1. Introduction
Despite tremendous progress in understanding the contractile mechanism in muscle and other myosin motors (Wulf et al. [
1]; Squire et al. [
2]), it is still uncertain exactly how many myosin heads interact at the same time with actin filaments in muscle to utilise the energy associated with ATP hydrolysis and produce muscular contraction. It is also not known how many different attached states there are and how much force is produced by each state. What is known is that in all muscles myosin (M) with ATP bound and hydrolysed to ADP and inorganic phosphate (Pi) in the form M.ADP.Pi can attach to actin filaments after the muscles have been switched on and that, while attached, the products Pi and then ADP are released [
3]. During some stage of this process force and movement (if the muscle is free to shorten) are produced. Binding of another ATP molecule splits the heads from actin and a further cycle of hydrolysis and actin attachment can then take place.
The myosin molecule has a long rod-shaped region, a two chain coiled-coil α-helix, on the end of which are two elongated globular heads which carry the ATP hydrolysis and actin binding sites of the molecule. The myosin rods form the shaft or backbone of myosin filaments and the myosin heads project from the surface of this backbone and can bind to and interact with adjacent actin filaments (see overview in [
2]). This much we know. However, what are the structural changes in the myosin heads that are associated with force production and movement? The myosin head has two major domains within it, a globular
motor domain which is the enzymatic and actin attachment part of the head, and a long
lever arm consisting of a long α–helix surrounded by two light chains. Crystal structures show that the motor and lever arm are linked through a small region called the
converter domain [
4,
5,
6,
7] and this has revealed two main kinds of structural difference between states. One is the marked difference in the angle at which the lever arm comes out of the motor domain. The other is the opening and closing of a cleft in the motor domain [
7,
8,
9]. It is now thought that movement is brought about by the swinging of the lever arm around a relatively fixed myosin motor domain bound to actin [
4,
5]. However, at what stage is force produced? What is the shape of the myosin head when it first attaches to actin? Does the lever arm of attached heads swing through a series of steps or are there, say, only two main structural attached states of the head, together with elasticity in some part of the head (e.g., in the lever arm or perhaps converter/transducer domains)?
Low-angle X-ray diffraction patterns from a variety of muscles have been recorded with ever increasing success over a number of decades ([
10,
11,
12,
13,
14,
15,
16,
17,
18,
19,
20,
21,
22] and many others; see [
23,
24,
25] for overviews and many additional references). Much of the improvement has been due to the exploitation of ever more powerful synchrotron radiation sources and the development of very fast 2D X-ray detectors [
26]. Important and unambiguous information has come from these studies. However, in no case have the diffraction data from the whole low-angle 2D diffraction pattern been modelled fully at every stage of the contractile cycle.
The disadvantage of the technique of X-ray diffraction is that it does not provide direct images of the diffracting object, the diffraction patterns need to be interpreted and modelled. These diffraction patterns carry a wealth of information, but they can sometimes be over-interpreted. For example, in a series of papers, the so-called M3 reflection of the meridian of diffraction patterns from frog muscles (the M3 corresponds to the 14.3 nm repeat of “crowns” of myosin heads in the myosin filaments), was used in an attempt to deduce the angles that the lever arms of myosin heads make on the motor domain under different conditions [
27,
28,
29,
30,
31,
32,
33,
34,
35,
36,
37,
38,
39,
40]; for further references see [
41]. Unfortunately, this was not unambiguous, and we showed [
41] that these results were simply reporting the relative axial positions of different motor domains in different populations of myosin heads without specifying the location of the lever arm.
Interpretation of diffraction data is not trivial and a very helpful pre-requisite is that the muscle doing the diffracting is very highly ordered so that it gives well-sampled X-ray diffraction patterns. Muscles of choice are the indirect flight muscles of insects such as the giant water bug
Lethocerus [
42,
43], and the muscles of bony fish which for the vertebrates are the most highly regular muscles [
44,
45]. Bony fish muscle order is such that all the myosin filaments in a given A-band have exactly the same rotation around their long axes, so that their crowns of heads make a quasi-crystalline three-dimensional (3D) array. This is a simple lattice muscle [
46]. Other vertebrate muscles such as those in frogs, rabbits, chickens, humans and all higher vertebrates have a so-called statistical superlattice A-band where the myosin filaments can have one of two rotations around their long axes, but these rotations are not regularly distributed across the A-band [
45,
46,
47,
48,
49]. Simple lattice muscles give highly sampled myosin diffraction patterns [
15,
44,
50], but superlattice muscles do not, and their X-ray patterns are, therefore, intrinsically more difficult to solve.
In previous work on bony fish muscles using either static muscles [
44] or in a time-resolved way [
15,
24,
44,
51,
52], we suggested (as have others using other muscles) that there is a population of heads, possibly weak-binding heads, which attach transiently to actin at an early stage in the contractile cycle, but which do not yet produce high levels of force. These changes were recorded without SL control and any muscle shortening during the contraction would complicate the interpretation of what is happening. The present paper outlines our approach to determining the full 2D low-angle X-ray diffraction pattern from Plaice fin muscle in a time-resolved way so that the contractile mechanism can be fully modelled. We sought to optimise as much as possible about this preparation, including the dissection step, the Ringer solution used and the nature of the electrical stimulus. We also tested the effects of sarcomere length control. In this paper, we present the methods and results from this optimization. The whole 2D diffraction pattern was recorded out to about
d = 60 Å. Here we describe the resulting diffraction data from the equator of the diffraction pattern and we model the results using a new procedure. Results from other parts of the diffraction pattern will be presented elsewhere.
4. Discussion
4.1. Plaice Fin Muscle Handling and Properties
The immediate aim of this work was to record the changing intensities of the equatorial peaks from bony fish muscle in a time-resolved way, improved by SL control and improved muscle treatment, and to model the results to learn about crossbridge behaviour. However, it was also to establish whole bony fish muscle as a viable preparation to study the whole of the low-angle X-ray diffraction patterns from myosin and actin filaments in active muscle. The muscle of choice for this project was that of bony fish skeletal muscle because of its simple lattice structure and well-sampled resting X-ray diffraction patterns [
44]. The performance of the active fin muscle specimens was improved to increase their tension output and to reduce their rate of fatigue (see Materials and Methods
Section 2.1 and the
Supplementary Materials Section S2). A method was also developed to set up a SL measurement and control system for active whole Plaice fin muscles; muscles large enough to give reasonable diffraction statistics even with relatively weak reflections.
We wanted to assess the effect of SL change in active bony fish muscle because SL is known to have an effect on both the time-course of tension development and the changes in equatorial X-ray intensities in other muscles [
59,
69]. Previous work carried out on bony fish muscle neglected SL changes when interpreting the relationship between tension and X-ray intensity time-courses from tetanically contracting muscle [
15,
24,
51,
80]. Until this current project the contractile SL change in bony fish muscle had not been measured and the extent to which such changes might affect the tension and X-ray time-courses and the conclusions drawn from them about the cross-bridge cycle was not known definitively.
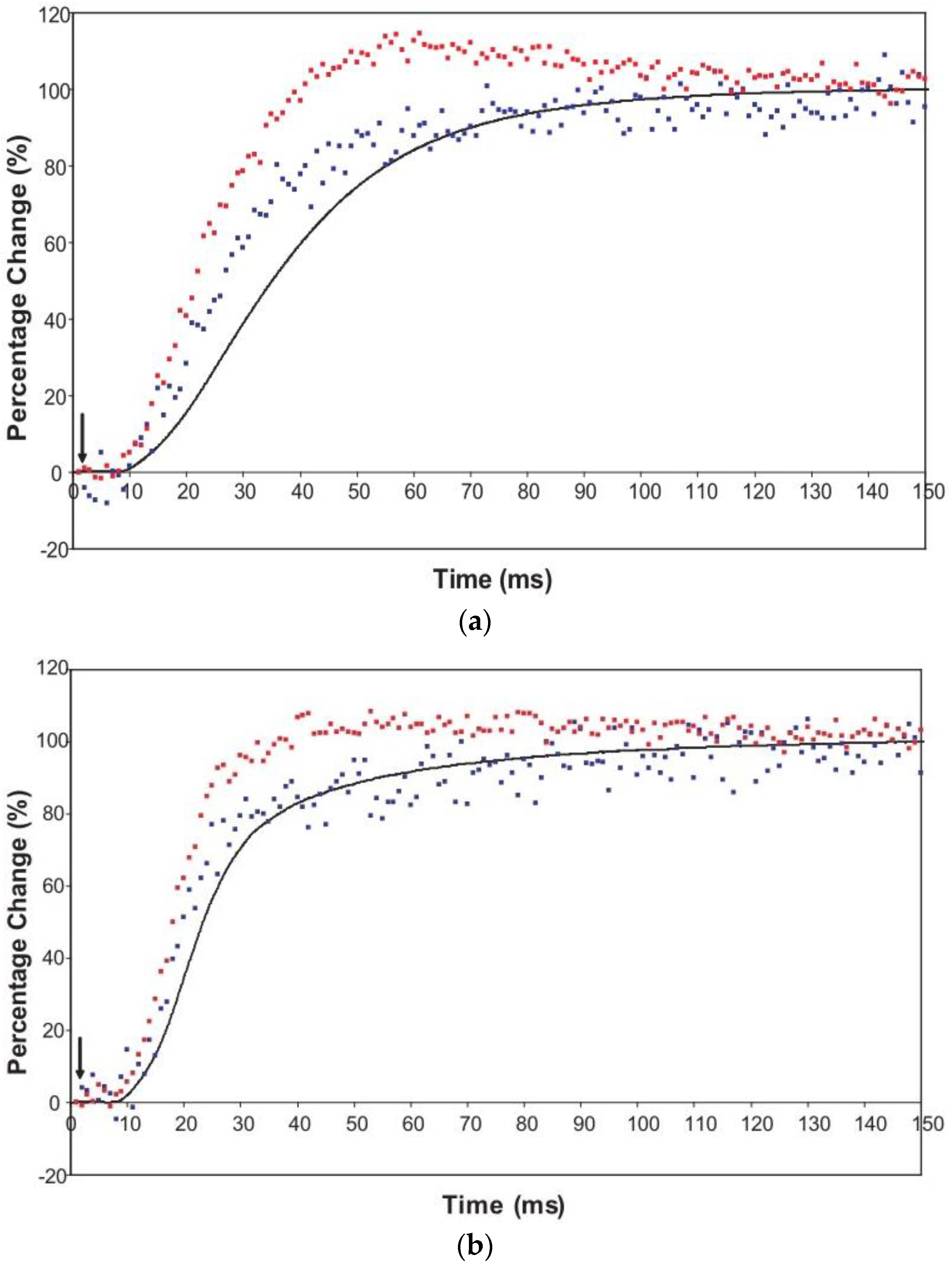
Table 1 compares the time responses of various muscles during the rising phase of tetanic contractions with the new results in the present paper. Immediately apparent is that Plaice fin muscles under SL control at 7 to 8 °C show a very fast response; the time to half tension change is 22.5 ± 0.8 ms compared to 30 to 50 ms in other intact, electrically stimulated, muscles.
4.2. Deductions about the Cross-Bridge Mechanism
Here, we evaluate what the observed equatorial intensity changes mean in terms of changes in myosin cross-bridge organisation during the contractile cycle. We are concerned to provide conclusions that are facts rather than interpretations. Equation (9), known as the structure factor, allows the amplitude of each X-ray reflection from a lattice to be calculated from the contents of the unit cell.
F(h, k, l) is the amplitude of a particular reflection with indices (h, k, l) from mass at fractional unit cell coordinates of (xj, yj, zj) and with scattering factor of Fj.
Equation (9) can be simplified for the 2D myofilament lattice (the 3D lattice projected down the filament long axis) to give the contribution of the actin and myosin scattering factors to each equatorial reflection, considered as simple rotationally symmetric densities (i.e., roughly cylindrical structures viewed down their long axes). In the case of the muscle A-band lattice, the unit cell contains one myosin filament at coordinates (0, 0, 0) and two actin filaments at (1/3, 2/3, 0) and (2/3, 1/3, 0). This gives a general expression for the amplitude of a muscle equatorial reflection as in Equation (10):
Here,
FM and
FA are the scattering factors of the myosin and actin filaments respectively in those particular directions. By calculating the contribution of the myosin and actin scattering factors to each equatorial reflection it is possible to deduce possible reasons why the changes in intensity described above occur. The reasoning is based upon evidence from electron density maps of the A-band lattice using Fourier synthesis, and time-courses of mass transfer in the 9 nm annulus around the thin filament centres during contraction [
15,
23,
79,
82,
83,
84]. These data indicate that as muscles contract, mass from around the myosin filament centres moves away from the thick filaments towards the thin filaments. This moving mass is thought to be the myosin cross-bridges as they attach to the thin filaments during contraction.
Table 2 shows the calculations of the multiplication factor of
FA in Equation (10) for each equatorial reflection (
e2πiθ has been converted to cos (2π
θ) +
isin (2π
θ) for the purpose of the calculations).
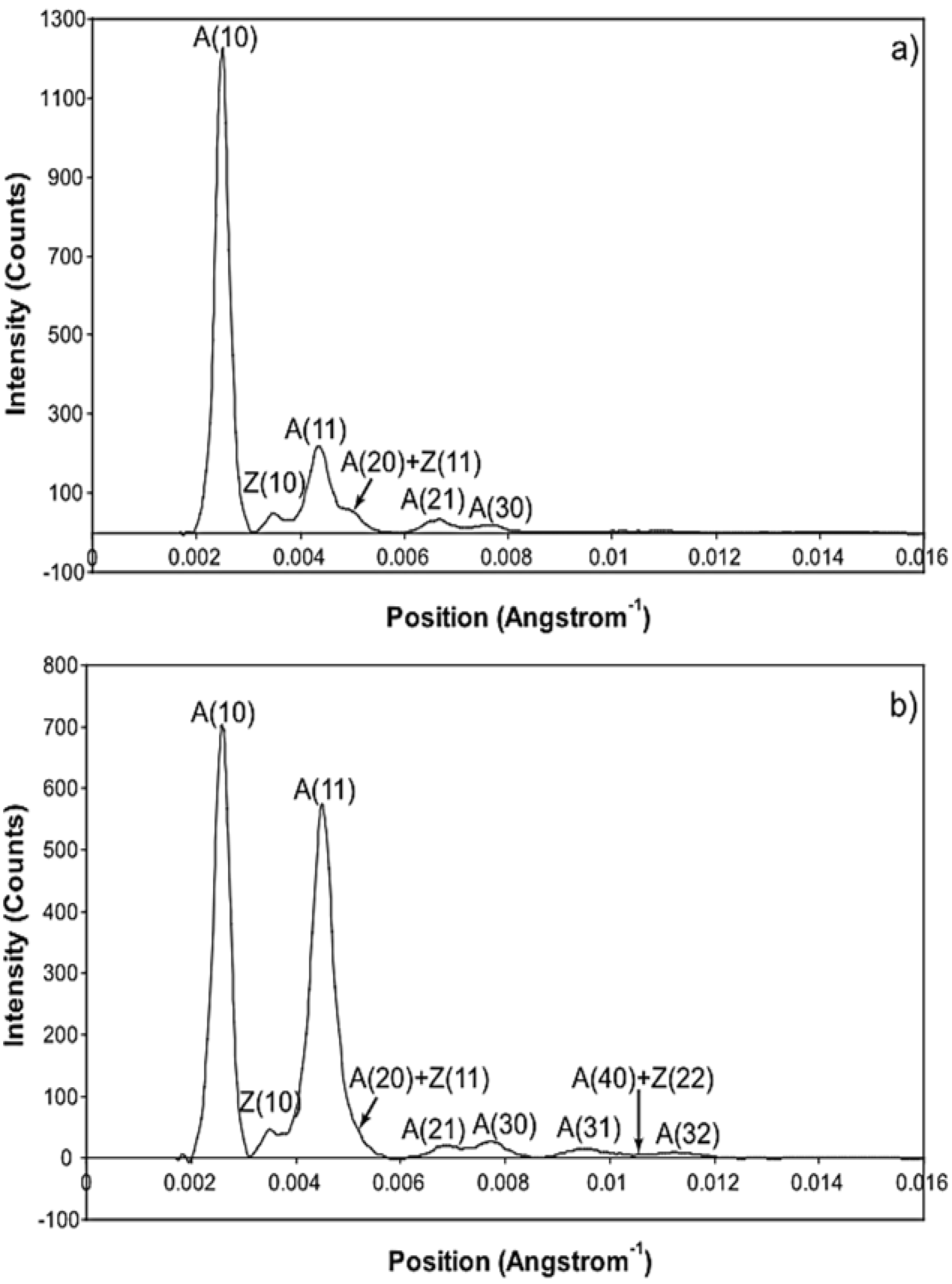
As shown in
Table 3, based on calculations of the simple model using Equation (10), each reflection out to the A(30) is observed to change in intensity in a way consistent with the general movement of cross-bridge mass to actin during the rising phase of the tetanus. If the myosin heads just moved out radially without clustering around actin the result would be different. In particular the A(11) peaks would not increase in such a marked way, as in rigor muscle where we know the heads are attached. The characteristic of equatorial X-ray diffraction patterns from rigor muscle, where most heads are actin-attached [
85,
86], is to have a relatively strong A(11) peak and a relatively weak A(10) peak, consistent with the analysis in
Table 3. Active muscle has intensities part way between the resting and rigor values. What is very different here, is that the outer reflections A(31), A(40) and A(32) change in a different way to that expected, indicating more complexity in the cross-bridge cycle than in the simple model used to calculate
Table 3, where the heads are just assumed to add mass to the actin positions [
87,
88].
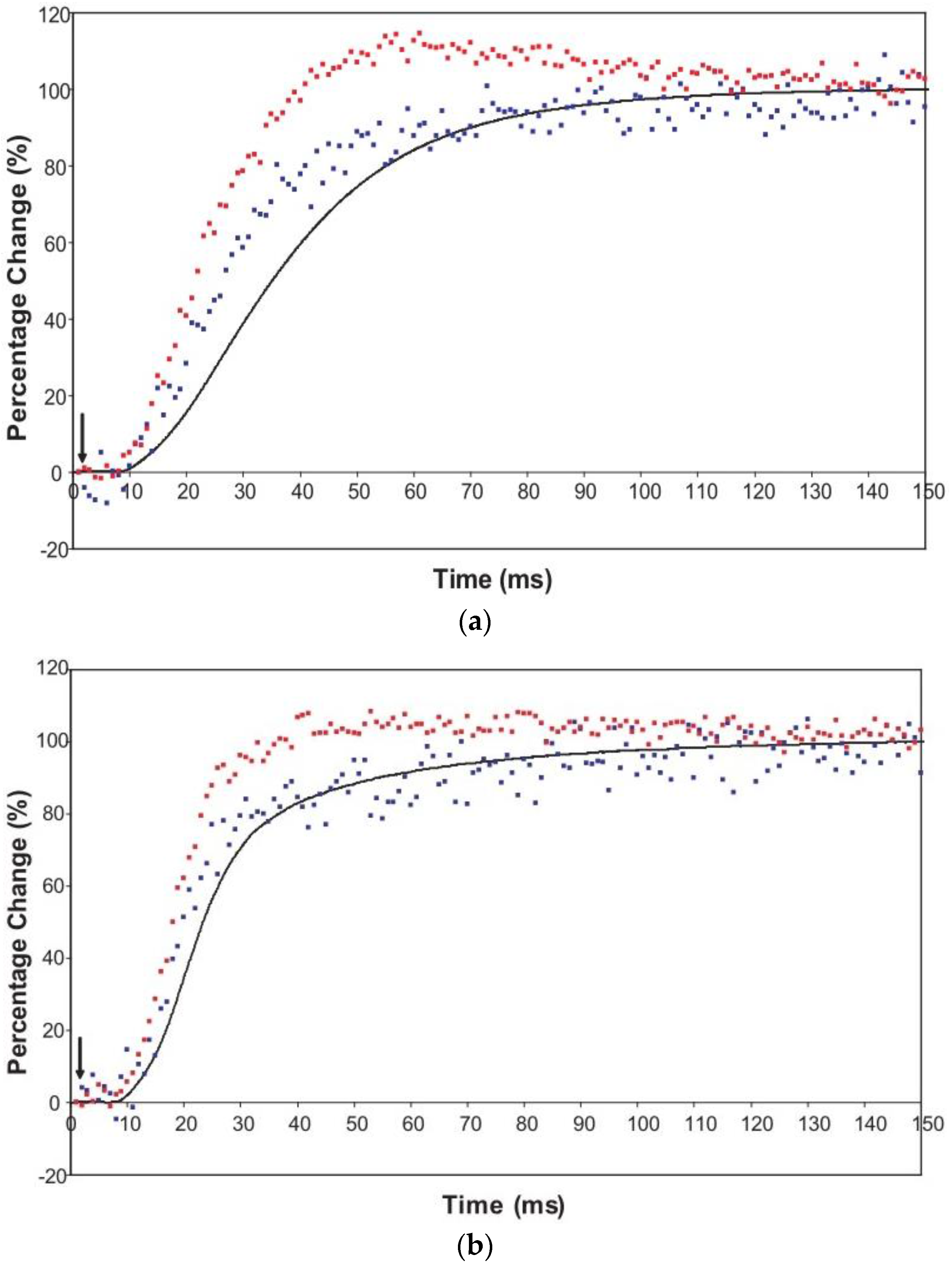
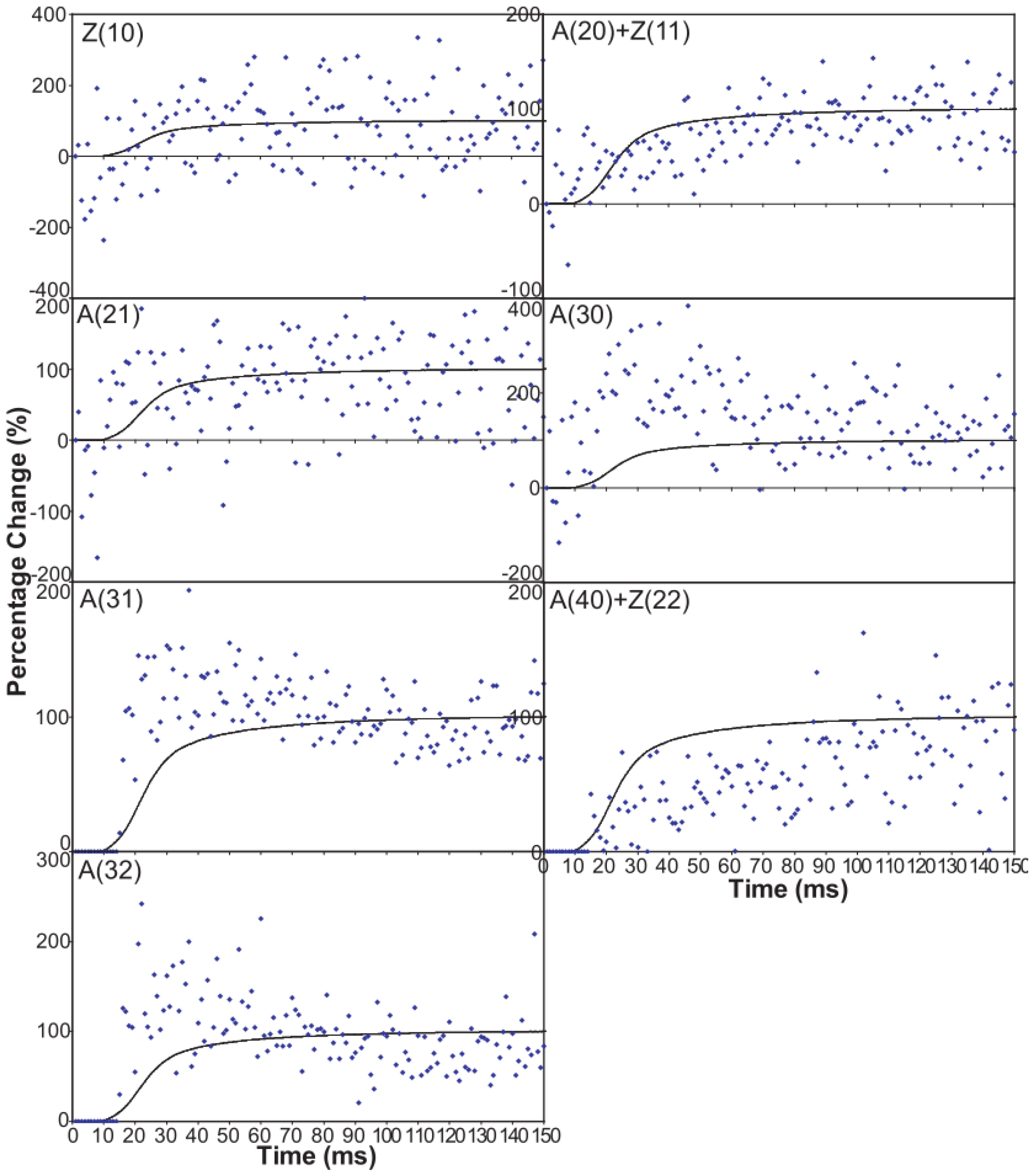
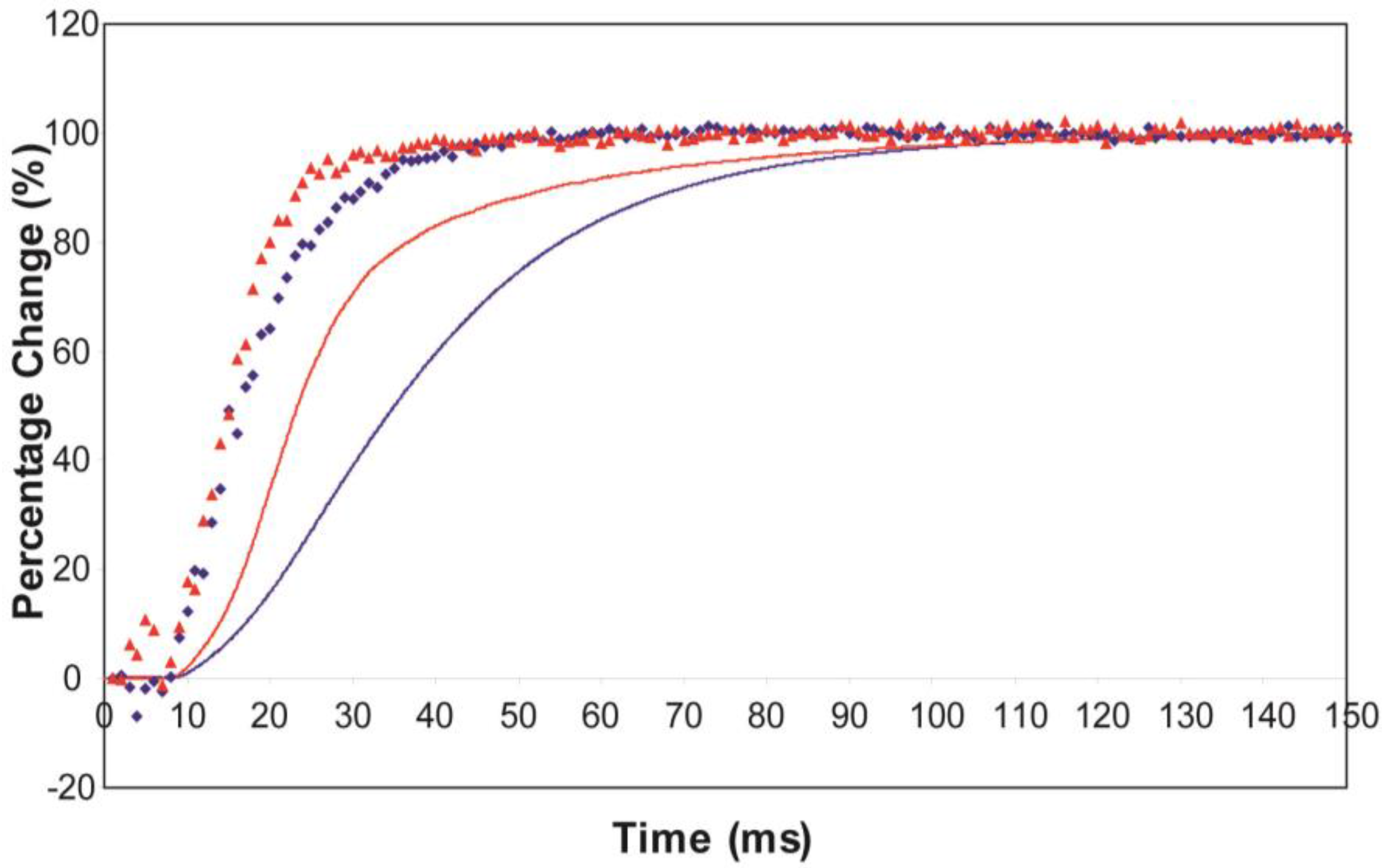
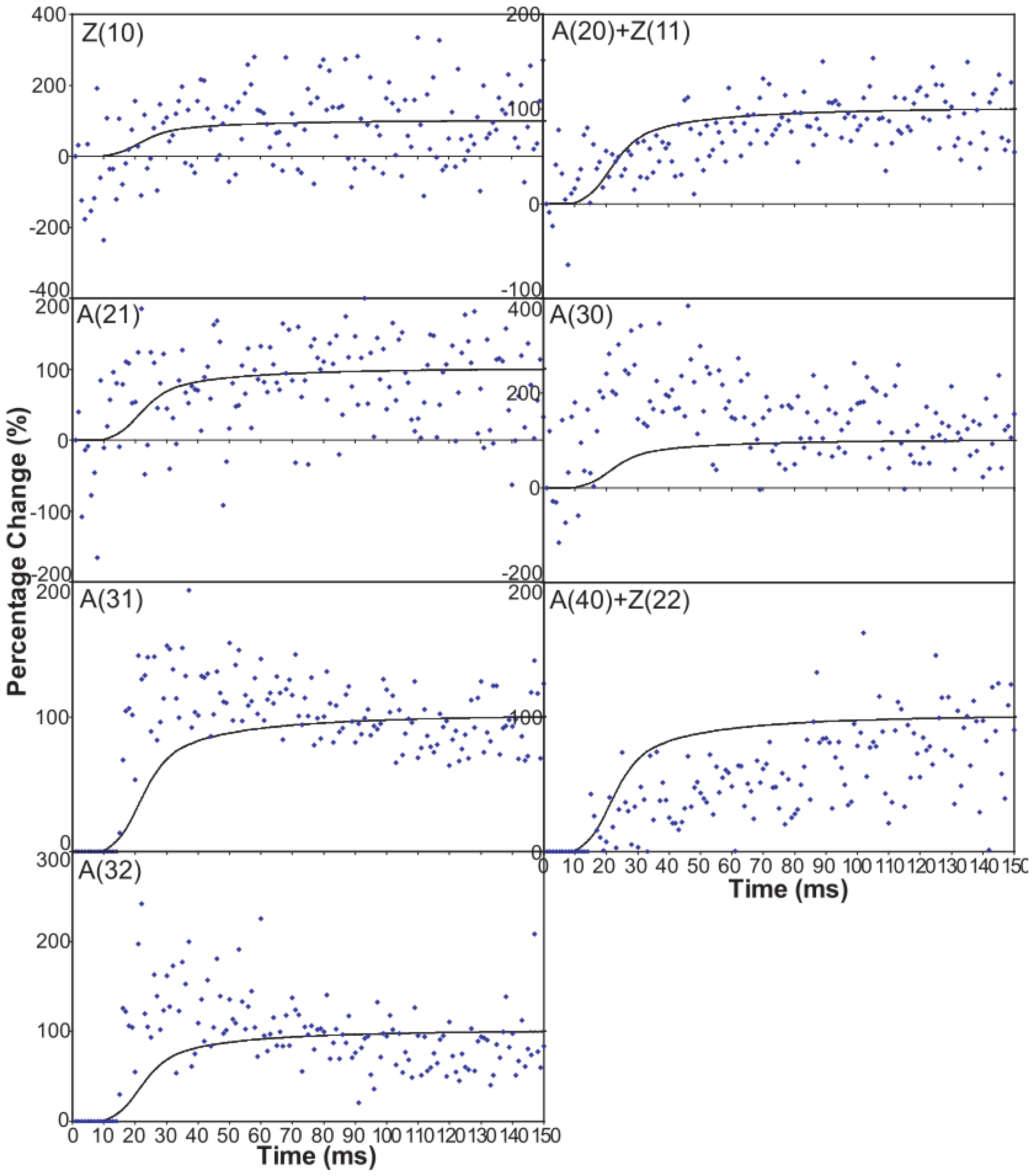
An indication of what this added complexity might be comes from the observation that not only does the A(11) intensity still lead tension and have a different shape to its time-course than the tension, even with partial SL control, but other weaker peaks like the A(30), A(31) and A(32) peaks all change very rapidly and overshoot their plateau values, in the case of the A(30) and A(32) peaks roughly by a factor of two, before the tension plateau is reached. Assuming (as above) that the observed intensity changes are solely due to myosin head movements, the time-courses of the equatorial peaks in
Figure 4 and
Figure 5, particularly the higher orders in
Figure 6, show that there is not just a change in the number of the attached heads, but that at an early stage in the development of tension there are heads in a different conformation or distribution of conformations to when the full tension (plateau) level is reached. This is unambiguous.
In thinking about what this might mean, we believe [
41] as do others that active muscle contains a mixture of at least three myosin head structural states; some strongly attached heads (AM.ADP.Pi, AM.ADP, AM), some detached, resetting heads (M.ATP to M.ADP.Pi) and thirdly some heads in a state (AM.ADP.Pi*; possibly weak binding, pre-powerstroke heads) which would give a relatively strong A(11) peak, an A(10) not far from its relaxed value and strong outer intensities at the A(30), A(31) and A(32) positions. This latter state may be the non-force-producing initial actin-attached state in fish muscle as suggested by Harford and Squire [
15]. It may be the elusive pre-powerstroke state that is often discussed in muscle cross-bridge scenarios [
9,
19].
4.3. Modelling the Equatorial Time-Courses
In an attempt to model objectively the equatorial time-courses we have supposed that what is being observed is a mixture of structurally different states. We assume that the myosin and actin filament backbones are virtually unchanged throughout a tetanus at the resolution we are considering (60 Å at best) and that all the observed changes are due to crossbridge movements and crossbridge shape changes.
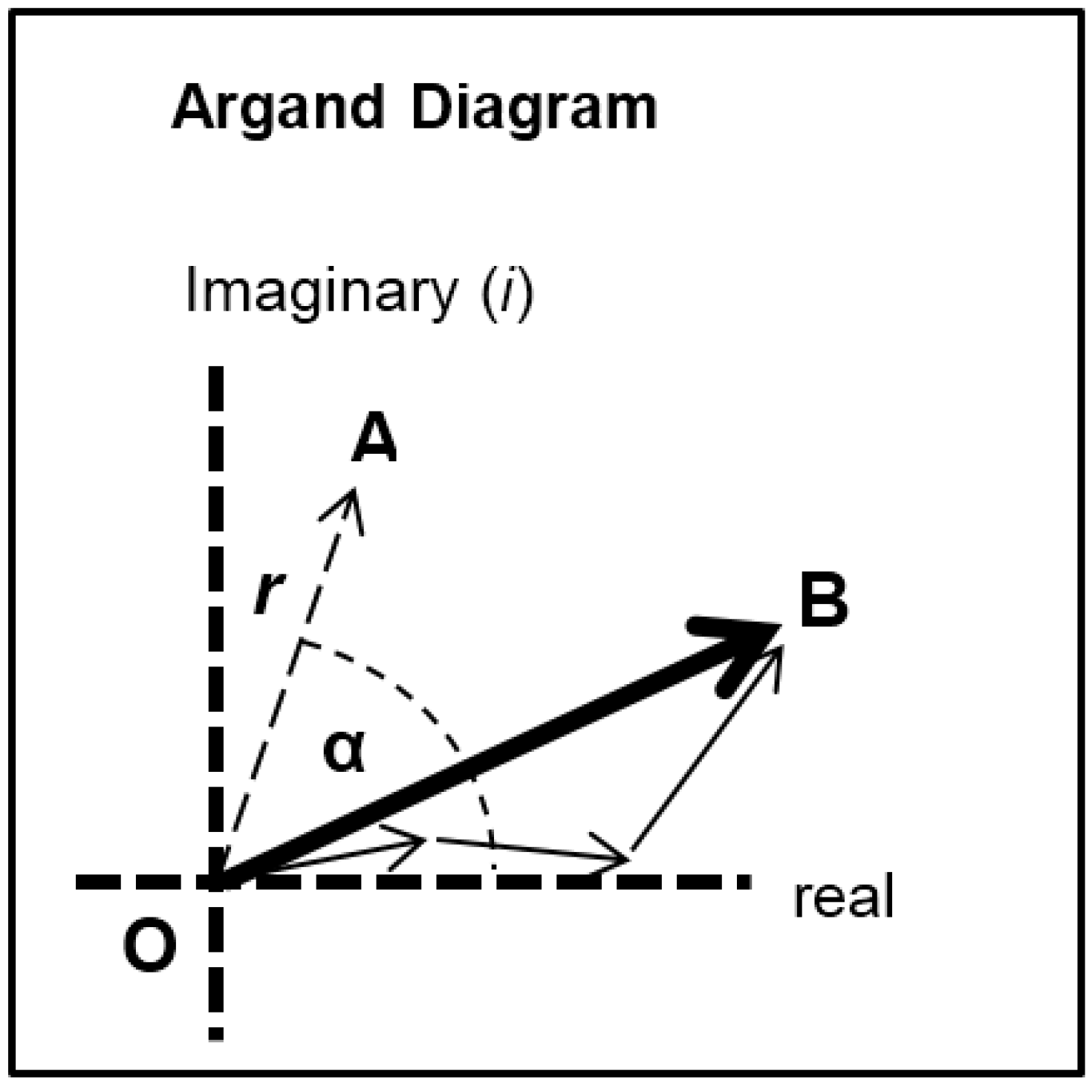
Each of the different structural states will make a specific contribution to the diffraction pattern. Let us consider some of the equatorial peaks. Each reflection will be described by the structure factor
F(
h,
k,
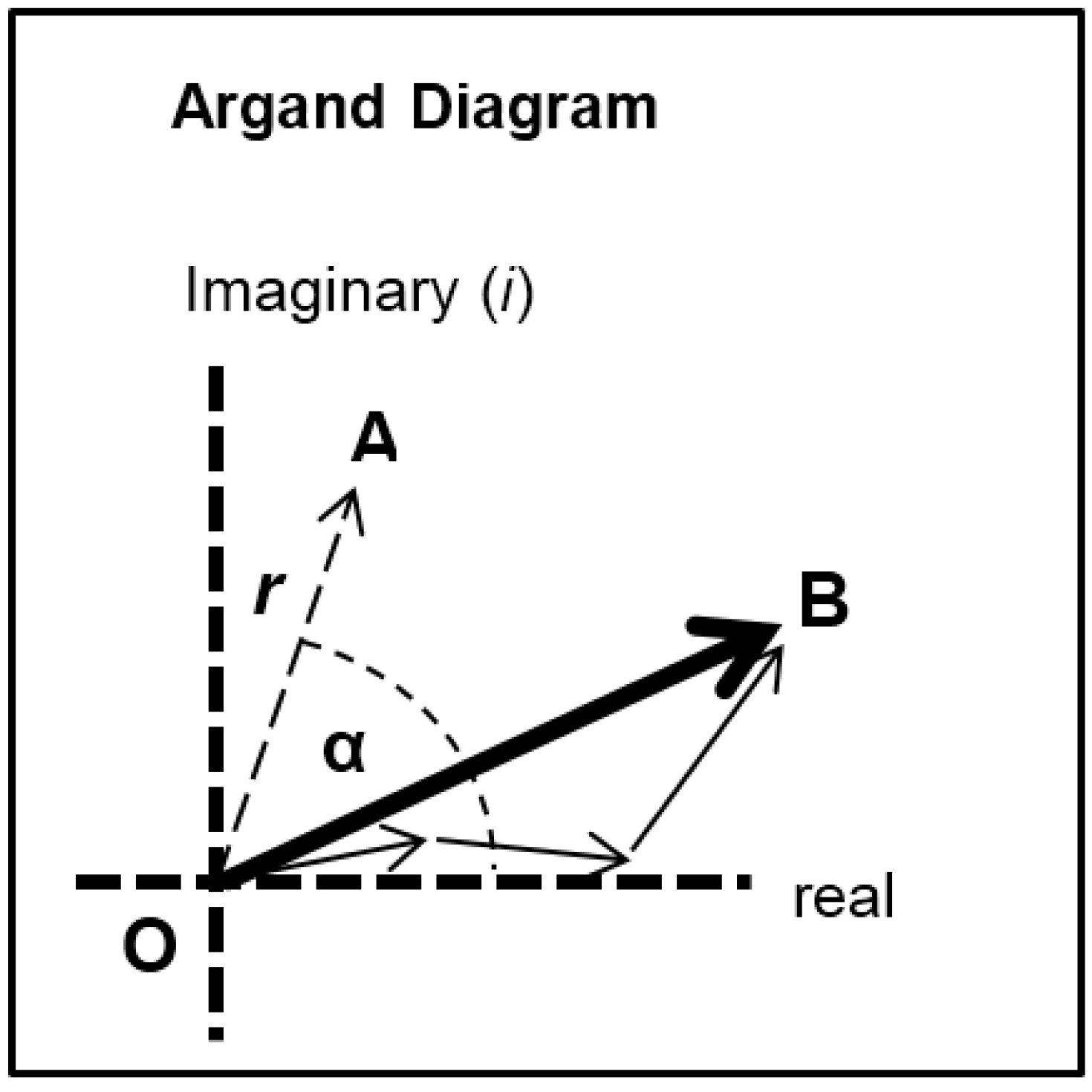
l) as in Equation (9). This is a complex quantity described by an amplitude (square root of the observed intensity) and a relative phase. This can be shown as a vector on an Argand diagram as in
Figure 7. For an arbitrary peak, the length OA in
Figure 7 represents the amplitude and the angle α with the horizontal axis represents the phase. If there is substitutional replacement of one myosin head state for another, then the structure factors for each state add vectorially as in
Figure 7 where each small arrow represents the contribution of a different state. In order to find the total resultant amplitude (OB), one needs to know the amplitudes and phases from each state and then to add them vectorially as in
Figure 7 to get the resultant OB. However, in muscle at low resolution, the muscle structure projected down the filament long axis, which the equatorial peaks tell us about, is likely to be approximately centrosymmetric. This means that for every projected density at
x,
y there is an identical density at −
x, −
y. The effect of this on the structure factor is that the phases (α) of the equatorial peaks, rather than being anywhere between 0° and 360°, can only be 0° or 180°. The corresponding vectors in
Figure 7 lie along the horizontal line (the real axis) through the origin and adding vectors is a relatively trivial exercise; the amplitudes are either added or subtracted along that line. Observations of the equatorial time-courses also suggest that the phases of the peaks do not change during the contraction; none of the amplitudes drops close to zero and then comes up again as might be expected if the contribution from one of the structures in the crossbridge cycle had a different phase (i.e., 0° instead of 180°).
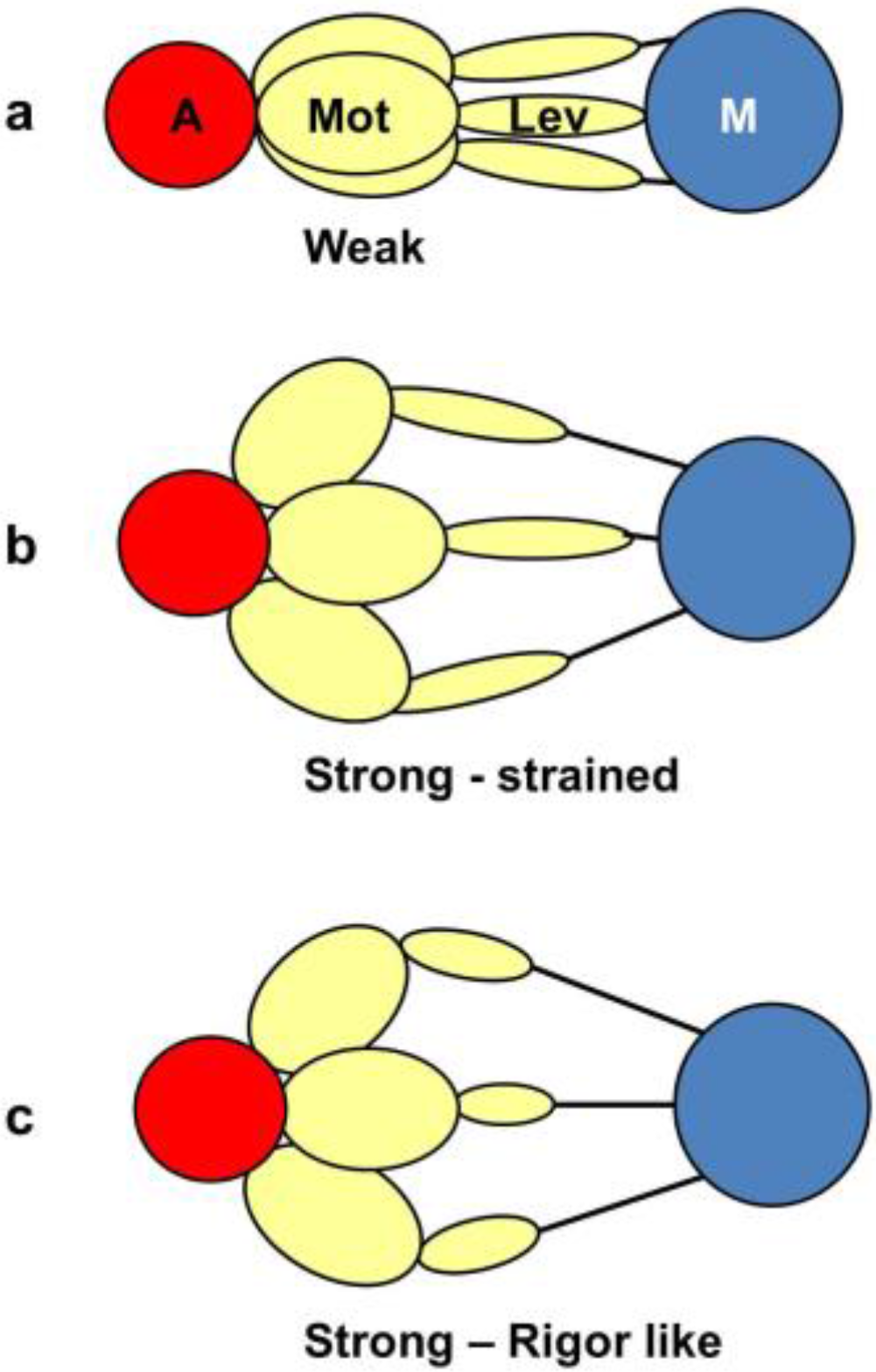
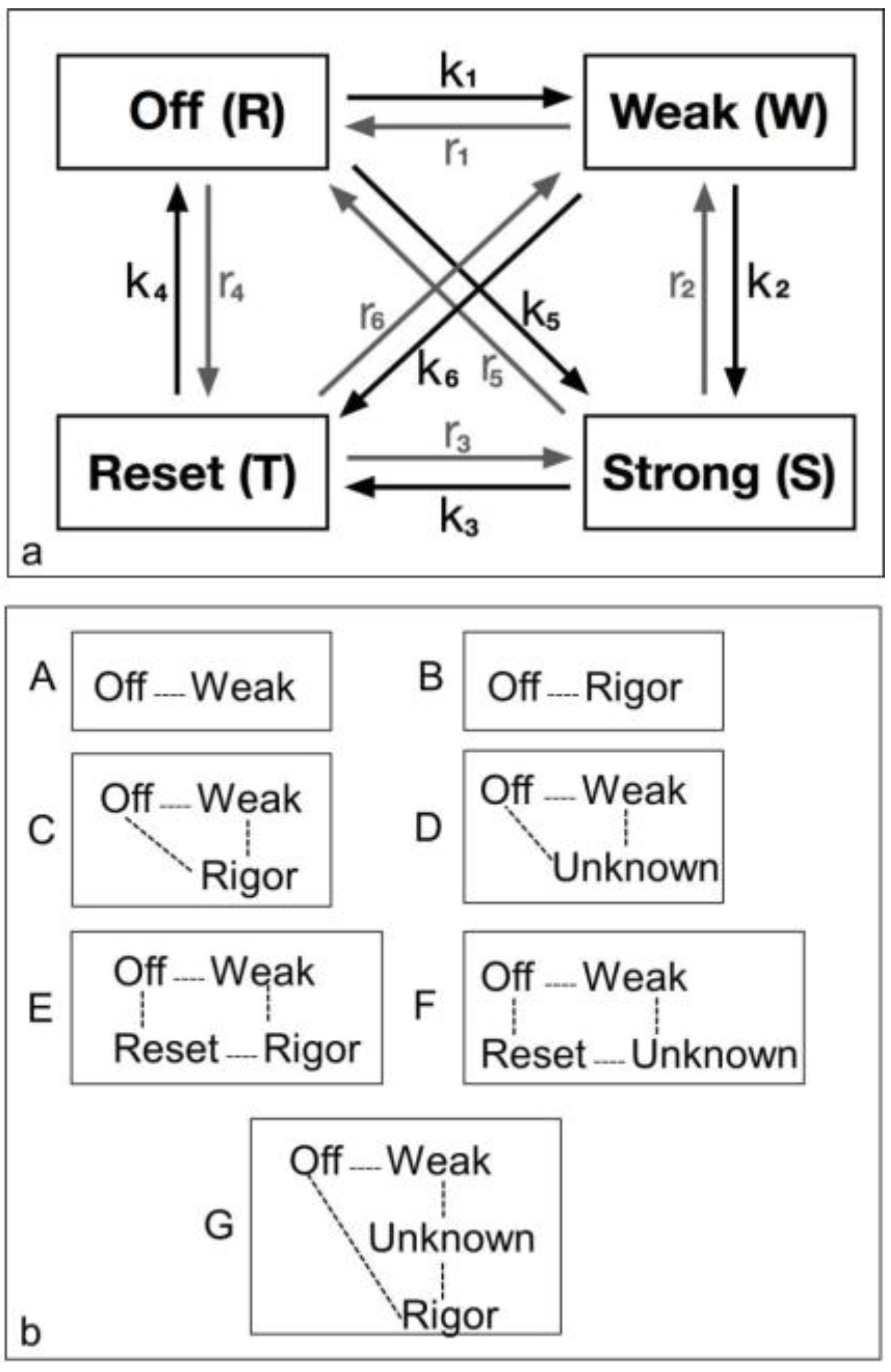
The logic of the analysis is that: (i) there might be several crossbridge states present during the rising phase of the tetanus; (ii) that each would have its own particular contribution to the observed diffraction pattern, depending on the particular structure and how many crossbridges are in that state; and (iii) that the observed amplitudes at low resolution will be a simple sum of the amplitudes (positive or negative) from each structural state. Because of this we set up a computer model consisting of several structural states with forward and backward rate constants between each (
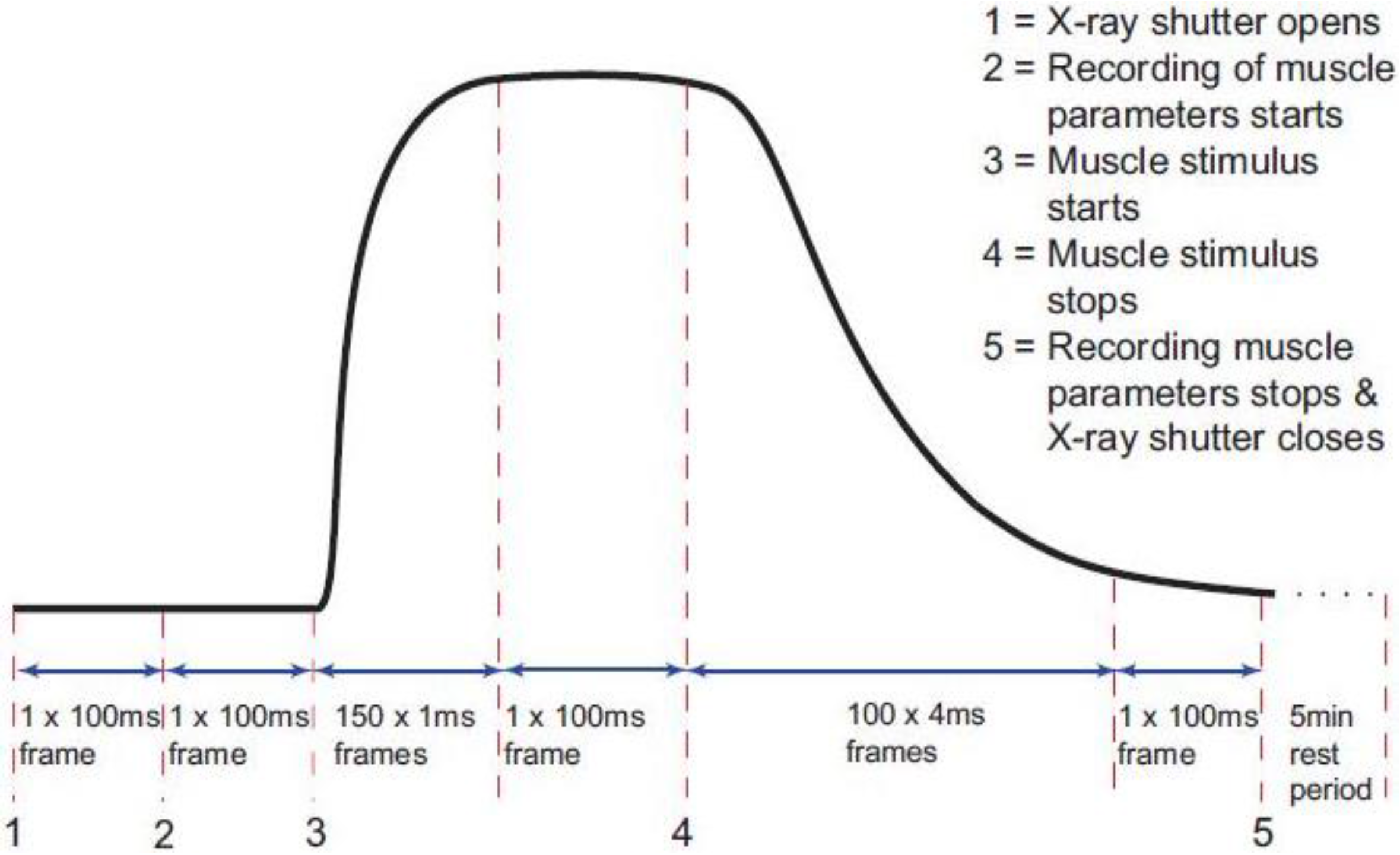
Figure 2a). We allowed the heads to start in a structure like resting muscle and to enter a cycle after activation and an appropriate delay (the latent period); the tension and the observed equatorial peaks do not change instantly when activation occurs—there is a delay (
Figure 4). With this model established we could define the structure amplitudes for some of the states (e.g., relaxed) and we could allow the computer program to use simulated annealing (see Methods) to search over other amplitude values (between limits) to give an optimal fit between the observed and calculated time-courses.
A factor to take into account in this kind of analysis is the number of observations being modelled and the number of parameters being searched over. Clearly the number of unknown parameters must be significantly less than the number of independent observations to give a reliable result. As we detail elsewhere [
88], one reason that the modelling of the interference effects on the meridian was not successful, as discussed in the introduction, was that the number of parameters being modelled was significantly more than the number of observations; the problem was under-determined.
Here we are modelling the time-courses of eight equatorial peaks and we estimate that each time-course contains at least four, possibly five, observations (the number of parameters required to model the time-course as a polynomial/exponential combination). Therefore there would be 32 to 40 observations. In parameter searches to find the crossbridge states, we need to keep the number of model parameters rather less than about 32.
Table 4 shows the results of this modelling.
4.3.1. Two-State Models
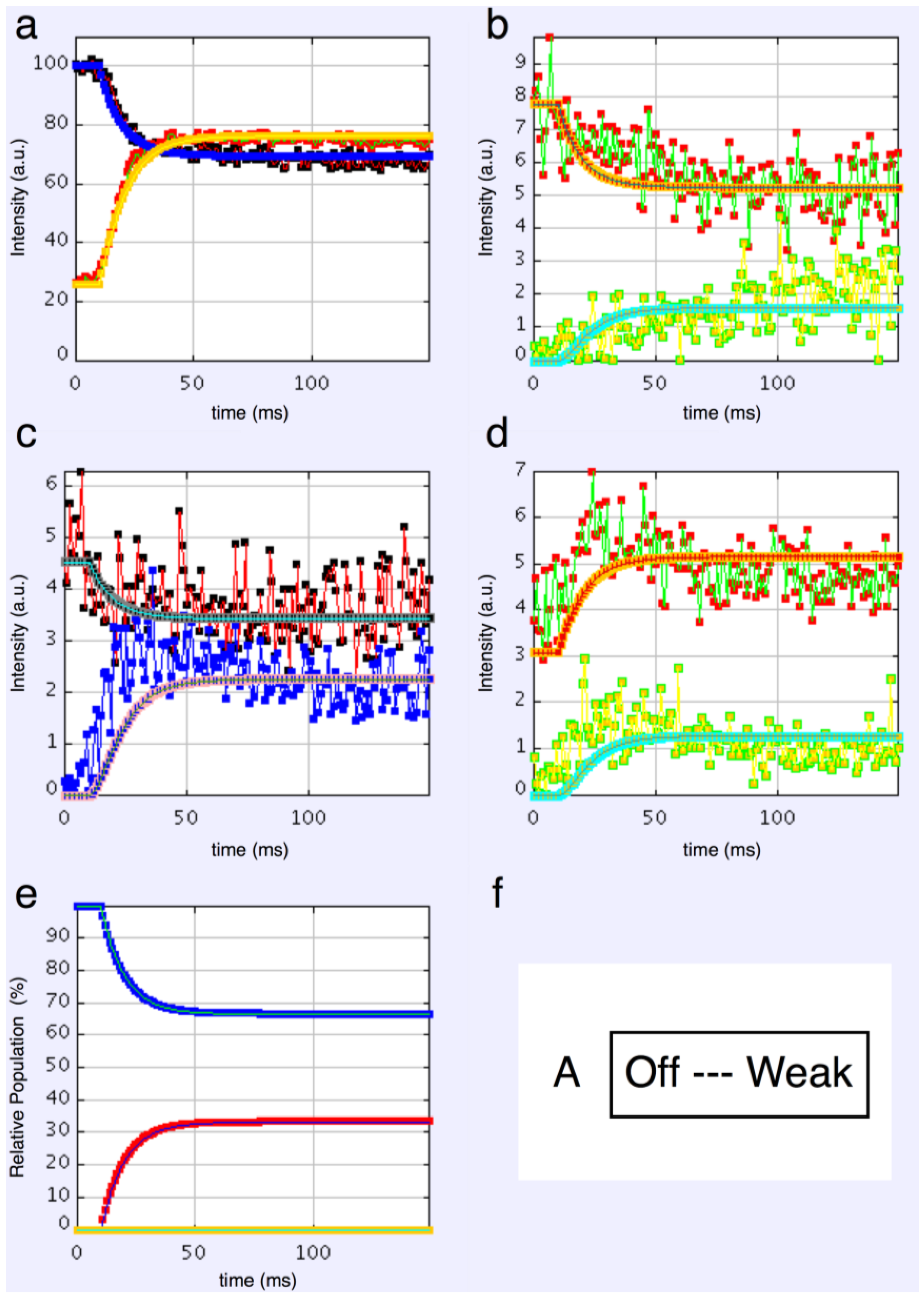
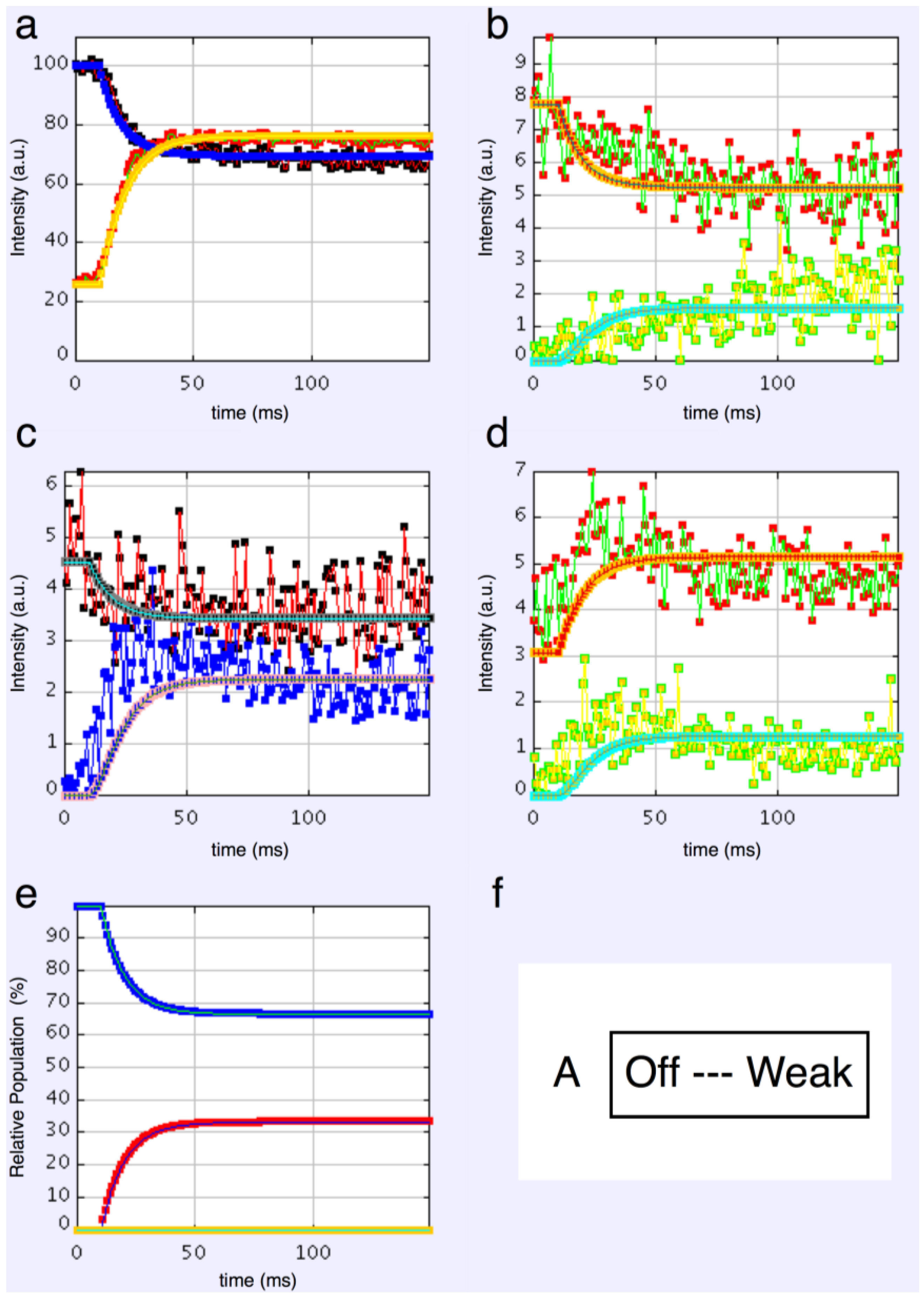
We started with simple 2-state models (Models A and B in
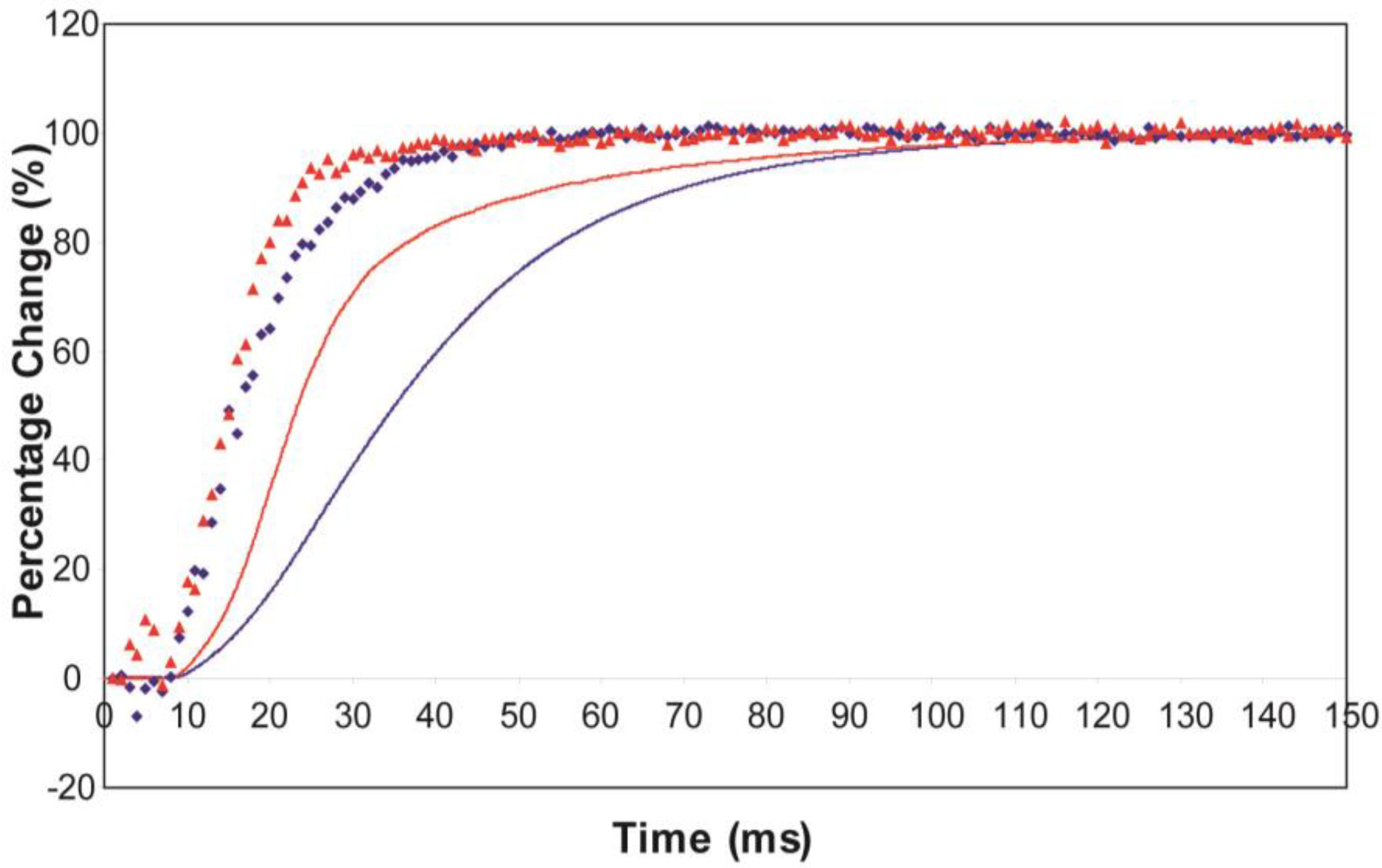
Figure 2b). The heads were either in the relaxed (off) state, or were attached and all in the same conformation. For Model A (off and weak), the number of parameters involved was: 2 rate constants, 1 delay (the latent period), and the eight unknown amplitudes of the attached structural state (11 parameters to model). The results from this very simple model were reasonable,
Figure 8, Chi = 1457 (
Table 4). Note that the modelling also gives the time-course of the changing populations of the states involved.
Here the steady state values for the active tetanus plateau were 67% detached and 33% attached. The latent period was about 10 ms. In the attached state the amplitudes are not typical of relaxed or rigor patterns; the best attached state had a very strong 11 peak (
Table 5). However, it is clear from such a simple scheme that none of the peaks will overshoot as observed—at least one intermediate attached state is needed to generate an overshoot. One can also do this calculation as a 2-state model which simply has relaxed heads as one state and rigor heads as the attached state (Model B in
Figure 2b). The agreement here was very poor indeed (Chi = 7802:
Table 4); the active structure is not a simple mix of relaxed and rigor heads.
4.3.2. Three-State Models
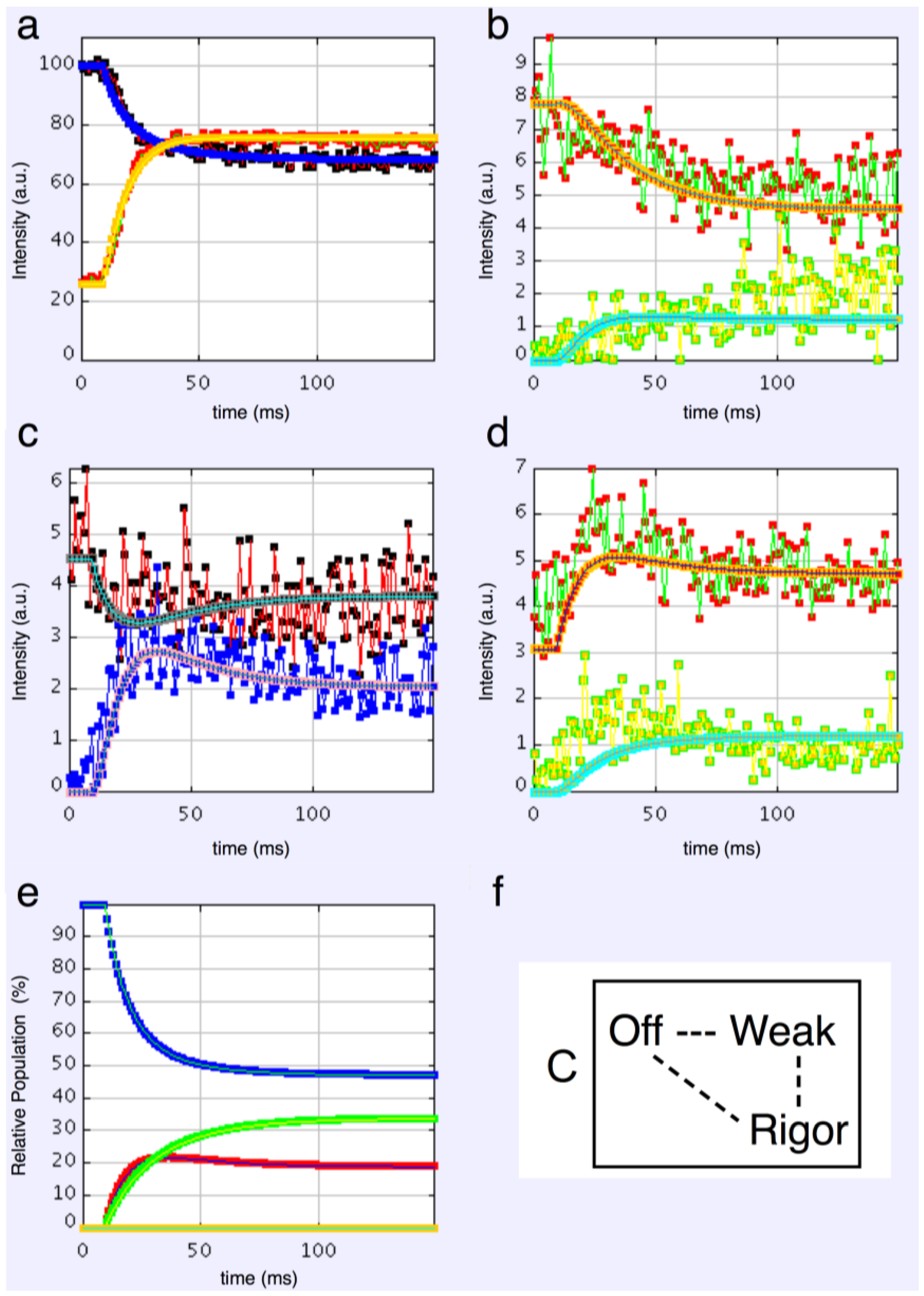
3-state models involved starting from the relaxed state and then having two actin-attached states (Model C & D in
Figure 2b). It might be assumed (Model C) from the Lymn-Taylor scheme [
3] that one attached state (the strong state in
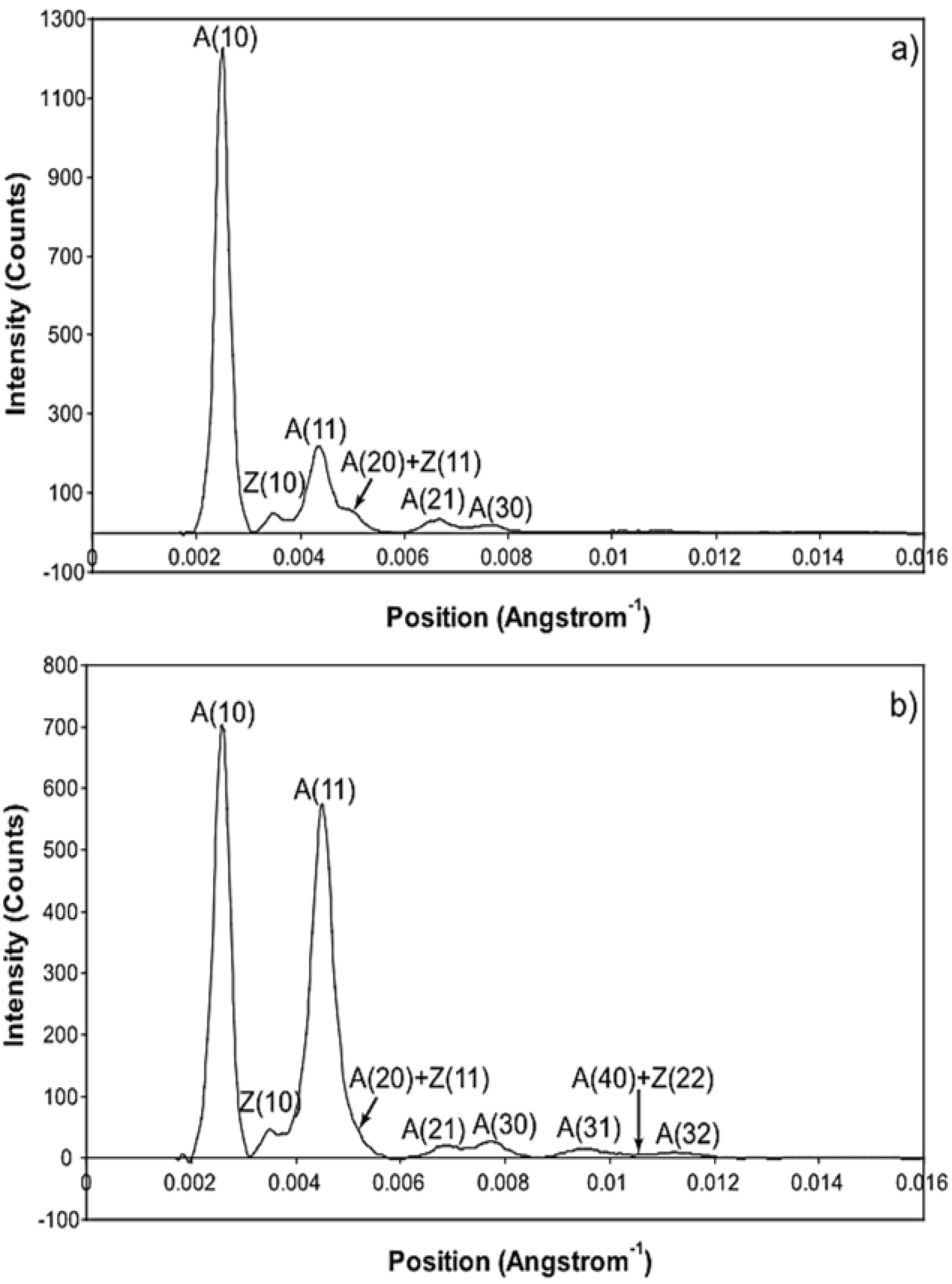
Figure 2) might possibly be like the rigor state and we know the equatorial intensities from rigor fish muscle at least out to the 30 peak (see
Supplementary Materials Figure S14). The weak state is unknown. On this basis the parameters involved are six rate constants, a delay, the eight amplitudes for the non-rigor (weak) attached state and the three outer rigor peaks (19 parameters to model). The results from this are quite good. However since we know that the 20 has a contribution from the Z-line (the Z11), the results were substantially improved if the A(20) in the rigor state was a free parameter. The final result in
Figure 9 was very good agreement, with as Chi of 1240.
If the strong structural state in
Figure 2b is not rigor (Model D), then the amplitudes from both of the two attached states are unknown. Parameters to find are six rate constants, one delay, and the eight amplitude parameters for each of the two attached states (23 parameters total). The best fits in this case were little different from the analysis with rigor in the cycle.
4.3.3. Four-State Models
At the risk of having too many unknown parameters, a fourth state can be included into the cycle. It is termed reset in
Figure 2a, but it could be either the first detached state of the heads prior to the hydrolysis step (Model E in
Figure 2b) or a third actin attached state (Model G in
Figure 2b). Is there any merit in including this fourth state? If one of the attached states is like rigor and the other two are unknown, then there are 28 parameters to fit. In this case the best Chi was 1270 (
Table 4). If the rigor state is not present and there are three unknown structures (Model F in
Figure 2b) then there are 33 parameters (really rather too many) and the best Chi was 1298 (
Table 4), not significantly better than other Models.
4.3.4. Modelling Summary
In summary, the results of the time-course fitting are as follows:
- (1)
The observed higher order reflection time-courses can be modelled quite well.
- (2)
There must be more than two states to explain the observed overshoot of the A(11) and particularly the higher order A(30) and A(31) peaks.
- (3)
Despite the rapidly increasing number of parameters to fit, there is no compelling reason to go from three to four states. The fit only changes marginally on going from 19 to 28 parameters or 23 to 33 parameters (
Table 4), where one might expect a substantially better fit if the extra step is important.
- (4)
Including rigor as one of the attached states gives as good results as anything else.
- (5)
The “weak” state that needs to be added, whether with 2-, 3- or 4-state models, always has the same sort of transform. The A(11) and other intensities in the rigor pattern are not enough to give the observed overshoots. The weak state (
Table 5;
Figure 10a,b) has an exceptionally strong A(11) reflection.
We have assumed that the structure is approximately centrosymmetric, giving equatorial phases of 0° or 180° only. Vertebrate striated muscle myosin filaments themselves have a somewhat 3-fold structure as shown by AL-Khayat et al. [
89], but the crossbridge arrays in the two halves of a single thick filament will have opposite hands when projected down the filament axis and there is also a roughly 40° rotation between equivalent crowns on each side of the bare zone, Luther et al. [
48].
Thus, at the resolution that we are considering the thick filaments are likely to appear approximately centrosymmetric in projection. However, note, finally, that even if the phases are off by a few degrees from 0° to 180°, this calculation is still valid. For example, a 5° error in each phase would in the worst case scenario change the sum of the amplitudes of a particular peak (OB in
Figure 7) in the 4-state model from 4 ×
R to 4 × cos(5) ×
R = 3.985
R, where
R is the amplitude from each state; a change of 0.375%. (Note that in the worst case scenario the Rs from the different states would all be the same). An average 10° phase error would in the worst case give 3.939R instead of 4, an error of 1.5% in OB. A 5% error in OB would arise from an average worstcase phase error of 18.2°. If there are fewer states then the error would be proportionately less.
With the modelling completed and the conclusion reached that a 3-state model including rigor is as good as anything in explaining the observations, there remain some obvious questions: (i) What is the “weak” state like? (ii) What are the rate constants around the cycle? (iii) What are the populations of the attached heads at the tetanus plateau? (iv) How much tension is generated by heads in each of the different attached structural states?
4.4. The Weak State
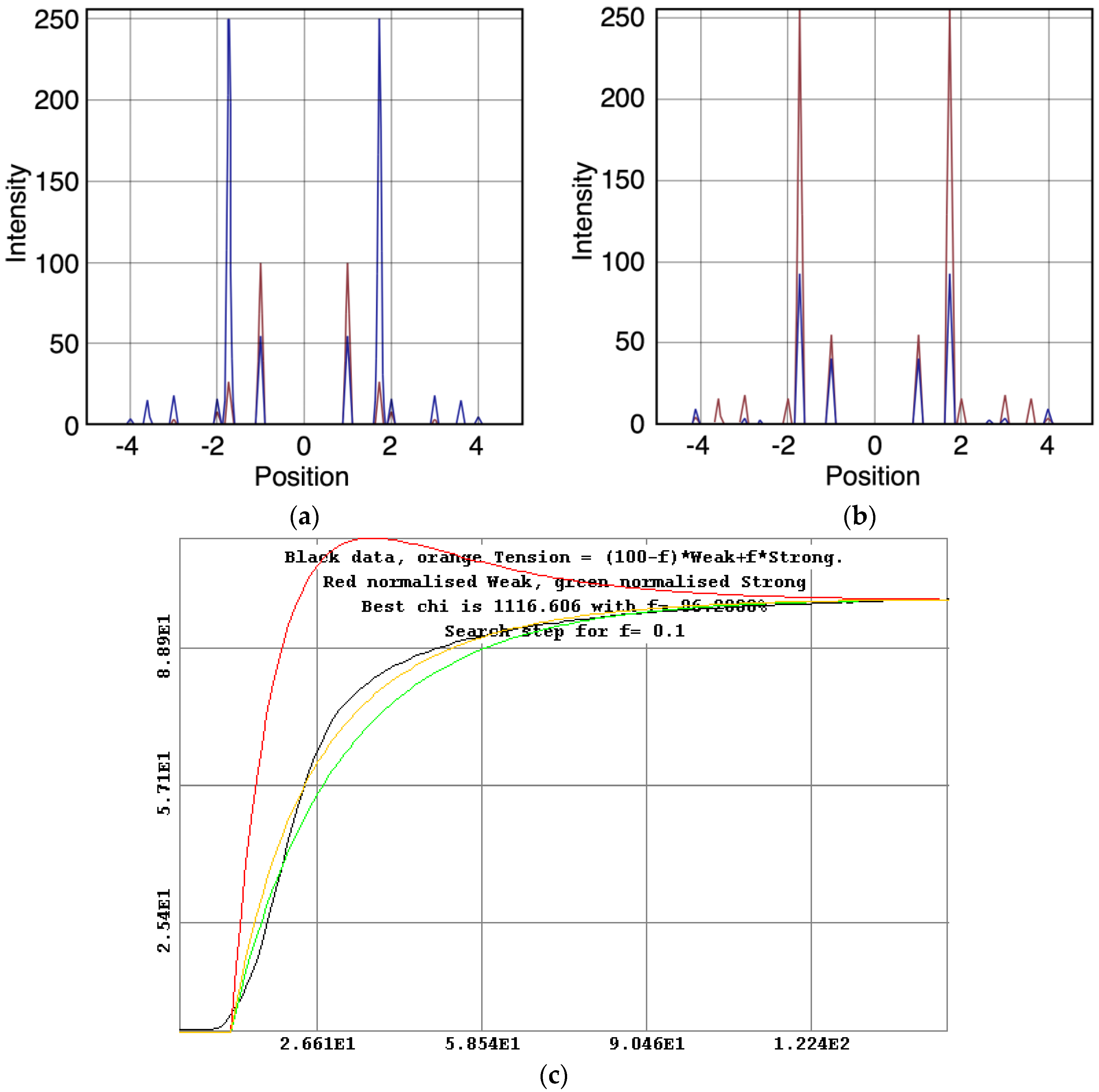
What is immediately apparent from
Table 5 is that, whatever model is chosen, in order to generate the observed higher order overshoot, the diffraction pattern from the weak state is always pretty much the same.
Table 5 compares the best modelled amplitudes from all of the Models that include a weak state, takes the average and calculates the standard deviation.
The consistency throughout is notable, so what is this state? The known structure with the nearest diffraction pattern to this is the weak binding state induced in rabbit psoas muscle fibres at low ionic strength (20 mM) observed by Brenner et al. [
64]. This state was characterised by them and in mechanical studies by, for example, Shoenberg [
90] as a relaxed state in the sense that ATP was present, but the muscle was not activated, and yet the heads were undergoing very rapid attachment-detachment steps such that the muscle stiffness rose rapidly from being low in relaxed and normal ionic strength fibres to gradually higher as the ionic strength was lowered if the stretches used to measure stiffness were fast enough.
In our case, the ionic strength is normal, but a state similar to the rapid equilibrium state of Brenner et al. [
64] appears to be present, indicating that such a state may be part of the normal contractile cycle of myosin heads on actin that increases in abundance when the muscle is activated.
4.5. The Rate Constants
The rate constants in the best models are summarised in
Table 4. For our preferred 3-state model including rigor, the off to weak rate constants,
k1 and
r1, are both quite large at 29.7 and 66.3 per second, consistent with a rapid equilibrium as in the weak-binding state. The transition from weak to strong structural states is a much slower process with
k2 at 7.5 and
r2 insignificant. The strong to off state is faster once again with
k5 and
r5 being 17.2 and 28.2, respectively. Although we are looking at structural states, which do not necessarily relate to different biochemical states (see later discussion), it appears that the rate limiting step is the weak to strong (force-producing) transition, as might be expected in an isometric contraction.
Of course, it is quite likely that some of the steps we are considering have rate constants that are strain-dependent. What we are modelling are the best simple rate constants to generate the observed variations in amplitude of the X-ray reflections. The fit is good, suggesting that the strain-dependence might not be too great in the quasi-equilibrium situation found in isometric contractions.
4.6. The Population of States
The modelling necessarily includes the time-courses of the populations of the different states. These populations are summarised in
Table 4. For the 2-state model not including rigor the two populations are 67% off and 33% attached. For the more realistic 3-state model including rigor, the off population is 48%, the weak population 20% and the rigor population 32%. However, as discussed below, these populations should not be taken to be the proportions in different biochemical states. It is interesting to note that the results from the paper of Wu et al. [
91] using a totally different approach with insect flight muscle tomograms came to the conclusion that 53% of the heads bind to actin with 29% in the strong conformation, which is very similar to our own conclusions.
4.7. The Tension Distribution and the Crossbridge Mechanism
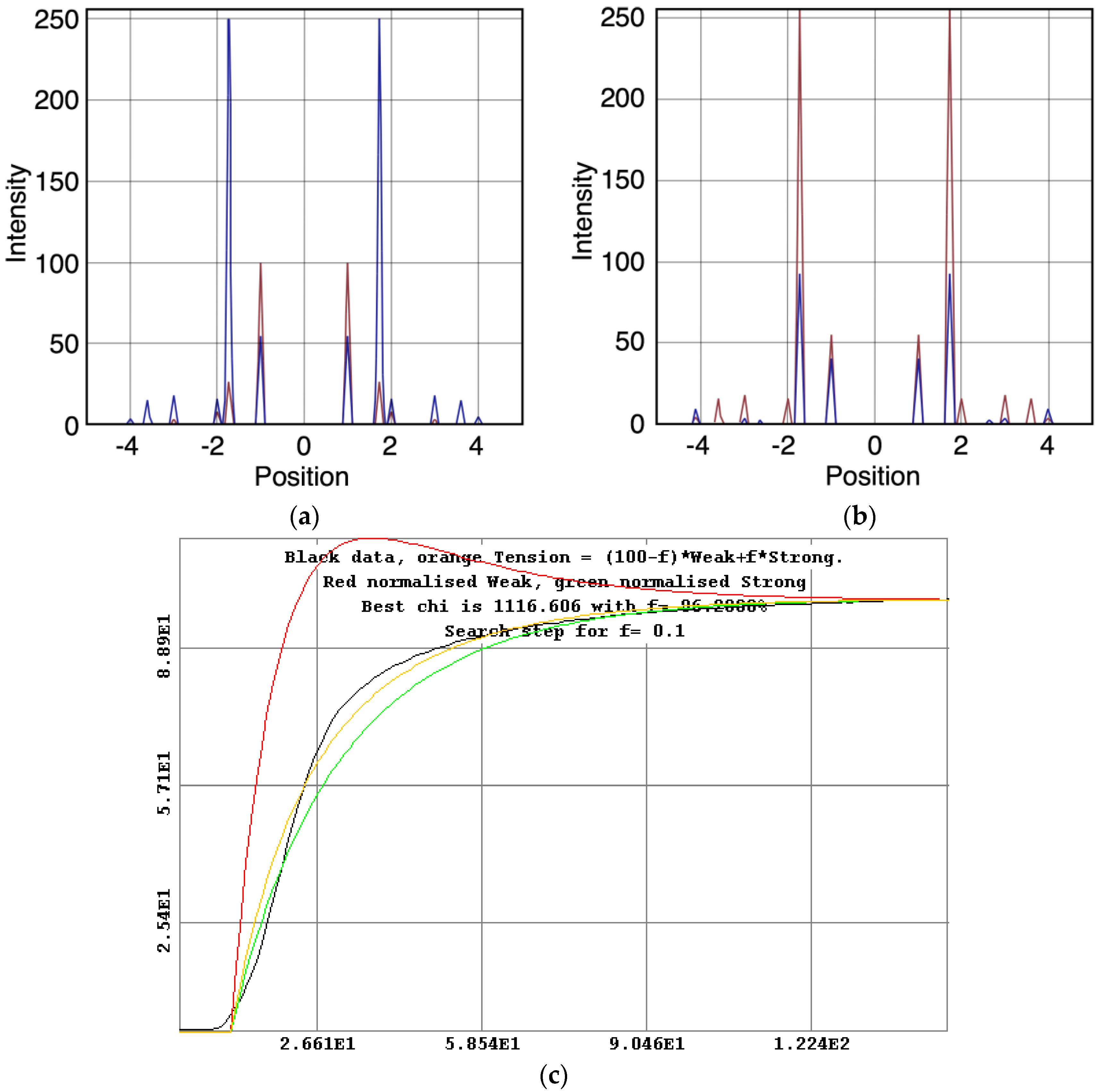
Since we know the time-course of the change in isometric tension and we know the time-courses and populations of the various states, it is possible to assign to each state a parameter (i.e., percentage
f) representing the tension associated with that state and then to optimise the sum of the individual state tensions against the observed total tension to find out how much each state contributes.
Figure 10c shows this for the 3-state model including rigor (Model C in
Figure 2b). Here the best fit is obtained as
f × strong and (100 −
f) × weak. The best fit to the observed tension is obtained if the rigor state provides (
f =) 96% of the tension and the weak state (100 −
f =) 4%, after a very clear overshoot in its time-course.
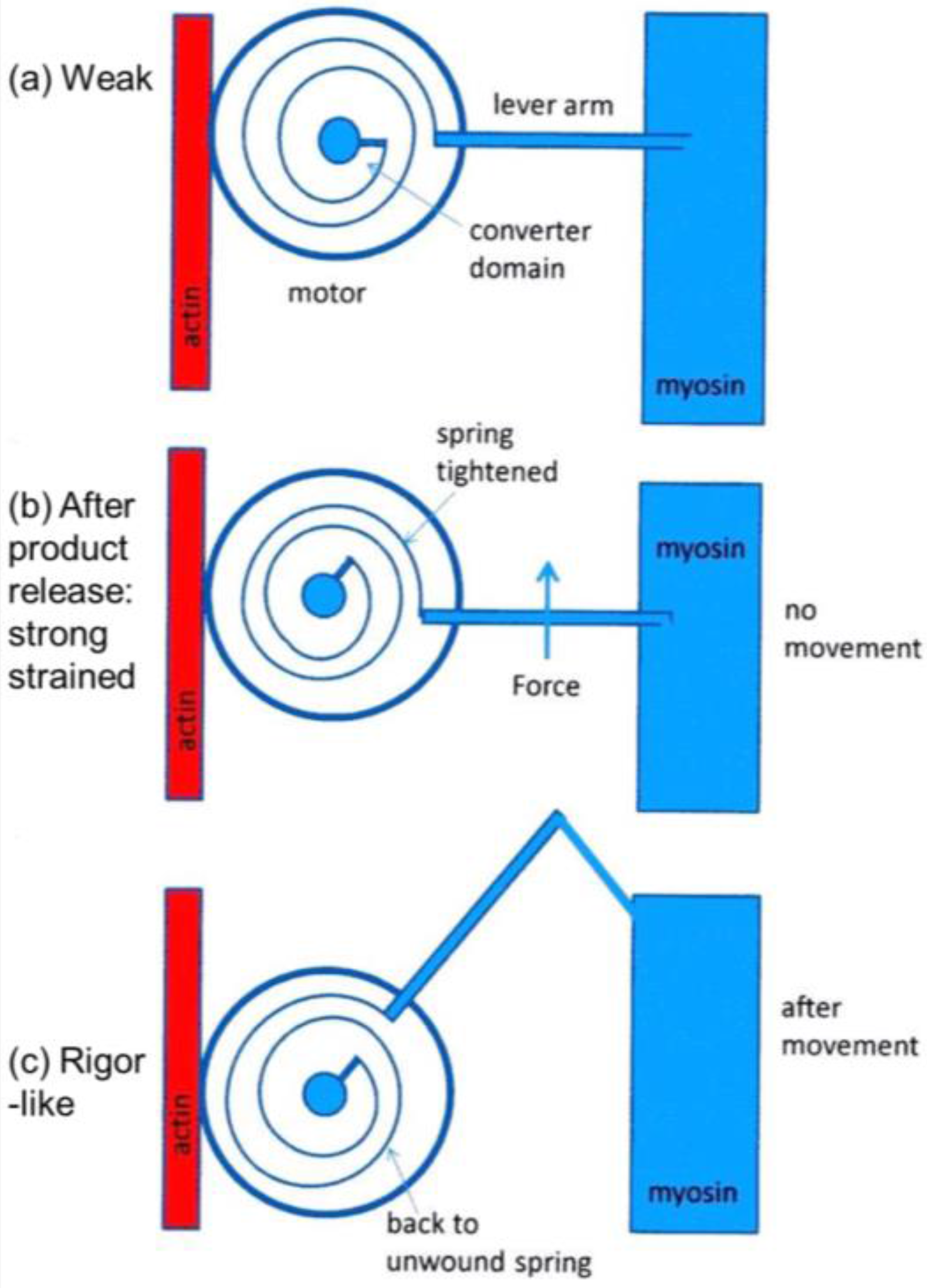
It was mentioned earlier that the structural states that we see are not necessarily correlated one-to-one with the known biochemical states in the crossbridge cycle. The reason for this can be seen in the kind of crossbridge mechanism illustrated schematically in
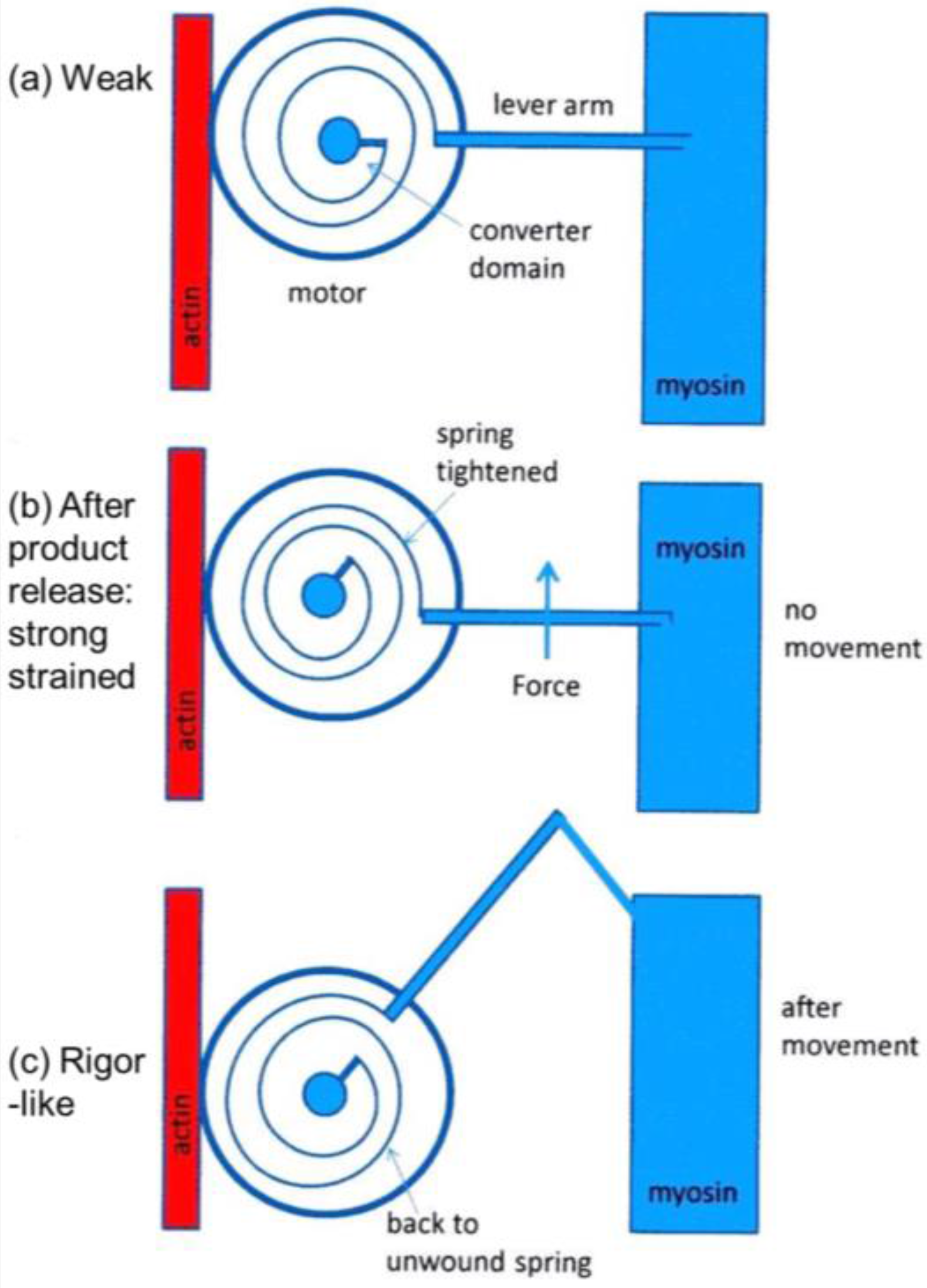
Figure 11. Here the myosin head is depicted as having elasticity (compliance) in the form of an internal spiral spring. In the weak state (
Figure 11a) the spring is assumed to be in its unstressed position. This would correspond to the weak state AM.ADP.Pi. With the release of products (Pi and perhaps ADP), the resulting internal structural rearrangement in the head would tighten the spring (
Figure 11b) to generate force. However, at this point, the lever arm need not be in a different place from where it is in the weak binding state in
Figure 11a. It will exert force between the filaments, but may not have moved. It will be a strained strong state. If the elasticity of the myosin is all in the head and not in S2, then the lever arm will only move if there is relative axial movement of the filaments (
Figure 11c), ending in the rigor conformation of the lever arm. If there is filament sliding and lever arm rotation, then the internal spring would gradually become unwound again, until at the end of the stroke the force between the filaments is zero.
From a side view, as in a longitudinal section, the weak state in our modelling would appear as a mixture of the states in
Figure 11a,b. We would expect some of those heads not to produce force (
Figure 11a) and some to produce force after product release (
Figure 11b) and these two states might look the same when viewed from the side as in
Figure 11. However, we are looking at the equator of the pattern; the view of the structure projected down the fibre axis. What would happen there?
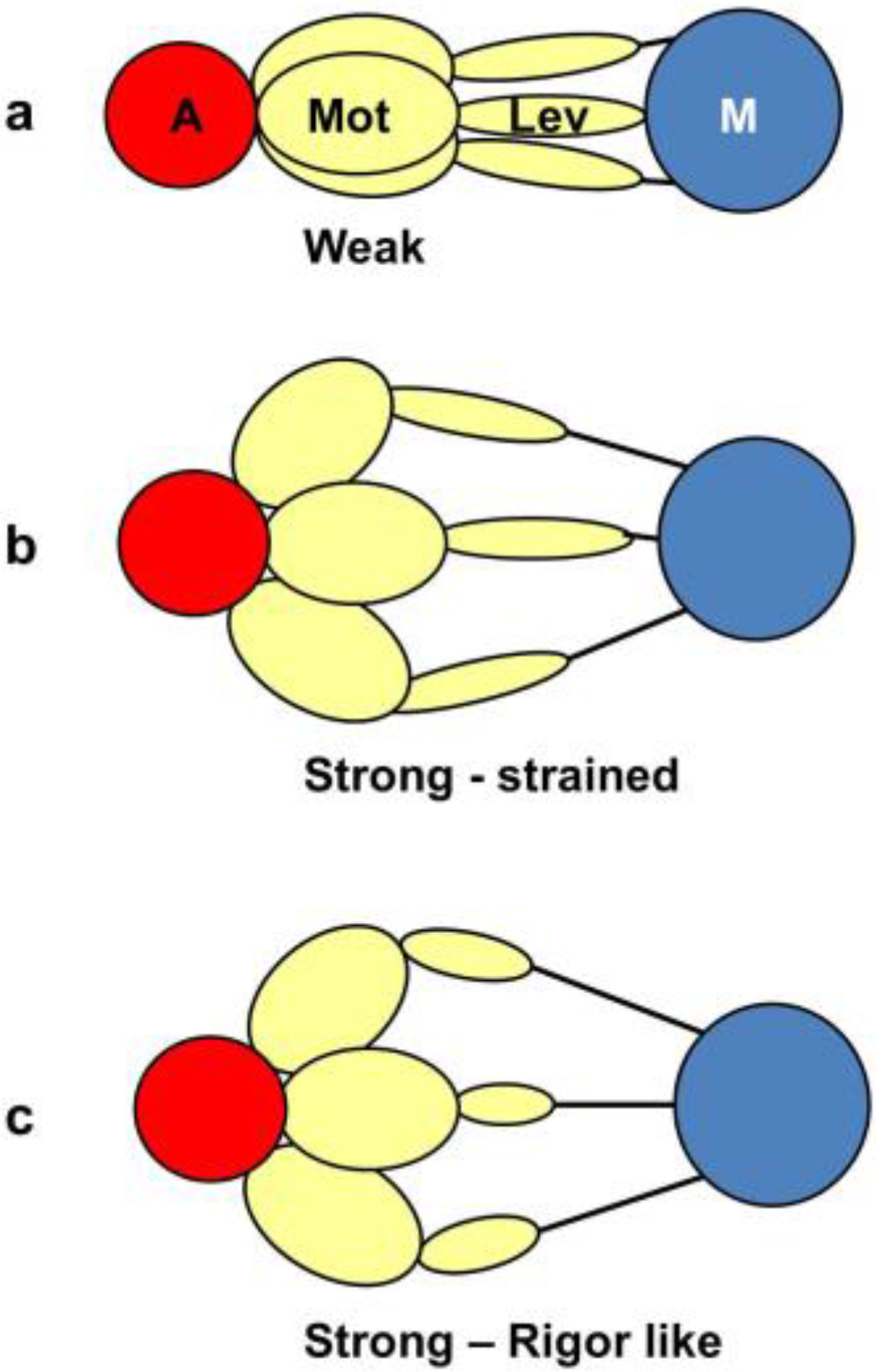
Figure 12 shows what might happen in projection down the axis. We would imagine that a weak state would be characterised by heads that attach non-stereospecifically to actin in such a way that they all point back to where they came from on myosin. In this way they would be lying along the 11 planes in the lattice and would give the observed strong A(11) reflection.
At some point in the cycle, after release of products, the heads would become stereospecifically attached to actin and because of the varying azimuths of the actin monomers the initial strongly bound heads would become more azimuthally distributed around the actin filament axis (
Figure 12b), no longer all pointing along the 11 planes. Thus, the A(11) reflection would be weaker, as in the rigor pattern. Filament movement and lever arm swinging would then result in a foreshortened lever arm in projection down the filament axis (
Figure 12c), but this would make little difference to the equatorial diffraction pattern. In other words, the states in
Figure 12b,c would look rather similar on the equator because of the relatively small mass of the lever arm compared to the motor domain. As discussed in the Introduction, we previously [
41] came to the same conclusion that the lever arm would make little contribution to the meridional diffraction pattern at low resolution because of its small mass.
What we observe as the rigor state will be a mixture of the initial stereospecifically-bound but strained high tension state in
Figure 11b, but viewed down the axis (
Figure 12b) and the final rigor-like (low tension) state (
Figure 11c and
Figure 12c) and everything in between. Thus, our results show for the 3-state model (C in
Figure 2b) that 48% + 20% of the heads are in the off and weak-binding states, that 32% of the heads are in the first stereospecific and rigor-like states and that these 32% of heads together produce about 96% of the tension. Note that in our previous analysis of the meridional diffraction pattern [
41] and the effects of length steps, we also concluded that the motor domains, which dominate the diffraction patterns, would have 70% (off and weak) not moving with the step and 30% (strong) moving as the filaments move. The two sets of results are closely compatible.
5. Conclusions
We have shown that the Plaice fin muscle is a viable preparation to use for recording time-resolved low-angle X-ray diffraction pattern from active whole muscle with sarcomere length control. Suitable equipment for recording and control of SL has been developed and used successfully. The tension in the rising phase of end-held tetanic contractions and the intensities of the many of the equatorial reflections all change much faster with partial SL control than without, but there is still evidence for a myosin head state at an early stage of the rising phase which has the characteristics of rapid equilibrium actin attachment, but where the force level is still low (see also Tanaka et al. [
92]). We have shown that: (i) a 3-state model including rigor fits the data as well as any other model; (ii) there is no need to have more than two structurally distinct attached states when viewed on the equator, although each state may include more than one biochemical state; (iii) we have defined the relative populations of states in this model; and (iv) we have determined the contribution that the structural states make to the total tension.
The analysis done here tests whether the observed changing equatorial intensities can be modelled as a sum of known or unknown contributions. If they are unknown it defines what these might be. One of the known patterns is that of rigor muscle and this seems to work pretty well as part of the cycle. We go on to discuss what the results might mean in terms of models in
Figure 11 and
Figure 12. What these show is that the rigor state that we are modelling appears to be showing the conformation and azimuthal distribution of the motor domains of the attached heads in the strong state. The lever arms in this scheme can be tilted away from the rigor conformation (
Figure 11b) where they would produce tension, or have tilted to the rigor position (
Figure 11c) where no tension would be produced. Both of these head conformations would look similar on the part of the equator that we are studying because of the relatively low mass of the lever arm.
We are now in a position to start to analyse the whole recorded 2D diffraction pattern in more detail in order to further refine myosin head behaviour in active fish muscle.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}