
Sensors of Infection: Viral Nucleic Acid PRRs in Fish

Abstract
:1. Introduction
2. Pattern Recognition Receptors (PRRs)



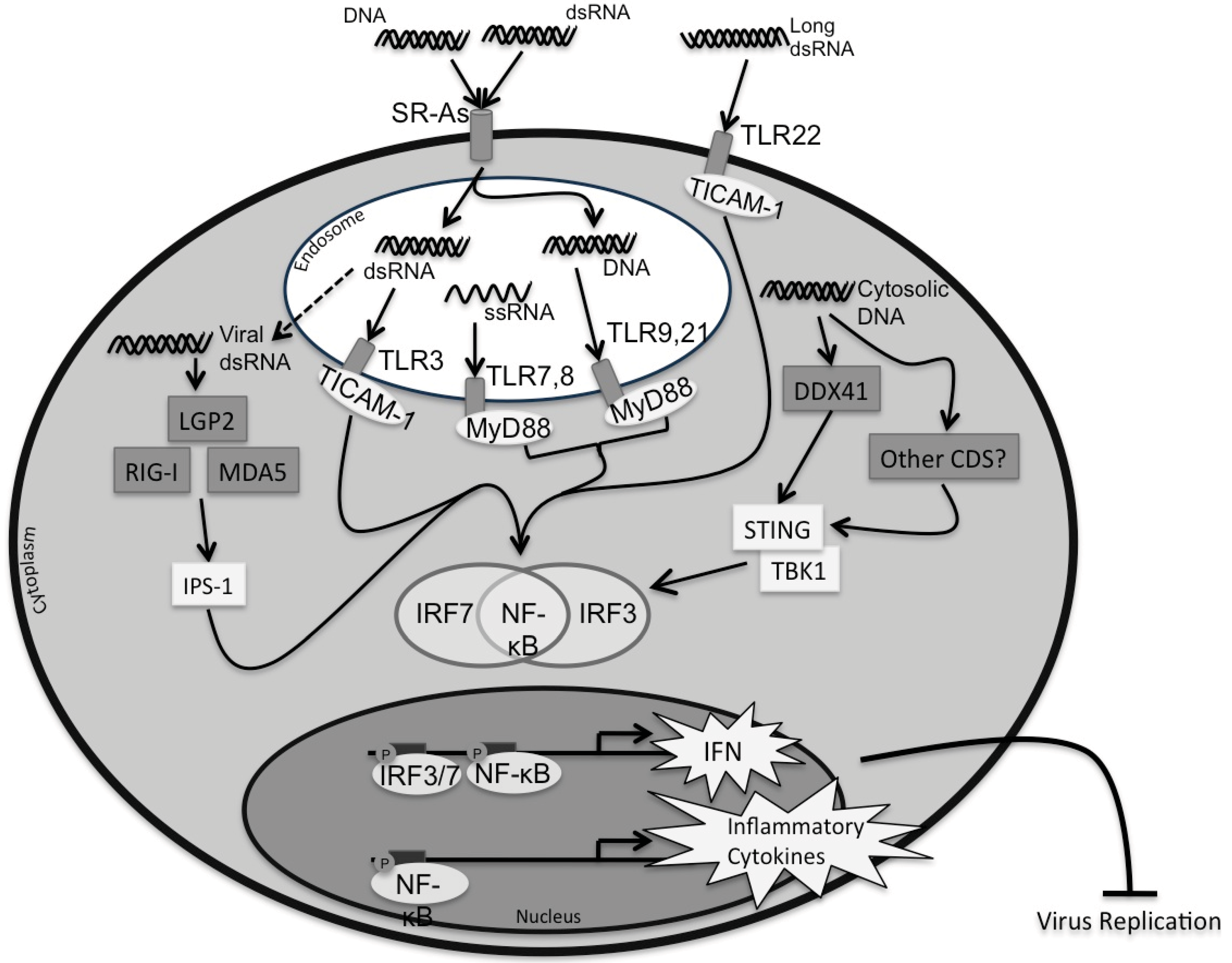{kind=link}
{kind=link}
| Pattern Recognition Receptor | Atlantic Salmon S. salar | Grass Carp C. idella | Channel Catfish I. punctatus | Fugu T. rubripes | Japanese Flounder P. olivaceus | Large Yellow Croaker L. crocea | Rainbow Trout O. mykiss | Zebrafish D. rerio | |
|---|---|---|---|---|---|---|---|---|---|
| TLRs | TLR3 | ✔[11] | ✔[19] | ✔[28] | ✔[33] | ✔[34] | ✔[42] | ✔[48] | ✔[53] |
| TLR7 | ✔[12] | ✔[20] | ✔[29] | ✔[33] | ✔[35] | ✔[43] | ✔[49] | ✔[53] | |
| TLR8 | ✔[13] | ✔[21] | ✔[30] | ✔[33] | ✔[36] ** | ✔[43] | ✔[49] | ✔[53] | |
| TLR9 | ✔[14] | ✔[22] | ✔[29] | ✔[33] | ✔[37] | ✔[44] | ✔[50] | ✔[53] | |
| TLR13 | ✔[11] | ✔[30] * | |||||||
| TLR21 | ✔[15] | ✔[23] | ✔[31] | ✔[33] | ✔[36] ** | ✔[53] | |||
| TLR22 | ✔[16] | ✔[24] | ✔[29] | ✔[33] | ✔[38] | ✔[45] | ✔[16] | ✔[53] | |
| RLRs | RIG-I | ✔[17] | ✔[25] | ✔[32] | ✔[54] | ||||
| MDA5 | ✔[18] | ✔[26] | ✔[32] | ✔[39] | ✔[46] | ✔[51] | ✔[55] | ||
| LGP2 | ✔[27] | ✔[32] | ✔[40] | ✔[51] | ✔[55] | ||||
| SRAs | SCARA3 | ✔[47] | |||||||
| SCARA4 | ✔[52] | ✔[56] | |||||||
| SCARA5 | ✔[47] | ||||||||
| MARCO | ✔[47] | ✔[57] | |||||||
| CDS | DDX41 | ✔[41] | |||||||
2.1. Toll-Like Receptors (TLRs)
2.1.1. Endosomal TLRs
2.1.1.1. TLR3
2.1.1.2. TLR7/8
2.1.1.3. TLR9
2.1.1.4. TLR13
2.1.1.5. TLR21
2.1.2. Surface TLRs
TLR22
2.1.3. Downstream Signaling: MyD88 and TICAM-1
2.2. RIG-I-Like Receptors (RLRs)
2.2.1. RIG-I and MDA5
2.2.2. LGP2
2.2.3. Downstream Signaling: IPS-1
2.2.4. Downstream Signaling: IRF3/7
2.3. Cytosolic DNA Sensors (CDSs)
Downstream Signaling: STING
2.4. Class A Scavenger Receptors (SRAs)
Downstream Signaling
2.5. The Type I Interferon Response
2.6. The Inflammatory Response
3. Future Directions
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Nellimarla, S.; Mossman, K.L. Extracellular dsRNA: Its function and mechanism of cellular uptake. J. Interferon Cytokine Res. 2014, 34, 419–426. [Google Scholar] [CrossRef]
- Jensen, S.; Thomsen, A.R. Sensing of RNA viruses: A review of innate immune receptors involved in recognizing RNA virus invasion. J. Virol. 2012, 86, 2900–2910. [Google Scholar] [CrossRef]
- Loo, Y.M.; Gale, M. Immune signaling by RIG-I-like receptors. Immunity 2011, 34, 680–692. [Google Scholar] [CrossRef]
- Jacobs, B.L.; Langland, J.O. When two strands are better than one: The mediators and modulators of the cellular responses to double-stranded RNA. Virology 1996, 219, 339–349. [Google Scholar] [CrossRef]
- DeWitte-Orr, S.J.; Mossman, K.L.; Mossman, K. The antiviral effects of extracellular dsRNA. In Viruses and Interferon: Current Research; Caister Academic Press: Norfolk, UK, 2011. [Google Scholar]
- Horan, K.A.; Hansen, K.; Jakobsen, M.R.; Holm, C.K.; Søby, S.; Unterholzner, L.; Thompson, M.; West, J.A.; Iversen, M.B.; Rasmussen, S.B. Proteasomal degradation of herpes simplex virus capsids in macrophages releases DNA to the cytosol for recognition by DNA sensors. J. Immunol. 2013, 190, 2311–2319. [Google Scholar] [CrossRef]
- Chiu, Y.-H.; MacMillan, J.B.; Chen, Z.J. RNA polymerase III detects cytosolic DNA and induces type I interferons through the RIG-I pathway. Cell 2009, 138, 576–591. [Google Scholar] [CrossRef]
- Whelan, F.J.; Meehan, C.J.; Golding, G.B.; McConkey, B.J.; Bowdish, D.M. The evolution of the class A scavenger receptors. BMC Evol. Biol. 2012. [Google Scholar] [CrossRef]
- Botos, I.; Segal, D.M.; Davies, D.R. The structural biology of Toll-like receptors. Structure 2011, 19, 447–459. [Google Scholar] [CrossRef]
- Bowie, A.G. Innate sensing of bacterial cyclic dinucleotides: More than just STING. Nat. Immunol. 2012, 13, 1137–1139. [Google Scholar] [CrossRef]
- Arnemo, M.; Kavaliauskis, A.; Gjøen, T. Effects of TLR agonists and viral infection on cytokine and TLR expression in atlantic salmon (Salmo salar). Dev. Comp. Immunol. 2014, 46, 139–145. [Google Scholar] [CrossRef]
- Lee, P.; Zou, J.; Holland, J.; Martin, S.; Kanellos, T.; Secombes, C. Identification and characterization of TLR7, TLR8a2, TLR8b1 and TLR8b2 genes in Atlantic salmon (Salmo salar). Dev. Comp. Immunol. 2013, 41, 295–305. [Google Scholar] [CrossRef]
- Skjaeveland, I.; Iliev, D.B.; Strandskog, G.; Jørgensen, J.B. Identification and characterization of TLR8 and MyD88 homologs in Atlantic salmon (Salmo salar). Dev. Comp. Immunol. 2009, 33, 1011–1017. [Google Scholar] [CrossRef]
- Skjaeveland, I.; Iliev, D.B.; Zou, J.; Jørgensen, T.; Jørgensen, J.B. A TLR9 homolog that is up-regulated by IFN-γ in Atlantic salmon (Salmo salar). Dev. Comp. Immunol. 2008, 32, 603–607. [Google Scholar] [CrossRef]
- Lee, P.; Zou, J.; Holland, J.; Martin, S.; Collet, B.; Kanellos, T.; Secombes, C. Identification and characterisation of TLR18–21 genes in Atlantic salmon (Salmo salar). Fish Shellfish Immunol. 2014, 41, 549–559. [Google Scholar] [CrossRef]
- Rebl, A.; Siegl, E.; Köllner, B.; Fischer, U.; Seyfert, H.-M. Characterization of twin toll-like receptors from rainbow trout (Oncorhynchus mykiss): Evolutionary relationship and induced expression by Aeromonas salmonicida salmonicida. Dev. Comp. Immunol. 2007, 31, 499–510. [Google Scholar] [CrossRef]
- Biacchesi, S.; LeBerre, M.; Lamoureux, A.; Louise, Y.; Lauret, E.; Boudinot, P.; Bremont, M. Mitochondrial antiviral signaling protein plays a major role in induction of the fish innate immune response against RNA and DNA viruses. J. Virol. 2009, 83, 7815–7827. [Google Scholar] [CrossRef]
- Sun, B.; Robertsen, B.; Wang, Z.; Liu, B. Identification of an Atlantic salmon IFN multigene cluster encoding three IFN subtypes with very different expression properties. Dev. Comp. Immunol. 2009, 33, 547–558. [Google Scholar] [CrossRef]
- Su, J.; Zhang, R.; Dong, J.; Yang, C. Evaluation of internal control genes for qRT-PCR normalization in tissues and cell culture for antiviral studies of grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2011, 30, 830–835. [Google Scholar] [CrossRef]
- Yang, C.; Su, J.; Zhang, R.; Peng, L.; Li, Q. Identification and expression profiles of grass carp Ctenopharyngodon idella TLR7 in responses to double-stranded RNA and virus infection. J. Fish Biol. 2012, 80, 2605–2622. [Google Scholar] [CrossRef]
- Su, J.; Su, J.; Shang, X.; Wan, Q.; Chen, X.; Rao, Y. SNP detection of TLR gene, association study with susceptibility/resistance to GCRV and regulation on mRNA expression in grass carp, Ctenopharyngodon idella. Fish Shellfish Immunol. 2015, 43, 1–12. [Google Scholar] [CrossRef]
- Yang, C.-R.; Su, J.-G.; Peng, L.-M.; Dong, J. Cloning and characterization of grass carp (Ctenopharyngodon idella) toll-like receptor 9. J. Fish. China 2011, 35, 641–649. [Google Scholar]
- Wang, W.; Shen, Y.; Pandit, N.P.; Li, J. Molecular cloning, characterization and immunological response analysis of toll-like receptor 21 (TLR21) gene in grass carp, Ctenopharyngodon idella. Dev. Comp. Immunol. 2013, 40, 227–231. [Google Scholar] [CrossRef]
- Lv, J.; Huang, R.; Li, H.; Luo, D.; Liao, L.; Zhu, Z.; Wang, Y. Cloning and characterization of the grass carp (Ctenopharyngodon idella) toll-like receptor 22 gene, a fish-specific gene. Fish Shellfish Immunol. 2012, 32, 1022–1031. [Google Scholar] [CrossRef]
- Yang, C.; Su, J.; Huang, T.; Zhang, R.; Peng, L. Identification of a retinoic acid-inducible gene I from grass carp (Ctenopharyngodon idella) and expression analysis in vivo and in vitro. Fish Shellfish Immunol. 2011, 30, 936–943. [Google Scholar] [CrossRef]
- Su, J.; Huang, T.; Dong, J.; Heng, J.; Zhang, R.; Peng, L. Molecular cloning and immune responsive expression of MDA5 gene, a pivotal member of the RLR gene family from grass carp Ctenopharyngodon idella. Fish Shellfish Immunol. 2010, 28, 712–718. [Google Scholar] [CrossRef]
- Huang, T.; Su, J.; Heng, J.; Dong, J.; Zhang, R.; Zhu, H. Identification and expression profiling analysis of grass carp Ctenopharyngodon idella LGP2 cDNA. Fish Shellfish Immunol. 2010, 29, 349–355. [Google Scholar] [CrossRef]
- Bilodeau, A.L.; Waldbieser, G.C. Activation of TLR3 and TLR5 in channel catfish exposed to virulent Edwardsiella ictaluri. Dev. Comp. Immunol. 2005, 29, 713–721. [Google Scholar] [CrossRef]
- Quiniou, S.M.; Boudinot, P.; Bengtén, E. Comprehensive survey and genomic characterization of toll-like receptors (TLRs) in channel catfish, Ictalurus punctatus: Identification of novel fish TLRs. Immunogenetics 2013, 65, 511–530. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, Y.; Zhou, Z.; Waldbieser, G.; Sun, F.; Lu, J.; Zhang, J.; Jiang, Y.; Zhang, H.; Wang, X. Efficient assembly and annotation of the transcriptome of catfish by RNA-seq analysis of a doubled haploid homozygote. BMC Genomics 2012. [Google Scholar] [CrossRef]
- Baoprasertkul, P.; Xu, P.; Peatman, E.; Kucuktas, H.; Liu, Z. Divergent toll-like receptors in catfish (Ictalurus punctatus): TLR5S, TLR20, TLR21. Fish Shellfish Immunol. 2007, 23, 1218–1230. [Google Scholar] [CrossRef]
- Rajendran, K.; Zhang, J.; Liu, S.; Peatman, E.; Kucuktas, H.; Wang, X.; Liu, H.; Wood, T.; Terhune, J.; Liu, Z. Pathogen recognition receptors in channel catfish: II. Identification, phylogeny and expression of retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs). Dev. Comp. Immunol. 2012, 37, 381–389. [Google Scholar] [CrossRef]
- Oshiumi, H.; Tsujita, T.; Shida, K.; Matsumoto, M.; Ikeo, K.; Seya, T. Prediction of the prototype of the human toll-like receptor gene family from the pufferfish, Fugu rubripes, genome. Immunogenetics 2003, 54, 791–800. [Google Scholar]
- Hwang, S.D.; Ohtani, M.; Hikima, J.-I.; Jung, T.S.; Kondo, H.; Hirono, I.; Aoki, T. Molecular cloning and characterization of toll-like receptor 3 in Japanese flounder, Paralichthys olivaceus. Dev. Comp. Immunol. 2012, 37, 87–96. [Google Scholar] [CrossRef]
- Avunje, S.; Kim, W.-S.; Park, C.-S.; Oh, M.-J.; Jung, S.-J. Toll-like receptors and interferon associated immune factors in viral haemorrhagic septicaemia virus-infected olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2011, 31, 407–414. [Google Scholar] [CrossRef]
- Hwang, S.D.; Fuji, K.; Takano, T.; Sakamoto, T.; Kondo, H.; Hirono, I.; Aoki, T. Linkage mapping of toll-like receptors (TLRs) in Japanese flounder, Paralichthys olivaceus. Mar. Biotechnol. 2011, 13, 1086–1091. [Google Scholar] [CrossRef]
- Takano, T.; Kondo, H.; Hirono, I.; Endo, M.; Saito-Taki, T.; Aoki, T. Molecular cloning and characterization of toll-like receptor 9 in Japanese flounder, Paralichthys olivaceus. Mol. Immunol. 2007, 44, 1845–1853. [Google Scholar] [CrossRef]
- Hirono, I.; Takami, M.; Miyata, M.; Miyazaki, T.; Han, H.-J.; Takano, T.; Endo, M.; Aoki, T. Characterization of gene structure and expression of two toll-like receptors from Japanese flounder, Paralichthys olivaceus. Immunogenetics 2004, 56, 38–46. [Google Scholar]
- Ohtani, M.; Hikima, J.-I.; Kondo, H.; Hirono, I.; Jung, T.-S.; Aoki, T. Characterization and antiviral function of a cytosolic sensor gene, MDA5, in Japanese flounder, Paralichthys olivaceus. Dev. Comp. Immunol. 2011, 35, 554–562. [Google Scholar] [CrossRef]
- Ohtani, M.; Hikima, J.-I.; Kondo, H.; Hirono, I.; Jung, T.-S.; Aoki, T. Evolutional conservation of molecular structure and antiviral function of a viral RNA receptor, LGP2, in Japanese flounder, Paralichthys olivaceus. J. Immunol. 2010, 185, 7507–7517. [Google Scholar] [CrossRef]
- Quynh, N.T.; Hikima, J.-I.; Kim, Y.-R.; Fagutao, F.F.; Kim, M.S.; Aoki, T.; Jung, T.S. The cytosolic sensor, DDX41, activates antiviral and inflammatory immunity in response to stimulation with double-stranded DNA adherent cells of the olive flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2015, 44, 576–583. [Google Scholar] [CrossRef]
- Huang, X.-N.; Wang, Z.-Y.; Yao, C.-L. Characterization of toll-like receptor 3 gene in large yellow croaker, Pseudosciaena crocea. Fish Shellfish Immunol. 2011, 31, 98–106. [Google Scholar] [CrossRef]
- Qian, T.; Wang, K.; Mu, Y.; Ao, J.; Chen, X. Molecular characterization and expression analysis of TLR 7 and TLR 8 homologs in large yellow croaker (Pseudosciaena crocea). Fish Shellfish Immunol. 2013, 35, 671–679. [Google Scholar] [CrossRef]
- Yao, C.-L.; Kong, P.; Wang, Z.-Y.; Ji, P.-F.; Cai, M.-Y.; Liu, X.-D.; Han, X.-Z. Cloning and expression analysis of two alternative splicing toll-like receptor 9 isoforms A and B in large yellow croaker, Pseudosciaena crocea. Fish Shellfish Immunol. 2008, 25, 648–656. [Google Scholar] [CrossRef]
- Xiao, X.; Qin, Q.; Chen, X. Molecular characterization of a toll-like receptor 22 homologue in large yellow croaker (Pseudosciaena crocea) and promoter activity analysis of its 5'-flanking sequence. Fish Shellfish Immunol. 2011, 30, 224–233. [Google Scholar] [CrossRef]
- Wang, X.; Wang, K.; Nie, P.; Chen, X.; Ao, J. Establishment and characterization of a head kidney cell line from large yellow croaker Pseudosciaena crocea. J. Fish Biol. 2014, 84, 1551–1561. [Google Scholar] [CrossRef]
- He, J.; Liu, H.; Wu, C. Identification of SCARA3, SCARA5 and MARCO of class A scavenger receptor-like family in Pseudosciaena crocea. Fish Shellfish Immunol. 2014, 41, 238–249. [Google Scholar] [CrossRef]
- Rodriguez, M.; Wiens, G.; Purcell, M.; Palti, Y. Characterization of toll-like receptor 3 gene in rainbow trout (Oncorhynchus mykiss). Immunogenetics 2005, 57, 510–519. [Google Scholar] [CrossRef]
- Palti, Y.; Gahr, S.A.; Purcell, M.K.; Hadidi, S.; Rexroad, C.E.; Wiens, G.D. Identification, characterization and genetic mapping of TLR7, TLR8a1 and TLR8a2 genes in rainbow trout (Oncorhynchus mykiss). Dev. Comp. Immunol. 2010, 34, 219–233. [Google Scholar] [CrossRef]
- Ortega-Villaizan, M.; Chico, V.; Falco, A.; Perez, L.; Coll, J.; Estepa, A. The rainbow trout TLR9 gene and its role in the immune responses elicited by a plasmid encoding the glycoprotein G of the viral haemorrhagic septicaemia rhabdovirus (VHSV). Mol. Immunol. 2009, 46, 1710–1717. [Google Scholar] [CrossRef]
- Chang, M.; Collet, B.; Nie, P.; Lester, K.; Campbell, S.; Secombes, C.J.; Zou, J. Expression and functional characterization of the RIG-I-like receptors MDA5 and LGP2 in Rainbow trout (Oncorhynchus mykiss). J. Virol. 2011, 85, 8403–8412. [Google Scholar] [CrossRef]
- Poynter, S.J.; Weleff, J.; Soares, A.B.; Dewitte-Orr, S.J. Class-A scavenger receptor function and expression in the rainbow trout (Oncorhynchus mykiss) epithelial cell lines RTgutGC and RTgill-W1. Fish Shellfish Immunol. 2015, 44, 138–146. [Google Scholar] [CrossRef]
- Meijer, A.H.; Krens, S.G.; Rodriguez, I.A.M.; He, S.; Bitter, W.; Snaar-Jagalska, B.E.; Spaink, H.P. Expression analysis of the toll-like receptor and TIR domain adaptor families of zebrafish. Mol. Immunol. 2004, 40, 773–783. [Google Scholar] [CrossRef]
- Nie, L.; Zhang, Y.-S.; Dong, W.-R.; Xiang, L.-X.; Shao, J.-Z. Involvement of zebrafish RIG-I in NF-κB and IFN signaling pathways: Insights into functional conservation of RIG-I in antiviral innate immunity. Dev. Comp. Immunol. 2015, 48, 95–101. [Google Scholar] [CrossRef]
- Chen, W.Q.; Hu, Y.W.; Zou, P.F.; Ren, S.S.; Nie, P.; Chang, M.X. MAVS splicing variants contribute to the induction of interferon and interferon-stimulated genes mediated by RIG-I-like receptors. Dev. Comp. Immunol. 2015, 49, 19–30. [Google Scholar] [CrossRef]
- Fukuda, M.; Ohtani, K.; Jang, S.-J.; Yoshizaki, T.; Mori, K.-I.; Motomura, W.; Yoshida, I.; Suzuki, Y.; Kohgo, Y.; Wakamiya, N. Molecular cloning and functional analysis of scavenger receptor zebrafish Cl-P1. Biochim. Biophys. Acta 2011, 1810, 1150–1159. [Google Scholar] [CrossRef]
- Benard, E.L.; Roobol, S.J.; Spaink, H.P.; Meijer, A.H. Phagocytosis of mycobacteria by zebrafish macrophages is dependent on the scavenger receptor marco, a key control factor of pro-inflammatory signalling. Dev. Comp. Immunol. 2014, 47, 223–233. [Google Scholar] [CrossRef]
- Bell, J.K.; Mullen, G.E.; Leifer, C.A.; Mazzoni, A.; Davies, D.R.; Segal, D.M. Leucine-rich repeats and pathogen recognition in toll-like receptors. Trends Immunol. 2003, 24, 528–533. [Google Scholar] [CrossRef]
- Palti, Y. Toll-like receptors in bony fish: From genomics to function. Dev. Comp. Immunol. 2011, 35, 1263–1272. [Google Scholar] [CrossRef]
- Zhang, J.; Kong, X.; Zhou, C.; Li, L.; Nie, G.; Li, X. Toll-like receptor recognition of bacteria in fish: Ligand specificity and signal pathways. Fish Shellfish Immunol. 2014, 41, 380–388. [Google Scholar] [CrossRef]
- Moresco, E.M.Y.; LaVine, D.; Beutler, B. Toll-like receptors. Curr. Biol. 2011, 21, R488–R493. [Google Scholar] [CrossRef]
- Li, X.-D.; Chen, Z.J. Sequence specific detection of bacterial 23S ribosomal RNA by TLR13. Elife 2012. [Google Scholar] [CrossRef]
- Yeh, D.-W.; Liu, Y.-L.; Lo, Y.-C.; Yuh, C.-H.; Yu, G.-Y.; Lo, J.-F.; Luo, Y.; Xiang, R.; Chuang, T.-H. Toll-like receptor 9 and 21 have different ligand recognition profiles and cooperatively mediate activity of CpG-oligodeoxynucleotides in zebrafish. PNAS 2013, 110, 20711–20716. [Google Scholar] [CrossRef]
- Matsuo, A.; Oshiumi, H.; Tsujita, T.; Mitani, H.; Kasai, H.; Yoshimizu, M.; Matsumoto, M.; Seya, T. Teleost TLR22 recognizes RNA duplex to induce IFN and protect cells from birnaviruses. J. Immunol. 2008, 181, 3474–3485. [Google Scholar] [CrossRef]
- Barton, G.M.; Kagan, J.C. A cell biological view of Toll-like receptor function: Regulation through compartmentalization. Nat. Rev. Immunol. 2009, 9, 535–542. [Google Scholar] [CrossRef]
- Matsumoto, M.; Kikkawa, S.; Kohase, M.; Miyake, K.; Seya, T. Establishment of a monoclonal antibody against human toll-like receptor 3 that blocks double-stranded RNA-mediated signaling. Biochem. Biophys. Res. Commun. 2002, 293, 1364–1369. [Google Scholar] [CrossRef]
- O’Neill, L.A.; Golenbock, D.; Bowie, A.G. The history of Toll-like receptors—Redefining innate immunity. Nat. Rev. Immunol. 2013, 13, 453–460. [Google Scholar] [CrossRef]
- Leifer, C.A.; Kennedy, M.N.; Mazzoni, A.; Lee, C.; Kruhlak, M.J.; Segal, D.M. TLR9 is localized in the endoplasmic reticulum prior to stimulation. J. Immunol. 2004, 173, 1179–1183. [Google Scholar] [CrossRef]
- Brownlie, R.; Zhu, J.; Allan, B.; Mutwiri, G.K.; Babiuk, L.A.; Potter, A.; Griebel, P. Chicken TLR21 acts as a functional homologue to mammalian TLR9 in the recognition of CpG oligodeoxynucleotides. Mol. Immunol. 2009, 46, 3163–3170. [Google Scholar] [CrossRef]
- Jørgensen, J.; Zou, J.; Johansen, A.; Secombes, C. Immunostimulatory CpG oligodeoxynucleotides stimulate expression of IL-1β and interferon-like cytokines in rainbow trout macrophages via a chloroquine-sensitive mechanism. Fish Shellfish Immunol. 2001, 11, 673–682. [Google Scholar] [CrossRef]
- Zhou, Z.-X.; Sun, L. Immune effects of R848: Evidences that suggest an essential role of TLR7/8-induced, MyD88-and NF-κB-dependent signaling in the antiviral immunity of Japanese flounder (Paralichthys olivaceus). Dev. Comp. Immunol. 2015, 49, 113–120. [Google Scholar] [CrossRef]
- DeWitte-Orr, S.J.; Mossman, K.L. dsRNA and the innate antiviral immune response. Future Virol. 2010, 5, 325–341. [Google Scholar] [CrossRef]
- Józefowski, S.; Sulahian, T.H.; Arredouani, M.; Kobzik, L. Role of scavenger receptor MARCO in macrophage responses to CpG oligodeoxynucleotides. J. Leukocyte Biol. 2006, 80, 870–879. [Google Scholar] [CrossRef]
- Delgado, M.; Deretic, V. Toll-like receptors in control of immunological autophagy. Cell Death Differ. 2009, 16, 976–983. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014. [Google Scholar] [CrossRef]
- Samanta, M.; Basu, M.; Swain, B.; Panda, P.; Jayasankar, P. Molecular cloning and characterization of toll-like receptor 3, and inductive expression analysis of type I IFN, Mx and pro-inflammatory cytokines in the Indian carp, rohu (Labeo rohita). Mol. Biol. Rep. 2013, 40, 225–235. [Google Scholar] [CrossRef]
- Phelan, P.E.; Mellon, M.T.; Kim, C.H. Functional characterization of full-length TLR3, IRAK-4, and TRAF6 in zebrafish (Danio rerio). Mol. Immunol. 2005, 42, 1057–1071. [Google Scholar] [CrossRef]
- Lin, K.; Ge, H.; Lin, Q.; Wu, J.; He, L.; Fang, Q.; Zhou, C.; Sun, M.; Huang, Z. Molecular characterization and functional analysis of toll-like receptor 3 gene in orange-spotted grouper (Epinephelus coioides). Gene 2013, 527, 174–182. [Google Scholar] [CrossRef]
- Zhou, Z.-X.; Zhang, B.-C.; Sun, L. Poly (I:C) induces antiviral immune responses in Japanese flounder (Paralichthys olivaceus) that require TLR3 and MDA5 and is negatively regulated by MyD88. PLoS ONE 2014, 9, e112918. [Google Scholar] [CrossRef]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef]
- Gantier, M.P.; Tong, S.; Behlke, M.A.; Xu, D.; Phipps, S.; Foster, P.S.; Williams, B.R. TLR7 is involved in sequence-specific sensing of single-stranded RNAs in human macrophages. J. Immunol. 2008, 180, 2117–2124. [Google Scholar] [CrossRef]
- Hemmi, H.; Kaisho, T.; Takeuchi, O.; Sato, S.; Sanjo, H.; Hoshino, K.; Horiuchi, T.; Tomizawa, H.; Takeda, K.; Akira, S. Small anti-viral compounds activate immune cells via the TLR7 MyD88-dependent signaling pathway. Nat. Immunol. 2002, 3, 196–200. [Google Scholar] [CrossRef]
- Gorden, K.B.; Gorski, K.S.; Gibson, S.J.; Kedl, R.M.; Kieper, W.C.; Qiu, X.; Tomai, M.A.; Alkan, S.S.; Vasilakos, J.P. Synthetic TLR agonists reveal functional differences between human TLR7 and TLR8. J. Immunol. 2005, 174, 1259–1268. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Q.; Yang, C.; Rao, Y.; Li, Q.; Wan, Q.; Peng, L.; Wu, S.; Su, J. Identification, expression profiling of a grass carp TLR8 and its inhibition leading to the resistance to reovirus in CIK cells. Dev. Comp. Immunol. 2013, 41, 82–93. [Google Scholar] [CrossRef]
- Lund, J.; Sato, A.; Akira, S.; Medzhitov, R.; Iwasaki, A. Toll-like receptor 9-mediated recognition of herpes simplex virus-2 by plasmacytoid dendritic cells. J. Exp. Med. 2003, 198, 513–520. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. TLR signaling. Cell Death Differ. 2006, 13, 816–825. [Google Scholar] [CrossRef]
- Roberts, T.L.; Sweet, M.J.; Hume, D.A.; Stacey, K.J. Cutting edge: Species-specific TLR9-mediated recognition of CpG and non-CpG phosphorothioate-modified oligonucleotides. J. Immunol. 2005, 174, 605–608. [Google Scholar] [CrossRef]
- Zelenay, S.; Elías, F.; Fló, J. Immunostimulatory effects of plasmid DNA and synthetic oligodeoxynucleotides. Eur. J. Immunol. 2003, 33, 1382–1392. [Google Scholar] [CrossRef]
- Cuesta, A.; Esteban, M.; Meseguer, J. The expression profile of TLR9 mRNA and CpG ODNs immunostimulatory actions in the teleost gilthead seabream points to a major role of lymphocytes. Cell. Mol. Life Sci. 2008, 65, 2091–2104. [Google Scholar] [CrossRef]
- Byadgi, O.; Puteri, D.; Lee, Y.-H.; Lee, J.-W.; Cheng, T.-C. Identification and expression analysis of cobia (Rachycentron canadum) toll-like receptor 9 gene. Fish Shellfish Immunol. 2014, 36, 417–427. [Google Scholar] [CrossRef]
- Strandskog, G.; Villoing, S.; Iliev, D.B.; Thim, H.L.; Christie, K.E.; Jørgensen, J.B. Formulations combining CpG containing oliogonucleotides and poly I:C enhance the magnitude of immune responses and protection against pancreas disease in Atlantic salmon. Dev. Comp. Immunol. 2011, 35, 1116–1127. [Google Scholar] [CrossRef]
- Jørgensen, J.B.; Johansen, A.; Stenersen, B.; Sommer, A.-I. CpG oligodeoxynucleotides and plasmid DNA stimulate Atlantic salmon (Salmo salar L.) leucocytes to produce supernatants with antiviral activity. Dev. Comp. Immunol. 2001, 25, 313–321. [Google Scholar] [CrossRef]
- Iliev, D.B.; Skjaeveland, I.; Jørgensen, J.B. CpG oligonucleotides bind TLR9 and RRM-containing proteins in Atlantic salmon (Salmo salar). BMC Immunol. 2013. [Google Scholar] [CrossRef]
- Broz, P.; Monack, D.M. Newly described pattern recognition receptors team up against intracellular pathogens. Nat. Rev. Immunol. 2013, 13, 551–565. [Google Scholar] [CrossRef]
- Shi, Z.; Cai, Z.; Sanchez, A.; Zhang, T.; Wen, S.; Wang, J.; Zhang, D. A novel Toll-like receptor that recognizes vesicular stomatitis virus. J. Biol. Chem. 2011, 286, 4517–4524. [Google Scholar] [CrossRef]
- Leong, J.S.; Jantzen, S.G.; von Schalburg, K.R.; Cooper, G.A.; Messmer, A.M.; Liao, N.Y.; Munro, S.; Moore, R.; Holt, R.A.; Jones, S.J. Salmo salar and Esox lucius full-length cDNA sequences reveal changes in evolutionary pressures on a post-tetraploidization genome. BMC Genomics 2010. [Google Scholar] [CrossRef]
- Keestra, A.M.; de Zoete, M.R.; Bouwman, L.I.; van Putten, J.P. Chicken TLR21 is an innate CpG DNA receptor distinct from mammalian TLR9. J. Immunol. 2010, 185, 460–467. [Google Scholar] [CrossRef]
- Sundaram, A.Y.; Kiron, V.; Dopazo, J.; Fernandes, J.M. Diversification of the expanded teleost-specific toll-like receptor family in Atlantic cod, Gadus morhua. BMC Evol. Biol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Priyathilaka, T.T.; Elvitigala, D.A.S.; Whang, I.; Lim, B.-S.; Jeong, H.-B.; Yeo, S.-Y.; Choi, C.Y.; Lee, J. Molecular characterization and transcriptional analysis of non-mammalian type toll like receptor (TLR21) from Rock bream (Oplegnathus fasciatus). Gene 2014, 553, 105–116. [Google Scholar] [CrossRef]
- Liu, L.; Botos, I.; Wang, Y.; Leonard, J.N.; Shiloach, J.; Segal, D.M.; Davies, D.R. Structural basis of toll-like receptor 3 signaling with double-stranded RNA. Science 2008, 320, 379–381. [Google Scholar] [CrossRef]
- Hu, G.-B.; Zhang, S.-F.; Yang, X.; Liu, D.-H.; Liu, Q.-M.; Zhang, S.-C. Cloning and expression analysis of a toll-like receptor 22 (TLR22) gene from turbot, Scophthalmus maximus. Fish Shellfish Immunol. 2015, 44, 399–409. [Google Scholar] [CrossRef]
- Muñoz, I.; Sepulcre, M.P.; Meseguer, J.; Mulero, V. Toll-like receptor 22 of gilthead seabream, Sparus aurata: Molecular cloning, expression profiles and post-transcriptional regulation. Dev. Comp. Immunol. 2014, 44, 173–179. [Google Scholar] [CrossRef]
- Samanta, M.; Swain, B.; Basu, M.; Mahapatra, G.; Sahoo, B.R.; Paichha, M.; Lenka, S.S.; Jayasankar, P. Toll-like receptor 22 in Labeo rohita: Molecular cloning, characterization, 3D modeling, and expression analysis following ligands stimulation and bacterial infection. Appl. Biochem. Biotech. 2014, 174, 309–327. [Google Scholar] [CrossRef]
- Su, J.; Huang, T.; Yang, C.; Zhang, R. Molecular cloning, characterization and expression analysis of interferon-β promoter stimulator 1 (IPS-1) gene from grass carp Ctenopharyngodon idella. Fish Shellfish Immunol. 2011, 30, 317–323. [Google Scholar] [CrossRef]
- Takano, T.; Kondo, H.; Hirono, I.; Saito-Taki, T.; Endo, M.; Aoki, T. Identification and characterization of a myeloid differentiation factor 88 (MyD88) cDNA and gene in Japanese flounder, Paralichthys olivaceus. Dev. Comp. Immunol. 2006, 30, 807–816. [Google Scholar] [CrossRef]
- Yao, C.-L.; Kong, P.; Wang, Z.-Y.; Ji, P.-F.; Liu, X.-D.; Cai, M.-Y.; Han, X.-Z. Molecular cloning and expression of MyD88 in large yellow croaker, Pseudosciaena crocea. Fish Shellfish Immunol. 2009, 26, 249–255. [Google Scholar] [CrossRef]
- Rebl, A.; Goldammer, T.; Fischer, U.; Köllner, B.; Seyfert, H.-M. Characterization of two key molecules of teleost innate immunity from rainbow trout (Oncorhynchus mykiss): MyD88 and SAA. Vet. Immunol. Immunopathol. 2009, 131, 122–126. [Google Scholar] [CrossRef]
- Van der Sar, A.M.; Stockhammer, O.W.; van der Laan, C.; Spaink, H.P.; Bitter, W.; Meijer, A.H. MyD88 innate immune function in a zebrafish embryo infection model. Infect. Immun. 2006, 74, 2436–2441. [Google Scholar] [CrossRef]
- Foley, J.F. TRAF6 phosphorylation inhibits inflammation. Sci. Signal. 2015. [Google Scholar] [CrossRef]
- Li, Y.-W.; Mo, X.-B.; Zhou, L.; Li, X.; Dan, X.-M.; Luo, X.-C.; Li, A.-X. Identification of IRAK-4 in grouper (Epinephelus coioides) that impairs MyD88-dependent NF-κB activation. Dev. Comp. Immunol. 2014, 45, 190–197. [Google Scholar] [CrossRef]
- Basu, M.; Swain, B.; Sahoo, B.R.; Maiti, N.K.; Samanta, M. Induction of toll-like receptor (TLR) 2, and MyD88-dependent TLR-signaling in response to ligand stimulation and bacterial infections in the Indian major carp, mrigal (Cirrhinus mrigala). Mol. Biol. Rep. 2012, 39, 6015–6028. [Google Scholar] [CrossRef]
- Iliev, D.B.; Sobhkhez, M.; Fremmerlid, K.; Jorgensen, J.B. MyD88 interacts with interferon regulatory factor (IRF)3 and IRF7 in Atlantic salmon (Salmo salar): Transgenic SsMyD88 modulates the IRF-induced type I interferon response and accumulates in aggresomes. J. Biol. Chem. 2011, 286, 42715–42724. [Google Scholar] [CrossRef]
- Baoprasertkul, P.; Peatman, E.; Somridhivej, B.; Liu, Z. Toll-like receptor 3 and TICAM genes in catfish: Species-specific expression profiles following infection with Edwardsiella ictaluri. Immunogenetics 2006, 58, 817–830. [Google Scholar] [CrossRef]
- Fan, S.; Chen, S.; Liu, Y.; Lin, Y.; Liu, H.; Guo, L.; Lin, B.; Huang, S.; Xu, A. Zebrafish TRIF, a Golgi-localized protein, participates in IFN induction and NF-κB activation. J. Immunol. 2008, 180, 5373–5383. [Google Scholar] [CrossRef]
- Zhang, Y.-B.; Gui, J.-F. Molecular regulation of interferon antiviral response in fish. Dev. Comp. Immunol. 2012, 38, 193–202. [Google Scholar] [CrossRef]
- Seya, T.; Matsumoto, M.; Ebihara, T.; Oshiumi, H. Functional evolution of the TICAM-1 pathway for extrinsic RNA sensing. Immunol. Rev. 2009, 227, 44–53. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. MDA5/RIG-I and virus recognition. Curr. Opin. Immunol. 2008, 20, 17–22. [Google Scholar] [CrossRef]
- Hornung, V.; Ellegast, J.; Kim, S.; Brzózka, K.; Jung, A.; Kato, H.; Poeck, H.; Akira, S.; Conzelmann, K.-K.; Schlee, M. 5'-triphosphate RNA is the ligand for RIG-I. Science 2006, 314, 994–997. [Google Scholar] [CrossRef]
- Goubau, D.; Schlee, M.; Deddouche, S.; Pruijssers, A.J.; Zillinger, T.; Goldeck, M.; Schuberth, C.; van der Veen, A.G.; Fujimura, T.; Rehwinkel, J. Antiviral immunity via RIG-I-mediated recognition of RNA bearing 5'-diphosphates. Nature 2014, 514, 372–375. [Google Scholar] [CrossRef]
- Kato, H.; Takahasi, K.; Fujita, T. RIG-I-like receptors: Cytoplasmic sensors for non-self RNA. Immunol. Rev. 2011, 243, 91–98. [Google Scholar] [CrossRef]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef]
- Zou, P.F.; Chang, M.X.; Li, Y.; Zhang, S.H.; Fu, J.P.; Chen, S.N.; Nie, P. Higher antiviral response of RIG-I through enhancing RIG-I/MAVs-mediated signaling by its long insertion variant in zebrafish. Fish Shellfish Immunol. 2015, 43, 13–24. [Google Scholar] [CrossRef]
- Chen, H.-Y.; Liu, W.; Wu, S.-Y.; Chiou, P.P.; Li, Y.-H.; Chen, Y.-C.; Lin, G.-H.; Lu, M.-W.; Wu, J.-L. RIG-I specifically mediates group II type I IFN activation in nervous necrosis virus infected zebrafish cells. Fish Shellfish Immunol. 2015, 43, 427–435. [Google Scholar] [CrossRef]
- Wang, W.; Asim, M.; Yi, L.; Hegazy, A.M.; Hu, X.; Zhou, Y.; Ai, T.; Lin, L. Abortive infection of snakehead fish vesiculovirus in ZF4 cells was associated with the RLRs pathway activation by viral replicative intermediates. Int. J. Mol. Sci. 2015, 16, 6235–6250. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Y.-B.; Liu, T.-K.; Shi, J.; Sun, F.; Gui, J.-F. Fish viperin exerts a conserved antiviral function through RLR-triggered IFN signaling pathway. Dev. Comp. Immunol. 2014, 47, 140–149. [Google Scholar] [CrossRef]
- Satoh, T.; Kato, H.; Kumagai, Y.; Yoneyama, M.; Sato, S.; Matsushita, K.; Tsujimura, T.; Fujita, T.; Akira, S.; Takeuchi, O. LGP2 is a positive regulator of RIG-I–and MDA5-mediated antiviral responses. PNAS 2010, 107, 1512–1517. [Google Scholar] [CrossRef]
- Yoneyama, M.; Fujita, T. Structural mechanism of RNA recognition by the RIG-I-like receptors. Immunity 2008, 29, 178–181. [Google Scholar] [CrossRef]
- Holland, J.; Bird, S.; Williamson, B.; Woudstra, C.; Mustafa, A.; Wang, T.; Zou, J.; Blaney, S.; Collet, B.; Secombes, C. Molecular characterization of IRF3 and IRF7 in rainbow trout, Oncorhynchus mykiss: Functional analysis and transcriptional modulation. Mol. Immunol. 2008, 46, 269–285. [Google Scholar] [CrossRef]
- Loo, Y.-M.; Fornek, J.; Crochet, N.; Bajwa, G.; Perwitasari, O.; Martinez-Sobrido, L.; Akira, S.; Gill, M.A.; García-Sastre, A.; Katze, M.G. Distinct RIG-I and MDA5 signaling by RNA viruses in innate immunity. J. Virol. 2008, 82, 335–345. [Google Scholar] [CrossRef]
- Bhat, A.; Paria, A.; Deepika, A.; Sreedharan, K.; Makesh, M.; Bedekar, M.K.; Purushothaman, C.; Rajendran, K. Molecular cloning, characterisation and expression analysis of melanoma differentiation associated gene 5 (MDA5) of green chromide, Etroplus suratensis. Gene 2015, 557, 172–181. [Google Scholar] [CrossRef]
- Zou, P.F.; Chang, M.X.; Xue, N.N.; Liu, X.Q.; Li, J.H.; Fu, J.P.; Chen, S.N.; Nie, P. Melanoma differentiation-associated gene 5 in zebrafish provoking higher interferon-promoter activity through signalling enhancing of its shorter splicing variant. Immunology 2014, 141, 192–202. [Google Scholar] [CrossRef]
- Yoneyama, M.; Onomoto, K.; Jogi, M.; Akaboshi, T.; Fujita, T. Viral RNA detection by RIG-I-like receptors. Curr. Opin. Immunol. 2015, 32, 48–53. [Google Scholar] [CrossRef]
- Bruns, A.M.; Leser, G.P.; Lamb, R.A.; Horvath, C.M. The innate immune sensor LGP2 activates antiviral signaling by regulating MDA5-RNA interaction and filament assembly. Mol. Cell 2014, 55, 771–781. [Google Scholar] [CrossRef]
- Venkataraman, T.; Valdes, M.; Elsby, R.; Kakuta, S.; Caceres, G.; Saijo, S.; Iwakura, Y.; Barber, G.N. Loss of DEXD/H box RNA helicase LGP2 manifests disparate antiviral responses. J. Immunol. 2007, 178, 6444–6455. [Google Scholar] [CrossRef]
- Chen, X.; Yang, C.; Su, J.; Rao, Y.; Gu, T. LGP2 plays extensive roles in modulating innate immune responses in Ctenopharyngodon idella kidney (cik) cells. Dev. Comp. Immunol. 2015, 49, 138–148. [Google Scholar] [CrossRef]
- Cao, X.; Chen, J.; Cao, Y.; Nie, G.; Wan, Q.; Wang, L.; Su, J. Identification and expression of the laboratory of genetics and physiology 2 gene in common carp Cyprinus carpio. J. Fish Biol. 2015, 86, 74–91. [Google Scholar] [CrossRef]
- Lauksund, S.; Svingerud, T.; Bergan, V.; Robertsen, B. Atlantic salmon IPS-1 mediates induction of IFNa1 and activation of NF-κB and localizes to mitochondria. Dev. Comp. Immunol. 2009, 33, 1196–1204. [Google Scholar] [CrossRef]
- Simora, R.M.C.; Ohtani, M.; Hikima, J.-I.; Kondo, H.; Hirono, I.; Jung, T.S.; Aoki, T. Molecular cloning and antiviral activity of IFN-β promoter stimulator-1 (IPS-1) gene in Japanese flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2010, 29, 979–986. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.-B.; Wu, M.; Wang, B.; Chen, C.; Gui, J.-F. Fish MAVs is involved in RLR pathway-mediated IFN response. Fish Shellfish Immunol. 2014, 41, 222–230. [Google Scholar] [CrossRef]
- Taniguchi, T.; Ogasawara, K.; Takaoka, A.; Tanaka, N. IRF family of transcription factors as regulators of host defense. Annu. Rev. Immunol. 2001, 19, 623–655. [Google Scholar] [CrossRef]
- Honda, K.; Takaoka, A.; Taniguchi, T. Type I interferon gene induction by the interferon regulatory factor family of transcription factors. Immunity 2006, 25, 349–360. [Google Scholar] [CrossRef]
- Sun, F.; Zhang, Y.-B.; Liu, T.-K.; Gan, L.; Yu, F.-F.; Liu, Y.; Gui, J.-F. Characterization of fish IRF3 as an IFN-inducible protein reveals evolving regulation of IFN response in vertebrates. J. Immunol. 2010, 185, 7573–7582. [Google Scholar] [CrossRef]
- Takaoka, A.; Wang, Z.; Choi, M.K.; Yanai, H.; Negishi, H.; Ban, T.; Lu, Y.; Miyagishi, M.; Kodama, T.; Honda, K. DAI (DLM-1ZBP1) is a cytosolic DNA sensor and an activator of innate immune response. Nature 2007, 448, 501–505. [Google Scholar] [CrossRef]
- Hornung, V.; Ablasser, A.; Charrel-Dennis, M.; Bauernfeind, F.; Horvath, G.; Caffrey, D.R.; Latz, E.; Fitzgerald, K.A. AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating inflammasome with ASC. Nature 2009, 458, 514–518. [Google Scholar] [CrossRef]
- Fernandes-Alnemri, T.; Yu, J.-W.; Datta, P.; Wu, J.; Alnemri, E.S. AIM2 activates the inflammasome and cell death in response to cytoplasmic DNA. Nature 2009, 458, 509–513. [Google Scholar] [CrossRef]
- Roberts, T.L.; Idris, A.; Dunn, J.A.; Kelly, G.M.; Burnton, C.M.; Hodgson, S.; Hardy, L.L.; Garceau, V.; Sweet, M.J.; Ross, I.L. HIN-200 proteins regulate caspase activation in response to foreign cytoplasmic DNA. Science 2009, 323, 1057–1060. [Google Scholar] [CrossRef]
- Ablasser, A.; Bauernfeind, F.; Hartmann, G.; Latz, E.; Fitzgerald, K.A.; Hornung, V. RIG-I-dependent sensing of poly (dA: dT) through the induction of an RNA polymerase III-transcribed RNA intermediate. Nat. Immunol. 2009, 10, 1065–1072. [Google Scholar] [CrossRef]
- Yang, P.; An, H.; Liu, X.; Wen, M.; Zheng, Y.; Rui, Y.; Cao, X. The cytosolic nucleic acid sensor LRRFIP1 mediates the production of type I interferon via a [beta]-catenin-dependent pathway. Nat. Immunol. 2010, 11, 487–494. [Google Scholar] [CrossRef]
- Zhang, Z.; Yuan, B.; Bao, M.; Lu, N.; Kim, T.; Liu, Y.-J. The helicase DDX41 senses intracellular DNA mediated by the adaptor STING in dendritic cells. Nat. Immunol. 2011, 12, 959–965. [Google Scholar] [CrossRef]
- Sun, L.; Wu, J.; Du, F.; Chen, X.; Chen, Z.J. Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 2013, 339, 786–791. [Google Scholar] [CrossRef]
- Unterholzner, L.; Keating, S.E.; Baran, M.; Horan, K.A.; Jensen, S.B.; Sharma, S.; Sirois, C.M.; Jin, T.; Latz, E.; Xiao, T.S. IFI16 is an innate immune sensor for intracellular DNA. Nat. Immunol. 2010, 11, 997–1004. [Google Scholar] [CrossRef]
- Kerur, N.; Veettil, M.V.; Sharma-Walia, N.; Bottero, V.; Sadagopan, S.; Otageri, P.; Chandran, B. IFI16 acts as a nuclear pathogen sensor to induce the inflammasome in response to Kaposi Sarcoma-associated herpesvirus infection. Cell Host Microbe 2011, 9, 363–375. [Google Scholar] [CrossRef]
- DeFilippis, V.R.; Alvarado, D.; Sali, T.; Rothenburg, S.; Früh, K. Human cytomegalovirus induces the interferon response via the DNA sensor ZBP1. J. Virol. 2010, 84, 585–598. [Google Scholar] [CrossRef]
- Rathinam, V.A.; Jiang, Z.; Waggoner, S.N.; Sharma, S.; Cole, L.E.; Waggoner, L.; Vanaja, S.K.; Monks, B.G.; Ganesan, S.; Latz, E. The AIM2 inflammasome is essential for host defense against cytosolic bacteria and DNA viruses. Nat. Immunol. 2010, 11, 395–402. [Google Scholar] [CrossRef]
- Biacchesi, S.; Mérour, E.; Lamoureux, A.; Bernard, J.; Brémont, M. Both STING and MAVS fish orthologs contribute to the induction of interferon mediated by RIG-I. PLoS ONE 2012, 7, e47737. [Google Scholar] [CrossRef]
- Burdette, D.L.; Vance, R.E. STING and the innate immune response to nucleic acids in the cytosol. Nat. Immunol. 2013, 14, 19–26. [Google Scholar] [CrossRef]
- Ishikawa, H.; Ma, Z.; Barber, G.N. STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 2009, 461, 788–792. [Google Scholar] [CrossRef]
- Jones, J.W.; Kayagaki, N.; Broz, P.; Henry, T.; Newton, K.; O’Rourke, K.; Chan, S.; Dong, J.; Qu, Y.; Roose-Girma, M. Absent in melanoma 2 is required for innate immune recognition of Francisella tularensis. PNAS 2010, 107, 9771–9776. [Google Scholar] [CrossRef]
- Prantner, D.; Darville, T.; Nagarajan, U.M. Stimulator of IFN gene is critical for induction of IFN-β during Chlamydia muridarum infection. J. Immunol. 2010, 184, 2551–2560. [Google Scholar] [CrossRef]
- Jin, L.; Hill, K.K.; Filak, H.; Mogan, J.; Knowles, H.; Zhang, B.; Perraud, A.-L.; Cambier, J.C.; Lenz, L.L. MPYS is required for IFN response factor 3 activation and type I IFN production in the response of cultured phagocytes to bacterial second messengers cyclic-di-AMP and cyclic-di-GMP. J. Immunol. 2011, 187, 2595–2601. [Google Scholar] [CrossRef]
- Sauer, J.-D.; Sotelo-Troha, K.; von Moltke, J.; Monroe, K.M.; Rae, C.S.; Brubaker, S.W.; Hyodo, M.; Hayakawa, Y.; Woodward, J.J.; Portnoy, D.A. The N-ethyl-N-nitrosourea-induced Goldenticket mouse mutant reveals an essential function of STING in the in vivo interferon response to Listeria monocytogenes and cyclic dinucleotides. Infect. Immun. 2011, 79, 688–694. [Google Scholar] [CrossRef]
- Abe, T.; Harashima, A.; Xia, T.; Konno, H.; Konno, K.; Morales, A.; Ahn, J.; Gutman, D.; Barber, G.N. STING recognition of cytoplasmic DNA instigates cellular defense. Mol. Cell 2013, 50, 5–15. [Google Scholar] [CrossRef]
- Sun, F.; Zhang, Y.-B.; Liu, T.-K.; Shi, J.; Wang, B.; Gui, J.-F. Fish MITA serves as a mediator for distinct fish IFN gene activation dependent on IRF3 or IRF7. J. Immunol. 2011, 187, 2531–2539. [Google Scholar] [CrossRef]
- Hornung, V.; Latz, E. Intracellular DNA recognition. Nat. Rev. Immunol. 2010, 10, 123–130. [Google Scholar] [CrossRef]
- Sullivan, C.; Postlethwait, J.H.; Lage, C.R.; Millard, P.J.; Kim, C.H. Evidence for evolving Toll-IL-1 receptor-containing adaptor molecule function in vertebrates. J. Immunol. 2007, 178, 4517–4527. [Google Scholar] [CrossRef]
- Chi, H.; Zhang, Z.; Bøgwald, J.; Zhan, W.; Dalmo, R.A. Cloning, expression analysis and promoter structure of TBK1 (tank-binding kinase 1) in Atlantic cod (Gadus morhua L.). Fish Shellfish Immunol. 2011, 30, 1055–1063. [Google Scholar] [CrossRef]
- Feng, H.; Liu, H.; Kong, R.; Wang, L.; Wang, Y.; Hu, W.; Guo, Q. Expression profiles of carp IRF-3/-7 correlate with the up-regulation of RIG-I/MAVS/TRAF3/TBK1, four pivotal molecules in RIG-I signaling pathway. Fish Shellfish Immunol. 2011, 30, 1159–1169. [Google Scholar] [CrossRef]
- Goldstein, J.L.; Ho, Y.; Basu, S.K.; Brown, M.S. Binding site on macrophages that mediates uptake and degradation of acetylated low density lipoprotein, producing massive cholesterol deposition. PNAS 1979, 76, 333–337. [Google Scholar] [CrossRef]
- DeWitte-Orr, S.J.; Collins, S.E.; Bauer, C.M.; Bowdish, D.M.; Mossman, K.L. An accessory to the “trinity”: SR-As are essential pathogen sensors of extracellular dsRNA, mediating entry and leading to subsequent type I IFN responses. PLoS Pathog. 2010, 6, e1000829. [Google Scholar] [CrossRef]
- Pearson, A.M. Scavenger receptors in innate immunity. Curr. Opin. Immunol. 1996, 8, 20–28. [Google Scholar] [CrossRef]
- Roach, J.C.; Glusman, G.; Rowen, L.; Kaur, A.; Purcell, M.K.; Smith, K.D.; Hood, L.E.; Aderem, A. The evolution of vertebrate toll-like receptors. PNAS 2005, 102, 9577–9582. [Google Scholar] [CrossRef]
- Doi, T.; Higashino, K.-I.; Kurihara, Y.; Wada, Y.; Miyazaki, T.; Nakamura, H.; Uesugi, S.; Imanishi, T.; Kawabe, Y.; Itakura, H. Charged collagen structure mediates the recognition of negatively charged macromolecules by macrophage scavenger receptors. J. Biol. Chem. 1993, 268, 2126–2133. [Google Scholar]
- Ojala, J.R.; Pikkarainen, T.; Tuuttila, A.; Sandalova, T.; Tryggvason, K. Crystal structure of the cysteine-rich domain of scavenger receptor MARCO reveals the presence of a basic and an acidic cluster that both contribute to ligand recognition. J. Biol. Chem. 2007, 282, 16654–16666. [Google Scholar] [CrossRef]
- Holmskov, U.; Malhotra, R.; Sim, R.B.; Jensenius, J.C. Collectins: Collagenous C-type lectins of the innate immune defense system. Immunol. Today 1994, 15, 67–74. [Google Scholar] [CrossRef]
- Martínez, V.G.; Moestrup, S.K.; Holmskov, U.; Mollenhauer, J.; Lozano, F. The conserved scavenger receptor cysteine-rich superfamily in therapy and diagnosis. Pharmacol. Rev. 2011, 63, 967–1000. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.S.; Goldstein, J.L. Lipoprotein metabolism in the macrophage: Implications for cholesterol deposition in atherosclerosis. Annu. Rev. Biochem. 1983, 52, 223–261. [Google Scholar] [CrossRef]
- Dumont, E.; Rallière, C.; Rescan, P.-Y. Identification of novel genes including Dermo-1, a marker of dermal differentiation, expressed in trout somitic external cells. J. Exp. Biol. 2008, 211, 1163–1168. [Google Scholar] [CrossRef]
- Meng, Z.; Zhang, X.-Y.; Guo, J.; Xiang, L.-X.; Shao, J.-Z. Scavenger receptor in fish is a lipopolysaccharide recognition molecule involved in negative regulation of NF-κB activation by competing with TNF receptor-associated factor 2 recruitment into the TNF-α signaling pathway. J. Immunol. 2012, 189, 4024–4039. [Google Scholar] [CrossRef]
- Martin-Armas, M.; Zykova, S.; Smedsrød, B. Effects of CpG-oligonucleotides, poly I:C and LPS on Atlantic cod scavenger endothelial cells (SEC). Dev. Comp. Immunol. 2008, 32, 100–107. [Google Scholar] [CrossRef]
- Kaur, H.; Jaso-Friedmann, L.; Evans, D.L. Identification of a scavenger receptor homologue on nonspecific cytotoxic cells and evidence for binding to oligodeoxyguanosine. Fish Shellfish Immunol. 2003, 15, 169–181. [Google Scholar] [CrossRef]
- Frøystad, M.K.; Rode, M.; Berg, T.; Gjøen, T. A role for scavenger receptors in phagocytosis of protein-coated particles in rainbow trout head kidney macrophages. Dev. Comp. Immunol. 1998, 22, 533–549. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Hajjar, D.P.; Khan, K.F.; Falcone, D.J. Ligand binding to macrophage scavenger receptor-A induces urokinase-type plasminogen activator expression by a protein kinase-dependent signaling pathway. J. Biol. Chem. 1998, 273, 1240–1246. [Google Scholar] [CrossRef]
- Whitman, S.C.; Daugherty, A.; Post, S.R. Regulation of acetylated low density lipoprotein uptake in macrophages by pertussis toxin-sensitive G proteins. J. Lipid Res. 2000, 41, 807–813. [Google Scholar]
- Hsu, H.-Y.; Chiu, S.-L.; Wen, M.-H.; Chen, K.-Y.; Hua, K.-F. Ligands of macrophage scavenger receptor induce cytokine expression via differential modulation of protein kinase signaling pathways. J. Biol. Chem. 2001, 276, 28719–28730. [Google Scholar] [CrossRef]
- Coller, S.P.; Paulnock, D.M. Signaling pathways initiated in macrophages after engagement of type A scavenger receptors. J. Leukocyte Biol. 2001, 70, 142–148. [Google Scholar]
- Kim, W.S.; Ordija, C.M.; Freeman, M.W. Activation of signaling pathways by putative scavenger receptor class A (SR-A) ligands requires CD14 but not SR-A. Biochem. Biophys. Res. Comm. 2003, 310, 542–549. [Google Scholar] [CrossRef]
- Murphy, J.E.; Tedbury, P.R.; Homer-Vanniasinkam, S.; Walker, J.H.; Ponnambalam, S. Biochemistry and cell biology of mammalian scavenger receptors. Atherosclerosis 2005, 182, 1–15. [Google Scholar] [CrossRef]
- Zou, J.; Secombes, C.J. Teleost fish interferons and their role in immunity. Dev. Comp. Immunol. 2011, 35, 1376–1387. [Google Scholar] [CrossRef]
- Fensterl, V.; Sen, G.C. Interferons and viral infections. Biofactors 2009, 35, 14–20. [Google Scholar] [CrossRef]
- González-Navajas, J.M.; Lee, J.; David, M.; Raz, E. Immunomodulatory functions of type I interferons. Nat. Rev. Immunol. 2012, 12, 125–135. [Google Scholar] [CrossRef]
- Goodbourn, S.; Didcock, L.; Randall, R. Interferons: Cell signalling, immune modulation, antiviral response and virus countermeasures. J. Gen. Virol. 2000, 81, 2341–2364. [Google Scholar]
- Parhi, J.; Mukherjee, S.; Saxena, G.; Sahoo, L.; Makesh, M. Molecular characterization and expression of type-I interferon gene in Labeo rohita. Mol. Biol. Rep. 2014, 41, 2979–2987. [Google Scholar] [CrossRef]
- Ohtani, M.; Hikima, J.-I.; Hwang, S.D.; Morita, T.; Suzuki, Y.; Kato, G.; Kondo, H.; Hirono, I.; Jung, T.-S.; Aoki, T. Transcriptional regulation of type I interferon gene expression by interferon regulatory factor-3 in Japanese flounder, Paralichthys olivaceus. Dev. Comp. Immunol. 2012, 36, 697–706. [Google Scholar] [CrossRef]
- Wan, Q.; Wicramaarachchi, W.N.; Whang, I.; Lim, B.-S.; Oh, M.-J.; Jung, S.-J.; Kim, H.C.; Yeo, S.-Y.; Lee, J. Molecular cloning and functional characterization of two duplicated two-cysteine containing type I interferon genes in rock bream Oplegnathus fasciatus. Fish Shellfish Immunol. 2012, 33, 886–898. [Google Scholar] [CrossRef]
- Li, D.; Tan, W.; Ma, M.; Yu, X.; Lai, Q.; Wu, Z.; Lin, G.; Hu, C. Molecular characterization and transcription regulation analysis of type I IFN gene in grass carp (Ctenopharyngodon idella). Gene 2012, 504, 31–40. [Google Scholar] [CrossRef]
- Adamek, M.; Rakus, K.Ł.; Chyb, J.; Brogden, G.; Huebner, A.; Irnazarow, I.; Steinhagen, D. Interferon type I responses to virus infections in carp cells: In vitro studies on Cyprinid herpesvirus 3 and Rhabdovirus carpio infections. Fish Shellfish Immunol. 2012, 33, 482–493. [Google Scholar] [CrossRef]
- Haller, O.; Staeheli, P.; Kochs, G. Interferon-induced Mx proteins in antiviral host defense. Biochimie 2007, 89, 812–818. [Google Scholar] [CrossRef]
- Leong, J.A.C.; Trobridge, G.D.; Kim, C.H.; Johnson, M.; Simon, B. Interferon-inducible Mx proteins in fish. Immunol. Rev. 1998, 166, 349–363. [Google Scholar] [CrossRef]
- Pavlovic, J.; Zürcher, T.; Haller, O.; Staeheli, P. Resistance to influenza virus and vesicular stomatitis virus conferred by expression of human MxA protein. J. Virol. 1990, 64, 3370–3375. [Google Scholar]
- Staeheli, P.; Haller, O.; Boll, W.; Lindenmann, J.; Weissmann, C. Mx protein: Constitutive expression in 3T3 cells transformed with cloned Mx cDNA confers selective resistance to influenza virus. Cell 1986, 44, 147–158. [Google Scholar] [CrossRef]
- Larsen, R.; Røkenes, T.P.; Robertsen, B. Inhibition of infectious pancreatic necrosis virus replication by Atlantic salmon Mx1 protein. J. Virol. 2004, 78, 7938–7944. [Google Scholar] [CrossRef]
- Caipang, C.M.A.; Hirono, I.; Aoki, T. In vitro inhibition of fish rhabdoviruses by Japanese flounder, Paralichthys olivaceus Mx. Virology 2003, 317, 373–382. [Google Scholar] [CrossRef]
- Skaug, B.; Chen, Z.J. Emerging role of ISG15 in antiviral immunity. Cell 2010, 143, 187–190. [Google Scholar] [CrossRef]
- Huang, X.; Huang, Y.; Cai, J.; Wei, S.; Ouyang, Z.; Qin, Q. Molecular cloning, expression and functional analysis of ISG15 in orange-spotted grouper, Epinephelus coioides. Fish Shellfish Immunol. 2013, 34, 1094–1102. [Google Scholar] [CrossRef]
- Langevin, C.; van der Aa, L.; Houel, A.; Torhy, C.; Briolat, V.; Lunazzi, A.; Harmache, A.; Bremont, M.; Levraud, J.-P.; Boudinot, P. Zebrafish ISG15 exerts a strong antiviral activity against RNA and DNA viruses and regulates the interferon response. J. Virol. 2013, 87, 10025–10036. [Google Scholar] [CrossRef]
- Zhang, Y.-B.; Jiang, J.; Chen, Y.-D.; Zhu, R.; Shi, Y.; Zhang, Q.-Y.; Gui, J.-F. The innate immune response to grass carp hemorrhagic virus (GCHV) in cultured Carassius auratus blastulae (CAB) cells. Dev. Comp. Immunol. 2007, 31, 232–243. [Google Scholar] [CrossRef]
- Sun, F.; Zhang, Y.-B.; Jiang, J.; Wang, B.; Chen, C.; Zhang, J.; Gui, J.-F. GIG1, a novel antiviral effector involved in fish interferon response. Virology 2014, 448, 322–332. [Google Scholar] [CrossRef]
- Sun, C.; Liu, Y.; Hu, Y.; Fan, Q.; Li, W.; Yu, X.; Mao, H.; Hu, C. GIG1 and GIG2 homologs (CiGIG1 and CiGIG2) from grass carp (Ctenopharyngodon idella) display good antiviral activities in an IFN-independent pathway. Dev. Comp. Immunol. 2013, 41, 477–483. [Google Scholar] [CrossRef]
- Franchi, L.; Muñoz-Planillo, R.; Núñez, G. Sensing and reacting to microbes through the inflammasomes. Nat. Immunol. 2012, 13, 325–332. [Google Scholar] [CrossRef]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef]
- Arend, W.P.; Palmer, G.; Gabay, C. IL-1, IL-18, and IL-33 families of cytokines. Immunol. Rev. 2008, 223, 20–38. [Google Scholar] [CrossRef]
- Secombes, C.J.; Wang, T.; Bird, S. The interleukins of fish. Dev. Comp. Immunol. 2011, 35, 1336–1345. [Google Scholar] [CrossRef]
- Wang, T.; Bird, S.; Koussounadis, A.; Holland, J.W.; Carrington, A.; Zou, J.; Secombes, C.J. Identification of a novel IL-1 cytokine family member in teleost fish. J. Immunol. 2009, 183, 962–974. [Google Scholar] [CrossRef]
- Bird, S.; Zou, J.; Wang, T.; Munday, B.; Cunningham, C.; Secombes, C.J. Evolution of interleukin-1β. Cytokine Growth Factor Rev. 2002, 13, 483–502. [Google Scholar] [CrossRef]
- Fujiki, K.; Shin, D.-H.; Nakao, M.; Yano, T. Molecular cloning and expression analysis of carp (Cyprinus carpio) interleukin-1β, high affinity immunoglobulin E Fc receptor γ subunit and serum amyloid A. Fish Shellfish Immunol. 2000, 10, 229–242. [Google Scholar] [CrossRef]
- Zou, J.; Grabowski, P.S.; Cunningham, C.; Secombes, C.J. Molecular cloning of interleukin 1β from rainbow trout Oncorhynchus mykiss reveals no evidence of an ice cut site. Cytokine 1999, 11, 552–560. [Google Scholar] [CrossRef]
- Ingerslev, H.; Cunningham, C.; Wergeland, H. Cloning and expression of TNF-α, IL-1β and COX-2 in an anadromous and landlocked strain of Atlantic salmon (Salmo salar L.) during the smolting period. Fish Shellfish Immunol. 2006, 20, 450–461. [Google Scholar] [CrossRef]
- Pelegrı́n, P.; Garcı́a-Castillo, J.; Mulero, V.; Meseguer, J. Interleukin-1β isolated from a marine fish reveals up-regulated expression in macrophages following activation with lipopolysaccharide and lymphokines. Cytokine 2001, 16, 67–72. [Google Scholar] [CrossRef]
- Scapigliati, G.; Buonocore, F.; Bird, S.; Zou, J.; Pelegrin, P.; Falasca, C.; Prugnoli, D.; Secombes, C. Phylogeny of cytokines: Molecular cloning and expression analysis of sea bass Dicentrarchus labrax interleukin-1β. Fish Shellfish Immunol. 2001, 11, 711–726. [Google Scholar] [CrossRef]
- Lee, D.-S.; Hong, S.H.; Lee, H.-J.; Jun, L.J.; Chung, J.-K.; Kim, K.H.; do Jeong, H. Molecular cDNA cloning and analysis of the organization and expression of the IL-1β gene in the Nile tilapia, Oreochromis niloticus. Comp. Biochem. Physiol. A 2006, 143, 307–314. [Google Scholar] [CrossRef]
- Lu, D.-Q.; Bei, J.-X.; Feng, L.-N.; Zhang, Y.; Liu, X.-C.; Wang, L.; Chen, J.-L.; Lin, H.-R. Interleukin-1β gene in orange-spotted grouper, Epinephelus coioides: Molecular cloning, expression, biological activities and signal transduction. Mol. Immunol. 2008, 45, 857–867. [Google Scholar] [CrossRef]
- Covello, J.; Bird, S.; Morrison, R.; Battaglene, S.; Secombes, C.; Nowak, B. Cloning and expression analysis of three striped trumpeter (Latris lineata) pro-inflammatory cytokines, TNF-α, IL-1β and IL-8, in response to infection by the ectoparasitic, Chondracanthus goldsmidi. Fish Shellfish Immunol. 2009, 26, 773–786. [Google Scholar] [CrossRef]
- Corripio-Miyar, Y.; Bird, S.; Tsamopoulos, K.; Secombes, C. Cloning and expression analysis of two pro-inflammatory cytokines, IL-1β and IL-8, in haddock (Melanogrammus aeglefinus). Mol. Immunol. 2007, 44, 1361–1373. [Google Scholar] [CrossRef]
- Tsutsui, S.; Iwamoto, T.; Nakamura, O.; Watanabe, T. LPS induces gene expression of interleukin-1β in conger eel (Conger myriaster) macrophages: First cytokine sequence within anguilliformes. Fish Shellfish Immunol. 2007, 23, 911–916. [Google Scholar] [CrossRef]
- Pleguezuelos, O.; Zou, J.; Cunningham, C.; Secombes, C.J. Cloning, sequencing, and analysis of expression of a second IL-1β gene in rainbow trout (Oncorhynchus mykiss). Immunogenetics 2000, 51, 1002–1011. [Google Scholar] [CrossRef]
- Engelsma, M.Y.; Stet, R.J.; Saeij, J.P.; Verburg-van Kemenade, B.L. Differential expression and haplotypic variation of two interleukin-1β genes in the common carp (Cyprinus carpio L.). Cytokine 2003, 22, 21–32. [Google Scholar] [CrossRef]
- Husain, M.; Bird, S.; van Zwieten, R.; Secombes, C.J.; Wang, T. Cloning of the IL-1β3 gene and IL-1β4 pseudogene in salmonids uncovers a second type of IL-1β gene in teleost fish. Dev. Comp. Immunol. 2012, 38, 431–446. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Baoprasertkul, P.; Peatman, E.; Liu, Z. Genomic organization, gene duplication, and expression analysis of interleukin-1β in channel catfish (Ictalurus punctatus). Mol. Immunol. 2006, 43, 1653–1664. [Google Scholar] [CrossRef]
- Hong, S.; Zou, J.; Collet, B.; Bols, N.C.; Secombes, C.J. Analysis and characterisation of IL-1β processing in rainbow trout, Oncorhynchus mykiss. Fish Shellfish Immunol. 2004, 16, 453–459. [Google Scholar] [CrossRef]
- Pelegrı́n, P.; Chaves-Pozo, E.; Mulero, V.; Meseguer, J. Production and mechanism of secretion of interleukin-1β from the marine fish gilthead seabream. Dev. Comp. Immunol. 2004, 28, 229–237. [Google Scholar] [CrossRef]
- Koussounadis, A.I.; Ritchie, D.W.; Kemp, G.J.; Secombes, C.J. Analysis of fish IL-1β and derived peptide sequences indicates conserved structures with species-specific IL-1 receptor binding: Implications for pharmacological design. Curr. Pharm. Des. 2004, 10, 3857–3871. [Google Scholar] [CrossRef]
- Bilen, S.; Biswas, G.; Otsuyama, S.; Kono, T.; Sakai, M.; Hikima, J.-I. Inflammatory responses in the Japanese pufferfish (Takifugu rubripes) head kidney cells stimulated with an inflammasome-inducing agent, nigericin. Dev. Comp. Immunol. 2014, 46, 222–230. [Google Scholar] [CrossRef]
- Martin, S.A.; Zou, J.; Houlihan, D.F.; Secombes, C.J. Directional responses following recombinant cytokine stimulation of rainbow trout (Oncorhynchus mykiss) RTS-11 macrophage cells as revealed by transcriptome profiling. BMC Genomics 2007. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Kwang, J. Carp interleukin-1β in the role of an immuno-adjuvant. Fish Shellfish Immunol. 2000, 10, 375–378. [Google Scholar] [CrossRef]
- Hong, S.; Peddie, S.; Campos-Pérez, J.J.; Zou, J.; Secombes, C.J. The effect of intraperitoneally administered recombinant IL-1β on immune parameters and resistance to Aeromonas salmonicida in the rainbow trout (Oncorhynchus mykiss). Dev. Comp. Immunol. 2003, 27, 801–812. [Google Scholar] [CrossRef]
- Angosto, D.; López-Castejón, G.; López-Muñoz, A.; Sepulcre, M.P.; Arizcun, M.; Meseguer, J.; Mulero, V. Evolution of inflammasome functions in vertebrates: Inflammasome and caspase-1 trigger fish macrophage cell death but are dispensable for the processing of IL-1β. Innate Immun. 2012, 18, 815–824. [Google Scholar] [CrossRef]
- Reis, M.I.; do Vale, A.; Pereira, P.J.; Azevedo, J.E.; dos Santos, N.M. Caspase-1 and IL-1β processing in a teleost fish. PLoS ONE 2012, 7, e50450. [Google Scholar] [CrossRef]
- Vojtech, L.N.; Scharping, N.; Woodson, J.C.; Hansen, J.D. Roles of inflammatory caspases during processing of zebrafish interleukin-1β in Francisella noatunensis infection. Infect. Immun. 2012, 80, 2878–2885. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poynter, S.; Lisser, G.; Monjo, A.; DeWitte-Orr, S. Sensors of Infection: Viral Nucleic Acid PRRs in Fish. Biology 2015, 4, 460-493. https://doi.org/10.3390/biology4030460
Poynter S, Lisser G, Monjo A, DeWitte-Orr S. Sensors of Infection: Viral Nucleic Acid PRRs in Fish. Biology. 2015; 4(3):460-493. https://doi.org/10.3390/biology4030460
Chicago/Turabian StylePoynter, Sarah, Graeme Lisser, Andrea Monjo, and Stephanie DeWitte-Orr. 2015. "Sensors of Infection: Viral Nucleic Acid PRRs in Fish" Biology 4, no. 3: 460-493. https://doi.org/10.3390/biology4030460