Multidrug Efflux Systems in Helicobacter cinaedi

Department of Microbiology, School of Pharmacy, Aichi Gakuin University/ 1-100 Kusumoto, Chikusa, Nagoya, Aichi 464-8650, Japan
*
Author to whom correspondence should be addressed.
Antibiotics 2012, 1(1), 29-43; https://doi.org/10.3390/antibiotics1010029
Submission received: 29 October 2012
/
Revised: 16 November 2012
/
Accepted: 16 November 2012
/
Published: 21 November 2012
(This article belongs to the Special Issue Efflux Pumps of Multi-Drug Resistant Pathogenic Bacteria: Their Genetic Regulation, Molecular Biology and Possible Control with Newly Designed Agents)
Abstract
:Helicobacter cinaedi causes infections, such as bacteremia, diarrhea and cellulitis in mainly immunocompromised patients. This pathogen is often problematic to analyze, and insufficient information is available, because it grows slowly and poorly in subculture under a microaerobic atmosphere. The first-choice therapy to eradicate H. cinaedi is antimicrobial chemotherapy; however, its use is linked to the development of resistance. Although we need to understand the antimicrobial resistance mechanisms of H. cinaedi, unfortunately, sufficient genetic tools for H. cinaedi have not yet been developed. In July 2012, the complete sequence of H. cinaedi strain PAGU 611, isolated from a case of human bacteremia, was announced. This strain possesses multidrug efflux systems, intrinsic antimicrobial resistance mechanisms and typical mutations in gyrA and the 23S rRNA gene, which are involved in acquired resistance to fluoroquinolones and macrolides, respectively. Here, we compare the organization and properties of the efflux systems of H. cinaedi with the multidrug efflux systems identified in other bacteria.
1. Introduction
Helicobacter cinaedi is a motile, Gram-negative, spiral bacterium belonging to the enterohepatic group of Helicobacter species of genus Helicobacter (the other group consists of gastric Helicobacter species, whose most well-known representative is the infamous H. pylori) [1]. During the last two decades, this bacterium has increasingly been recognized as a human pathogen that causes infections such as bacteremia, diarrhea and cellulitis in mainly immune-compromised patients and occasionally in immunocompetent ones with a high potential for recurrence [2,3]. A possible association between H. cinaedi and atrial arrhythmias and atherosclerosis was also suggested [4].
This pathogen grows slowly over several days on blood agar, even at its optimal conditions, such as a wet microaerobic atmosphere at 37 °C, and often appears as a swarming thin film that is difficult to observe [1,5]. Therefore, it is often problematic to isolate, detect and sub-culture [5,6]. Antimicrobial chemotherapy has been used successfully to treat such infections, but prolonged courses of multiple antimicrobials for at least 2–3 weeks may be required [1]. Recently, molecular epidemiological analysis in Japan showed that all H. cinaedi isolates since 2000 had acquired resistance to clarithromycin (macrolides) and ciprofloxacin (quinolones), for which the MIC90 (μg/mL) was >128 and 128, respectively, and contained typical mutations in gyrA and the 23S rRNA gene, respectively [7,8]. Unlike H. pylori, enteric Helicobacter species, such as H. cinaedi, are intrinsically resistant to amoxicillin (penicillin) [9]. High-level resistance and intrinsic resistance often require the presence of endogenous multidrug efflux pumps [10,11], which have not yet been analyzed in H. cinaedi.
2. Multidrug Efflux Systems in Bacteria
Multidrug efflux transporters are fundamental antimicrobial resistance mechanisms in Gram-negative bacteria [12]. Multidrug efflux transport has been studied extensively in bacteria, including ε-proteobacteria, such as H. pylori and Campylobacter jejuni, but not H. cinaedi [e.g., 13,14]. Most bacterial multidrug efflux pumps function as secondary transporters coupled with the proton-motive force (e.g., AcrB of Escherichia coli [15], MdfA of E. coli [16], and EmrE of E. coli [17]) and, although very rare, the sodium-motive force (e.g., NorM of Vibrio parahaemolyticus [18]), while some pumps hydrolyze ATP (e.g., MacB of E. coli [19] and VcaM of Vibrio cholerae [20]). Multidrug efflux transporters can be single component transporters that act at the cytoplasmic membrane (e.g., MdfA and EmrE of E. coli and NorM of V. parahaemolyticus) in both Gram-negative and -positive bacteria or three component transporters that span the entirety of the Gram-negative cell envelope (e.g., AcrAB-TolC of E. coli and MexXY-OprM of Pseudomonas aeruginosa [21]), i.e., cytoplasmic membrane transporter component (e.g., AcrB and MexY), outer membrane factor (OMF) component (e.g., TolC and OprM) and periplasmic component belonging to the membrane fusion protein (MFP) family (e.g., AcrA and MexX) [13,14]. Although bacterial multidrug efflux transporters fall into five families, i.e., resistance nodulation cell division (RND) (e.g., AcrB of E. coli and MexY of P. aeruginosa), major facilitator (MF) (e.g., MdfA of E. coli), small multidrug resistance (SMR) (e.g., EmrE of E. coli), multi-antimicrobial and toxic extrusion (MATE) (e.g., NorM of V. parahaemolyticus) and ABC (ATP binding cassette) (e.g., VcaM of V. cholerae)), the RND family is the most clinically relevant in Gram-negative bacteria [14].
3. Antibacterial Resistance Revealed by the Complete Genome of H. cinaedi
Very recently, we announced the complete genome sequence of H. cinaedi PAGU 611 isolated from a case of human bacteremia in Japan [22]. The clinical microbiological aspect of this strain was described as H. cinaedi-case 1; strain 923 [3]. Three months after our original report, another group published the sequence of the strain ATCC BAA-847, which was isolated in the 1980s in the USA [23]. The genome sequence of H. cinaedi CCUG 18818, although just a whole genome assembly and not complete, is also available from the Human Microbiome Project [24]. H. cinaedi PAGU 611 had a threonine to isoleucine mutation at position 84 of GyrA and adenine to guanine at position 2060 in PAGU 611 and ATCC BAA-847 (position 2018 in CCUG 18818) in the 23S rRNA gene, both of which are the same mutations identified in recent ciprofloxacin- and clarithromycin-resistant H. cinaedi isolates in Japan [7,8]. In addition to the slow, poor and, sometimes, failed growth described above, genetic tools for H. cinaedi are not sufficiently developed to take full advantage of the wealth of information generated by genome sequencing and to elucidate the function of unknown genes identified through sequencing. Fortunately, gene replacement via homologous replacement in H. cinaedi is possible by electroporation; however, no complementation system, e.g., a plasmid vector, is currently available for this organism [25]. We identified 10 putative drug transporter genes (2 RND, 1 MF, 2 MATE, 1 ABC, 4 SMR) in the genome of H. cinaedi PAGU 611 [22] (Figure 1). All transporters have homologues in H. hepaticus ATCC 51449, while only two-fifths are in H. pylori 26695 (Table 1). Interestingly C. jejuni subsp. jejuni NCTC 11168 has, rather, the most homologues (Table 1). Here, we compare the organization and properties of the multidrug efflux systems of H. cinaedi with the characterized and uncharacterized pumps available in the database.
4. RND Efflux Gene Operons of H. cinaedi
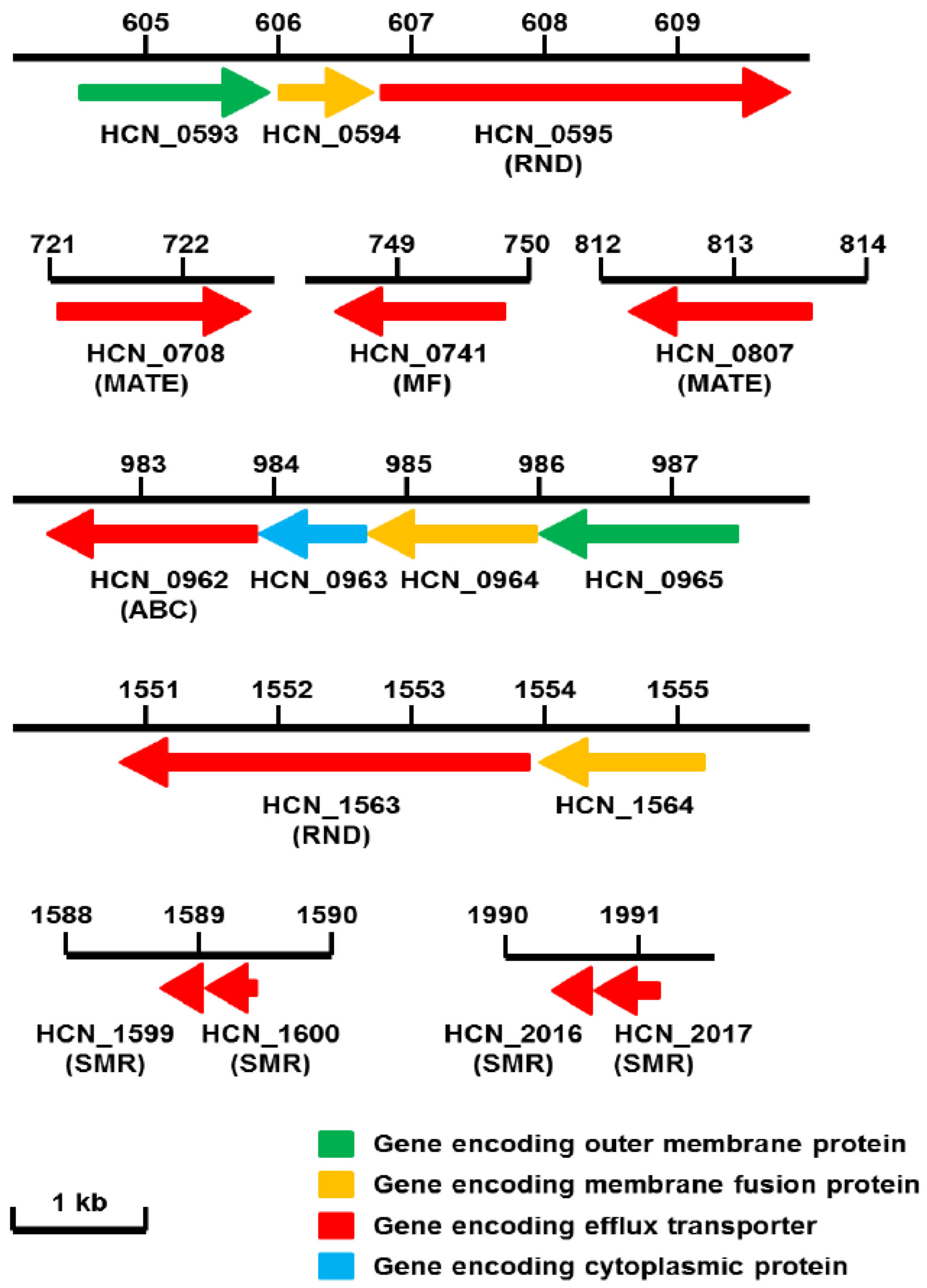
We identified two open reading frames (ORFs) belonging to the hydrophobe/amphiphile efflux-1 (HAE1) sub-family [26] of the RND family (locus-tags HCN_0595 and HCN_1563) encoded in the 2.08 Mbp chromosome of H. cinaedi PAGU 611 (Figure 1). One consists of three genes (HCN_0593-HCN_0594-HCN_0595) that encode OMP, MFP and RND, respectively, and the other consists of two genes (HCN_1564-HCN_1563) that encode MFP and RND, respectively. The ORFs were obtained from the chromosomes of ATCC BAA-847 and CCUG 18818. Both a three-gene operon (MFP, RND, and OMF) and a two-gene operon (MFP and RND) are genetically common as a multidrug efflux operon, while the latter is functionally associated with an OMF component that is encoded by a separate gene that is physically unattached to the other two members on the chromosome. For example, in P. aeruginosa PAO1, mexAB-oprM and mexXY encode two multidrug efflux pumps (MexAB-OprM and MexXY-OprM, respectively) and contribute to natural antimicrobial resistance [27]. However, three-gene RND-type multidrug efflux operons (e.g., mexAB-oprM of P. aeruginosa [28] and cmeABC of C. jejuni [29]) are usually in the order MFP-RND-OMF, unlike H. cinaedi, H. pylori and H. hepaticus [9,30].
Figure 1.
Drug efflux genes encoded in the genome of H. cinaedi PAGU 611. Chromosomal positions of drug efflux genes coding for putative inner membrane efflux transporters (red), outer membrane proteins (green), membrane fusion proteins (orange), and cytoplasmic proteins (light blue) are indicated by the kb (kilobase pair) in the H. cinaedi PAGU 611 genome [22]. Arrows correspond to the lengths and directions of the genes.
Figure 1.
Drug efflux genes encoded in the genome of H. cinaedi PAGU 611. Chromosomal positions of drug efflux genes coding for putative inner membrane efflux transporters (red), outer membrane proteins (green), membrane fusion proteins (orange), and cytoplasmic proteins (light blue) are indicated by the kb (kilobase pair) in the H. cinaedi PAGU 611 genome [22]. Arrows correspond to the lengths and directions of the genes.
5. Structure of the RND Components (HCN_0595 and HCN_1563) of H. cinaedi
The RND components of RND-type tripartite multidrug efflux pumps determine their substrate specificity [31,32]; therefore, we focused on the structure and function of the RND components of H. cinaedi (HCN_0595 (YP_006638872) and HCN_1563 (YP_006235870)), rather than the OMPs or MFPs. BLAST analysis [33] showed that HCN_0595, with a calculated molecular mass of 112 kDa, exhibited strong sequence homology to the uncharacterized RND component of the HH0222 pump (NP_859753, 86% (94%) identity (positive)) of HH0224-HH0223-HH00224 (named HefABC after those of H. pylori [9]) of H. hepaticus ATCC 51449 and significant homology to the HefC (NP_207402, 58% (78%) identity (positive)) RND component of the HefABC pump of H. pylori 26695 [30] (Table 1)and uncharacterized pumps (49%–58% (70%–78%) identity (positive)) of various other Helicobacter species (e.g., H. acinonychis, H. cetorum, H. mustelae, H. bizzozeronii, H. bilis, H. suis, H felis, H. pullorum, H. winghamensis, and H. canadensis) and other ε-proteobacteria (e.g., Wolinella succinogenes). The HefC pump of H. pylori was shown to play a critical role in resistance to bile salts and ceragenins, non-peptide mimics of antimicrobial peptides [30]. This pump might also be involved, to some extent, in antimicrobial resistance, including metronidazole [e.g., 34], although genetic evidence for the HefC pump has not been provided [35]. In C. jejuni subsp. jejuni NCTC 11168, the best-studied organism for efflux systems in ε-proteobacteria, CmeF (YP_002344428), the RND component of the CmeDEF pump [36], but not CmeB (YP_002343803), the RND component of the CmeABC pump, showed significant similarity (38% (59%) identity (positive)) to HCN_0595. The contribution of CmeDEF to intrinsic resistance is likely to be small or secondary compared with that of the major multidrug efflux system CmeABC [36,37]. We could not find any other characterized pumps that were significantly similar to HCN_0595.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Homologues in the other representative ε-proteobacteria for the putative drug efflux transporters of H. cinaedi PAGU 611. Homologues in H. hepaticus ATCC 51449, H. pylori 26695 and C. jejuni NCTC 11168 for the putative efflux transporters of H. cinaedi PAGU 611 are shown using BLAST analysis.
| Putative drug transporter | Family | Homologue (Identity (%) (Positives (%))) | ||
|---|---|---|---|---|
| H. hepaticus | H. pylori | C. jejuni | ||
| ATCC 51449 | 26695 | NCTC 11168 | ||
| HCN_0595 | RND | HH0222 (86 (94)) | HefC (58 (78)) | CmeF (38 (59)) |
| HCN_0708 | MATE | HH0167 (81 (90)) | HP1184 (48 (73) | Cj0560 (29 (49)) |
| HCN_0741 | MF | HH1614 (80 (90)) | HP1181 (47 (64)) | CmeG (43 (64)) |
| HCN_0807 | MATE | HH0031 (76 (87)) | HP0759 (40 (62)) | |
| HCN_0962 | ABC | HH1856 (87 (93)) | Cj0607 (32 (57)) | |
| HCN_1563 | RND | HH0174 (88 (95)) | CmeB (53 (73)) | |
| HCN_1599 | SMR | HH0508 (59 (74)) | Cj1174 (54 (76)) | |
| HCN_1600 | SMR | HH0509 (61 (73)) | Cj1173 (40 (64)) | |
| HCN_2016 | SMR | HH1452 (61 (73)) | Cj0309c (57 (73)) | |
| HCN_2017 | SMR | HH1451 (56 (69)) | Cj0309c (57 (73)) | |
HCN_1563, with a calculated molecular mass of 113 kDa, exhibited strong sequence homology to uncharacterized RND pumps of enterohepatic Helicobacter species, including HH0174 (NP_859705, 88% (95%) identity (positive)) of the HH0175-HH0174 pump (named CmeAB after those of C. jejuni [9]) of H. hepaticus ATCC 51449 and HRAG_01727 (ZP_04580572, 80% (90%) identity (positive)) of H. bilis ATCC 43879, but not of gastric Helicobacter species, such as H. pylori (Table 1). Actually, HCN_1563 exhibited high similarity with major RND multidrug efflux pumps (CmeBs) of Campylobacter species (e.g., CmeB of C. jejuni subsp. jejuni NCTC 11168 (YP_002343803, 53% (73%) identity (positive)) (Table 1). The genome of H. cinaedi was the most similar to that of H. hepaticus [22], which exhibits a unique combination of features mainly from H. pylori and C. jejuni [38]. HCN_1563 might be a pump required to survive in the gut environment, but not the gastric environment. The contribution of the CmeABC efflux pump to acquired resistance of C. jejuni with target mutations to macrolides and fluoroquinolones has been described [10,39,40,41], which is similar to the ciprofloxacin- and clarithromycin-resistant H. cinaedi clinical isolates identified in Japan since 2000 [7,8]. With the exception of ε-proteobacteria, the BepE (NP_697326, 55% (73%) identity (positive)) and BepG pumps (NP_699529, 43% (65%) identity (positive)) of Brucella suis 1330 are taxonomically classified within α-proteobacteria [42], the TtgB pump (YP_006536083, 47% (68%) identity (positive)) of Pseudomonas putida DOT-T1E [43] and the AheB pump (YP_857414, 43% (65%) identity (positive)) of Aeromonas hydrophila subsp. hydrophila ATCC 7966 [44] are noteworthy as very similar pumps. HCN_1563 also exhibited significant similarities, as judged from phylogenetic distance, with well-studied multidrug efflux pumps (AcrB/AcrD/AcrF pumps (41% (61%–64%) identity (positive)) of E. coli and MexB/MexF/MexD/MexY pumps (39%–43% (61%–65%)) of P. aeruginosa [13,14].
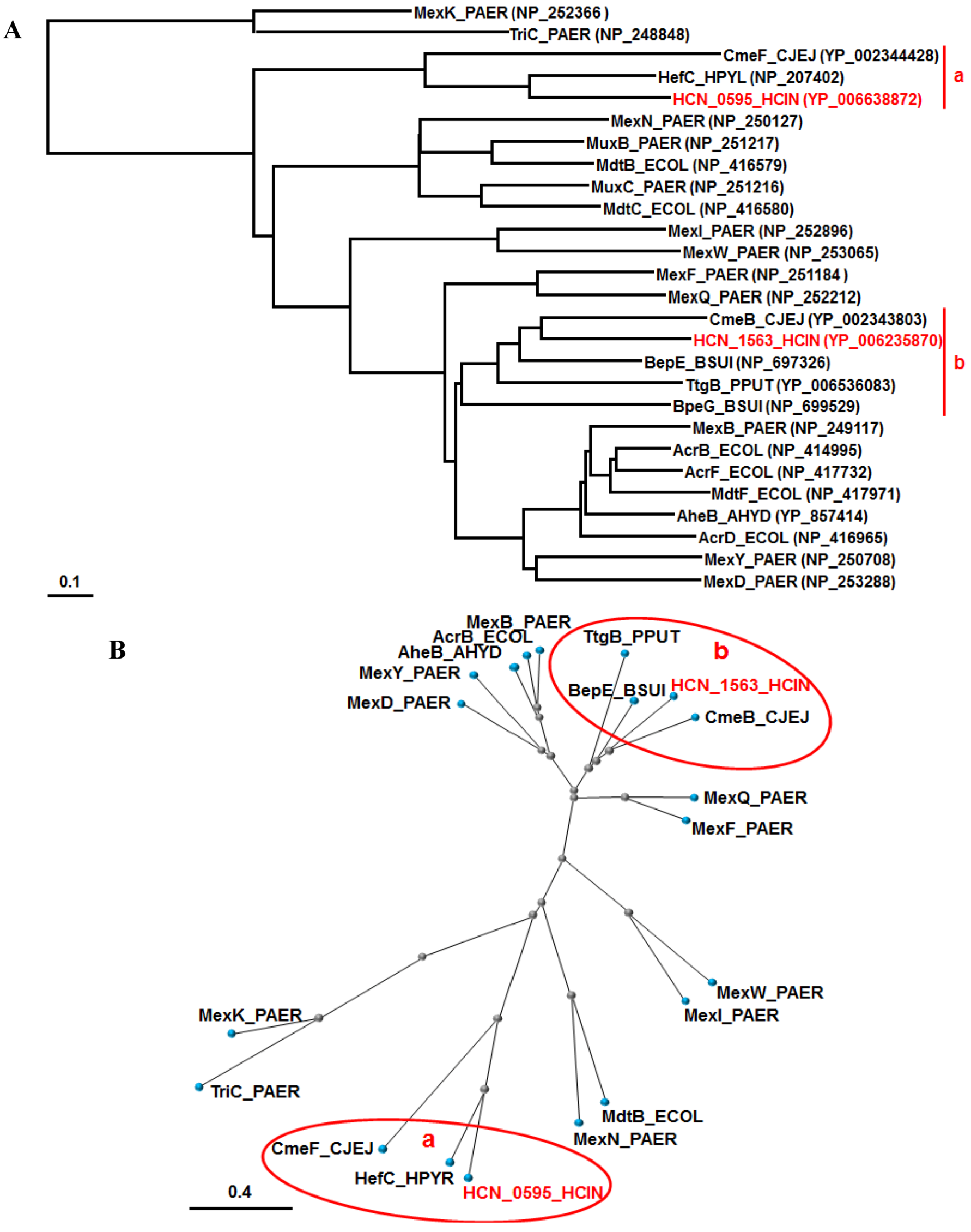
COBALT analysis [45] of representative RND pumps in Gram-negative bacteria, including all RND pumps from P. aeruginosa PAO1 and E. coli K12, characterized their relationships, and we focused on two branches containing the two RND pumps of H. cinaedi (Figure 2). The branch belonging to the HCN_0595 pump only includes HefC of H. pylori and CmeF of C. jejuni, while the branch containing the HCN_1563 pump includes not only CmeB of C. jejuni, but also the BepE/G pumps of B. suis and TtgB of P. putida (Figure 2).
Taken together, we assume that the HCN_0595 pump of H. cinaedi plays a similar role to HefC of H. pylori and CmeF of C. jejuni, while the HCN_1563 pump has a similar role as CmeB of C. jejuni. In addition, the two pumps of H. cinaedi must play very similar roles to those of H. hepaticus, which is not surprising, according to their biological and genomic similarities. Recently, HefA (HH0224), the OMF component of HefABC of H. hepaticus ATCC 51449, was shown to be involved in resistance to amoxicillin and some antimicrobials, as well as bile acids [9]. As the authors failed to isolate a mutant RND pump (HH0222 (HCN_0595 orthologue) and HH0174 (HCN_1563 orthologue) in Figure 2), we do not know if the resistance to amoxicillin and bile acids is caused by HH0222 or HH0174, because the HH0174 gene is a two-gene operon, like the HCN_1563 gene [9]. It is noteworthy that the HefC pump of H. pylori played a role in cholesterol-dependent resistance in the bile salt-rich enterohepatic environment [30]. Cholesterol enhanced H. pylori resistance to various antibiotics, such as clarithromycin, amoxicillin and ciprofloxacin, as well as bile salts (e.g., deoxycholate) [30,46]. It is intriguing to determine if H. cinaedi resistance is enhanced by cholesterol and if the RND pumps of H. cinaedi play a role in cholesterol-dependent resistance. Actually, hefABC of H. hepaticus and H. pylori and cmeABC of C. jejuni were inducible by bile acids [9,30,47]. It is of note that CmeABC of C. jejuni plays a critical role in colonization in vivo [48].
Figure 2.
Phylogenetic trees for RND pumps of various bacteria. According to the COBALT program, the trees were constructed using the Fast evolution method and rendered with (A) Rectangle and (B) Radical. The accession numbers are shown in parentheses. The branches belonging to HCN_0595 and HCN_1563 of H. cinaedi PAGU 611 are shown in red and named “a” and “b”, respectively. The proteins are abbreviated (e.g., “AcrB_ECOL” stands for “AcrB of E. coli”). Abbreviations; PAER, Pseudomonas aeruginosa; CJEJ, Campylobacter jejuni; HPYR, Helicobacter pyroli; HCIN, Helicobacter cinaedi; ECOL, Escherichia coli; BSUI, Brucella suis; PPUT, Pseudomonas putida; AHYD, Aeromonas hydrophila.
Figure 2.
Phylogenetic trees for RND pumps of various bacteria. According to the COBALT program, the trees were constructed using the Fast evolution method and rendered with (A) Rectangle and (B) Radical. The accession numbers are shown in parentheses. The branches belonging to HCN_0595 and HCN_1563 of H. cinaedi PAGU 611 are shown in red and named “a” and “b”, respectively. The proteins are abbreviated (e.g., “AcrB_ECOL” stands for “AcrB of E. coli”). Abbreviations; PAER, Pseudomonas aeruginosa; CJEJ, Campylobacter jejuni; HPYR, Helicobacter pyroli; HCIN, Helicobacter cinaedi; ECOL, Escherichia coli; BSUI, Brucella suis; PPUT, Pseudomonas putida; AHYD, Aeromonas hydrophila.

6. A Possible Regulator Gene of Multidrug Efflux Systems in H. cinaedi
Although cognate regulators (e.g., repressors, activators, or two-component systems) located upstream of the RND efflux genes often exist, no cognate regulator was found upstream or downstream of the RND efflux operons of H. cinaedi, H. hepaticus and H. pylori. In C. jejuni, cmeR, which is a transcriptional repressor located immediately upstream of the cmeABC operon, encodes a 210 amino-acid protein that shares sequence and structural similarities with the members of the TetR family of transcriptional repressors [49]. BLAST analysis did not identify a homologue of CmeR in the genomes of Helicobacter species. Actually, H. cinaedi possesses only a small set of genes encoding transcriptional regulators, very similar to H. hepaticus [22,38].
Very recently, CosR, an oxidative stress responsive global regulator essential for viability [50], was shown to regulate the cmeABC operon negatively by binding directly upstream of cmeABC in C. jejuni NCTC 11168 [51]. CosR homologues are found mostly in ε-proteobacteria [51]. BLAST analysis showed that a quite similar CosR homologue (HCN_1079, YP_006235418.1) exists in H. cinaedi PAGU 611 (74% (86%) identity (positive)) and the strains ATCC BAA-847 and CCUG 18818. This homologue might also be involved in the expression of an efflux gene in H. cinaedi. In Gram-negative bacteria, oxidative stress responses are linked to the development of antimicrobial resistance, resulting from the activation of a resistance mechanism in which the RND multidrug efflux system is an important component [52]. For example, exposure to reactive oxygen species, such as peroxide, leads to MexXY-dependent aminoglycoside resistance in P. aeruginosa [52,53]. We point out that the putative start codon of all CosR homologues (HCN_1079, HCBAA847_0895, and HCCG_01220) of the H. cinaedi strains is TTG, which is a minor start codon [54], and found that an ATG codon located 3 codons before this TTG is also a possible start codon that is preceded by ribosome binding site-like sequences [55].
7. C. jejuni CmeG Homologue Identified in H. cinaedi
CmeG homologues (e.g., HCN_0741 (YP_006235115) of PAGU 611) found in the three H. cinaedi strains showed significant homology (43% (64%) identity (positive)) (Figure 1, Table 1). BLAST analysis showed that HH1614 of H. hepaticus ATCC 51449 is a strong homologue (80% (90%) identity (positive)) (Table 1). CmeG was shown to function as a multidrug efflux transporter of the MF family that contributes to antimicrobial resistance and oxidative defense (hydrogen peroxide) in C. jejuni [56]. Mutations of cmeG significantly reduced resistance to various classes of antimicrobials, including ciprofloxacin, tetracycline, gentamicin, ethidium bromide and cholic acid, and overexpression of cmeG in the wild-type background increased resistance to fluoroquinolones [56]. CmeG shows significant homology to well-known MF-type multidrug efflux transporters of Gram-positive bacteria, such as NorA of Staphylococcus aureus (27% identity) and Bmr of Bacillus subtilis (27% identity) [56].
8. Other Probable Drug Efflux Systems in H. cinaedi
Finally, we discuss other probable drug efflux systems found in H. cinaedi, although the clinical significance and natural function of their homologues in other characterized bacteria remain unknown.
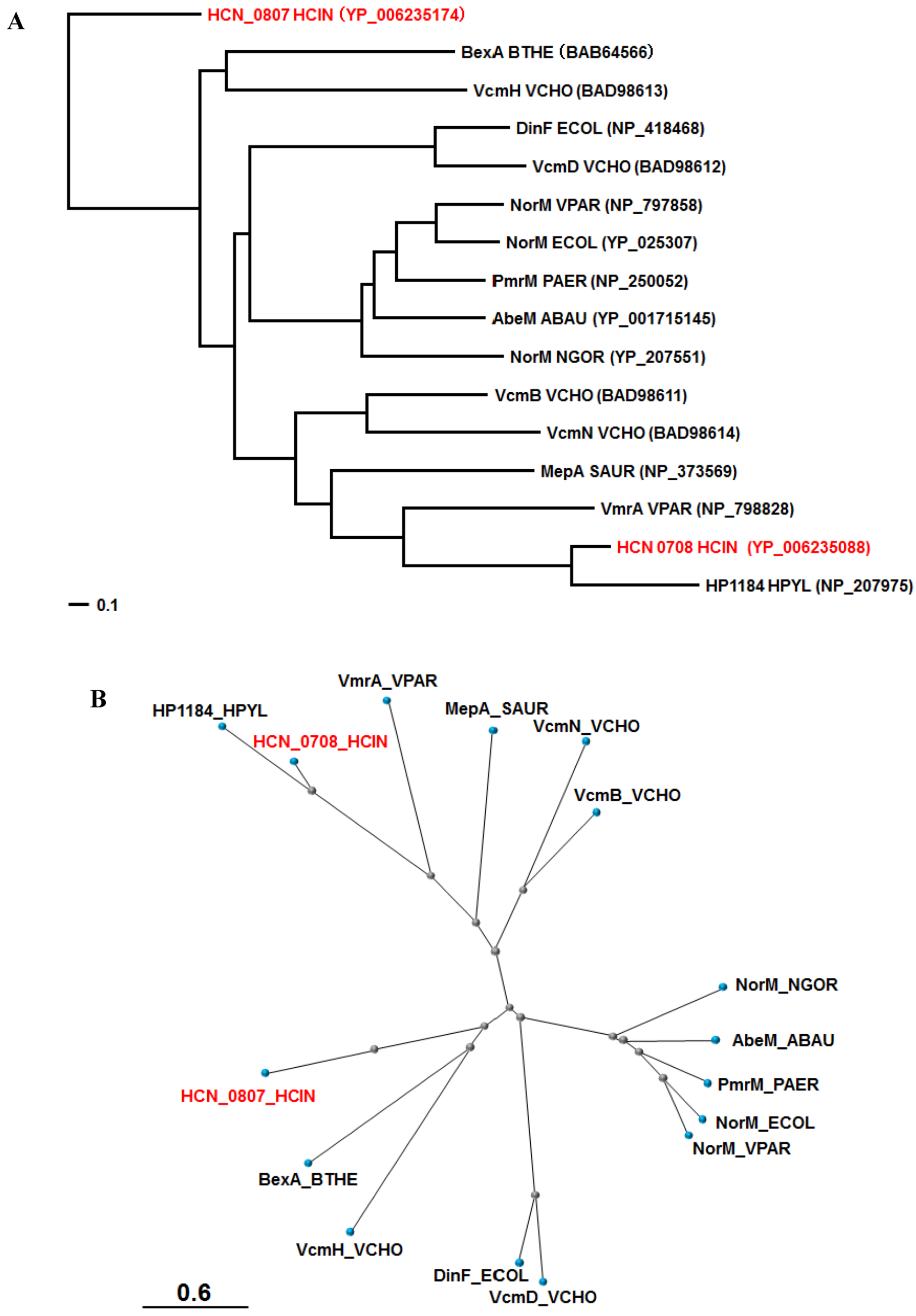
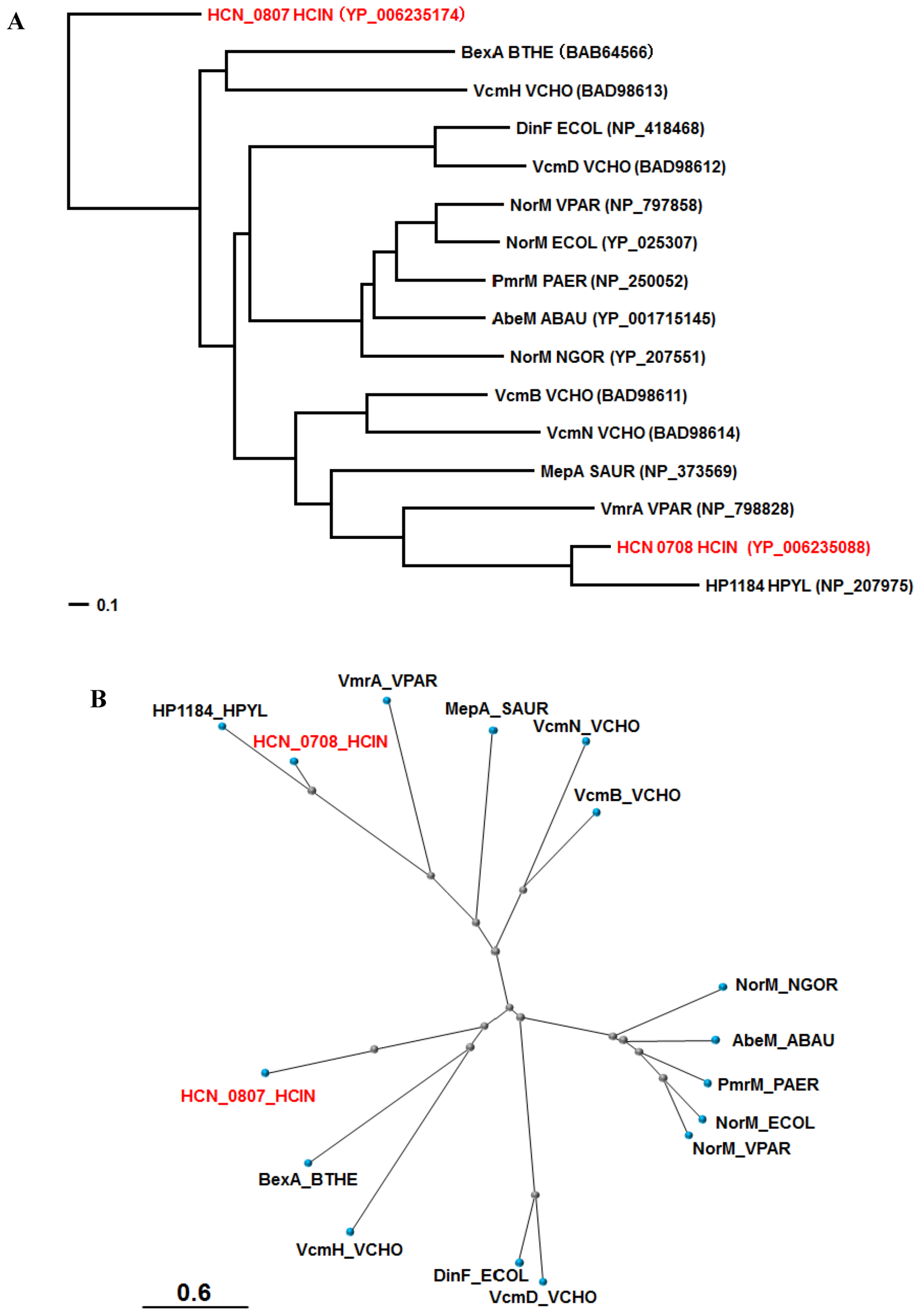
Two MATE family multidrug efflux family transporters (HCN_0708 and HCN_0807) were found in H. cinaedi PAGU 611 and the other two H. cinaedi strains (Figure 1). BLAST analysis showed strong homologues of HCN_0708 and HCN_0807 are HH0167 (NP_859698) and HH0031 (NP_859562) of H. hepaticus ATCC 51449 (81% (90%) and 76% (87%) identity (positive)), respectively (Table 1). COBALT analysis with MATE pumps characterized in other bacteria showed that HCN_0708 was close to HP1184 of H. pylori [57], followed by VmrA of Vibrio parahaemolyticus [58], while HCN_0807 seemed unique, but comparably close to BexA of Bacteroides thetaiotaomicron [59] and VcmH of Vibrio cholerae [60] (Figure 3). Both BexA and VcmH gave resistance to hydrophilic quinolones (e.g., norfloxacin and ciprofloxacin) when expressed in an E. coli mutant lacking an acrB gene encoding the major RND multidrug efflux pump [59,60].
Figure 3.
Phylogenetic trees for the MATE pumps of various bacteria. According to the COBALT program, the trees were constructed using the Fast evolution method and rendered with (A) Rectangle and (B) Radical. The proteins are abbreviated (e.g., “NorM_ECOR” stands for “NorM of E. coli”). The accession numbers are shown in parentheses. HCN_0708 and HCN_0807 of H cinaedi PAGU 611 are shown in red. Abbreviations; HCIN, Helicobacter cinaedi; BTHE, Bacteroides thetaiotaomicron ; VCHO, Vibrio cholera; ECOL, Escherichia coli; VPAR, Vibrio parahaemolyticus; PAER, Pseudomonas aeruginosa; ABAU, Acinetobacter baumannii; NGOR, Neisseria gonorrhoeae; SAUR, Staphylococcus aureus; HPYR, Helicobacter pyroli.
Figure 3.
Phylogenetic trees for the MATE pumps of various bacteria. According to the COBALT program, the trees were constructed using the Fast evolution method and rendered with (A) Rectangle and (B) Radical. The proteins are abbreviated (e.g., “NorM_ECOR” stands for “NorM of E. coli”). The accession numbers are shown in parentheses. HCN_0708 and HCN_0807 of H cinaedi PAGU 611 are shown in red. Abbreviations; HCIN, Helicobacter cinaedi; BTHE, Bacteroides thetaiotaomicron ; VCHO, Vibrio cholera; ECOL, Escherichia coli; VPAR, Vibrio parahaemolyticus; PAER, Pseudomonas aeruginosa; ABAU, Acinetobacter baumannii; NGOR, Neisseria gonorrhoeae; SAUR, Staphylococcus aureus; HPYR, Helicobacter pyroli.
Two putative SMR family efflux systems (HCN_2017-HCN_2016 and HCN_1600-HCN_1599), both of which encode two SMR components, were found in H. cinaedi PAGU 611 and the other two H. cinaedi strains (Figure 1). BLAST analysis showed that strong homologues of HCN_2017-HCN_2016 and HCN_1600-HCN_1599 are HRAG_00571-HRAG_00572 (ZP_04582237-ZP_04582238) of H. bilis ATCC 43879 (94% (96%) and 98% (99%) identity (positive)) and HH509-HH508 (NP_860040-NP_860039) of H. hepaticus ATCC 51449 (59% (74%) and 67% (86%) identity (positive)), respectively (Table 1). Of note, a strong homologue of HCN_2017-HCN_2016 was HH1451-HH1452 (NP_860982-NP_860983) of H. hepaticus ATCC 51449 (56% (69%) and 61% (73%) identity (positive)) (Table 1). Both components appear to be necessary for pump activity, e.g., EbrAB and YkkCD of Bacillus subtilis [61,62]. BLAST analysis with E. coli K12, P. aeruginosa PAO1, B. subtilis 168 and Staphylococcus aureus N315 suggested that HCN_2017-HCN_2016 showed significant similarity to YkkCD (NP_389192 and NP_389193; 43% (60%) and 48% (68%) identity (positive), respectively) of B. subtilis, while HCN_1599-HCN_1600 showed significant similarity to MdtJI (NP_416117 and NP_416116; 38% (57%) and 38% (63%) identity (positive), respectively) of E. coli K12. YkkCD is a multidrug efflux pump that gives rise to broad specificity, including to cationic (e.g., streptomycin, tetracycline and ethidium bromide), neutral (e.g., chloramphenicol), and anionic compounds (e.g., phosphonomycin), when expressed in E. coli [62]. In addition, MdtJI overexpression conferred resistance to deoxycholate when expressed in an E. coli mutant lacking acrB, a major RND multidrug efflux pump [63], and rescued cell toxicity and growth inhibition due to the over-accumulation of spermidine in a spermidine acetyltransferase-deficient E. coli mutant [64].
One ABC family efflux system was found in H. cinaedi PAGU 611 (Figure 1). It consists of four genes (HCN_0962-HCN_0963-HCN_0964-HCN_0965) encoding an inner membrane transporter, ATP binding protein, MFP and OMF, respectively, which means that it is an ABC transporter that spans the entirety of the Gram-negative cell envelope. The same efflux system was observed in the two other H. cinaedi strains. BLAST analysis showed that HH1856 (NP_861387) of H. hepaticus ATCC 51449 was a strong homologue of HCN_0962 (87% (93%) identity (positive)) (Table 1). BLAST analysis with E. coli K12 and P. aeruginosa PAO1 suggested that HCN_0962 was significantly similar to the inner membrane domains of both MacB (NP_415400; 34% (56%) identity (positive)) of E. coli K12 and PvdT (33% (54%)) of P. aeruginosa PAO1. MacB and PvdT are inner membrane components of the macrolide-specific ABC transporter MacAB of E. coli [19] and of the de novo synthesized pyoverdine secretion system PvdRT-OpmQ of P. aeruginosa, respectively [65].
9. Future Perspective
The genome of H. cinaedi possesses probable uncharacterized drug efflux systems consisting of two RND pumps, one MF pump, two MATE pumps, two SMR pumps and one ABC pump, all of which are very similar to those of H. hepaticus. Because multidrug efflux pumps have roles in not only bacterial drug resistance, but also in other systems, including virulence and the stress response [52,63], characterizing the multidrug efflux pumps of H. cinaedi should lead to the understanding of various physiological aspects of this organism and, ultimately, conquering H. cinaedi infections. To do so, it is necessary to develop genetic tools and improve the culture method for this organism, while we can also use multiplex technologies, such as real-time PCR, DNA microarrays, proteomics and metagenomics. In the meantime, each pump can be cloned and characterized in organisms that lack a homologue, such as E. coli, C. jejuni and H. pylori, but some uncertainties will remain. Interestingly, H. cinaedi PAGU 611, but not ATCC BAA-847, possesses one plasmid, pHci1 (~23 kbp, 29 predicted coding sequences, of which 27 are hypothetical proteins) [22,23]. As such, it may represent a diamond in the rough that can be developed into a stable shuttle vector, although no replication protein or origin of replication have yet been found in this plasmid.
Acknowledgments
This work was supported in part by a Grant-in-Aid for Young Scientists (B) (KAKENHI 23790106) from the Japan Society for the Promotion of Science and a research grant from the Institute of Pharmaceutical Life Sciences, Aichi Gakuin University.
Conflict of Interest
The authors declare no conflict of interest.
References
- Lawson, A.J. Helicobacter. In Manual of Clinical Microbiology, 10th; Versalovic, J., Carroll, K.C., Funke, G., Jorgensen, J.H., Landry, M.L., Warnock, D.W., Eds.; ASM Press: Washington D.C., USA, 2011; Volume 1, pp. 900–915. [Google Scholar]
- Uckay, I.; Garbino, J.; Dietrich, P.Y.; Ninet, B.; Rohner, P.; Jacomo, V. Recurrent Bacteremia with Helicobacter cinaedi: Case Report and Review of the Literature. BMC Infect. Dis. 2006, 6, e86. [Google Scholar] [CrossRef]
- Kitamura, T.; Kawamura, Y.; Ohkusu, K.; Masaki, T.; Iwashita, H.; Sawa, T.; Fujii, S.; Okamoto, T.; Akaike, T. Helicobacter cinaedi Cellulitis and Bacteremia in Immunocompetent Hosts after Orthopedic Surgery. J. Clin. Microbiol. 2007, 45, 31–38. [Google Scholar] [CrossRef]
- Khan, S.; Okamoto, T.; Enomoto, K.; Sakashita, N.; Oyama, K.; Fujii, S.; Sawa, T.; Takeya, M.; Ogawa, H.; Yamabe, H.; et al. Potential Association of Helicobacter cinaedi with Atrial Arrhythmias and Atherosclerosis. Microbiol. Immunol. 2012, 56, 145–154. [Google Scholar]
- Tomida, J.; Kashida, M.; Oinishi, K.; Endo, R.; Morita, Y.; Kawamura, Y. Evaluation of Various Media for Rapid Detection of Helicobacter spp. J. Jpn. Soc. Clin. Microbiol. 2008, 18, 227–235. [Google Scholar]
- Oyama, K.; Khan, S.; Okamoto, T.; Fujii, S.; Ono, K.; Matsunaga, T.; Yoshitake, J.; Sawa, T.; Tomida, J.; Kawamura, Y.; et al. Identification of and Screening for Human Helicobacter cinaedi Infections and Carriers via Nested PCR. J. Clin. Microbiol. 2012, 50, 3893–3900. [Google Scholar] [CrossRef]
- Rimbara, E.; Mori, S.; Matsui, M.; Suzuki, S.; Wachino, J.; Kawamura, Y.; Shen, Z.; Fox, J.G.; Shibayama, K. Molecular Epidemiologic Analysis and Antimicrobial Resistance of Helicobacter cinaedi Isolated from Seven Hospitals in Japan. J. Clin. Microbiol. 2012, 50, 2553–2560. [Google Scholar]
- Tomida, J.; Morita, Y.; Kawamura, Y. Antimicrobial Susceptibility Tests and Resistant Mechanisms of Helicobacter cinaedi. Jan. J. Bacteriol. 2012, 67, 127. [Google Scholar]
- Belzer, C.; Stoof, J.; Breijer, S.; Kusters, J.G.; Kuipers, E.J.; van Vliet, A.H. The Helicobacter hepaticus hefA Gene is Involved in Resistance to Amoxicillin. Helicobacter 2009, 14, 72–79. [Google Scholar] [CrossRef]
- Gibreel, A.; Wetsch, N.M.; Taylor, D.E. Contribution of the CmeABC Efflux Pump to Macrolide and Tetracycline Resistance in Campylobacter jejuni. Antimicrob. Agents Chemother. 2007, 51, 3212–3216. [Google Scholar] [CrossRef]
- Morita, Y.; Tomida, J.; Kawamura, Y. Primary Mechanisms Mediating Aminoglycoside Resistance in the Multidrug-resistant Pseudomonas aeruginosa Clinical Isolate PA7. Microbiology 2012, 158, 1071–1083. [Google Scholar] [CrossRef]
- Nikaido, H. Prevention of Drug Access to Bacterial Targets: Permeability Barriers and Active Rfflux. Science 1994, 264, 382–388. [Google Scholar]
- Li, X.Z.; Nikaido, H. Efflux-mediated Drug Resistance in Bacteria: An Update. Drugs 2009, 69, 1555–1623. [Google Scholar]
- Poole, K. Efflux-mediated Antimicrobial Resistance. In Antibiotic Discovery and Development; Dougherty, T.J., Pucci, M.J., Eds.; Springer: New York, NY, USA, 2012; Volume 1, pp. 349–395. [Google Scholar]
- Zgurskaya, H.I.; Nikaido, H. Bypassing the Periplasm: Reconstitution of the AcrAB Multidrug Efflux Pump of Escherichia coli. Proc. Natl. Acad. Sci. USA 1999, 96, 7190–7195. [Google Scholar] [CrossRef]
- Mine, T.; Morita, Y.; Kataoka, A.; Mizushima, T.; Tsuchiya, T. Evidence for Chloramphenicol/H+ Antiport in Cmr (MdfA) System of Escherichia coli and Properties of the Antiporter. J. Biochem. 1998, 124, 187–193. [Google Scholar] [CrossRef]
- Yerushalmi, H.; Lebendiker, M.; Schuldiner, S. EmrE, an Escherichia coli 12-kDa Multidrug Transporter, Exchanges Toxic Cations and H+ and is Soluble in Organic Solvents. J. Biol. Chem. 1995, 270, 6856–6863. [Google Scholar]
- Morita, Y.; Kataoka, A.; Shiota, S.; Mizushima, T.; Tsuchiya, T. NorM of Vibrio parahaemolyticus is an Na(+)-driven Multidrug Efflux Pump. J. Bacteriol. 2000, 182, 6694–6697. [Google Scholar] [CrossRef]
- Kobayashi, N.; Nishino, K.; Yamaguchi, A. Novel Macrolide-specific ABC-type Efflux Transporter in Escherichia coli. J. Bacteriol. 2001, 183, 5639–5644. [Google Scholar]
- Huda, N.; Lee, E.W.; Chen, J.; Morita, Y.; Kuroda, T.; Mizushima, T.; Tsuchiya, T. Molecular Cloning and Characterization of an ABC Multidrug Efflux Pump, VcaM, in Non-O1 Vibrio cholerae. Antimicrob. Agents Chemother. 2003, 47, 2413–2417. [Google Scholar] [CrossRef]
- Mine, T.; Morita, Y.; Kataoka, A.; Mizushima, T.; Tsuchiya, T. Expression in Escherichia coli of a New Multidrug Efflux Pump, MexXY, from Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1999, 43, 415–417. [Google Scholar]
- Goto, T.; Ogura, Y.; Hirakawa, H.; Tomida, J.; Morita, Y.; Akaike, T.; Hayashi, T.; Kawamura, Y. Complete Genome Sequence of Helicobacter cinaedi Strain PAGU 611, Isolated in a Case of Human Bacteremia. J. Bacteriol. 2012, 194, 3744–3745. [Google Scholar] [CrossRef]
- Miyoshi-Akiyama, T.; Takeshita, N.; Ohmagari, N.; Kirikae, T. Complete Genome Sequence of Helicobacter cinaedi Type Strain ATCC BAA-847. J. Bacteriol. 2012, 194, 5692. [Google Scholar] [CrossRef]
- The Human Microbiome Project Consortium. A Framework for Human Microbiome Research. Nature 2012, 486, 215–221.
- Charoenlap, N.; Shen, Z.; McBee, M.E.; Muthupalani, S.; Wogan, G.N.; Fox, J.G.; Schauer, D.B. Alkyl Hydroperoxide Reductase is Required for Helicobacter cinaedi Intestinal Colonization and Survival under Oxidative Stress in BALB/c and BALB/c Interleukin-10−/− mice. Infect. Immun. 2012, 80, 921–928. [Google Scholar] [CrossRef]
- Tseng, T.T.; Gratwick, K.S.; Kollman, J.; Park, D.; Nies, D.H.; Goffeau, A.; Saier, M.H., Jr. The RND Permease Superfamily: An Ancient, Ubiquitous and Diverse Family that Includes Human Disease and Development Proteins. J. Mol. Microbiol. Biotechnol. 1999, 1, 107–125. [Google Scholar]
- Morita, Y.; Kimura, N.; Mima, T.; Mizushima, T.; Tsuchiya, T. Roles of MexXY- and MexAB-Multidrug Efflux Pumps in Intrinsic Multidrug Resistance of Pseudomonas aeruginosa PAO1. J. Gen. Appl. Microbiol. 2001, 47, 27–32. [Google Scholar] [CrossRef]
- Poole, K.; Krebes, K.; McNally, C.; Neshat, S. Multiple Antibiotic Resistance in Pseudomonas aeruginosa: Evidence for Involvement of an Efflux Operon. J. Bacteriol. 1993, 175, 7363–7372. [Google Scholar]
- Lin, J.; Michel, L.O.; Zhang, Q. CmeABC Functions as a Multidrug Efflux System in Campylobacter jejuni. Antimicrob. Agents Chemother. 2002, 46, 2124–2131. [Google Scholar]
- Trainor, E.A.; Horton, K.E.; Savage, P.B.; Testerman, T.L.; McGee, D.J. Role of the HefC Efflux Pump in Helicobacter pylori Cholesterol-dependent Resistance to Ceragenins and Bile Salts. Infect. Immun. 2011, 79, 88–97. [Google Scholar] [CrossRef]
- Srikumar, R.; Li, X.Z.; Poole, K. Inner Membrane Efflux Components are Responsible for Beta-Lactam Specificity of Multidrug Efflux Pumps in Pseudomonas aeruginosa. J. Bacteriol. 1997, 179, 7875–7881. [Google Scholar]
- Nakashima, R.; Sakurai, K.; Yamasaki, S.; Nishino, K.; Yamaguchi, A. Structures of the Multidrug Exporter AcrB Reveal a Proximal Multisite Drug-binding Pocket. Nature 2011, 480, 565–569. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar]
- Tsugawa, H.; Suzuki, H.; Muraoka, H.; Ikeda, F.; Hirata, K.; Matsuzaki, J.; Saito, Y.; Hibi, T. Enhanced bacterial efflux system is the first step to the development of metronidazole resistance in Helicobacter pylori. Biochem. Biophys. Res. Commun. 2011, 404, 656–660. [Google Scholar] [CrossRef]
- Francesco, V.D.; Zullo, A.; Hassan, C.; Giorgio, F.; Rosania, R.; Ierardi, E. Mechanisms of Helicobacter pylori Antibiotic Resistance: An Updated Appraisal. World J. Gastrointest. Pathophysiol. 2011, 2, 35–41. [Google Scholar] [CrossRef]
- Pumbwe, L.; Randall, L.P.; Woodward, M.J.; Piddock, L.J. Evidence for Multiple-antibiotic Resistance in Campylobacter jejuni not Mediated by CmeB or CmeF. Antimicrob. Agents Chemother. 2005, 49, 1289–1293. [Google Scholar] [CrossRef]
- Akiba, M.; Lin, J.; Barton, Y.W.; Zhang, Q. Interaction of CmeABC and CmeDEF in Conferring Antimicrobial Resistance and Maintaining Cell Viability in Campylobacter jejuni. J. Antimicrob. Chemother. 2006, 57, 52–60. [Google Scholar]
- Suerbaum, S.; Josenhans, C.; Sterzenbach, T.; Drescher, B.; Brandt, P.; Bell, M.; Droge, M.; Fartmann, B.; Fischer, H.P.; Ge, Z.; et al. The Complete Genome Sequence of the Carcinogenic Bacterium Helicobacter hepaticus. Proc. Natl. Acad. Sci. USA 2003, 100, 7901–7906. [Google Scholar]
- Cagliero, C.; Mouline, C.; Payot, S.; Cloeckaert, A. Involvement of the CmeABC Efflux Pump in the Macrolide Resistance of Campylobacter coli. J. Antimicrob. Chemother. 2005, 56, 948–950. [Google Scholar] [CrossRef]
- Yan, M.; Sahin, O.; Lin, J.; Zhang, Q. Role of the CmeABC Efflux Pump in the Emergence of Fluoroquinolone-resistant Campylobacter under Selection Pressure. J. Antimicrob. Chemother. 2006, 58, 1154–1159. [Google Scholar] [CrossRef]
- Lin, J.; Yan, M.; Sahin, O.; Pereira, S.; Chang, Y.J.; Zhang, Q. Effect of Macrolide Usage on Emergence of Erythromycin-resistant Campylobacter Isolates in Chickens. Antimicrob. Agents Chemother. 2007, 51, 1678–1686. [Google Scholar]
- Martin, F.A.; Posadas, D.M.; Carrica, M.C.; Cravero, S.L.; O'Callaghan, D.; Zorreguieta, A. Interplay Between Two RND Systems Mediating Antimicrobial Resistance in Brucella suis. J. Bacteriol. 2009, 191, 2530–2540. [Google Scholar] [CrossRef]
- Teran, W.; Felipe, A.; Segura, A.; Rojas, A.; Ramos, J.L.; Gallegos, M.T. Antibiotic-dependent Induction of Pseudomonas putida DOT-T1E TtgABC Efflux Pump is Mediated by the Drug Binding Repressor TtgR. Antimicrob. Agents Chemother. 2003, 47, 3067–3072. [Google Scholar]
- Hernould, M.; Gagne, S.; Fournier, M.; Quentin, C.; Arpin, C. Role of the AheABC Efflux Pump in Aeromonas hydrophila Intrinsic Multidrug Resistance. Antimicrob. Agents Chemother. 2008, 52, 1559–1563. [Google Scholar] [CrossRef]
- Papadopoulos, J.S.; Agarwala, R. COBALT: Constraint-based Alignment Tool for Multiple Protein Sequences. Bioinformatics 2007, 23, 1073–1079. [Google Scholar] [CrossRef]
- McGee, D.J.; George, A.E.; Trainor, E.A.; Horton, K.E.; Hildebrandt, E.; Testerman, T.L. Cholesterol Enhances Helicobacter pylori Resistance to Antibiotics and ll-37. Antimicrob. Agents Chemother. 2011, 55, 2897–2904. [Google Scholar] [CrossRef]
- Lin, J.; Cagliero, C.; Guo, B.; Barton, Y.W.; Maurel, M.C.; Payot, S.; Zhang, Q. Bile Salts Modulate Expression of the CmeABC Multidrug Efflux Pump in Campylobacter jejuni. J. Bacteriol. 2005, 187, 7417–7424. [Google Scholar] [CrossRef]
- Lin, J.; Martinez, A. Effect of Efflux Pump Inhibitors on Bile Resistance and in Vivo Colonization of Campylobacter jejuni. J. Antimicrob. Chemother. 2006, 58, 966–972. [Google Scholar] [CrossRef]
- Lin, J.; Akiba, M.; Sahin, O.; Zhang, Q. CmeR Functions as a Transcriptional Repressor for the Multidrug Efflux Pump CmeABC in Campylobacter jejuni. Antimicrob. Agents Chemother. 2005, 49, 1067–1075. [Google Scholar] [CrossRef]
- Hwang, S.; Kim, M.; Ryu, S.; Jeon, B. Regulation of Oxidative Stress Response by CosR, an Essential Response Regulator in Campylobacter jejuni. PLoS One 2011, 6, e22300. [Google Scholar]
- Hwang, S.; Zhang, Q.; Ryu, S.; Jeon, B. Transcriptional Regulation of the CmeABC Multidrug Efflux Pump and the KatA Catalase by CosR in Campylobacter jejuni. J. Bacteriol. 2012, in press. [Google Scholar]
- Poole, K. Stress Responses as Determinants of Antimicrobial Resistance in Gram-negative Bacteria. Trends Microbiol. 2012, 20, 227–234. [Google Scholar] [CrossRef]
- Morita, Y.; Tomida, J.; Kawamura, Y. MexXY Multidrug Efflux System of Pseudomonas aeruginosa. Front. Microbiol. 2012, 3, e408. [Google Scholar]
- Hannenhalli, S.S.; Hayes, W.S.; Hatzigeorgiou, A.G.; Fickett, J.W. Bacterial Start Site Prediction. Nucleic Acids Res. 1999, 27, 3577–3582. [Google Scholar]
- Shine, J.; Dalgarno, L. The 3'-terminal Sequence of Escherichia coli 16S Ribosomal RNA: Complementarity to Nonsense Triplets and Ribosome Binding Sites. Proc. Natl. Acad. Sci. USA 1974, 71, 1342–1346. [Google Scholar] [CrossRef]
- Jeon, B.; Wang, Y.; Hao, H.; Barton, Y.W.; Zhang, Q. Contribution of CmeG to Antibiotic and Oxidative Stress Resistance in Campylobacter jejuni. J. Antimicrob. Chemother. 2011, 66, 79–85. [Google Scholar] [CrossRef]
- Van Amsterdam, K.; Bart, A.; van der Ende, A. A Helicobacter pylori TolC Efflux Pump Confers Resistance to Metronidazole. Antimicrob. Agents Chemother. 2005, 49, 1477–1482. [Google Scholar] [CrossRef]
- Chen, J.; Morita, Y.; Huda, M.N.; Kuroda, T.; Mizushima, T.; Tsuchiya, T. VmrA, a Member of a Novel Class of Na(+)-coupled Multidrug Efflux Pumps from Vibrio parahaemolyticus. J. Bacteriol. 2002, 184, 572–576. [Google Scholar] [CrossRef]
- Miyamae, S.; Ueda, O.; Yoshimura, F.; Hwang, J.; Tanaka, Y.; Nikaido, H. A MATE Family Multidrug Efflux Transporter Pumps out Fluoroquinolones in Bacteroides thetaiotaomicron. Antimicrob. Agents Chemother. 2001, 45, 3341–3346. [Google Scholar] [CrossRef]
- Begum, A.; Rahman, M.M.; Ogawa, W.; Mizushima, T.; Kuroda, T.; Tsuchiya, T. Gene Cloning and Characterization of Four MATE Family Multidrug Efflux Pumps from Vibrio cholerae Non-O1. Microbiol. Immunol. 2005, 49, 949–957. [Google Scholar]
- Masaoka, Y.; Ueno, Y.; Morita, Y.; Kuroda, T.; Mizushima, T.; Tsuchiya, T. A Two-component Multidrug Efflux Pump, EbrAB, in Bacillus subtilis. J. Bacteriol. 2000, 182, 2307–2310. [Google Scholar] [CrossRef]
- Jack, D.L.; Storms, M.L.; Tchieu, J.H.; Paulsen, I.T.; Saier, M.H., Jr. A Broad-specificity Multidrug Efflux Pump Requiring a Pair of Homologous SMR-type Proteins. J. Bacteriol. 2000, 182, 2311–2313. [Google Scholar] [CrossRef]
- Nishino, K.; Nikaido, E.; Yamaguchi, A. Regulation and Physiological Function of Multidrug Efflux Pumps in Escherichia coli and Salmonella. Biochim. Biophys. Acta 2009, 1794, 834–843. [Google Scholar] [CrossRef]
- Higashi, K.; Ishigure, H.; Demizu, R.; Uemura, T.; Nishino, K.; Yamaguchi, A.; Kashiwagi, K.; Igarashi, K. Identification of a Spermidine Excretion Protein Complex (MdtJI) in Escherichia coli. J. Bacteriol. 2008, 190, 872–878. [Google Scholar] [CrossRef]
- Imperi, F.; Tiburzi, F.; Visca, P. Molecular basis of Pyoverdine Siderophore Recycling in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2009, 106, 20440–20445. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Morita, Y.; Tomida, J.; Kawamura, Y. Multidrug Efflux Systems in Helicobacter cinaedi. Antibiotics 2012, 1, 29-43. https://doi.org/10.3390/antibiotics1010029
AMA Style
Morita Y, Tomida J, Kawamura Y. Multidrug Efflux Systems in Helicobacter cinaedi. Antibiotics. 2012; 1(1):29-43. https://doi.org/10.3390/antibiotics1010029
Chicago/Turabian StyleMorita, Yuji, Junko Tomida, and Yoshiaki Kawamura. 2012. "Multidrug Efflux Systems in Helicobacter cinaedi" Antibiotics 1, no. 1: 29-43. https://doi.org/10.3390/antibiotics1010029