gerhardt, G.A. Second-by-second measures of l-glutamate and other neurotransmitters using enzyme-based microelectrode arrays. In Electrochemical Methods for Neuroscience; Michael, A.C., Borland, L.M., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2007. [Google Scholar]
- Mishra, D.; Harrison, N.R.; Gonzales, C.B.; Schilstrom, B.; Konradsson-Geuken, A. Effects of age and acute ethanol on glutamatergic neurotransmission in the medial prefrontal cortex of freely moving rats using enzyme-based microelectrode amperometry. PLoS ONE 2015, 10, e0125567. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.M.; Quintero, J.E.; Pomerleau, F.; Huettl, P.; Gerhardt, G.A.; Glaser, P.E. Simultaneous glutamate recordings in the frontal cortex network with multisite biomorphic microelectrodes: New tools for adhd research. J. Neurosci. Methods 2015, 252, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, J.J.; Pomerleau, F.; Huettl, P.; Gash, C.R.; Werner, C.E.; Bruno, J.P.; Gerhardt, G.A. Ceramic-based multisite microelectrode arrays for simultaneous measures of choline and acetylcholine in CNS. Biosens. Bioelectron. 2008, 23, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, C.F.; Ledo, A.; Gerhardt, G.A.; Laranjinha, J.; Barbosa, R.M. Neurometabolic and electrophysiological changes during cortical spreading depolarization: Multimodal approach based on a lactate-glucose dual microbiosensor arrays. Sci. Rep. 2017, 7, 6764. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lin, S.C.; Nicolelis, M.A. Acquiring local field potential information from amperometric neurochemical recordings. J. Neurosci. Methods 2009, 179, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Ledo, A.; Lourenco, C.F.; Laranjinha, J.; Gerhardt, G.A.; Barbosa, R.M. Combined in vivo amperometric oximetry and electrophysiology in a single sensor: A tool for epilepsy research. Anal. Chem. 2017, 89, 12383–12390. [Google Scholar] [CrossRef] [PubMed]
- Hascup, E.R.; af Bjerken, S.; Hascup, K.N.; Pomerleau, F.; Huettl, P.; Stromberg, I.; Gerhardt, G.A. Histological studies of the effects of chronic implantation of ceramic-based microelectrode arrays and microdialysis probes in rat prefrontal cortex. Brain Res. 2009, 1291, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Fantinato, S.; Pollegioni, L.; Pilone, M.S. Engineering, expression and purification of a his-tagged chimeric d-amino acid oxidase from rhodotorula gracilis. Enzyme Microb. Technol. 2001, 29, 407–412. [Google Scholar]
- Konradsson-Geuken, A.; Gash, C.R.; Alexander, K.; Pomerleau, F.; Huettl, P.; Gerhardt, G.A.; Bruno, J.P. Second-by-second analysis of alpha 7 nicotine receptor regulation of glutamate release in the prefrontal cortex of awake rats. Synapse 2009, 63, 1069–1082. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, J.J.; Pomerleau, F.; Palmer, M.; Day, B.K.; Huettl, P.; Gerhardt, G.A. Improved ceramic-based multisite microelectrode for rapid measurements of l-glutamate in the cns. J. Neurosci. Methods 2002, 119, 163–171. [Google Scholar] [CrossRef]
- Day, B.K.; Pomerleau, F.; Burmeister, J.J.; Huettl, P.; Gerhardt, G.A. Microelectrode array studies of basal and potassium-evoked release of l-glutamate in the anesthetized rat brain. J. Neurochem. 2006, 96, 1626–1635. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, E.C.; Pomerleau, F.; Huettl, P.; Stromberg, I.; Gerhardt, G.A. Chronic second-by-second measures of l-glutamate in the central nervous system of freely moving rats. J. Neurochem. 2007, 102, 712–722. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press/Elsevier: Amsterdam, The Netherlands; Boston, MA, USA, 2007. [Google Scholar]
- Hinzman, J.M.; Thomas, T.C.; Burmeister, J.J.; Quintero, J.E.; Huettl, P.; Pomerleau, F.; Gerhardt, G.A.; Lifshitz, J. Diffuse brain injury elevates tonic glutamate levels and potassium-evoked glutamate release in discrete brain regions at two days post-injury: An enzyme-based microelectrode array study. J. Neurotrauma 2010, 27, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Bortz, D.M.; Jorgensen, C.V.; Mikkelsen, J.D.; Bruno, J.P. Transient inactivation of the ventral hippocampus in neonatal rats impairs the mesolimbic regulation of prefrontal glutamate release in adulthood. Neuropharmacology 2014, 84, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Grunewald, R.A. Ascorbic acid in the brain. Brain Res. Brain Res. Rev. 1993, 18, 123–133. [Google Scholar] [CrossRef]
- Harrison, F.E.; May, J.M. Vitamin c function in the brain: Vital role of the ascorbate transporter SVCT2. Free Radic. Biol. Med. 2009, 46, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, V.; Storm-Mathisen, J.; Bergersen, L.H. Neuroglial transmission. Phys. Rev. 2015, 95, 695–726. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, N.R.; Ledo, A.; Laranjinha, J.; Gerhardt, G.A.; Barbosa, R.M. Simultaneous measurements of ascorbate and glutamate in vivo in the rat brain using carbon fiber nanocomposite sensors and microbiosensor arrays. Bioelectrochemistry 2018, 121, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Hamase, K.; Konno, R.; Morikawa, A.; Zaitsu, K. Sensitive determination of d-amino acids in mammals and the effect of d-amino-acid oxidase activity on their amounts. Biol. Pharm. Bull. 2005, 28, 1578–1584. [Google Scholar] [CrossRef] [PubMed]
- Pollegioni, L.; Ghisla, S.; Pilone, M.S. Studies on the active centre of rhodotorula gracilis d-amino acid oxidase and comparison with pig kidney enzyme. Biochem. J. 1992, 286 Pt 2, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Molla, G.; Vegezzi, C.; Pilone, M.S.; Pollegioni, L. Overexpression in escherichia coli of a recombinant chimeric rhodotorula gracilis d-amino acid oxidase. Protein Expr. Purif. 1998, 14, 289–294. [Google Scholar] [CrossRef] [PubMed]
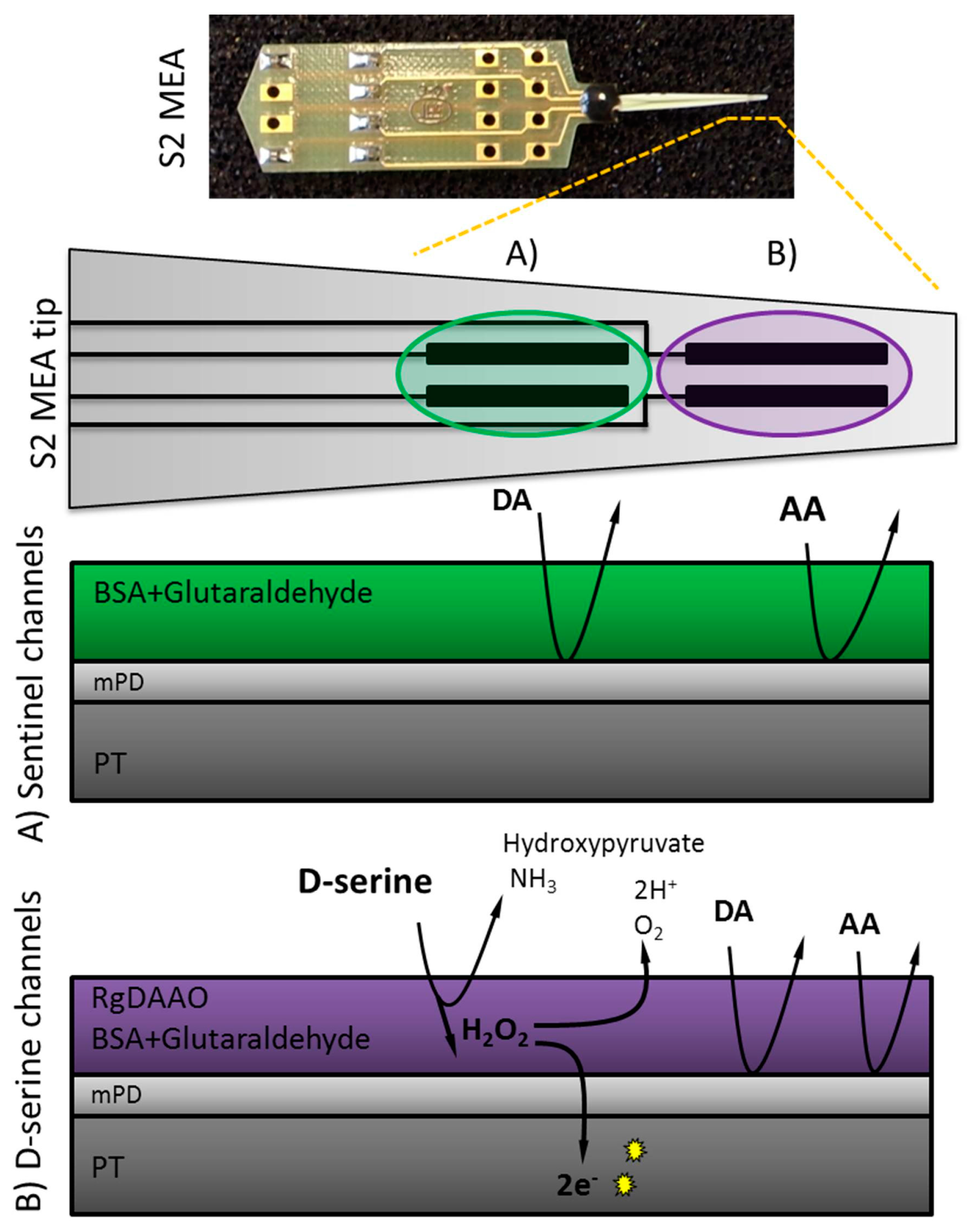
Figure 1.
Scheme for D-serine detection using S2 style Microelectrode Array (MEAs) consisting of two pairs of platinum (PT) sites. When D-serine comes into contact with Rhodotorula gracilis (RgDAAO) (platinum sites B: D-serine recording channels) H2O2 is produced by the enzymatic reaction, crosses the mPD barrier and is then further oxidized, yielding two electrons per molecule. At the amperometric fixed potential of +0.7 V, the sentinel channels (platinum sites A) detect only electrochemically active interferents and background noise, whereas the RgDAAO coated channels measure in addition D-serine.
Figure 1.
Scheme for D-serine detection using S2 style Microelectrode Array (MEAs) consisting of two pairs of platinum (PT) sites. When D-serine comes into contact with Rhodotorula gracilis (RgDAAO) (platinum sites B: D-serine recording channels) H2O2 is produced by the enzymatic reaction, crosses the mPD barrier and is then further oxidized, yielding two electrons per molecule. At the amperometric fixed potential of +0.7 V, the sentinel channels (platinum sites A) detect only electrochemically active interferents and background noise, whereas the RgDAAO coated channels measure in addition D-serine.
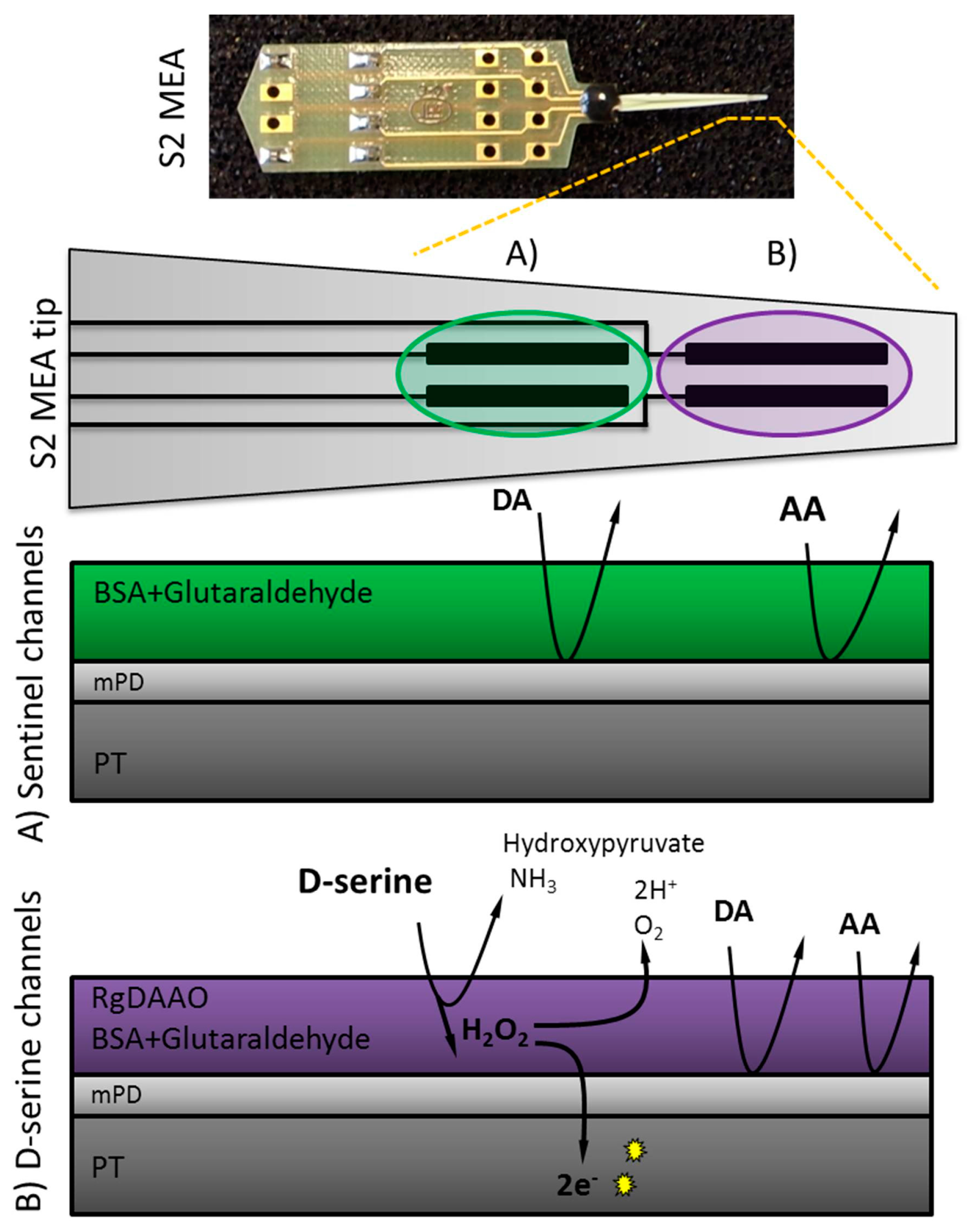
Figure 2.
In vitro calibration of one D-serine detecting Microelectrode Array (MEA). Arrows indicate when substances were added and the resultant concentration obtained in the calibration media. The figure reflects the current (nA) and corresponding concentration (µM) measured by the MEA after each challenge. Channels 1 and 2 are D-serine detecting, and channels 3 and 4 are sentinel channels, respectively. See Table S2 and Figure S2 for parameters of this calibration, and Figure S1 and Table S1 for the calibration using 1 mM D-serine.
Figure 2.
In vitro calibration of one D-serine detecting Microelectrode Array (MEA). Arrows indicate when substances were added and the resultant concentration obtained in the calibration media. The figure reflects the current (nA) and corresponding concentration (µM) measured by the MEA after each challenge. Channels 1 and 2 are D-serine detecting, and channels 3 and 4 are sentinel channels, respectively. See Table S2 and Figure S2 for parameters of this calibration, and Figure S1 and Table S1 for the calibration using 1 mM D-serine.

Figure 3.
D-serine transients from a freely moving recording in the rat prefrontal cortex (A,B) and a representative example of the self-referencing method used (C).
Figure 3.
D-serine transients from a freely moving recording in the rat prefrontal cortex (A,B) and a representative example of the self-referencing method used (C).

{kind=link}
{kind=link}
{kind=link}
Table 1.
In vitro measurements for the RgDAAO coated D-serine detecting MEAs as compared to threshold criteria (n = 12; mean ± SEM). To test for selectivity against L-Glutamate 10 calibrations were performed.
Table 1.
In vitro measurements for the RgDAAO coated D-serine detecting MEAs as compared to threshold criteria (n = 12; mean ± SEM). To test for selectivity against L-Glutamate 10 calibrations were performed.
| Parameters | Sensitivity | LOD | Linearity (R2) | Selectivity Against AA | Selectivity Against L-Glutamate |
|---|---|---|---|---|---|
| Threshold criteria | ≥2 pA/µM | ≤0.5 µM | 1 | ≥80 | ≥80 |
| Measurements | 8.61 ± 0.83 pA/µM | 0.17 ± 0.01 µM | 0.9986 ± 0.0005 | 191.75 ± 19.55 | 521.3 ± 112.23 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Campos-Beltrán, D.; Konradsson-Geuken, Å.; Quintero, J.E.; Marshall, L. Amperometric Self-Referencing Ceramic Based Microelectrode Arrays for D-Serine Detection. Biosensors 2018, 8, 20. https://doi.org/10.3390/bios8010020
AMA Style
Campos-Beltrán D, Konradsson-Geuken Å, Quintero JE, Marshall L. Amperometric Self-Referencing Ceramic Based Microelectrode Arrays for D-Serine Detection. Biosensors. 2018; 8(1):20. https://doi.org/10.3390/bios8010020
Chicago/Turabian StyleCampos-Beltrán, Diana, Åsa Konradsson-Geuken, Jorge E. Quintero, and Lisa Marshall. 2018. "Amperometric Self-Referencing Ceramic Based Microelectrode Arrays for D-Serine Detection" Biosensors 8, no. 1: 20. https://doi.org/10.3390/bios8010020
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.