
Dual Regulation of the Small RNA MicC and the Quiescent Porin OmpN in Response to Antibiotic Stress in Escherichia coli


UMR_MD1, Aix-Marseille Univ & Institut de Recherche Biomédicale des Armées, 27 Boulevard Jean Moulin, 13005 Marseille, France
*
Author to whom correspondence should be addressed.
Antibiotics 2017, 6(4), 33; https://doi.org/10.3390/antibiotics6040033
Submission received: 27 October 2017
/
Revised: 27 November 2017
/
Accepted: 3 December 2017
/
Published: 6 December 2017
(This article belongs to the Special Issue Top 35 of Antibiotics Travel Awards 2017)
Abstract
:Antibiotic resistant Gram-negative bacteria are a serious threat for public health. The permeation of antibiotics through their outer membrane is largely dependent on porin, changes in which cause reduced drug uptake and efficacy. Escherichia coli produces two major porins, OmpF and OmpC. MicF and MicC are small non-coding RNAs (sRNAs) that modulate the expression of OmpF and OmpC, respectively. In this work, we investigated factors that lead to increased production of MicC. micC promoter region was fused to lacZ, and the reporter plasmid was transformed into E. coli MC4100 and derivative mutants. The response of micC–lacZ to antimicrobials was measured during growth over a 6 h time period. The data showed that the expression of micC was increased in the presence of β-lactam antibiotics and in an rpoE depleted mutant. Interestingly, the same conditions enhanced the activity of an ompN–lacZ fusion, suggesting a dual transcriptional regulation of micC and the quiescent adjacent ompN. Increased levels of OmpN in the presence of sub-inhibitory concentrations of chemicals could not be confirmed by Western blot analysis, except when analyzed in the absence of the sigma factor σE. We suggest that the MicC sRNA acts together with the σE envelope stress response pathway to control the OmpC/N levels in response to β-lactam antibiotics.
1. Introduction
Antibacterial resistance is broadly recognized as a growing threat for human health [1,2,3]. As such, increasing antibiotic treatment failures due to multidrug resistant (MDR) bacteria have stirred the urgent need to better understand the underlying molecular mechanisms and promote innovation, with the development of new antibiotics and alternative therapies [4,5]. The efficacy of antibacterial compounds depends on their capacity to reach inhibitory concentrations at the vicinity of their target. This is particularly challenging for drugs directed against Gram-negative bacteria, which exhibit a complex envelope comprising two membranes and transmembrane efflux pumps [6]. The Gram-negative envelope comprises an inner membrane (IM), which is a symmetric phospholipid bilayer; a thin peptidoglycan (PG) layer ensuring the cell shape; and an outer membrane (OM) that is an asymmetric bilayer, composed of an inner phospholipid leaflet and an outer leaflet of lipopolysaccharide (LPS) [7]. First, the OM is a barrier to both hydrophobic and hydrophilic compounds, including necessary nutrients, metabolic substrates and antibiotics, but access is provided by the water filled β-barrel channels called porins [8,9]. In Escherichia coli, the channels of the general porins OmpF and OmpC, are size restricted, and show a preference for passage of hydrophilic charged compounds, including antibiotics such as β-lactams and fluoroquinolones. Second, constitutive tripartite RND (resistance–nodulation–cell division) efflux pumps, such as the AcrAB–TolC pump of E. coli, play a major role in removing antibiotics from the periplasm [10]. Importantly, it has been noted that the efflux pumps are synergized by the OM, since, once ejected into the extracellular space, compounds must re-traverse the restricted-permeability OM barrier [10]. Not surprisingly, MDR clinical isolates of Enterobacteriaceae generally exhibit porin loss and/or increased efflux, which both contribute to reduce the intracellular accumulation of antibiotics below the threshold that would be efficient for activity [9,10,11].
Given the importance of the OM in controlling the uptake of beneficial as well as toxic compounds, one can expect that the expression of porins depends on environmental factors, and is well-coordinated at the transcriptional and post-transcriptional levels. Best studied transcriptional regulators are the IM sensor kinase EnvZ and its cognate response regulator OmpR [12]. EnvZ autophosphorylates in response to a specific envelope stress, such as high osmolarity, then transfers its phosphate group to OmpR. OmpR and OmpR-P have different binding affinities to the porin promoters. At low osmolarity, OmpR activates ompF transcription, whereas at high osmolarity, OmpR-P represses ompF transcription and activates ompC transcription. This differential regulation of OmpF and OmpC is consistent with that in high osmolarity environments, such as in a host where nutrients are abundant, the small pore porin OmpC is predominant, thus limiting the uptake of toxic bile salts; whereas in low osmolarity environments where nutrients are scarce, the large pore porin OmpF is expressed [8]. EnvZ–OmpR [12] and CpxA–CpxR [13] are the main two-component systems involved in the transcriptional control of OmpF and OmpC. Interestingly, the two systems are interconnected [14], and mutations have been found in response to antibiotic stresses [15] (Masi M, Pagès J.-M and Kohler T, personal observations).
The post-transcriptional repression of OmpF by the small regulatory RNA (sRNA) MicF has been discovered in 1984 [16,17,18]. This 93 nucleotide (nt) RNA is divergent to the ompC gene, and acts by direct base-pairing to a region that encompasses the ribosome binding site (RBS) and the start codon of the ompF mRNA, thus preventing translation initiation [19]. The expression of the MicF sRNA is subjected to multiple signals and regulatory pathways [20]. Positive regulation includes EnvZ–OmpR in high osmolarity conditions [21], SoxS in response to oxidative stress [22], and MarA in response to antibiotic stress [23]. The 109 nt MicC sRNA has been discovered more recently, and shown to repress OmpC by direct base-pairing to a 5′ untranslated region of the ompC mRNA [24]. Interestingly, MicC is transcribed clockwise, and is opposite to the adjacent ompN gene that encodes a quiescent porin homologous to OmpF and OmpC [25]. Due to the similar genetic organization of ompN–micC and ompC–micF, and the co-induction of ompC and micF under specific conditions (i.e., high osmolarity via EnvZ–OmpR), it has been suggested that ompN and micC could also be subjected to dual regulation [24]. With the recent interest in post-transcriptional regulators, additional sRNAs that modulate expression of abundant OM proteins have been found. As yet, the ompC mRNA is targeted by multiple sRNAs MicC [24], RybB [26], RseX [27], and IpeX [28,29,30]. To date, external growth conditions and regulatory factors that control the expression of MicC and/or OmpN remain largely unknown.
In this work, we first examined the transcription of micC and ompN in E. coli MC4100 cells grown under a series of external conditions by using lacZ transcriptional fusions and β-galactosidase assays. We optimized the assay by using 96-well microtiter plates, and screened the entire collection of compounds provided by the Biolog Phenotype MicroArraysTM for bacterial chemical susceptibility, in order to extend the range of putative inducing cues. Results showed that high concentrations of carbapenems and cephalosporins, two clinically relevant classes of β-lactams, induce both micC and ompN. Then, the impact of carefully chosen inducing conditions on the expression levels of OmpC and OmpN was tested by Western blotting with appropriate antisera. Because the OmpN protein was undetectable in the presence of mild antibiotic stress conditions, we reasoned to investigate the transcription of micC and ompN in a series of MC4100 derivatives carrying null mutations or multicopy plasmids in order to identify putative transcriptional regulators. Interestingly, we found that OmpN was specifically expressed when the envelope stress sigma factor σE was depleted by the overexpression of the anti-sigma RseA, or when the hns gene encoding the histone nucleoid structuring protein, H-NS, was inactivated. Finally, we examined the functional relevance of OmpN as compared to OmpC and OmpF, with respect to drug translocation.
All these data are discussed considering the current knowledge on the Gram-negative envelope stress response pathways.
2. Results
2.1. Screening of MicC and OmpN Inducing Conditions Using LacZ Transcriptional Fusions and BiologTM Plates
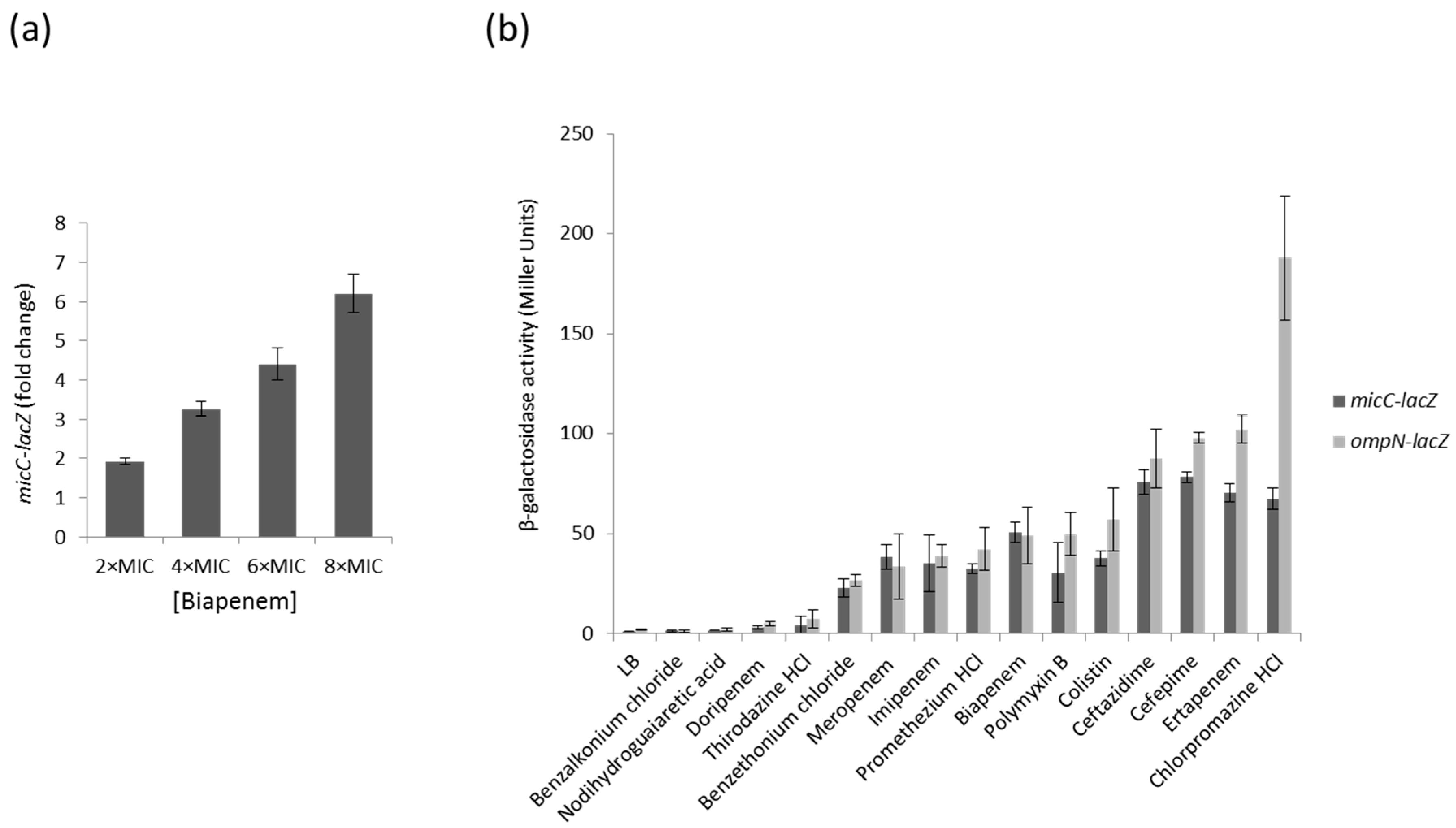
Changes in porin expression play a major role in the development of antibacterial resistance. Because increased levels of MicC are associated with a decreased expression of OmpC, we aimed to examine the expression profile of the MicC sRNA by using a micC–lacZ transcriptional fusion in MC4100 cells grown under a series of growth conditions and β-galactosidase assays. First, we selected a number of representative growth conditions, some of which are sensed by known regulatory factors: growth phase (stationary phase accumulates RpoS), exposure to heat shock, high osmolarity (activates EnvZ–OmpR), iron or nitrogen starvation, or exposure to chemicals, such as salicylate (activates MarA), paraquat (activates SoxR/S), or different classes of antibiotics (β-lactams and fluoroquinolones). To determine whether MicC and OmpN are co-regulated, the β-galactosidase activity of an ompN–lacZ transcriptional fusion was also tested in MC4100 grown under the same conditions. These preliminary assays showed that growth conditions that are known to induce specific regulatory factors, such as RpoS, EnvZ–OmpR, MarA and SoxR/S, do not significantly affect the activity of the micC– and ompN–lacZ fusions, suggesting that the expression of MicC and OmpN is not controlled by these regulators. Instead, these assays allowed the identification of β-lactams potent inducers of both the micC– and ompN–lacZ fusions. As an example, Figure 1a shows that increasing concentrations of the carbapenem biapenem were accompanied with increased β-galactosidase activities. In order to extend the range of putative inducing compounds, we optimized the β-galactosidase assay using preloaded 96-well microtiter plates, and then screened Phenotype MicroArraysTM plates (Biolog PM11 to PM19) for bacterial chemical susceptibility (Supplementary Data 1). A total of 18 compounds were found to increase the activity of the micC– and ompN–lacZ fusions more than 10 times, and 6 of them were selected for further investigations. Concentrations of compounds for β-galactosidase assays adapted to microtiter plates were determined with respect to their MICs (Supplementary Data 2). The data showed that the activity of the micC– and ompN–lacZ fusions were strongly increased when cells were exposed to carbapenems (i.e., biapenem and ertapenem) or cephalosporins (i.e., ceftazidime and cefepime) (Figure 1b). Interestingly, these compounds belong to the most potent subclasses of clinically used β-lactams used for treating Gram-negative infections. Other strong inducers include antiseptics (e.g., benzalkonium chloride and benzethonium chloride) and anesthetics (e.g., chlorpromazine HCl), which are also used in the clinics (Figure 1b).
2.2. Effects of MicC and OmpN Inducing Conditions on the Expression Levels of OmpC and OmpN
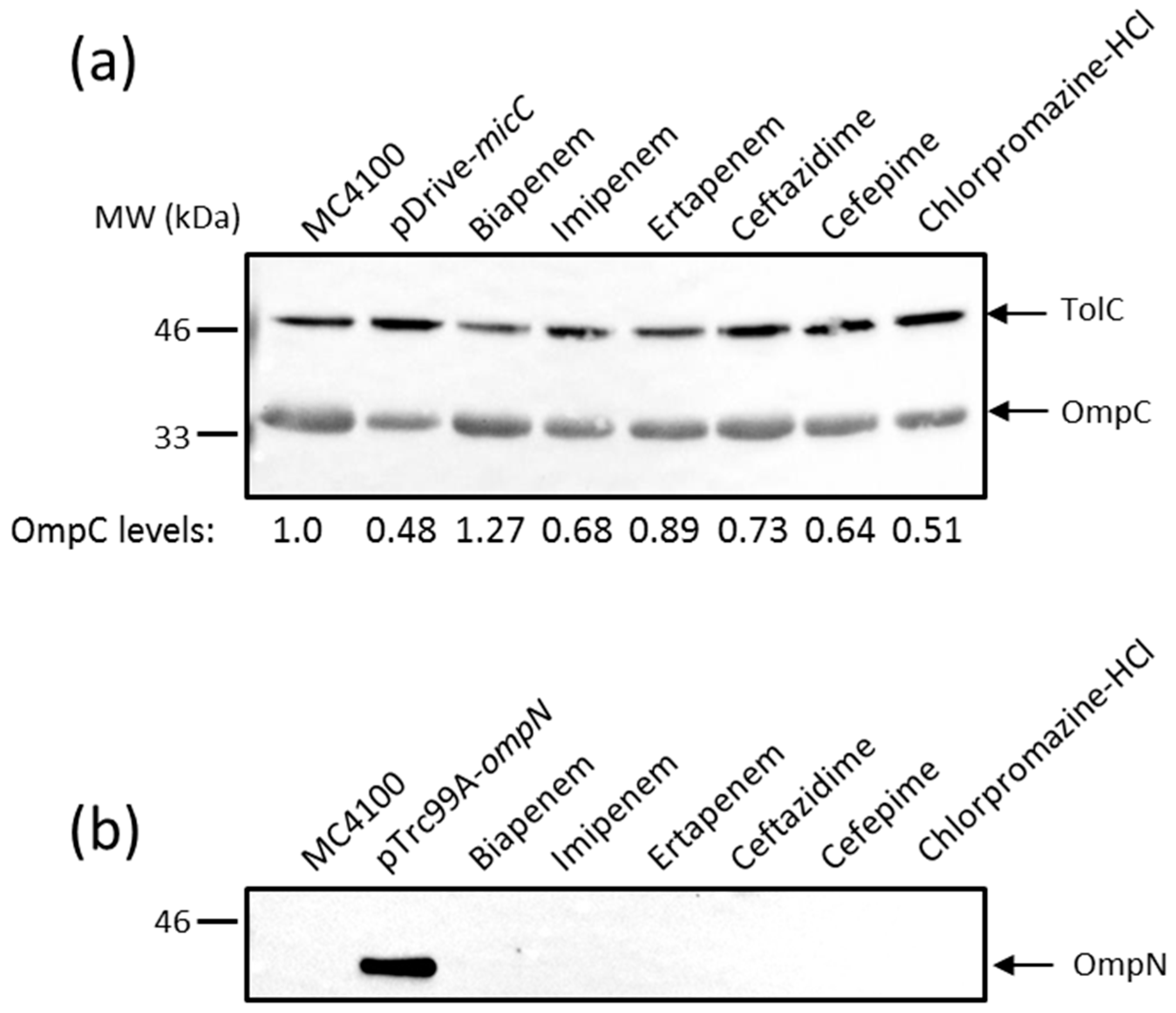
The effect of MicC overexpression on ompC expression was first examined by monitoring OmpC protein levels directly. MC4100 was transformed with the MicC overexpression plasmid (pSD01) and the corresponding empty vector (pDrive). Cultures were induced with IPTG to allow MicC expression, OM extracts were prepared, and levels of OmpC were analyzed by Western blot (WB) with specific anti-peptide antibodies. As shown in Figure 2a, the overexpression of MicC clearly resulted in reduced OmpC levels, confirming that the MicC sRNA represses ompC expression. As noted in the section above, high micC–lacZ activities were obtained in the presence of high concentrations of compounds, which were detrimental for the cell growth. Therefore, MC4100 was cultured in the presence of sub-inhibitory concentrations of inducing compounds—namely biapenem, imipenem, ertapenem, ceftazidime, cefepime, and chlorpromazine HCl—in order to obtain exponentially grown cells and examine their effect on OmpC protein levels. As shown in Figure 2a, these conditions only weakly altered OmpC levels.
Given the co-induction of micC and ompN, we also tested whether OmpN expression was increased in the same samples. As a control, MC4100 was transformed with the OmpN overexpression plasmid (pSD04) and the corresponding empty vector (pTrc99A). Cultures were induced with IPTG to allow OmpN expression; OM extracts were prepared and tested for OmpN expression by WB. For this, we generated antibodies against a peptide in loop 7 present in OmpN, but absent in OmpF and OmpC. A single protein of about 39 kDa was detected in the OM extracts of MC4100 (pSD04), but not in that of MC4100 (pTrc99A), suggesting that the detected band is OmpN without cross-reactivity to other porins, and that OmpN production from the chromosome is undetectable (Figure 2b). However, OmpN production was also undetectable in OM extracts prepared from cells grown in the presence of sub-inhibitory concentrations of micC inducing compounds (Figure 2b).
These results suggest that transient exposure of the cells to sub-inhibitory concentrations of micC–lacZ inducing compounds was not sufficient to yield high levels of MicC and concomitant changes in the porin expression profile. Moreover, it is worth to note that Western blot analysis only provides steady-state levels of OmpC and OmpN. Additional reverse transcription PCR and pulse-chase experiments are needed to conclude the effects of micC and ompN inducing conditions on the expression of OmpC and OmpN at the transcriptional and post-transcriptional levels, respectively.
2.3. Identification of Genetic Factors That Impact on MicC and OmpN Expression
micC–lacZ and ompN–lacZ transcriptional fusions were transformed into MC4100 derivatives carrying either chromosomal null mutations or overexpression plasmids of several regulatory factors, in order to identify putative repressors or activators, respectively. In Enterobacteriaceae, global regulators MarA and RamA have been reported to induce MDR associated with an increase in efflux pump production and a decrease in OmpF expression levels [31,32]. We detected no induction of the reporter fusions, either when these factors were overexpressed from multicopy plasmids or when the corresponding genes were inactivated (data not shown). This observation suggests that the micC–ompN operon is not part of the MarA and RamA regulatory pathways, or is strongly silenced by an upstream repressor.
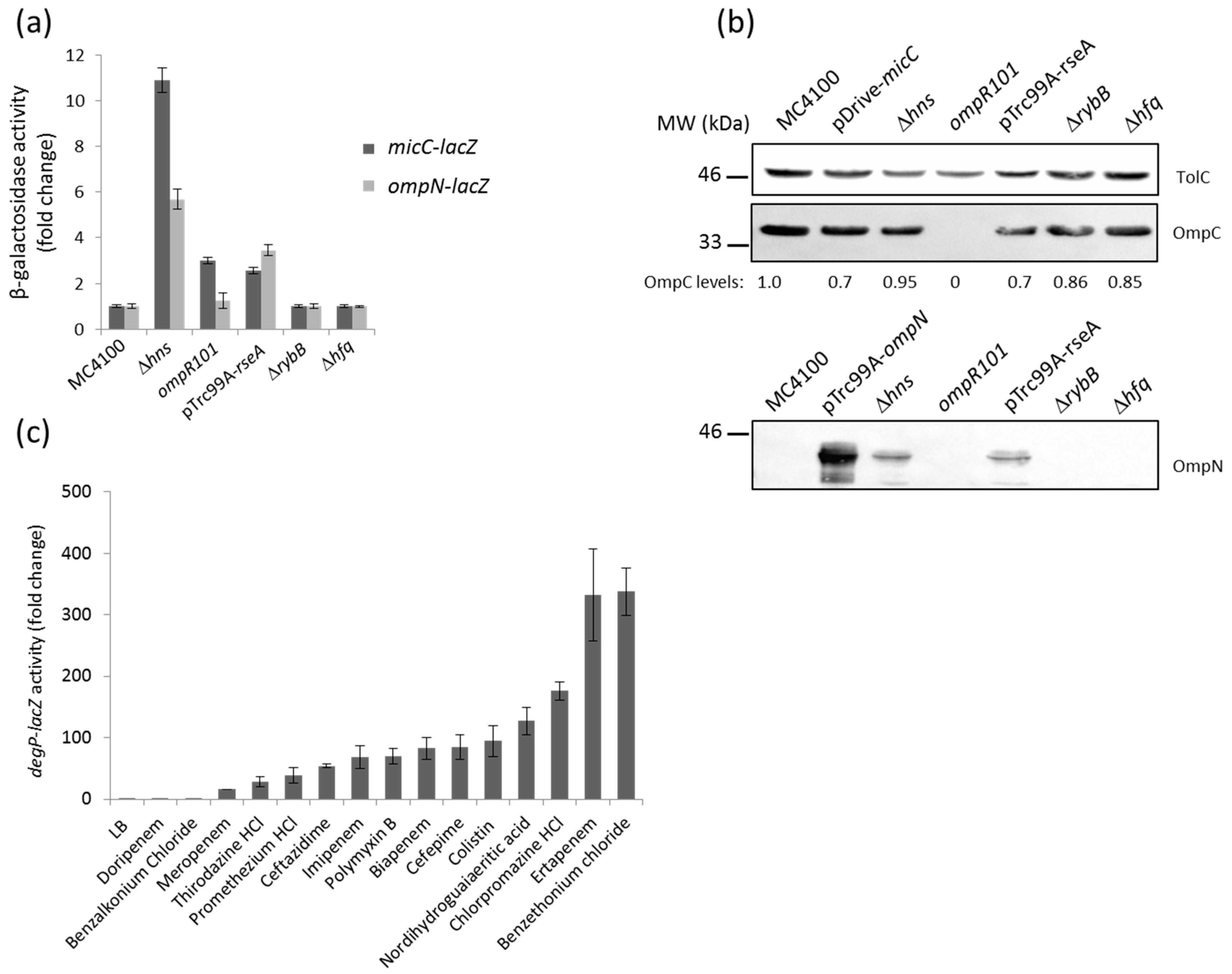
Previous Northern blotting analysis showed that the expression of MicF (repressor of OmpF) was opposite to that of MicC (repressor of OmpC) under most of the tested conditions [24]. Because the osmoregulator OmpR is known to modulate MicF and control the opposite expression of OmpF and OmpC, we tested the impact of an ompR mutation on micC and ompN expression. Here, the activity of the micC–lacZ, but not that of the ompN-lacZ fusion, was slightly increased in the ompR null mutant, thus confirming that OmpR represses MicC (Figure 3a). Whether this regulation is direct or indirect is still unknown.
The last decade has been marked by the identification of several sRNAs. These are differentially expressed, and have been assigned to various important regulons of E. coli and Salmonella. Examples include the RyhB sRNA as a member of the iron-responsive Fur regulon [33]; MicA and RybB, which are activated by the envelope stress sigma factor, σE [26,34,35]; CyaR, whose transcription is governed by the cAMP-CRP complex [36,37]; ArcZ and FnrS, which respond to oxygen availability via the ArcA/B or Fnr systems [38,39]; MgrR, which is a member of the Mg2+-responsive PhoP/Q regulon [40]; SdsR, which is selectively transcribed by the major stationary phase and stress sigma factor, σS [41]; and CpxQ, which responds to the CpxA/R two-component envelope stress system [42,43]. Focusing on envelope stress responses and expression of OM proteins, we examined the impact of CpxA/R and σE on micC and ompN induction. Constitutive activation of the Cpx stress response, by multicopy plasmids expressing an autoactivated CpxA [15] or the signaling lipoprotein NlpE [44], did not increase the activity of the reporter fusion (data not shown). In the opposing scenario, when cells were depleted of σE upon the overexpression of its cognate anti-sigma RseA, the activity of both the micC– and ompN–lacZ fusions resulted in a 3–4-fold increase (Figure 3a). Additionally, OmpN was detected in OM extracts of cells grown under the same conditions (Figure 3b). We suggest this regulation is most likely indirect, as the micC–ompN intergenic region does not contain a σE core promoter motif [45]. Because RybB is one of the most abundant sRNA, represses OmpC as well as other OM proteins, and is part of the σE regulon in E. coli [46], we hypothesized that OmpN could be silenced by RybB. However, the activity of the ompN–lacZ fusion did not increase in rybB and hfq mutants, suggesting that the ompN mRNA is not targeted by RybB or any other Hfq-dependent sRNA (Figure 3a,b).
In order to explore the connection between σE and the MicC/OmpN inducing compounds, we examined the effect of the latter on the expression of DegP, a periplasmic protease/chaperone member of the σE regulon, by using a degP–lacZ fusion [47]. Interestingly, all the compounds that had been identified as inducers of micC– and ompN–lacZ also activated degP–lacZ (Figure 3c). These results suggest a strong link between toxic compounds that target the bacterial envelope, the envelope stress σE pathway, and MicC/OmpN expression [48].
Previous studies on porin regulation reported that the H-NS nucleoid protein binds to the micF–ompC intergenic region. Expression of the major OM proteins, OmpF and OmpC, is affected by hns mutations, such that OmpC expression increases via direct effect at the transcriptional level, while OmpF expression decreases via indirect regulation by the MicF sRNA at the post-transcriptional level [49,50]. Comparative transcriptomic and proteomic studies further confirmed the influence of H-NS on the expression of OmpF and OmpC, but also indicated that ompN was upregulated in an hns mutant [51]. Here, the activity of both the micC– and ompN–lacZ fusions was significantly increased (approximately by 11- and 6-fold, respectively) in an hns mutant (Figure 3a). The OM profile of this mutant is shown and indicates that the expression level of both OmpC and OmpN is increased by 2–3-fold (Figure 3b). Considering that MicC functions as a repressor of OmpC, negative regulation of non-identified OmpC repressors by H-NS could explain upregulation of OmpC in the hns mutant.
2.4. Role of OmpN in Antibiotic Translocation
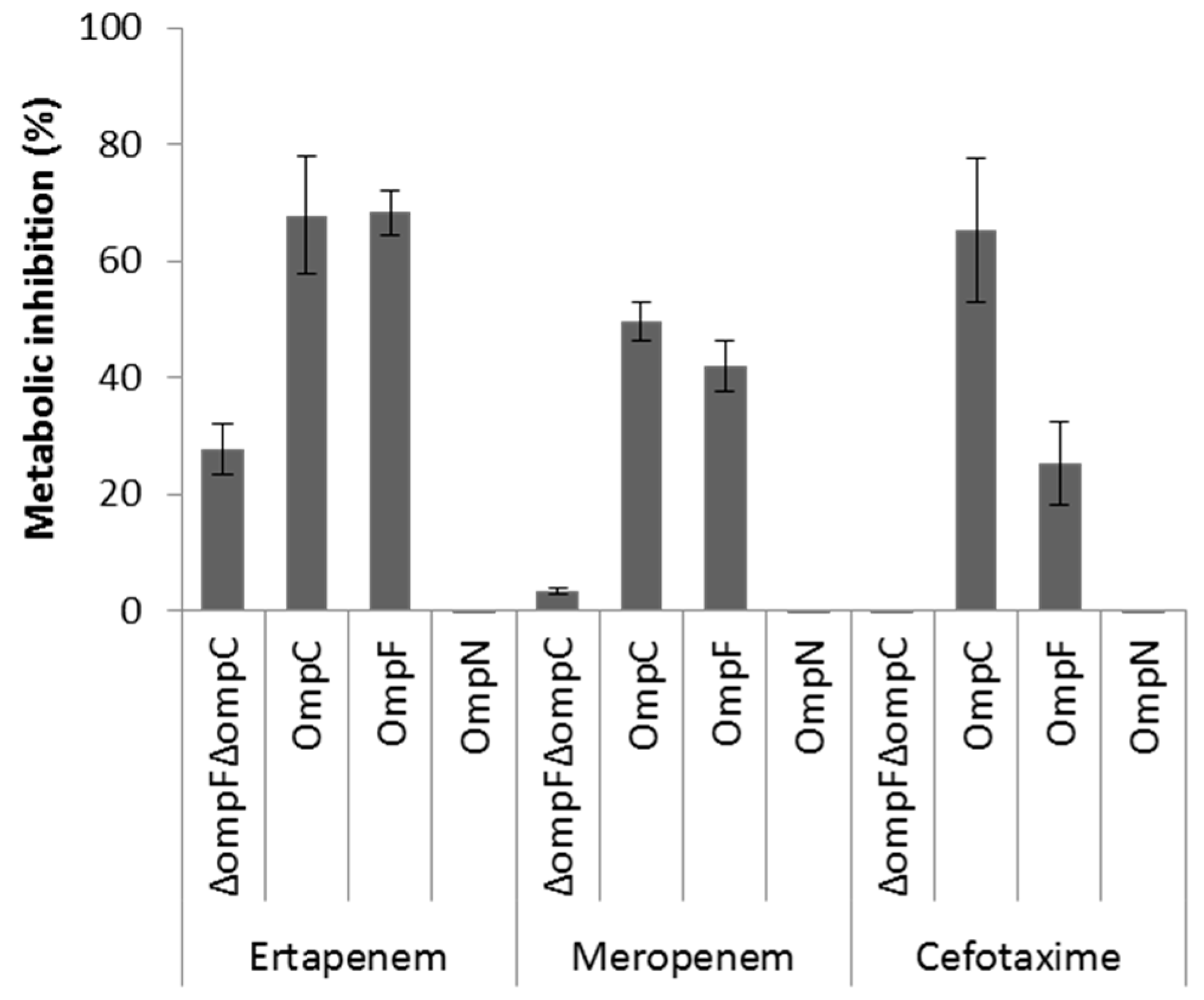
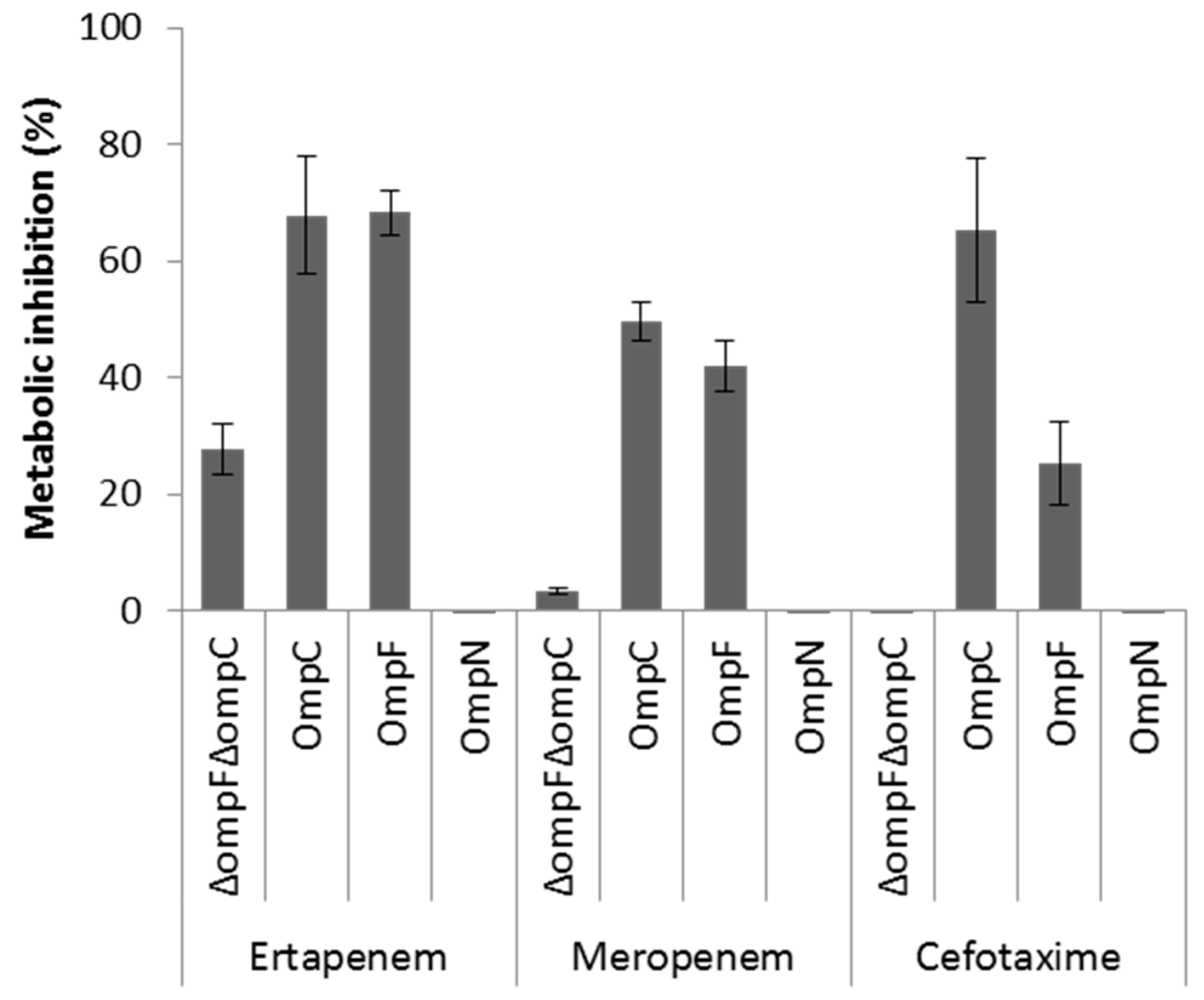
OmpF and OmpC porins represent the preferred route for the uptake of β-lactam antibiotics across the OM of E. coli [6,8,9]. Although OmpN is quiescent porin in E. coli [25], the orthologous OmpK37 of Klebsiella pneumoniae has been shown to be expressed at low levels under standard laboratory growth conditions, but highly expressed in β-lactam-resistant clinical isolates [52]. As a first step to investigate the role of MicC/OmpN in antibiotic susceptibility profile, we examined the expression levels of OmpF, OmpC, and OmpN in a collection of E. coli β-lactam-resistant clinical isolates by WB analysis. None of these isolates produced detectable OmpF, OmpC, or OmpN (Supplementary Data 3). Here, it should be noted that the anti-OmpN antibodies are directed against amino acid residues of the extracellular loop 7, which are specific of E. coli OmpN, but also submitted to variability between strains of this species. The impact of MicC in the downregulation of OmpC in these isolates is not known, and should be further investigated by Northern blot analysis. Second, we used a whole cell-based assay to compare the role of OmpN to that of OmpF and OmpC in the uptake of β-lactam antibiotics. To do this, the metabolic activity of E. coli W3100ΔompF(pTrc99A) (OmpF− OmpC+), W3100ΔompC(pTrc99A) (OmpF+ OmpC−), W3100ΔompFΔompC(pTrc99A) (OmpF− OmpC−) and W3100ΔompFΔompC(pSD04) (OmpF+ OmpC− OmpN+) was monitored in the absence and in presence of representative β-lactams added at inhibitory concentrations, with regards to their capacity to inhibit the reduction of the viability dye resazurin [6]. The results showed that the metabolic activity of E. coli expressing either OmpF or OmpC, but not OmpN, was significantly inhibited upon exposure to β-lactams, suggesting that OmpN is not competent for translocation of this class of antibiotics (Figure 4). However, other approaches, such as liposome swelling assays with reconstituted OmpN, are necessary to conclude on this point.
OmpF and OmpC channels are also used for the translocation of various colicins across the OM of E. coli [53]. We examined the sensitivity of E. coli strains expressing OmpF, OmpC, or OmpN to colicins E2 and E3, by spotting serial 2-fold dilutions onto cell lawns. Interestingly, the expression of any of the three porins yields similar sensitivity (titers of 2 × 10−7), suggesting that OmpN channels are able to bind and transport porin-dependent group A colicins across the OM of E. coli (data not shown). This also points to the different mechanism of antibiotic versus colicin translocation through OM porin channels.
3. Discussion
sRNAs have become important players in bacterial gene regulation. To date, systematic genome-wide searches have led to the identification of approximately 80 sRNAs in E. coli, the majority of which are conserved in Salmonella and other closely related species. About one-third of the reported sRNAs repress synthesis of OM proteins. Evidence for important roles of sRNAs in this post-transcriptional regulation was previously established by the fact that the loss of Hfq, the sRNA chaperone [54], results in the overproduction of OM proteins [24,26,27,36,37,41].
In E. coli, the conserved Hfq-associated sRNA, MicC, was identified as a repressor of the synthesis of OmpC [24,54]. MicC inhibits the 30S ribosome binding through a conserved 22 bp RNA duplex near the start codon of the ompC mRNA [24]. Many parallels have been drawn between the MicC and MicF sRNAs. Both repress the expression of abundant porins by base pairing near the RBS, thereby blocking translation. Both are encoded opposite to another porin gene. Both are also conserved, together with their omp target sequences in Salmonella, K. pneumoniae, and Enterobacter spp. However, major questions such as (i) environmental conditions and/or intracellular regulatory pathways that promote maximal expression of MicC; (ii) the co-regulation of MicC and OmpN; (iii) the impact of such regulation on antibiotic susceptibility; and (iv) the prevalence of MicC/OmpN in MDR clinical isolates remain unanswered. In this work, we used lacZ transcriptional fusions and β-galactosidase assays to show that the expression of micC and ompN is co-regulated in response to antibiotic stress. In particular, β-lactam antibiotics are among the most potent inducers of both micC and ompN. Interestingly, we found that expression of OmpN from a plasmid could not restore the susceptibility of an E. coli porin-less strain to β-lactams. In addition, other studies have demonstrated that strains expressing OmpN, but not OmpF or OmpC, were less susceptible to β-lactams [52,55].
Our results also identified that envelop stress sigma factor σE and H-NS are two major negative regulators of MicC/OmpN. σE is widespread among pathogenic and non-pathogenic bacteria, and becomes activated when bacterial envelope homeostasis is perturbed due to misfolding of OM proteins in the periplasm, or severe OM damage by external stresses [56]. In both cases, the bacteria must decrease the synthesis of major OM proteins. It has been shown that MicA and RybB are the two most abundant sRNAs responsible for the rapid decay of omp mRNAs upon activation of the σE envelope stress response [46,57]. Although β-lactams were found to be potent inducers of the σE envelope stress response, RybB nor any other Hfq-dependent sRNA could be responsible for ompN silencing. This suggests that ompN is not subjected to sRNA post-transcriptional regulation. On the other hand, H-NS is a major component of the bacterial nucleoid, and has pleiotropic effects on gene expression, genome stability, and DNA recombination. Previous work has shown that H-NS was required for full expression of OmpF, and that this involves a role for H-NS in repressing the expression of MicF sRNA [48]. Our results also showed that H-NS had a role in repressing the expression of MicC and OmpN.
4. Materials and Methods
4.1. Plasmids and Bacterial Strains
All the E. coli strains and plasmids used in this study are listed in Table 1. E. coli MC4100 and derivatives were used for lacZ reporter gene assays and protein expression analysis. Knockout mutants were generated by P1 transduction from different sources and cured by using the FLP helper plasmid pCP20 to remove the kanamycin resistance cassette [58]. Strains were routinely grown in Luria Bertani (LB) broth (Sigma, Saint Quentin Fallavier, France), supplemented with the following antibiotics when necessary: ampicillin, 100 μg/mL (Amp); kanamycin, 50 μg/mL (Kan); chloramphenicol (Cam), 30 μg/mL; streptomycin 50 μg/mL (Str). E. coli W3110 and derivatives were used for translocation assays.
4.2. Plasmid Construction
Genomic DNA was extracted from MC4100 by using the Wizard® Genomic purification kit (Promega, Charbonnières-les-Bains, France) according to the manufacturer’s instructions, and used as a template for all PCR-amplifications. micC– and ompN–lacZ transcriptional fusions were constructed in the promoter-less lacZ containing vector pFus2K [59]. A 184 nt fragment containing the MicC promoter was amplified by using the primer pair SD1 (5′-TTACGTATCGGATCCTCGGGGAGTGAAAACATCCT-3′) and SD2 (5′-GCGGATCCCCGCGCAGAATAACGTAT-3′), which contain BamHI restriction sites (underlined) for classic restriction/ligation cloning into BamHI restricted pFus2K (Supplementary data 4) in the orientation of micC–lacZ (pSD02). Because the transcription start of ompN is only based on promoter prediction, the entire intergenic region between MicC and OmpN was PCR-amplified by using the primer pair SD3 (5′-GAGCTCGCATGCGGATCCTGAATAAATCCTTTAGTTATT-3′) and SD4 (5′-CAGGACTCTAGAGGATCCCCGCGCAGAATAACGTAT-3′). This generated a 227 nt fragment, which contained BamHI restriction sites (underlined) and extension homologous to BamHI restricted pFus2K for cloning using the In-Fusion ™ cloning kit (Clontech, Saint Germaine n Laye, France), in the orientation of ompN–lacZ (pSD03) (Supplementary Data 4). For overexpression of the MicC sRNA, a 410 nt PCR fragment was generated by using the primer pair SD1 and SD5 (5′-AGGCTCGAGAAGCTT AGATGCTGCAGCTGAATTTG-3′) inserted into the pDrive vector restricted with BamHI and HindIII under the control of an IPTG inducible promoter by using the In-Fusion ™ cloning kit (pSD01) (Supplementary Data 1). Recombinant plasmids pSD04 and pSD05 were obtained by InFusion cloning of fragments into the pTrc99A vector after digestion with appropriate restriction enzymes. pSD04 contains ompN, which was PCR-amplified by using the primer set SD6 (5′-CATGGAATTCATGAAAAGCAAAGTACTGGCAC-3′) and SD7 (5′-CGACTCAGAGGATCCTTAGAACTGATAAACCAGACCTAAAGCG-3′) that contain the EcoRI and BamHI restriction sites respectively. pSD05 contains rseA, which was PCR-amplified by using the primer pair SD8 (5′-GGTATTAGCCATGGAGAAAG-3′) and SD9 (5′-CTGTGCCGCCCCGGGTACTTTCTG-3′) that contain the NcoI and SmaI restriction sites, respectively. All the plasmid constructs were confirmed by sequencing.
4.3. β-Galactosidase Assays
β-Galactosidase activity was routinely assayed on log-phase bacterial cultures, as described by Miller [69].
4.4. Determination of Minimal Inhibitory Concentrations (MIC)
MIC values of antibiotics were determined by the microdilution method in Mueller Hinton II broth (MHIIB) (Sigma). Susceptibilities were determined in 96-well microtiter plates with an inoculum of 2 × 105 cfu in 200 µL containing two-fold serial dilutions of each compound. The MIC was defined as the lowest concentration of each compound for which no visible growth was observed after 18 h of incubation at 37 °C. Each assay was systematically performed in triplicate. The average of three independent assays was considered in μg/mL.
4.5. Preparation of the Microtiter Plates for β-Galactosidase Assays
The standard β-galactosidase assay was adapted for compound screening by using 96-well microtiter plates and a SUNRISETM Tecan for absorbance readings. Briefly, strains were grown to an OD600 of 0.6. Cultures were diluted to an OD600 of 0.2, and added (200 µL) to the Phenotype MicroArrays ™ test plates (Biolog plates PM11 to PM19) (Supplementary Data 2). After overnight incubation at 37 °C, cells were centrifuged, washed, and treated with ONPG (2-nitrophenyl β-d-galactoside, Sigma) (4 mg/mL). Curves of OD420 were plotted over the time (30 min) to identify optimal inducers (Supplementary data 2). Similar experiments were repeated in 96-well microtiter plates preloaded with a chosen concentration range for each compound: each well was loaded with 20 μL of ONPG (4 mg/mL) and 10 μL of compound dilutions (Supplementary Data 3), then cells (170 µL at an OD600 of 0.2) were added. The plates were incubated at 37 °C inside the reader, and curves of OD420 were plotted over the time (6 h). The obtained readings in presence of ONPG were used to calculate Miller units and for determining the fold change in lacZ activity, relatively to standard growth conditions. Experiments were independently repeated at least three times.
4.6. Preparation of OM Extracts
Bacterial cultures (50 mL), grown in the presence or absence of stress, were incubated according to the optimum micC/ompN induction conditions determined by the β-galactosidase assay. The cells were washed and concentrated 12.5 fold in 20 mM sodium phosphate buffer (pH 7.4), and lysed by one passage through a cell disruptor (Constant Systems) at 2 kbar. After removal of cell debris by centrifugation (7000× g, 20 min, 4 °C) the supernatant was ultracentrifuged (100,000× g, 60 min, 4 °C) to collect the whole cell envelopes. These were resuspended in 0.3% N-laurylsarcosinate, and incubated for 30 min at room temperature to solubilize the IM. The insoluble OM extracts were pelleted by centrifugation (100,000× g, 60 min, 4 °C).
4.7. SDS-PAGE and Western Blot Analysis
OM were prepared as described above, resuspended in 20 mM sodium phosphate buffer (pH 7.4), and kept at −20 °C until use. All samples were diluted in Laemmli buffer (2×: 4% SDS, 20% glycerol, 10% 2-mercaptoethanol, 0.004% bromophenol blue, 125 mM Tris-HCl, pH 6.8) and heated for 5 min at 100 °C before loading. Samples corresponding to 0.2 OD units were separated on 10% SDS-PAGE. To better resolve OmpF and OmpC, 4 M urea was added to the running gel. Proteins were either visualized after straining with Coomassie Brilliant Blue R250 or transferred onto nitrocellulose blotting membranes (GE Healthcare, Aulnay-sous-Bois, France). Primary rabbit antibodies and dilutions were: TolC (1:5000), OmpFd (1:5000), OmpC1 (1:5000), and OmpN (1:1000). Goat anti-rabbit HRP-conjugated secondary antibodies and Clarity Max™ Western ECL Blotting substrates (Bio-Rad, Marnes-la-Coquette, France) were used for detection. Protein bands were visualized with a molecular imager Chemidoc-XRS System (Bio-Rad) and quantified using the Image Lab software (Bio-Rad) by using the TolC band as a standard. Peptide-specific antibodies were used to avoid cross-detection of OmpC and OmpN: OmpC1 antibodies are directed against KNGNPSGEGTSGVTNNG amino acid sequence present in loop 4 [70], and OmpN1 antibodies are directed against the GGADNPAGVDDKDLVKYAD amino acid sequence found in loop 7 (Thermo Scientific Pierce custom antibody service, Villebon-sur-Yvette, France).
4.8. Whole Cell-Based Viability Assay
Resazurin-based CellTiter-Blue® Cell Viability Assay (Promega) was used to determine the metabolic inhibition of cells expressing single porins in the presence of clinically relevant antibiotics as an indicator of porin permeation properties [6]. These assays were performed on W3110 derivatives, i.e., W3110ΔF (expressing OmpC), W3110ΔC (expressing OmpF), and W3110ΔFC transformed with pTrc99A-ompN (expressing OmpN). Overnight cultures were diluted to 1:100 and grown until mid-log phase in MHIIB. Strain containing pTrc99A-ompN was grown in the presence of Amp, and OmpN expression was induced with 0.1 mM IPTG for 1 h at 37 °C. When tested for β-lactam permeation, cultures were diluted to 107 cells/mL in fresh MHIIB containing 10% of CellTiter Viability Reagent. For strains containing pTrc99A-ompN, MHIIB was supplemented with 0.1 mM IPTG, and β-lactamase inhibitors tazobactam and clavulanic acid (4 µg/mL each), to inhibit the activity of the plasmidic AmpC, but not Amp. Microtiter plates (96 well) with black sides and a clear bottom were preloaded with 10 μL of 20× concentrated antibiotic solutions. For each antibiotic, the final concentration in the wells was defined as the maximal concentration that did not alter the metabolism of the porin-less strain, i.e., ertapenem, 0.125 µg/mL; meropenem, 0.125 µg/mL; cefotaxime, 0.0625 µg/mL. Cells (190 µL) were then added to separate wells. Control wells also contained cells with resazurin, but no antibiotic, and resaruzin with antibiotics without cells. Fluorescent signals of resorufin were measured with a TECAN Infinite Pro M200 spectrofluorometer (excitation wavelength 530 nm and emission wavelength 590 nm). Kinetic readings were taken at 37 °C every 10 min for 300 min. The % of metabolic inhibition for each strain exposed to each antibiotic was calculated from the measured difference of relative fluorescence units (RFUs) in the presence (RFUATB) as compared to in the absence (RFUMAX) of antibiotic. All experiments were performed at least four times.
4.9. Colicin Killing Assays
LB agar plates were overlaid with 4 mL of soft agar (with a final agar concentration of 0.75%) containing 100 µL of E. coli overnight cultures. Serial two-fold dilutions of ColE2 or ColE3 (laboratory collection), were spotted in 5 µL drops onto the lawns, and the plates were incubated overnight at 37 °C. Efficiencies of killing were taken as the reciprocal of the highest dilution that gave complete clearing of the lawn.
5. Conclusions
Altogether, these data suggest that exposure to β-lactams induce a complex stress response to reduce the translocation of these antibiotics across the OM in Enterobacteriaceae. Further work will analyze how external stresses, such as β-lactams, interact with the σE envelope stress response and H-NS in laboratory strains, as well as in MDR clinical isolates.
Supplementary Materials
The following are available online at www.mdpi.com/2079-6382/6/4/33/s1, Supplementary Data 1: Screening of micC expression by β-galactosidase assay using preloaded 96-well Phenotype MicroArraysTM plates (Biolog PM11 to PM19) for bacterial chemical susceptibility, Supplementary Data 2: Fifteen compounds were selected to investigate their effects on MicC and OmpN, Supplementary Data 3: The expression of OmpN was evaluated in laboratory and clinical strains of E. coli by Western blot analysis, Supplementary Data 4: Partial pfo(ybdK)-micC-ompN genetic region.
Acknowledgments
We thank E. Dumont, J. Vergalli and members of the laboratory for helpful discussions throughout this work. The research leading to the discussions presented here was conducted as part of the Marie Curie Initial Training Network TRANSLOCATION consortium and has received support from the ITN-2013-607694-Translocation (SD). This work was also supported by Aix-Marseille Univ and Service de Santé des Armées.
Author Contributions
Jean-Marie Pagès and Muriel Masi conceived and designed the experiments; Sushovan Dam performed the experiments; Sushovan Dam, Jean-Marie Pagès and Muriel Masi analyzed the data; Sushovan Dam, Jean-Marie Pagès and Muriel Masi wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- The Review on Antimicrobial Resistance Chaired by Jim O’Neill. Tackling Drug-Resistant Infections Globally: Final Report and Recommandations. Available online: https://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf (accessed on 3 March 2016).
- World Health Organization (WHO). Antimicrobial Resistance: Global Report on Surveillance. Available online: http://apps.who.int/iris/bitstream/10665/112642/1/9789241564748_eng.pdf (accessed on 3 March 2016).
- National Institute of Allergy and Infectious Diseases (NIAID). NIAID’s Antibacterial Resistance Program: Currtent Status and Future Directions. Available online: https://www.niaid.nih.gov/sites/default/files/arstrategicplan2014.pdf (accessed on 3 March 2016).
- Stavenger, R.A.; Winterhalter, M. Translocation project: How to get good drugs into bad bugs. Sci. Transl. Med. 2014, 6, 228ed7. [Google Scholar] [CrossRef] [PubMed]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.; Wertheim, H.F.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic resistance-the need for global solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef]
- Masi, M.; Réfregiers, M.; Pos, K.M.; Pagès, J.-M. Mechanisms of envelope permeability and antibiotic influx and efflux in Gram-negative bacteria. Nat. Microbiol. 2017, 2, 17001. [Google Scholar] [CrossRef] [PubMed]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed]
- Pagès, J.-M.; James, C.E.; Winterhalter, M. The porin and the permeating antibiotic: A selective diffusion barrier in Gram-negative bacteria. Nat. Rev. Microbiol. 2008, 6, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H.; Pagès, J.-M. Broad-specificity efflux pumps and their role in multidrug resistance of Gram-negative bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef] [PubMed]
- Davin-Regli, A.; Bolla, J.-M.; James, C.E.; Lavigne, J.-P.; Chevalier, J.; Garnotel, E.; Molitor, A. Membrane permeability and regulation of drug “influx and efflux” in enterobacterial pathogens. Curr. Drug Targets 2008, 9, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Pratt, L.A.; Hsing, W.; Gibson, K.E.; Silhavy, T.J. From acids to osmZ: Multiple factors influence synthesis of the OmpF and OmpC porins in Escherichia coli. Mol. Microbiol. 1996, 20, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Guest, R.L.; Raivio, T.L. Role of the Gram-negative envelope stress response in the presence of antimicrobial agents. Trends Microbiol. 2016, 24, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Gerken, H.; Charlson, E.S.; Cicirelli, E.M.; Kenney, L.J.; Misra, R. MzrA: A novel modulator of the EnvZ/OmpR two-component regulon. Mol. Microbiol. 2009, 72, 1408–1422. [Google Scholar] [CrossRef] [PubMed]
- Philippe, N.; Maigre, L.; Santini, S.; Pinet, E.; Claverie, J.-M.; Davin-Régli, A.V.; Pagès, J.-M.; Masi, M. In vivo evolution of bacterial resistance in two cases of Enterobacter aerogenes infections during treatment with imipenem. PLoS ONE 2015, 10, e0138828. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, T.; Chou, M.Y.; Inouye, M. A unique mechanism regulating gene expression: Translational inhibition by a complementary RNA transcript (micRNA). Proc. Natl. Acad. Sci. USA 1984, 81, 1966–1970. [Google Scholar] [CrossRef] [PubMed]
- Delihas, N. Discovery and characterization of the first non-coding RNA that regulates gene expression, micF RNA: A historical perspective. World J. Biol. Chem. 2015, 6, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Inouye, M. The first demonstration of RNA interference to inhibit mRNA function. Gene 2016, 592, 332–333. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.; Delihas, N. micF RNA binds to the 5′ end of ompF mRNA and to a protein from Escherichia coli. Biochemistry 1990, 29, 9249–9256. [Google Scholar] [CrossRef] [PubMed]
- Delihas, N.; Forst, S. MicF: An antisense RNA gene involved in response of Escherichia coli to global stress factors. J. Mol. Biol. 2001, 313, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ramani, N.; Hedeshian, M.; Freundlich, M. micF antisense RNA has a major role in osmoregulation of OmpF in Escherichia coli. J. Bacteriol. 1994, 176, 5005–5010. [Google Scholar] [CrossRef] [PubMed]
- Chou, J.H.; Greenberg, J.T.; Demple, B. Posttranscriptional repression of Escherichia coli OmpF protein in response to redox stress: Positive control of the micF antisense RNA by the soxRS locus. J. Bacteriol. 1993, 175, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.P.; McMurry, L.M.; Levy, S.B. MarA locus causes decreased expression of OmpF porin in multiple-antibiotic-resistant (Mar) mutants of Escherichia coli. J. Bacteriol. 1988, 170, 5416–5422. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhang, A.; Blyn, L.B.; Storz, G. MicC, a second small-RNA regulator of Omp protein expression in Escherichia coli. J. Bacteriol. 2004, 186, 6689–6697. [Google Scholar] [CrossRef] [PubMed]
- Prilipov, A.; Phale, P.S.; Koebnik, R.; Widmer, C.; Rosenbusch, J.-P. Identification and characterization of two quiescent porin genes, nmpC and ompN, in Escherichia coli BE. J. Bacteriol. 1998, 180, 3388–3392. [Google Scholar] [PubMed]
- Johansen, J.; Rasmussen, A.A.; Overgaard, M.; Valentin-Hansen, P. Conserved small non-coding RNAs that belong to the sigmaE regulon: Role in down-regulation of outer membrane proteins. J. Mol. Biol. 2006, 364, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Douchin, V.; Bohn, C.; Bouloc, P. Down-regulation of porins by a small RNA bypasses the essentiality of the regulated intramembrane proteolysis protease RseP in Escherichia coli. J. Biol. Chem. 2006, 281, 12253–12259. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Keller, M.; Vuong, P.; Misra, R. Novel mechanism of Escherichia coli porin regulation. J. Bacteriol. 2006, 188, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.; Papenfort, K. Small non-coding RNAs and the bacterial outer membrane. Curr. Opin. Microbiol. 2006, 9, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Guillier, M.; Gottesman, S.; Storz, G. Modulating the outer membrane with small RNAs. Genes Dev. 2006, 20, 2338–2348. [Google Scholar] [CrossRef] [PubMed]
- Chollet, R.; Bollet, C.; Chevalier, J.; Malléa, M.; Pagès, J.-M.; Davin-Regli, A. mar operon involved in multidrug resistance of Enterobacter aerogenes. Antimicrob. Agents Chemother. 2002, 46, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Chollet, R.; Chevalier, J.; Bollet, C.; Pages, J.-M.; Davin-Regli, A. RamA is an alternate activator of the multidrug resistance cascade in Enterobacter aerogenes. Antimicrob. Agents Chemother. 2004, 48, 2518–2523. [Google Scholar] [CrossRef] [PubMed]
- Massé, E.; Gottesman, S. A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 99, 4620–4625. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.M.; Rhodius, V.A.; Gottesman, S. SigmaE regulates and is regulated by a small RNA in Escherichia coli. J. Bacteriol. 2007, 189, 4243–4256. [Google Scholar] [CrossRef] [PubMed]
- Skovierova, H.; Rowley, G.; Rezuchova, B.; Homerova, D.; Lewis, C.; Roberts, M.; Kormanec, J. Identification of the sigmaE regulon of Salmonella enterica serovar Typhimurium. Microbiology 2006, 152, 1347–1359. [Google Scholar] [CrossRef] [PubMed]
- Johansen, J.; Eriksen, M.; Kallipolitis, B.; Valentin-Hansen, P. Down-regulation of outer membrane proteins by noncoding RNAs: Unraveling the cAMP-CRP- and sigmaE-dependent CyaR-OmpX regulatory case. J. Mol. Biol. 2008, 383, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Pfeiffer, V.; Lucchini, S.; Sonawane, A.; Hinton, J.C.; Vogel, J. Systematic deletion of Salmonella small RNA genes identifies CyaR, a conserved CRP-dependent riboregulator of OmpX synthesis. Mol. Microbiol. 2008, 68, 890–906. [Google Scholar] [CrossRef] [PubMed]
- Mandin, P.; Gottesman, S. Integrating anaerobic/aerobic sensing and the general stress response through the ArcZ small RNA. EMBO J. 2010, 29, 3094–3107. [Google Scholar] [CrossRef] [PubMed]
- Durand, S.; Storz, G. Reprogramming of anaerobic metabolism by the FnrS small RNA. Mol. Microbiol. 2010, 75, 1215–1231. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.; Six, D.A.; Lee, H.J.; Raetz, C.R.; Gottesman, S. Complex transcriptional and post-transcriptional regulation of an enzyme for lipopolysaccharide modification. Mol. Microbiol. 2013, 89, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, K.S.; Papenfort, K.; Berger, A.A.; Vogel, J. A conserved RpoS-dependent small RNA controls the synthesis of major porin OmpD. Nucleic Acids Res. 2012, 40, 3623–3640. [Google Scholar] [CrossRef] [PubMed]
- Grabowicz, M.; Koren, D.; Silhavy, T.J. The CpxQ sRNA negatively regulates Skp to prevent mistargeting of β-barrel outer membrane proteins into the cytoplasmic membrane. mBio 2016, 7, e00312-16. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.; Vogel, J. A 3′ UTR-derived small RNA provides the regulatory noncoding arm of the inner membrane stress response. Mol. Cell 2016, 61, 352–363. [Google Scholar] [CrossRef] [PubMed]
- Snyder, W.B.; Davis, L.J.; Danese, P.N.; Cosma, C.L.; Silhavy, T.J. Overproduction of NlpE, a new outer membrane lipoprotein, suppresses the toxicity of periplasmic LacZ by activation of the Cpx signal transduction pathway. J. Bacteriol. 1995, 177, 4216–4223. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, G.; Blankschien, M.; Herman, C.; Gross, C.A.; Rhodius, V.A. Regulon and promoter analysis of the E. coli heat-shock factor, sigma32, reveals a multifaceted cellular response to heat stress. Genes Dev. 2006, 20, 1776–1789. [Google Scholar] [CrossRef] [PubMed]
- Gogol, E.B.; Rhodius, V.A.; Papenfort, K.; Vogel, J.; Gross, C.A. Small RNAs endow a transcriptional activator with essential repressor functions for single-tier control of a global stress regulon. Proc. Natl. Acad. Sci. USA 2011, 108, 12875–12880. [Google Scholar] [CrossRef] [PubMed]
- Raivio, T.L.; Silhavy, T.J. Transduction of envelope stress in Escherichia coli by the Cpx two-component system. J. Bacteriol. 1997, 179, 7724–7733. [Google Scholar] [CrossRef] [PubMed]
- Viveiros, M.; Dupont, M.; Rodrigues, L.; Couto, I.; Davin-Regli, A.; Martins, M.; Pagès, J.-M.; Amaral, L. Antibiotic stress, genetic response and altered permeability of E. coli. PLoS ONE 2007, 2, e365. [Google Scholar] [CrossRef] [PubMed]
- Deighan, P.; Free, A.; Dorman, C.J. A role for the Escherichia coli H-NS-like protein StpA in OmpF porin expression through modulation of micF RNA stability. Mol. Microbiol. 2000, 38, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Ueguchi, C.; Mizuno, T. H-NS regulates OmpF expression through micF antisense RNA in Escherichia coli. J. Bacteriol. 1996, 178, 3650–3653. [Google Scholar] [CrossRef] [PubMed]
- Hommais, F.; Krin, E.; Laurent-Winter, C.; Soutourina, O.; Malpertuy, A.; Le Caer, J.P.; Danchin, A.; Bertin, P. Large-scale monitoring of pleiotropic regulation of gene expression by the prokaryotic nucleoid-associated protein, H-NS. Mol. Microbiol. 2001, 40, 20–36. [Google Scholar] [CrossRef] [PubMed]
- Doménech-Sánchez, A.; Hernández-Allés, S.; Martínez-Martínez, L.; Benedí, V.J.; Albertí, S. Identification and characterization of a new porin gene of Klebsiella pneumoniae: Its role in beta-lactam antibiotic resistance. J. Bacteriol. 1999, 181, 2726–2732. [Google Scholar] [PubMed]
- Housden, N.G.; Kleanthous, C. Colicin translocation across the Escherichia coli outer membrane. Biochem. Soc. Trans. 2012, 40, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Wassarman, K.M.; Rosenow, C.; Tjaden, B.C.; Storz, G.; Gottesman, S. Global analysis of small RNA and mRNA targets of Hfq. Mol. Microbiol. 2003, 50, 1111–1124. [Google Scholar] [CrossRef] [PubMed]
- Fàbrega, A.; Rosner, J.L.; Martin, R.G.; Solé, M.; Vila, J. SoxS-dependent coregulation of ompN and ydbK in a multidrug-resistant Escherichia coli strain. FEMS Microbiol. Lett. 2012, 332, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Raivio, T.L.; Silhavy, T.J. Periplasmic stress and ECF sigma factors. Annu. Rev. Microbiol. 2001, 55, 591–624. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Pfeiffer, V.; Mika, F.; Lucchini, S.; Hinton, J.C.; Vogel, J. SigmaE-dependent small RNAs of Salmonella respond to membrane stress by accelerating global omp mRNA decay. Mol. Microbiol. 2006, 62, 1674–1688. [Google Scholar] [CrossRef] [PubMed]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Pagès, J.-M.; Villard, C.; Pradel, E. The eefABC multidrug efflux pump operon is repressed by H-NS in Enterobacter aerogenes. J. Bacteriol. 2005, 187, 3894–3897. [Google Scholar] [CrossRef] [PubMed]
- Casadaban, M.J. Transposition and fusion of the lac genes to selected promoters in Escherichia coli using bacteriophage lambda and Mu. J. Mol. Biol. 1976, 104, 541–555. [Google Scholar] [CrossRef]
- Sarma, V.; Reeves, P. Genetic locus (ompB) affecting a major outer-membrane protein in Escherichia coli K-12. J. Bacteriol. 1977, 132, 23–27. [Google Scholar] [PubMed]
- Bachmann, B.J. Pedigrees of some mutant strains of Escherichia coli K-12. Bacteriol. Rev. 1972, 36, 525–557. [Google Scholar] [PubMed]
- Klein, G.; Lindner, B.; Brade, H.; Raina, S. Molecular basis of lipopolysaccharide heterogeneity in Escherichia coli: Envelope stress-responsive regulators control the incorporation of glycoforms with a third 3-deoxy-α-d-manno-oct-2-ulosonic acid and rhamnose. J. Biol. Chem. 2011, 286, 42787–42807. [Google Scholar] [CrossRef] [PubMed]
- Bertin, P.; Terao, E.; Lee, E.H.; Lejeune, P.; Colson, C.; Danchin, A.; Collatz, E. The H-NS protein is involved in the biogenesis of flagella in Escherichia coli. J. Bacteriol. 1994, 176, 5537–5540. [Google Scholar] [CrossRef] [PubMed]
- George, A.M.; Levy, S.B. Amplifiable resistance to tetracycline, chloramphenicol, and other antibiotics in Escherichia coli: Involvement of a non-plasmid-determined efflux of tetracycline. J. Bacteriol. 1983, 155, 531–540. [Google Scholar] [PubMed]
- Rosner, J.L.; Slonczewski, J.L. Dual regulation of inaA by the multiple antibiotic resistance (mar) and superoxide (soxRS) stress response systems of Escherichia coli. J. Bacteriol. 1994, 176, 6262–6269. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006. [Google Scholar] [CrossRef] [PubMed]
- Guzman, L.M.; Belin, D.; Carson, M.J.; Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995, 177, 4121–4130. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1972. [Google Scholar]
- Simonet, V.; Malle, M.; Fourel, D.; Bolla, J.-M.; Pages, J.-M. Crucial domains are conserved in Enterobacteriaceae porins. FEMS Microbiol. Lett. 1996, 136, 91–97. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a) Dose dependent micC–lacZ activity in presence of increasing concentrations of biapenem (MIC of 0.32 µg/mL); (b) β-galactosidase activity of the micC- and ompN-lacZ fusions in the presence of selected compounds. Values are means from three independent determinations, and standard deviation is represented.
Figure 1.
(a) Dose dependent micC–lacZ activity in presence of increasing concentrations of biapenem (MIC of 0.32 µg/mL); (b) β-galactosidase activity of the micC- and ompN-lacZ fusions in the presence of selected compounds. Values are means from three independent determinations, and standard deviation is represented.
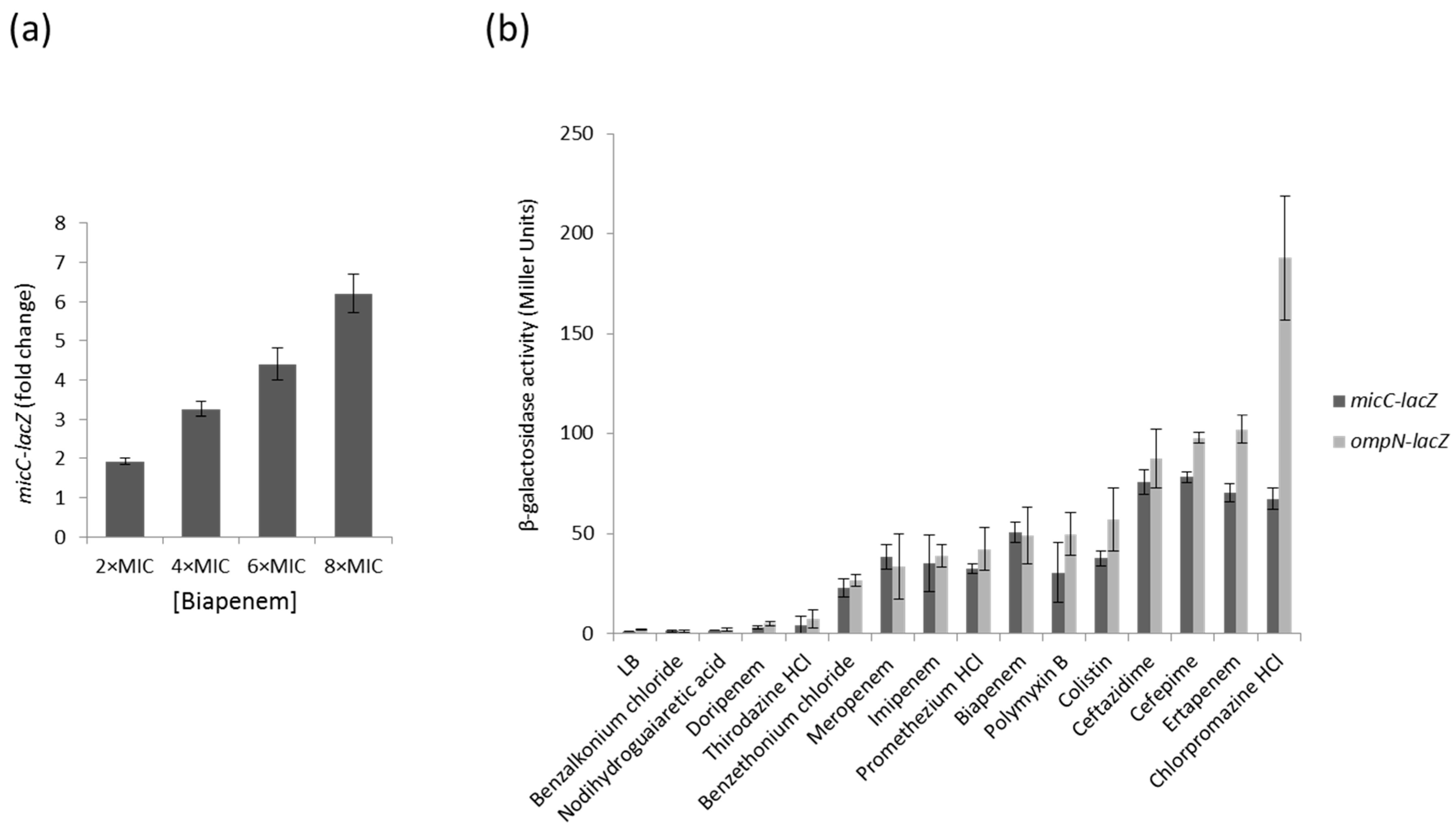
Figure 2.
Western blot (WB) analysis of outer membrane (OM) proteins. Cells were grown, and OM extracts were prepared as described in the Materials and Methods. OM proteins equivalent to 0.2 OD600 units of cultures were separated by SDS-PAGE, electrotransferred on nitrocellulose membranes, and blotted with the appropriate anti-sera. Data show the production of OmpC (a) and OmpN (b). Both the positive controls pDrive-micC and pTrc99A-ompN were induced by 0.4 mM IPTG for 3 h. TolC expression was used for normalizing sample loading, and the expression of normalized OmpC has been expressed in mean values from three independent experiments.
Figure 2.
Western blot (WB) analysis of outer membrane (OM) proteins. Cells were grown, and OM extracts were prepared as described in the Materials and Methods. OM proteins equivalent to 0.2 OD600 units of cultures were separated by SDS-PAGE, electrotransferred on nitrocellulose membranes, and blotted with the appropriate anti-sera. Data show the production of OmpC (a) and OmpN (b). Both the positive controls pDrive-micC and pTrc99A-ompN were induced by 0.4 mM IPTG for 3 h. TolC expression was used for normalizing sample loading, and the expression of normalized OmpC has been expressed in mean values from three independent experiments.
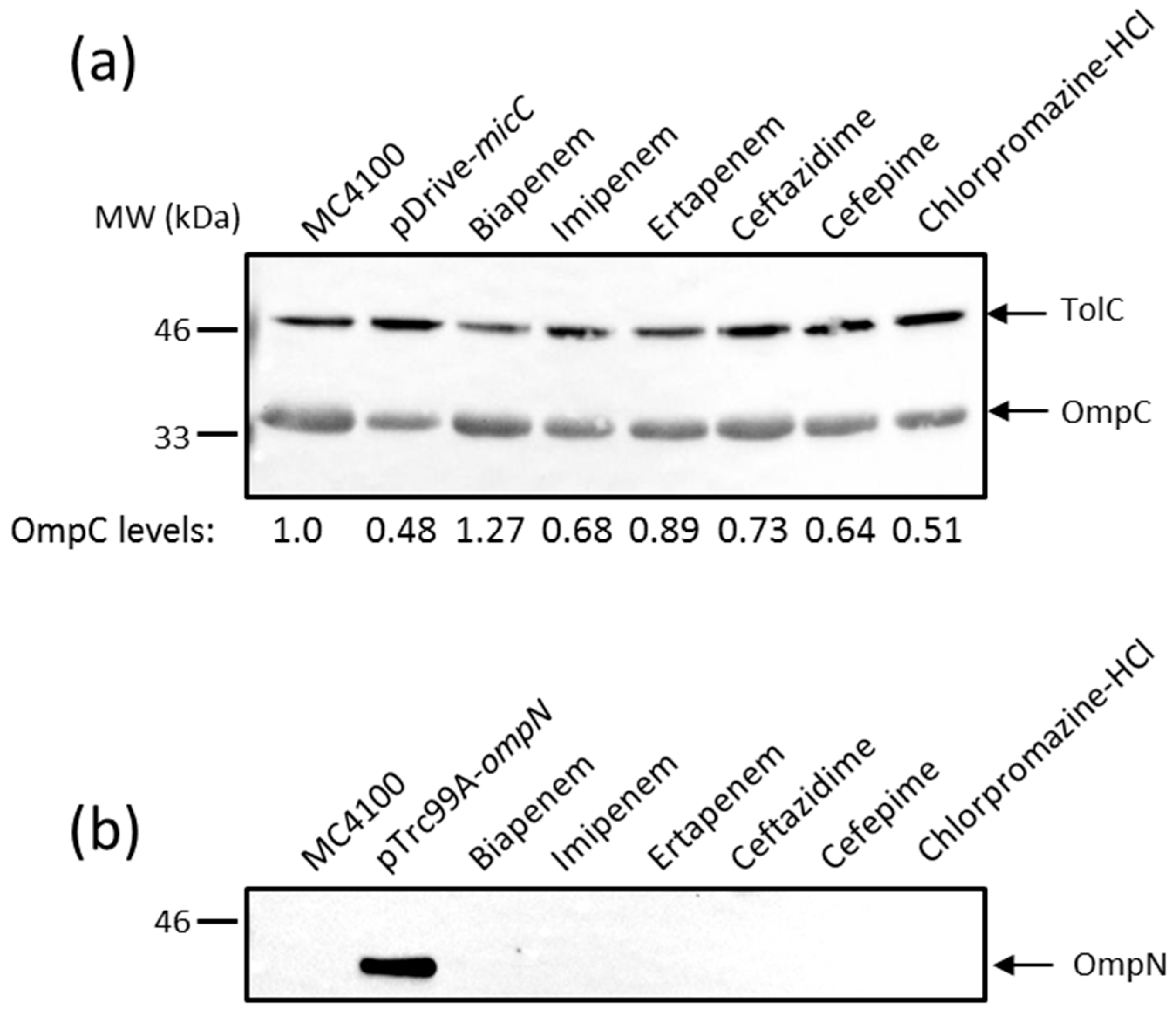
Figure 3.
(a) β-Galactosidase activity of the micC– and ompN–lacZ fusions in different genetic backgrounds. Envelope stress sigma factor σE is essential in Escherichia coli. Therefore, cells were temporarily depleted of σE by the overexpression of the anti-sigma factor RseA with 0.4 mM IPTG under heat shock conditions at 42 °C; (b) WB analysis of OM proteins. Cells were grown, and OM extracts were prepared as described in the Materials and Methods. OM proteins equivalent to 0.2 ODU of cultures were separated by SDS-PAGE, electrotransferred on nitrocellulose membranes, and blotted with the appropriate anti-sera. Data show the production of OmpC (upper panel) and OmpN (lower panel). TolC expression was evaluated for normalizing sample loading and the expression of normalized OmpC has been expressed in numerical values below the bands; (c) β-galactosidase activity of a degP–lacZ chromosomal fusion in response to various external stresses.
Figure 3.
(a) β-Galactosidase activity of the micC– and ompN–lacZ fusions in different genetic backgrounds. Envelope stress sigma factor σE is essential in Escherichia coli. Therefore, cells were temporarily depleted of σE by the overexpression of the anti-sigma factor RseA with 0.4 mM IPTG under heat shock conditions at 42 °C; (b) WB analysis of OM proteins. Cells were grown, and OM extracts were prepared as described in the Materials and Methods. OM proteins equivalent to 0.2 ODU of cultures were separated by SDS-PAGE, electrotransferred on nitrocellulose membranes, and blotted with the appropriate anti-sera. Data show the production of OmpC (upper panel) and OmpN (lower panel). TolC expression was evaluated for normalizing sample loading and the expression of normalized OmpC has been expressed in numerical values below the bands; (c) β-galactosidase activity of a degP–lacZ chromosomal fusion in response to various external stresses.
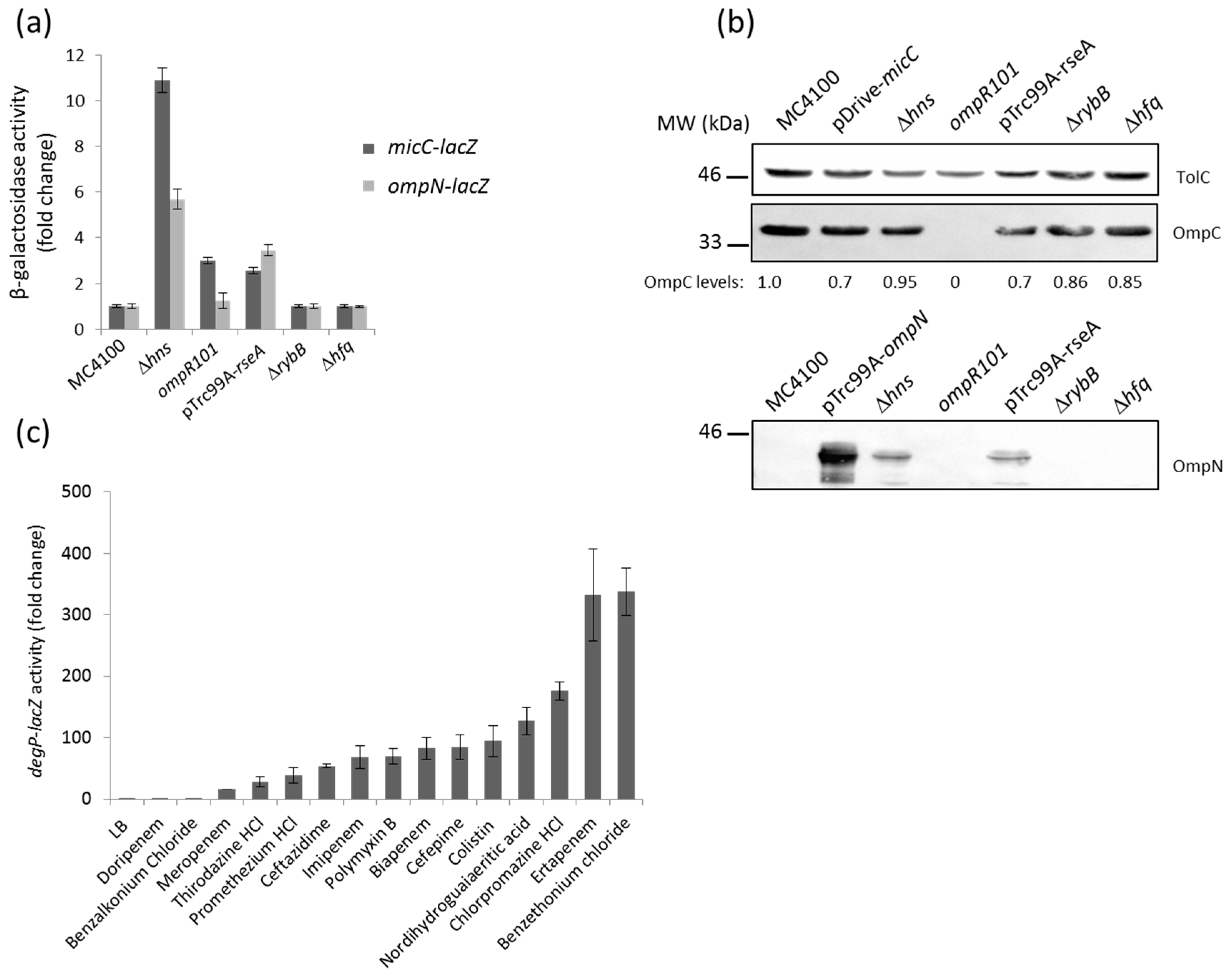
Figure 4.
Metabolic inhibition of intact cells expressing OmpF, OmpC, or OmpN in the presence of selected β-lactam antibiotics using a resazurin-reduction-based assay. Actively metabolizing bacterial cells are able to reduce blue resazurin into red resofurin, which emits fluorescence at 590 nm. The experiment was performed in a microtiter plate, and fluorescence was measured every 10 min with an excitation wavelength of 530 nm and an emission wavelength of 590 nm. Inhibition of resazurin reduction in the presence of appropriate concentrations of each antibiotic was translated into % metabolic inhibition.
Figure 4.
Metabolic inhibition of intact cells expressing OmpF, OmpC, or OmpN in the presence of selected β-lactam antibiotics using a resazurin-reduction-based assay. Actively metabolizing bacterial cells are able to reduce blue resazurin into red resofurin, which emits fluorescence at 590 nm. The experiment was performed in a microtiter plate, and fluorescence was measured every 10 min with an excitation wavelength of 530 nm and an emission wavelength of 590 nm. Inhibition of resazurin reduction in the presence of appropriate concentrations of each antibiotic was translated into % metabolic inhibition.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Strains and plasmids used in this study.
| Strain or Plasmid | Description | Source or Reference |
|---|---|---|
| E. coli strains | ||
| MC4100 | F− [araD139]B/r Δ(argF-lac)169 λ− e14 flhD5301 Δ(fruK-yeiR)725(fruA25) relA1 rpsL150(StrR) rbsR22 Δ(fimB-fimE)632(::IS1) deoC1 | [60] |
| MH1160 | MC4100 ompR101 | [61] |
| TR49 | MC4100 λRS88[degP–lacZ] | [47] |
| W3110 | F− λ− IN(rrnD-rrnE)1 rph-1 | [62] |
| SR8265 | W3110 rybB< >aph, KanR, source for P1 transduction | [63] |
| PS2209 | W3110 ΔlacZ169 | [64] |
| PS2652 | ΔlacZ169 zch-506::TnlO hns-1001::Tnseq1, KanR, source for P1 transduction | [64] |
| AG100 | F− glnX44(AS) galK2(Oc) rpsL704(StrR) xylA5 mtl-1 argE3(Oc) thiE1 tfr-3 | [65] |
| CH164 | AG100 marA zdd-230::Tn9, CamR, source for P1 transduction | [66] |
| BW25113 | F− Δ(araD–araB)567 ΔlacZ4787(::rrnB-3) λ− rph-1 Δ(rhaD–rhaB)568 hsdR514 | [67] |
| JW4130 | BW25113 hfq::kan, KanR, source for P1 transduction | GE Healthcare |
| SD01 | MC4100 ΔrybB | This study |
| SD02 | MC4100 marA zdd-230::Tn9, CamR, | This study |
| SD03 | MC4100 Δhfq | This study |
| SD04 | MC4100 Δhns | This study |
| SD05 | MC4100 ΔrpoS | This study |
| W3110ΔompF | W3110 ompF::kan | M.G. Page |
| W3110ΔompC | W3110 ompC::kan | M.G. Page |
| W3110ΔompFΔompC | W3110 ΔompFΔompC | M.G. Page |
| Plasmids | ||
| pDrive | PCR cloning vector; AmpR, KanR | Qiagen |
| pRC1 | pDrive containing Enterobacter aerogenes MarA | [31] |
| pRC2 | pDrive containing Enterobacter aerogenes RamA | [32] |
| pSD01 | pDrive encoding MicC sRNA | This study |
| pFus2K | Cloning vector with promoter-less lacZ, KanR | [59] |
| pSD02 | pFus2K containing the micC–lacZ fusion | This study |
| pSD03 | pFus2K containing the ompN–lacZ fusion | This study |
| pTrc99A | Expression vector with the inducible PTRC promoter, AmpR | Pharmacia |
| pSD04 | pTrc99A containing OmpN | This study |
| pSD05 | pTrc99A containing RseA | This study |
| pBAD24 | Expression vector with the inducible PBAD promoter, AmpR | [68] |
| pBAD24-NlpE | pBAD24 containing NlpE | M. Masi |
| pBAD33 | Expression vector with the inducible PBAD promoter, CamR | [68] |
| pBAD33-CpxA* | pBAD33 containing an autoactivated (*) CpxA | M. Masi |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dam, S.; Pagès, J.-M.; Masi, M. Dual Regulation of the Small RNA MicC and the Quiescent Porin OmpN in Response to Antibiotic Stress in Escherichia coli. Antibiotics 2017, 6, 33. https://doi.org/10.3390/antibiotics6040033
AMA Style
Dam S, Pagès J-M, Masi M. Dual Regulation of the Small RNA MicC and the Quiescent Porin OmpN in Response to Antibiotic Stress in Escherichia coli. Antibiotics. 2017; 6(4):33. https://doi.org/10.3390/antibiotics6040033
Chicago/Turabian StyleDam, Sushovan, Jean-Marie Pagès, and Muriel Masi. 2017. "Dual Regulation of the Small RNA MicC and the Quiescent Porin OmpN in Response to Antibiotic Stress in Escherichia coli" Antibiotics 6, no. 4: 33. https://doi.org/10.3390/antibiotics6040033
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.