
Amino Carbonylation of Epidermal Basement Membrane Inhibits Epidermal Cell Function and Is Suppressed by Methylparaben


1
Bionics Program, Tokyo University of Technology Graduate School, Tokyo 192-0982, Japan
2
School of Bioscience and Biotechnology, Tokyo University of Technology, Tokyo 192-0982, Japan
*
Author to whom correspondence should be addressed.
Cosmetics 2017, 4(4), 38; https://doi.org/10.3390/cosmetics4040038
Submission received: 29 July 2017
/
Revised: 4 September 2017
/
Accepted: 21 September 2017
/
Published: 25 September 2017
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:This study investigated the effect of amino carbonylation (Maillard reaction) on the function of the epidermal basement membrane (BM) by analyzing epidermal cell proliferation and keratinization and stratum corneum barrier function using a three-dimensional human epidermal BM model treated with glyceraldehyde. Intracellular ATP levels were lower in cells cultured on amino-carbonylated epidermal BM as compared to those in normal epidermal BM (control). Moreover, trans-epidermal water loss was increased by culturing on amino-carbonylated BM relative to the control; this was accompanied by downregulation of filaggrin, transglutaminase-1, and serine palmitoyltransferase 2 mRNA levels. p-Hydroxybenzoic acid methyl ester (methylparaben) abrogated the decrease in ATP production and filaggrin expression in human keratinocytes induced by amino-carbonylated collagen. Thus, amino carbonylation of the epidermal BM inhibits moisture retention, keratinization, and ceramide synthesis and disrupts the barrier function of the stratum corneum. These findings suggest that methylparaben can be an effective additive to cosmetics for improving epidermal function that is compromised by amino carbonylation.
1. Introduction
Aldehyde groups in reducing sugars are modified by and form crosslinks with the amino groups of lysine or arginine residues in proteins, leading to alterations in the three-dimensional structure, physical properties, and activity of the latter. This amino carbonyl reaction is known as glycation or the Maillard reaction and generates advanced glycation end products (AGEs) via oxidation, dehydration, and condensation [1]. Collagen is a structural protein that is a component of all organs including the skin, blood vessel walls, and bones, and is subject to amino carbonylation [2]. AGEs are generated not only from glucose but also from its metabolic intermediates and degradation products as well as from Maillard reaction intermediates. AGEs derived from glyceraldehyde—a metabolic intermediate of glucose—have been linked to the development and progression of some diseases [3,4].
AGEs accumulate during aging, and are especially abundant for proteins with a slow turnover, such as collagen [5]. Studies of skin AGEs have mostly focused on the dermis, which has a high collagen content. Collagen fibers have a triple-helical structure and maintain the visco-elasticity of the skin, which is lost when the fibers are crosslinked as a result of the amino carbonylation of lysine and arginine residues in collagen [6], leading to skin hardening and wrinkling [7,8]. Nε-(carboxymethyl) lysine (CML) is a typical AGE in the skin that does not form crosslinks or exhibit fluorescence [9]. Accumulation of AGEs in the dermis gives the skin a yellowish color [10], causes nuclear factor-κB activation, and induces inflammation [11,12]. AGEs also promote melanin synthesis [13]. Aminoguanidine [14], N-phenacylthiazolium bromide [15], pyridoxamine [16], cerivastatin [3], metformin [17], pioglitazone [17], pentoxifylline [17], longistatin [18], quercetin [19], plantamajoside [20], and N-acetyl-l-cysteine [21] have been shown to inhibit AGE formation.
Decreases in the amounts of natural moisturizing factor (NMF), intracellular lipid, and ceramide in the stratum corneum decreases moisture and the barrier function of the stratum corneum, resulting in dry skin [22,23]. Filaggrin (FLG) is a protein involved in NMF production and water retention in the stratum corneum [24], which consists of thick cell membranes that are lined with an intracellular insoluble membrane, known as a cornified cell envelope (CE), that strengthens the cell membrane [25,26]. Transglutaminase-1 (TGase1) is involved in cornification [27,28] and catalyzes the cross-linking of proteins by condensing a primary amine of lysine to the amino group of glutamine and transferring a substituent on the former to the latter [29,30]. Omega-hydroxy ceramides attach to the outside of CEs and are required for cornification. Thus, both CEs and ceramides play important roles in maintaining the normal barrier function of the stratum corneum [25,31,32]. Ceramides are synthesized by enzymes such as serine palmitoyltransferase 2 (SPTLC2), which catalyzes the condensation of l-serine and palmitoyl-CoA during the keratinization of epidermal cells. Sphinganine is subsequently converted to sphingomyelin and glucosylceramide, which are then reconverted to ceramides by β-glucocerebrosidase (βGCase) and acid sphingomyelinase (aSMase) [33].
The present study investigated whether collagen in the epidermal basement membrane (BM) is a target of amino carbonylation and how this affects epidermal cell proliferation and keratinization and stratum corneum barrier function using a three-dimensional (3D) human epidermal BM model. We evaluated 34 types of cosmetic material for their ability to inhibit AGE production. Methylparaben showed the most potent inhibition of AGEs at a concentration that was non-toxic to cultured cells. We investigated the effects of methylparaben on the function of keratinocytes cultured on amino-carbonylated collagen.
2. Materials and Methods
2.1. Materials
Human keratinocyte and culture medium for the 3D human epidermis model (LabCyte EPI-KIT; Japan Tissue Engineering Co., Tokyo, Japan) were purchased from Japan Tissue Engineering (Gamagori, Japan). The HaCaT human keratinocyte cell line was obtained from Cell Lines Service GmbH (Eppelheim, Germany). Glyceraldehyde was from Wako Pure Chemical Industries (Osaka, Japan). p-Hydroxybenzoic acid methyl ester (methylparaben) was from Ueno Fine Chemicals Industry (Osaka, Japan). Native collagen acid solution (IAC-30 Native collagen bovine dermis) was from Koken (Tokyo, Japan). Other reagents used in the experiments were obtained from Wako Pure Chemical Industries (Osaka, Japan).
2.2. Determination of Effects of Glyceraldehyde on the Relative Fluorescence Intensity Associated with Amino Carbonylation of Epidermal BM
After adding 15 μL of 40 mmol/L or 100 mmol/L glyceraldehyde solution or phosphate-buffered saline (PBS) to 285 μL of a diluted BM extract, 300 μL of the BM extract was added to a 24-well culture plate (Falcon; Thermo Fisher Scientific; Waltham, MA, USA), and was allowed to gel for 30 min at 37 °C in an incubator. The 24-well plate was incubated in a humid chamber for two, four, and six days at 37 °C in the incubator. Relative fluorescence intensity (excitation/emission, 360/460 nm) was measured with a fluorescence microplate reader (Multi-Detection Microplate POWERSAN HT; BioTek, Winooski, VT, USA). Results are expressed as the mean ± standard deviation of three experiments.
2.3. Culture of Epidermal Keratinocytes on Amino-Carbonylated BM
After adding 2.5 μL of 100 mmol/L glyceraldehyde solution or PBS to 47.5 μL of the diluted BM extract, 50 μL of the BM extract was added to a 24-well culture insert (Falcon; Thermo Fisher Scientific; Waltham, MA, USA), and was allowed to gel for 30 min at 37 °C in the incubator. The 24-well plate was incubated in the humid chamber for six days at 37 °C in the incubator. After three washes with PBS, the culture medium for the 3D epidermis was added to the 24-well culture plate and followed by incubation for 24 h. The 3D human epidermis model (LabCyte EPI-KIT) was cultured for 10 days at 37 °C in a CO2 incubator. The relative fluorescence intensity (excitation/emission, 360/460 nm) was measured with the fluorescence microplate reader. ATP measurement reagent (“Cell” ATP Assay reagent; Toyo Ink Co., LTD., Tokyo, Japan) was added, and after 30 min the relative luminescence (RLU/sec) was measured as the amount of ATP. The experiment was carried out with triplicate samples.
2.4. Measurement of Transepidermal Water Loss (TEWL) in the 3D Human Epidermis Model
The 24-well culture insert containing the epidermis was inserted into an agar plate maintained at 37 °C, and TEWL was measured using a Tewameter TM210 (Courage and Khazaka Electronic GmbH, Köln, Germany). TEWL was measured at room temperature (22 °C) and 40% humidity. Probes were well-suited for TEWL measurements on the culture insert. To obtain stable TEWL measurements, the insert was placed in a suitable hole on an agar plate warmed at 32 °C.
2.5. Measurement of FLG, TGase1, SPTLC2, βGCase, and aSMase mRNA Levels
After measuring TEWL, total RNA was isolated from the 3D model of human epidermis using an mRNA Extraction kit (Qiagen K.K., Tokyo, Japan) according to the manufacturer’s instructions. The expression levels of FLG, TGase1, and SPTLC2 were determined using a real-time reverse transcription PCR kit (Takara Bio, Otsu, Japan) on an ABI PRISM 7900HT system (Applied Biosystems, Foster City, CA, USA) according to the manufacturer’s protocols, with β-actin used as an endogenous control. Primers used to amplify FLG, TGase1, SPTLC2, βGCase, aSMase, and β-actin were purchased from Qiagen. Relative changes in mRNA expression levels were calculated with the 2−ΔΔCt method, and the levels were normalized to that of β-actin. The experiment was performed with triplicate samples.
2.6. Immunohistochemical Analysis of FLG and TGase1 Expression
The 3D model of the human epidermis was embedded in Tissue-Tek optimum cutting temperature (O.C.T.) medium (Sakura Finetek, Tokyo, Japan) and cut on a cryostat into 10-μm-thick frozen sections that were incubated in methanol for 5 min at 0 °C, dried, washed three times with PBS supplemented with 0.01% bovine serum albumin (BSA), then incubated with 10% goat serum in PBS in the humid chamber for 1 h at room temperature. After three washes with PBS supplemented with 0.01% BSA, the sections were subsequently incubated overnight at 4 °C in the humid chamber with polyclonal anti-FLG (Cusabio, College Park, MD, USA) and anti-TGase1 (Harber Bio-Product, Norwood, MA, USA) antibodies diluted 500-fold in PBS. The sections were washed three times with PBS supplemented with 0.05% Tween 20 and then incubated overnight at 4 °C in the humid chamber with the Alexa Fluor 488-conjugated F(ab’) fragment of goat anti-rabbit IgG/IgM(H+M) (1:1000) and Alexa Fluor 546-conjugated goat anti-mouse IgG/IgM (H+M) (1:1000) (Invitrogen/Life Technologies, Carlsbad, CA, USA). After washing five times with PBS supplemented with 0.05% Tween 20 and once with PBS, the sections were mounted onto slides using Fluoromount (Japan Tanner, Osaka, Japan), covered with cover slips, and examined under a fluorescence microscope (BX51; Olympus, Tokyo, Japan).
2.7. Determination of Effects of Glyceraldehyde on the Relative Fluorescence Intensity Associated with Amino Carbonylation of Collagen
Native collagen acid solution (3 mg/mL) was added to a 24-well culture plate (180 μL per well), and was allowed to gel for 30 min at 37 °C in the incubator. After adding 20 μL of glyceraldehyde solution (6.25, 12.5, 25, 50, 100, 250, and 500 mmol/L), or PBS to collagen gel-coated culture wells, the 24-well plate was incubated in the humid chamber for four days at 37 °C in the incubator. Relative fluorescence intensity (excitation/emission, 360/460 nm) was measured with the fluorescence microplate reader. Results are expressed as the mean ± standard deviation of three experiments.
2.8. Measurement of ATP Content
Native collagen acid solution (3 mg/mL) was added to a 96-well plate at 90 μL/well. After incubation for 1 h at room temperature, 10 μL of glyceraldehyde (500 mmol/L) was added followed by incubation for three days. After three washes with PBS, 100 μL of 2% fetal bovine serum (FBS) containing Dulbecco’s Modified Eagle’s Medium (DMEM) was added followed by incubation for one day to remove residual glyceraldehyde. A 100-μL volume of HaCaT cells (100,000 cells/mL) in DMEM containing 2% FBS was added to the well followed by incubation for one day at 37 °C in the CO2 incubator. The solution was replaced with 100 μL/well of DMEM containing 2% FBS (control) or DMEM containing 2% FBS and methylparaben (0.05, 0.1 and 0.2 mmol/L) and the cells were cultured for four days. ATP measurement reagent (“cell” ATP assay reagent) was added, and after 30 min the relative luminescence (RLU/sec) was measured as the amount of ATP. The experiment was carried out with triplicate samples.
2.9. Measurement of FLG mRNA Levels in Cultured HaCaT
A 35-mm dish was coated with native collagen acid solution (0.5 mL for 1 h). After washing with PBS, the collagen was amino carbonylated by treatment with 50 mmol/L glyceraldehyde solution for three days at 37 °C in the CO2 incubator. After three washes with PBS, 1 mL of DMEM was added and the incubation was carried out for one day. After removing the DMEM in the well, 500 μL of HaCaT cells (300,000 cells/mL) were seeded in each well. After incubation for one day, the solution was replaced with 2% FBS-DMEM or 0.1 mmol/L methylparaben containing 2% FBS-DMEM. After culturing for two days at 37 °C in the CO2 incubator, RNA was extracted and evaluated as described above. The experiment was carried out with triplicate samples.
2.10. Data Analysis
mRNA and ATP levels are expressed as mean ± standard deviation. An unpaired t test was performed to compare group means; p < 0.05 was considered statistically significant.
3. Results
3.1. Effects of Glyceraldehyde on the Relative Fluorescence Intensity of Amino-carbonylated BM and Effect of Amino Carbonylation of the BM on TEWL
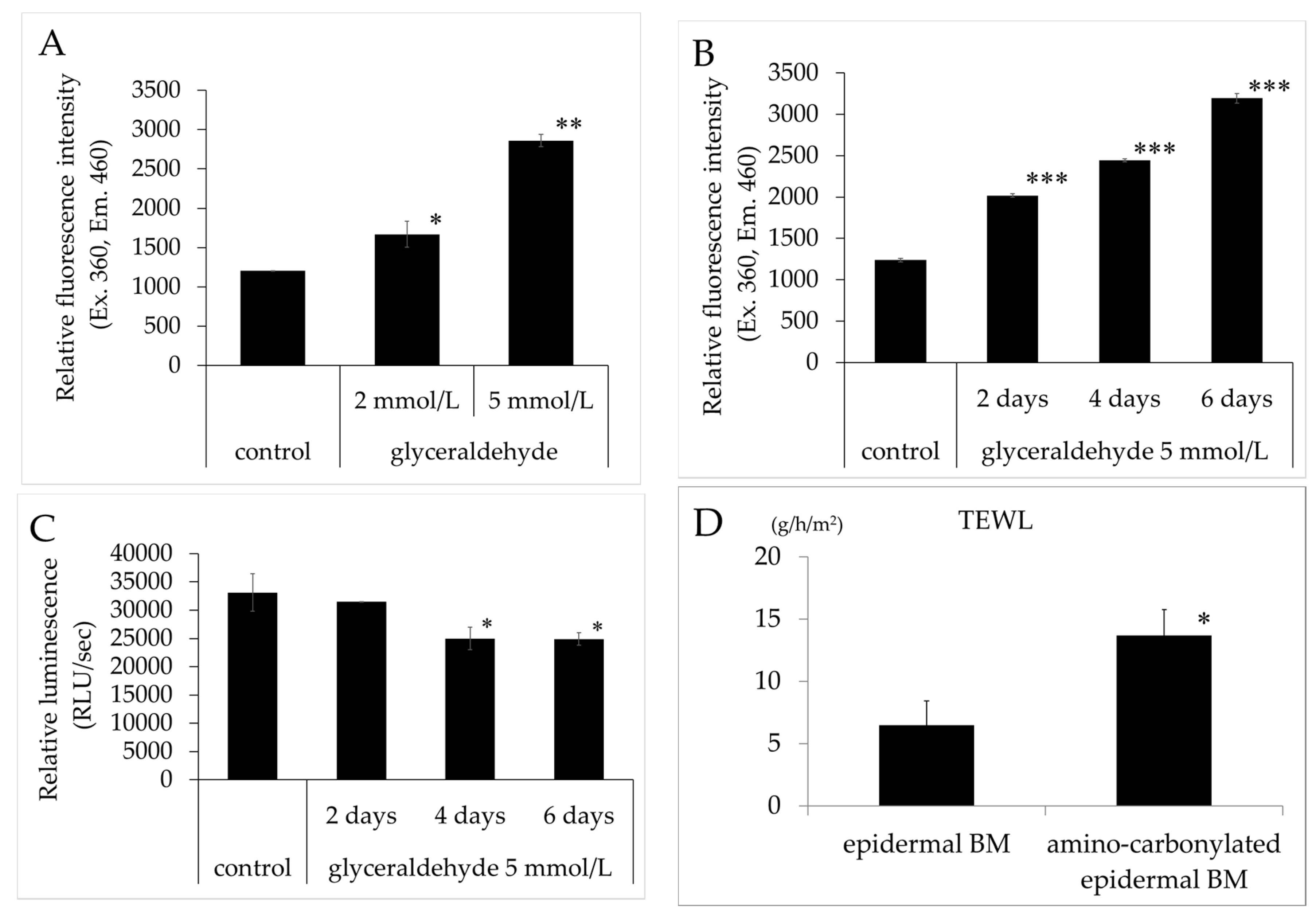
The relative fluorescence intensity of BM treated with glyceraldehyde (2 and 5 mmol/L) was significantly increased concentration-dependently compared to that of the untreated control (Figure 1A). Furthermore, the relative fluorescence intensity of BM treated with 5 mmol/L glyceraldehyde (two, four, and six days) was significantly increased time-dependently compared to that of the untreated control (Figure 1B). ATP levels were lower in keratinocyte cultured on BM treated with 5 mmol/L glyceraldehyde (two, four, and six days) than in cells grown on untreated collagen gel (Figure 1C). TEWL in the 3D model of human epidermis was increased relative to that in the control when the BM was treated with glyceraldehyde for four days (Figure 1D), indicating that the barrier function of the epidermis was compromised.
3.2. Effect of Amino Carbonylation of BM on the mRNA Expressions of FLG, TGase1, SPTLC2, GCase, and aSMase
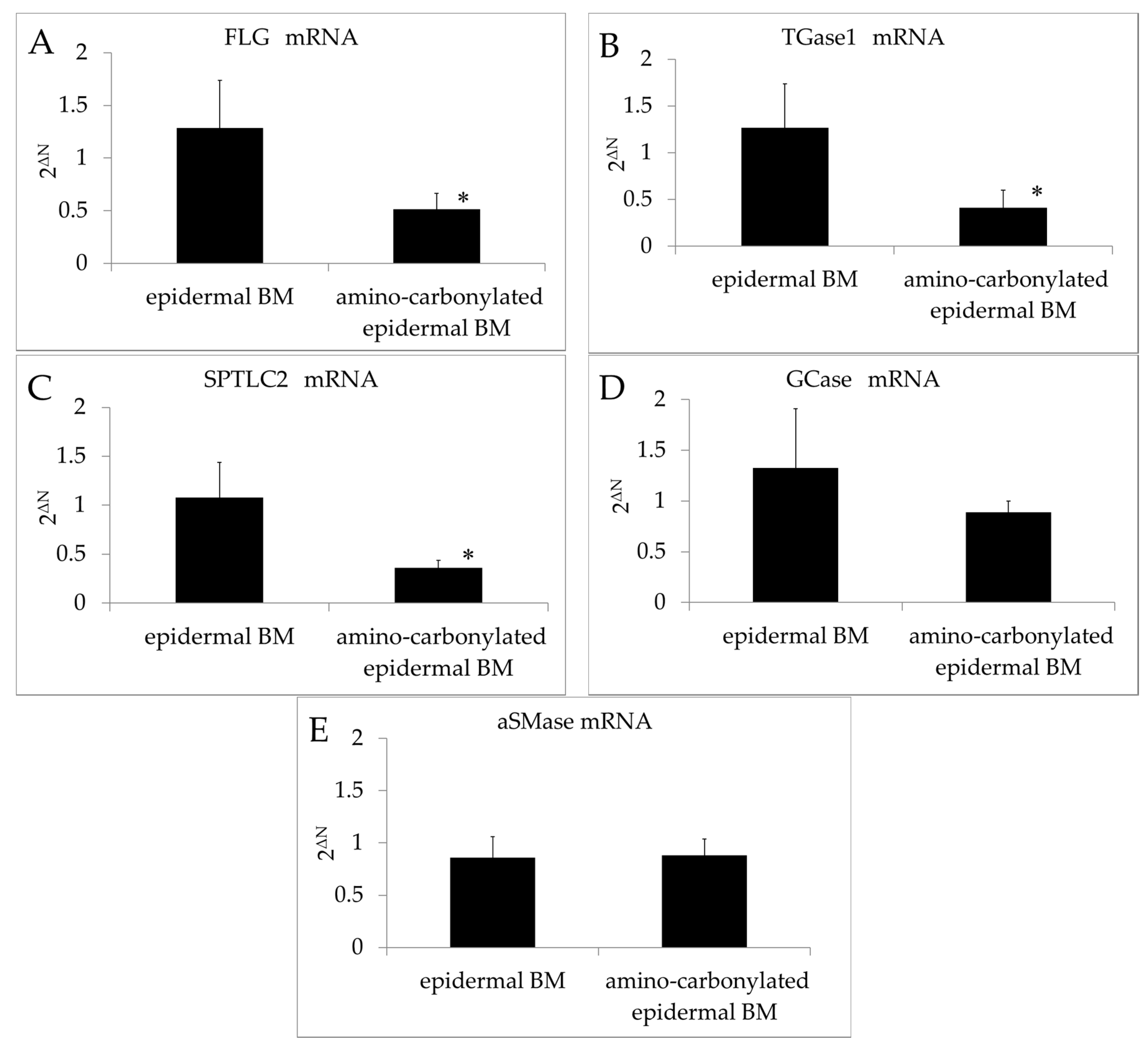
The FLG and TGase1 transcript levels in the 3D model of the human epidermis were downregulated relative to those in the control upon BM amino carbonylation (Figure 2A,B). Filaggrin is an important protein in the stratum corneum that participates in the production of NMF and is a determinant of water-retention capacity [24]. The cell membrane in the stratum corneum is thicker than that in other layers of the skin, a lining known as the cornified cell envelope (CE) on the inner side of corneocytes, suggesting that the CE is involved in cell membrane reinforcement. TGase1 is required for the formation of the CE in corneocytes, which are found in the stratum corneum. Thus, amino carbonylation of the BM decreased the moisturizing function of the stratum corneum and prevented of cornification.
3.3. Effect of Amino Carbonylation of the BM on FLG and TGase1 Protein Levels
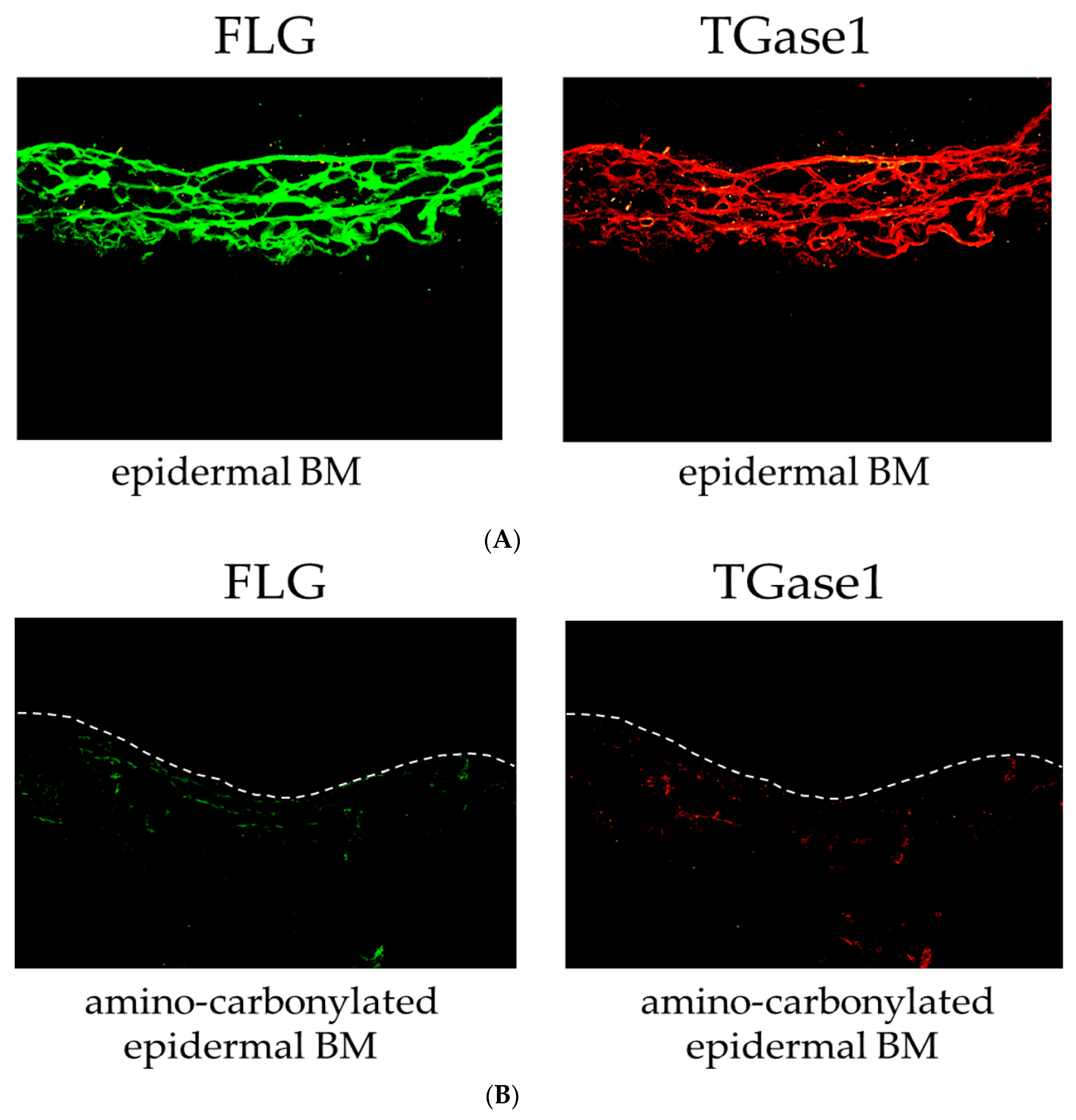
We also examined the effect of the amino carbonylation of the BM on the protein levels of FLG and TGase1 in the 3D model of the human epidermis by immunohistochemistry. FLG and TGase1 were primarily localized to the stratum corneum layer of the epidermis cultured on the epidermal BM (normal BM) (Figure 3A). The 3D model of the human epidermis cultured on the amino-carbonylated BM showed decreased FLG and TGase1 protein levels and was labeled along with the stratum corneum layer of the epidermis cultured on the BM treated with glyceraldehyde (Figure 3B).
3.4. Effects of Glyceraldehyde on the Relative Fluorescence Intensity of Collagen Gel
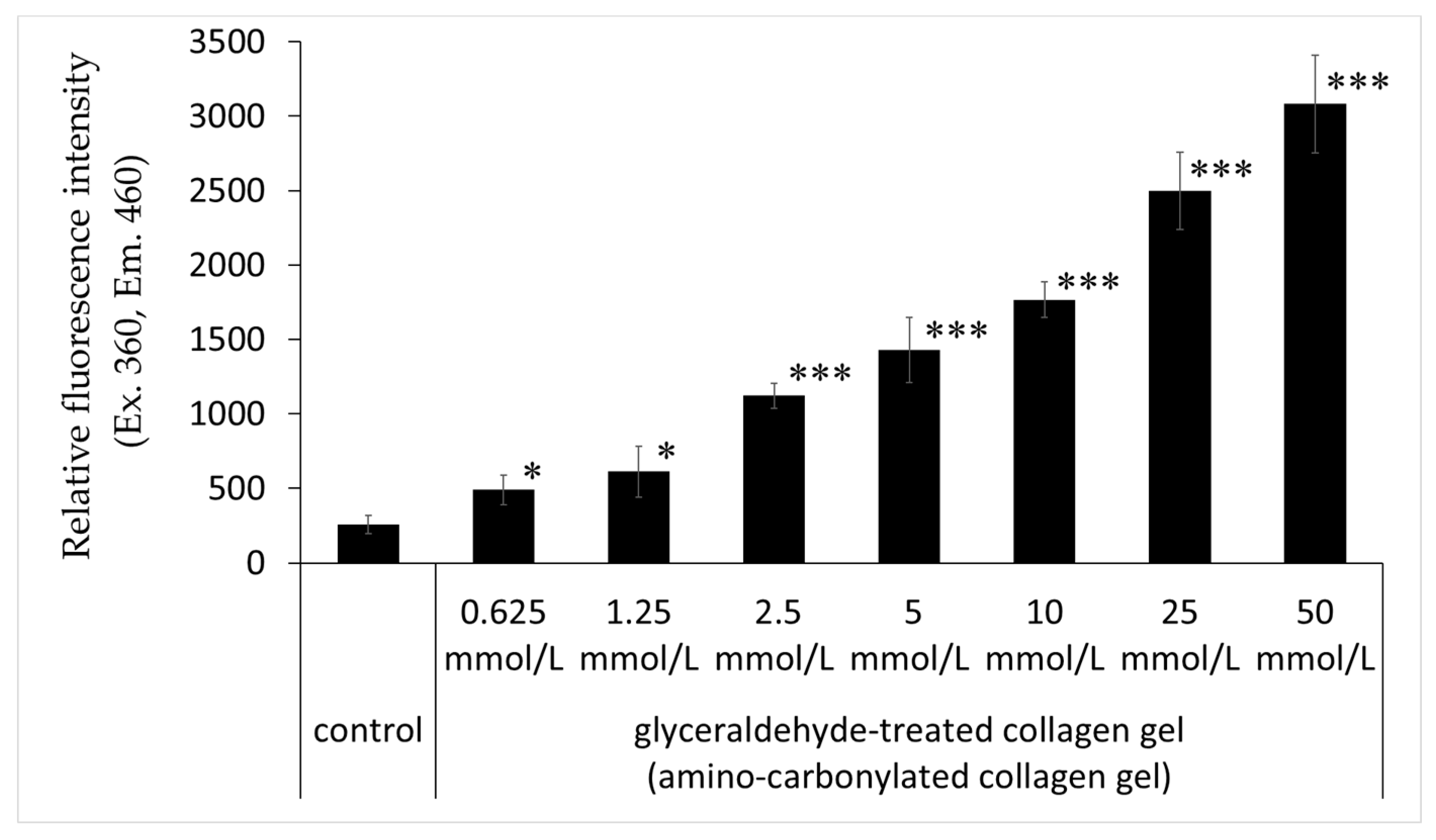
The relative fluorescence intensity of collagen gel with treated glyceraldehyde (0.625, 1.25, 2.5, 5, 10, 25, and 50 mmol/L) was significantly higher than that of the untreated control; this effect was concentration-dependent (Figure 4).
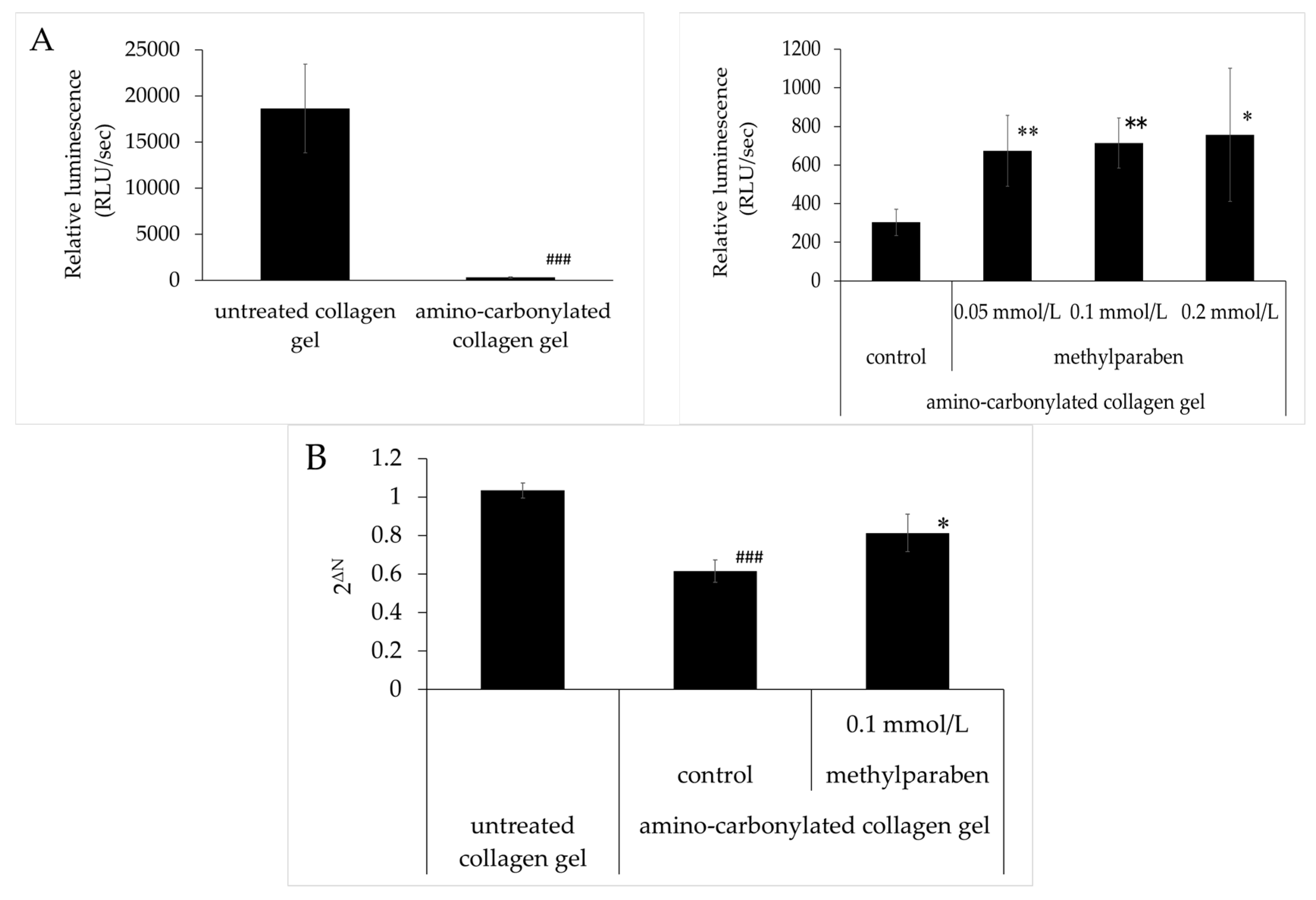
3.5. Effect of Methylparaben on the Decrease in ATP and FLG mRNA Levels Caused by Amino Carbonylation of Collagen
ATP levels were lower in HaCaT cells cultured on amino-carbonylated collagen gel than in cells grown on untreated collagen gel (Figure 5A). The inhibitory effect of the amino carbonylation of collagen on ATP levels was abrogated by treatment with methylparaben at concentrations of 0.05, 0.1, and 0.2 mmol/L. These effects were significant relative to the control and were concentration-dependent. A similar trend was observed for filaggrin mRNA levels (Figure 5B). These decreases were abolished by treatment with methylparaben at a concentration of 0.1 mmol/L.
4. Discussion
Various amino-carbonylated proteins are generated via the non-enzymatic reaction of aldehydes derived from glucose and lipids [1]. Although most amino-carbonylated proteins are degraded by the proteasome in the cell, collagen fibers that undergo amino carbonylation accumulate in the extracellular matrix, given that the half-life of collagen fiber in the skin is as long as 14.8 years [5]. If this leads to inflammation and matrix metalloproteinase activation, the amino-carbonylated collagen will be degraded, returning the skin to its original state. In the present study, we found that the ATP content in epidermal keratinocytes was reduced when the cells were cultured on amino-carbonylated collagen. Accumulation of amino-carbonylated collagen in the epidermal BM implies that the metabolic activity of mitochondria in epidermal keratinocytes is reduced. Moreover, TEWL in the 3D model of the human epidermis cultured on amino-carbonylated BM was increased, which was associated with reduced mRNA levels of filaggrin, TGase1, and SPTLC2. These results suggest that amino carbonylation of epidermal BM undermines the stratum corneum, specifically by decreasing its water retention capacity and barrier function.
Accumulation of AGEs has been detected in various tissues during aging and diabetes, including in collagen, skeletal and vascular smooth muscle, and the BM [34]. Furthermore, it has been reported that laminin and type IV collagen, which are important BM components, were non-enzymatically glycosylated in vitro [35]. Epidermal BM consists of lamina lucida, lamina densa, and lamina fibroreticularis layers on the epidermis side; keratinocyte hemidesmosomes in the epidermal basal layer are connected to type XVII collagen in the lamina densa via the lamina lucida. α6/β4 Integrin in the keratinocyte membrane binds the glycoprotein laminin and associates with type IV collagen in the lower lamina densa, which bind to reticular fibers (consisting of type III collagen) of connective tissue through anchoring fibrils composed of type VII collagen. We speculate that amino carbonylation of laminin, and type XVII and IV collagen in the BM affects epidermal function, but cannot rule out the possibility that it alters the structure and function of epidermal BM proteins.
CML combined with collagen is known to induce apoptosis in human skin fibroblasts [36]. Pentosidine—a cross-linking agent that exhibits fluorescence in AGEs produced from ribose, arginine, and lysine—is also present in the collagen of the dermis, reportedly at higher levels in patients with diabetes than in age-matched healthy subjects [37]. Advanced glycation end products of the glyceraldehyde-related Maillard reaction with glyceraldehyde and Nα-acetyllysine, 3-hydroxy-5-hydroxymethyl-pyridinium were previously reported as exhibiting fluorescence at 298 nm (excitation) and 388 nm (emission) [38]. Given that the fluorescence wavelengths of collagen amino-carbonylated with glyceraldehyde were 360 nm (excitation) and 460 nm (emission), it is possible that other fluorescent molecules are generated. We found that the fluorescence intensity of the BM increased upon treatment with glyceraldehyde, indicating that a fluorescent cross-linked material was formed with collagen fibrils.
Aminoguanidine, OPB-9195 (2-isopropylidenehydrazono-4-oxo-thiazolidine-5-ylacetanilide), and LR-90 (methylene bis [4,4-(2 chlorophenylureido phenoxyisobutyric acid)]) are AGEs inhibitors, although they are not used in clinical settings [39,40]. In the present study, we found that methylparaben—a widely used ingredient in cosmetics—abrogated the decrease in ATP and filaggrin mRNA levels caused by the culture of keratinocytes on amino-carbonylated collagen. A similar effect was observed with spermidine at concentrations of 1.7 and 3.4 mmol/L (data not shown). In addition, methylparaben suppressed filaggrin mRNA levels in HaCaT cultured on amino-carbonylated collagen gel at a concentration 0.1 mmol/L, with the effect of methylparaben being 35 times more potent than that of spermidine.
5. Conclusions
The results of this study indicate that amino carbonylation of the epidermal BM decreased moisture retention, keratinization, and ceramide synthesis in the stratum corneum, thereby reducing its barrier function; these effects were reversed by methylparaben. Thus, cosmetics containing methylparaben can potentially improve epidermal function that has been compromised by amino carbonylation of epidermal BM.
Acknowledgments
The authors gratefully acknowledge the technical assistance of Ms. Miha Kurihara.
Author Contributions
Haruka Morimoto, Lihao Gu, Haifeng Zeng and Kazuhisa Maeda performed the experiments. Kazuhisa Maeda designed the study and performed the data analysis. Haruka Morimoto, Lihao Gu, Haifeng Zeng and Kazuhisa Maeda interpreted the data and drafted the manuscript. Kazuhisa Maeda supervised the study and critically revised the manuscript. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vistoli, G.; De Maddis, D.; Cipak, A.; Zarkovic, N.; Carini, M.; Aldini, G. Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): An overview of their mechanisms of formation. Free Radic. Res. 2013, 47, 3–27. [Google Scholar] [CrossRef] [PubMed]
- Haus, J.M.; Carrithers, J.A.; Trappe, S.W.; Trappe, T.A. Collagen, cross-linking, and advanced glycation end products in aging human skeletal muscle. J. Appl. Physiol. 2007, 103, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Yamagishi, S.; Inagaki, Y.; Amano, S.; Koga, K.; Abe, R.; Takeuchi, M.; Ohno, S.; Yoshimura, A.; Makita, Z. Angiogenesis induced by advanced glycation end products and its prevention by cerivastatin. FASEB J. 2002, 16, 1928–1930. [Google Scholar] [CrossRef] [PubMed]
- Abe, R.; Shimizu, T.; Sugawara, H.; Watanabe, H.; Nakamura, H.; Choei, H.; Sasaki, N.; Yamagishi, S.; Takeuchi, M.; Shimizu, H. Regulation of human melanoma growth and metastasis by AGE-AGE receptor interactions. J. Investig. Dermatol. 2004, 122, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Verzijl, N.; DeGroot, J.; Thorpe, S.R.; Bank, R.A.; Shaw, J.N.; Lyons, T.J.; Bijlsma, J.W.; Lafeber, F.P.; Baynes, J.W.; TeKoppele, J.M. Effect of collagen turnover on the accumulation of advanced glycation end products. J. Biol. Chem. 2000, 275, 39027–39031. [Google Scholar] [CrossRef] [PubMed]
- Airaksinen, K.E.; Salmela, P.I.; Linnaluoto, M.K.; Ikäheimo, M.J.; Ahola, K.; Ryhänen, L.J. Diminished arterial elasticity in diabetes: Association with fluorescent advanced glycosylation end products in collagen. Cardiovasc. Res. 1993, 27, 942–945. [Google Scholar] [CrossRef] [PubMed]
- Pageon, H.; Asselineau, D. An in vitro approach to the chronological aging of skin by glycation of the collagen: The biological effect of glycation on the reconstructed skin model. Ann. N. Y. Acad. Sci. 2005, 1043, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Pageon, H.; Técher, M.P.; Asselineau, D. Reconstructed skin modified by glycation of the dermal equivalent as a model for skin aging and its potential use to evaluate anti-glycation molecules. Exp. Gerontol. 2008, 43, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Mizutari, K.; Ono, T.; Ikeda, K.; Kayashima, K.; Horiuchi, S. Photo-enhanced modification of human skin elastin in actinic elastosis by N(epsilon)-(carboxymethyl)lysine, one of the glycoxidation products of the Maillard reaction. J. Investig. Dermatol. 1997, 108, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, H.; Oyobikawa, M.; Tada, A.; Maeda, T.; Takiwaki, H.; Itoh, M.; Kanto, H. Melanin and facial skin fluorescence as markers of yellowish discoloration with aging. Skin Res. Technol. 2009, 15, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Ren, M.; Yang, C.; Hu, Y.X.; Ran, J.M.; Yan, L. Involvement of RAGE, MAPK and NF-κB pathways in AGEs-induced MMP-9 activation in HaCaT keratinocytes. Exp. Dermatol. 2012, 21, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Serban, A.I.; Stanca, L.; Geicu, O.I.; Munteanu, M.C.; Dinischiotu, A. RAGE and TGF-β1 Cross-Talk Regulate Extracellular Matrix Turnover and Cytokine Synthesis in AGEs Exposed Fibroblast Cells. PLoS ONE 2016, 11, e0152376. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Kim, J.Y.; Oh, S.H. Advanced glycation end products (AGEs) promote melanogenesis through receptor for AGEs. Sci. Rep. 2016, 6, 27848. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M.; Vlassara, H.; Kooney, A.; Ulrich, P.; Cerami, A. Aminoguanidine prevents diabetes-induced arterial wall protein cross-linking. Science 1986, 232, 1629–1632. [Google Scholar] [CrossRef] [PubMed]
- Vasan, S.; Zhang, X.; Zhang, X.; Kapurniotu, A.; Bernhagen, J.; Teichberg, S.; Basgen, J.; Wagle, D.; Shih, D.; Terlecky, I.; et al. An agent cleaving glucose-derived protein crosslinks in vitro and in vivo. Nature 1996, 382, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Litchfield, J.E.; Baynes, J.W. AGE-breakers cleave model compounds, but do not break Maillard crosslinks in skin and tail collagen from diabetic rats. Arch. Biochem. Biophys. 2003, 412, 42–46. [Google Scholar] [CrossRef]
- Rahbar, S.; Figarola, J.L. Novel inhibitors of advanced glycation endproducts. Arch. Biochem. Biophys. 2003, 419, 63–79. [Google Scholar] [CrossRef] [PubMed]
- Anisuzzaman; Hatta, T; Miyoshi, T; Matsubayashi, M; Islam, M.K; Alim, M.A; Anas, M.A; Hasan, M.M; Matsumoto, Y; Yamamoto, Y; et al. Longistatin in tick saliva blocks advanced glycation end-product receptor activation. J. Clin. Investig. 2014, 124, 4429–4444. [Google Scholar] [CrossRef] [PubMed]
- Karuppagounder, V.; Arumugam, S.; Thandavarayan, R.A.; Pitchaimani, V.; Sreedhar, R.; Afrin, R.; Harima, M.; Suzuki, H.; Nomoto, M.; Miyashita, S.; et al. Modulation of HMGB1 translocation and RAGE/NFκB cascade by quercetin treatment mitigates atopic dermatitis in NC/Nga transgenic mice. Exp. Dermatol. 2015, 24, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Han, A.R.; Nam, M.H.; Lee, K.W. Plantamajoside Inhibits UVB and advanced glycation end products-induced MMP-1 wxpression by suppressing the MAPK and NF-κB pathways in HaCaT cells. Photochem. Photobiol. 2016, 92, 708–719. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.T.; Meng, F.H.; Chen, L.; Li, X.; Cen, L.J.; Wen, Y.H.; Zhang, H.; Li, C.C. Inhibition of Methylglyoxal-Induced AGEs/RAGE Expression Contributes to Dermal Protection by N-Acetyl-l-Cysteine. Cell Physiol. Biochem. 2017, 41, 742–754. [Google Scholar] [CrossRef] [PubMed]
- Menon, G.K.; Norlén, L. Stratum corneum ceramides and their role in skin barrier function. In Skin Moisturization, 2nd ed.; Rawlings, A.V., Leyden, J.J., Eds.; CRC Press: Boca Raton, FL, USA, 2002; pp. 31–60. [Google Scholar]
- Tagami, H. Functional characteristics of the stratum corneum in photoaged skin in comparison with those found in intrinsic aging. Arch. Dermatol. Res. 2008, 300, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kezic, S.; Kemperman, P.M.; Koster, E.S.; de Jongh, C.M.; Thio, H.B.; Campbell, L.E.; Irvine, A.D.; McLean, W.H.; Puppels, G.J.; Caspers, P.J. Loss-of-function mutations in the filaggrin gene lead to reduced level of natural moisturizing factor in the stratum corneum. J. Investig. Dermatol. 2008, 128, 2117–2119. [Google Scholar] [CrossRef] [PubMed]
- Holleran, W.M.; Feingold, K.R.; Man, M.Q.; Gao, W.N.; Lee, J.M.; Elias, P.M. Regulation of epidermal sphingolipid synthesis by permeability barrier function. J. Lipid Res. 1991, 32, 1151–1158. [Google Scholar] [PubMed]
- Rogers, J.; Harding, C.; Mayo, A.; Banks, J.; Rawlings, A. Stratum corneum lipids: The effect of ageing and the seasons. Arch. Dermatol. Res. 1996, 288, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Matsuki, M.; Yamashita, F.; Ishida-Yamamoto, A.; Yamada, K.; Kinoshita, C.; Fushiki, S.; Ueda, E.; Morishima, Y.; Tabata, K.; Yasuno, H.; et al. Defective stratum corneum and early neonatal death in mice lacking the gene for transglutaminase 1 (keratinocyte transglutaminase). Proc. Natl. Acad. Sci. USA 1998, 95, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Candi, E.; Schmidt, R.; Melino, G. The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell Biol. 2005, 6, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Griffin, M.; Casadio, R.; Bergamini, C.M. Transglutaminases: Nature’s biological glues. Biochem. J. 2002, 368, 377–396. [Google Scholar] [CrossRef] [PubMed]
- Lorand, L.; Graham, R.M. Transglutaminases: Crosslinking enzymes with pleiotropic functions. Nat. Rev. Mol. Cell Biol. 2003, 4, 140–156. [Google Scholar] [CrossRef] [PubMed]
- Stachowitz, S.; Alessandrini, F.; Abeck, D.; Ring, J.; Behrendt, H. Permeability barrier disruption increases the level of serine palmitoyltransferase in human epidermis. J. Investig. Dermatol. 2002, 119, 1048–1052. [Google Scholar] [PubMed]
- Doering, T.; Brade, H.; Sandhoff, K. Sphingolipid metabolism during epidermal barrier development in mice. J. Lipid Res. 2002, 43, 1727–1733. [Google Scholar] [CrossRef] [PubMed]
- Hamanaka, S.; Hara, M.; Nishio, H.; Otsuka, F.; Suzuki, A.; Uchida, Y. Human epidermal glucosylceramides are major precursors of stratum corneum ceramides. J. Investig. Dermatol. 2002, 119, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Sell, D.R.; Carlson, E.C.; Monnier, V.M. Differential effects of type 2 (non-insulin-dependent) diabetes mellitus on pentosidine formation in skin and glomerular basement membrane. Diabetologia 1993, 36, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Haitoglou, C.S.; Tsilibary, E.C.; Brownlee, M.; Charonis, A.S. Altered cellular interactions between endothelial cells and nonenzymatically glucosylated laminin/type IV collagen. J. Biol. Chem. 1992, 267, 12404–12407. [Google Scholar] [PubMed]
- Alikhani, Z.; Alikhani, M.; Boyd, C.M.; Nagao, K.; Trackman, P.C.; Graves, D.T. Advanced glycation end products enhance expression of pro-apoptotic genes and stimulate fibroblast apoptosis through cytoplasmic and mitochondrial pathways. J. Biol. Chem. 2005, 280, 12087–12095. [Google Scholar] [CrossRef] [PubMed]
- Dyer, D.G.; Dunn, J.A.; Thorpe, S.R.; Bailie, K.E.; Lyons, T.J.; McCance, D.R.; Baynes, J.W. Accumulation of Maillard reaction products in skin collagen in diabetes and aging. J. Clin. Investig. 1993, 91, 2463–2469. [Google Scholar] [CrossRef] [PubMed]
- Usui, T.; Hayase, F. Isolation and identification of the 3-hydroxy-5-hydroxymethyl-pyridinium compound as a novel advanced glycation end product on glyceraldehyde-related Maillard reaction. Biosci. Biotechnol. Biochem. 2003, 67, 930–932. [Google Scholar] [CrossRef] [PubMed]
- Bolton, W.K.; Cattran, D.C.; Williams, M.E.; Adler, S.G.; Appel, G.B.; Cartwright, K.; Foiles, P.G.; Freedman, B.I.; Raskin, P.; Ratner, R.E.; et al. ACTION I Investigator Group. Randomized trial of an inhibitor of formation of advanced glycation end products in diabetic nephropathy. Am. J. Nephrol. 2014, 24, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Nenna, A.; Nappi, F.; Avtaar Singh, S.S.; Sutherland, F.W.; Di Domenico, F.; Chello, M.; Spadaccio, C. Pharmacologic approaches against advanced glycation end products (AGEs) in diabetic cardiovascular disease. Res. Cardiovasc. Med. 2015, 4, e26949. [Google Scholar] [CrossRef]
Figure 1.
(A,B) Effects of glyceraldehyde on the relative fluorescence intensity of epidermal BM. Relative fluorescence intensity (excitation/emission, 360/460 nm) was compared between epidermal BM treated with glyceraldehyde (A, 2 and 5 mmol/L for 4 days; B, 5 mmol/L for 2, 4, and 6 days) and untreated epidermal BM (control). (C) ATP levels in human keratinocyte cultured on epidermal BM treated with 5 mmol/L glyceraldehyde (for 2, 4, and 6 days) and untreated epidermal BM. Results are expressed as mean ± standard deviation of three experiments. * p < 0.05 vs. control, ** p < 0.01 vs. control, *** p < 0.001 vs. control. (D) Effects of amino carbonylation of the epidermal BM on TEWL in the 3D model of human keratinocyte. The 3D model was cultured on epidermal BM treated with glyceraldehyde (5 mmol/L) for 4 days. Fold changes in TEWL levels are expressed as mean ± standard deviation of three experiments. * p < 0.05 vs. epidermal BM.
Figure 1.
(A,B) Effects of glyceraldehyde on the relative fluorescence intensity of epidermal BM. Relative fluorescence intensity (excitation/emission, 360/460 nm) was compared between epidermal BM treated with glyceraldehyde (A, 2 and 5 mmol/L for 4 days; B, 5 mmol/L for 2, 4, and 6 days) and untreated epidermal BM (control). (C) ATP levels in human keratinocyte cultured on epidermal BM treated with 5 mmol/L glyceraldehyde (for 2, 4, and 6 days) and untreated epidermal BM. Results are expressed as mean ± standard deviation of three experiments. * p < 0.05 vs. control, ** p < 0.01 vs. control, *** p < 0.001 vs. control. (D) Effects of amino carbonylation of the epidermal BM on TEWL in the 3D model of human keratinocyte. The 3D model was cultured on epidermal BM treated with glyceraldehyde (5 mmol/L) for 4 days. Fold changes in TEWL levels are expressed as mean ± standard deviation of three experiments. * p < 0.05 vs. epidermal BM.
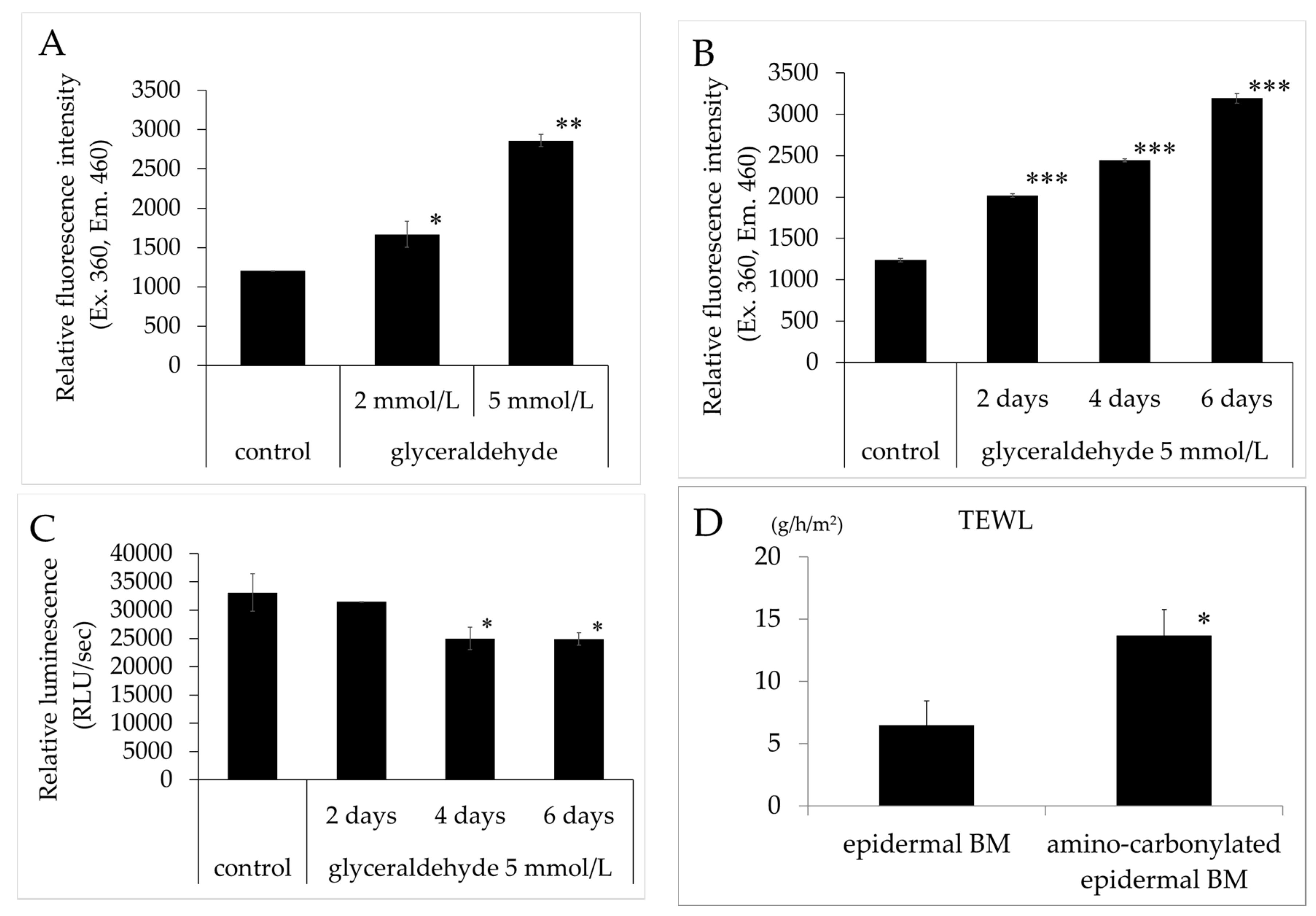
Figure 2.
Effects of amino carbonylation of the epidermal BM on FLG, TGase1, SPTLC2, GCase, and aSMase mRNA levels in the human epidermis model. (A) FLG; (B) TGase1; (C) SPTLC2; (D) GCase; and (E) aSMase levels were determined by real-time reverse transcription PCR. The 3D model of human epidermis was cultured on epidermal BM treated with glyceraldehyde (5 mmol/L for 4 days). Expression levels of target genes relative to the internal control (β-actin) were determined with the 2−ΔΔCt method. Fold changes in gene expression levels are expressed as mean ± standard deviation of three experiments. * p < 0.05 vs. epidermal BM.
Figure 2.
Effects of amino carbonylation of the epidermal BM on FLG, TGase1, SPTLC2, GCase, and aSMase mRNA levels in the human epidermis model. (A) FLG; (B) TGase1; (C) SPTLC2; (D) GCase; and (E) aSMase levels were determined by real-time reverse transcription PCR. The 3D model of human epidermis was cultured on epidermal BM treated with glyceraldehyde (5 mmol/L for 4 days). Expression levels of target genes relative to the internal control (β-actin) were determined with the 2−ΔΔCt method. Fold changes in gene expression levels are expressed as mean ± standard deviation of three experiments. * p < 0.05 vs. epidermal BM.
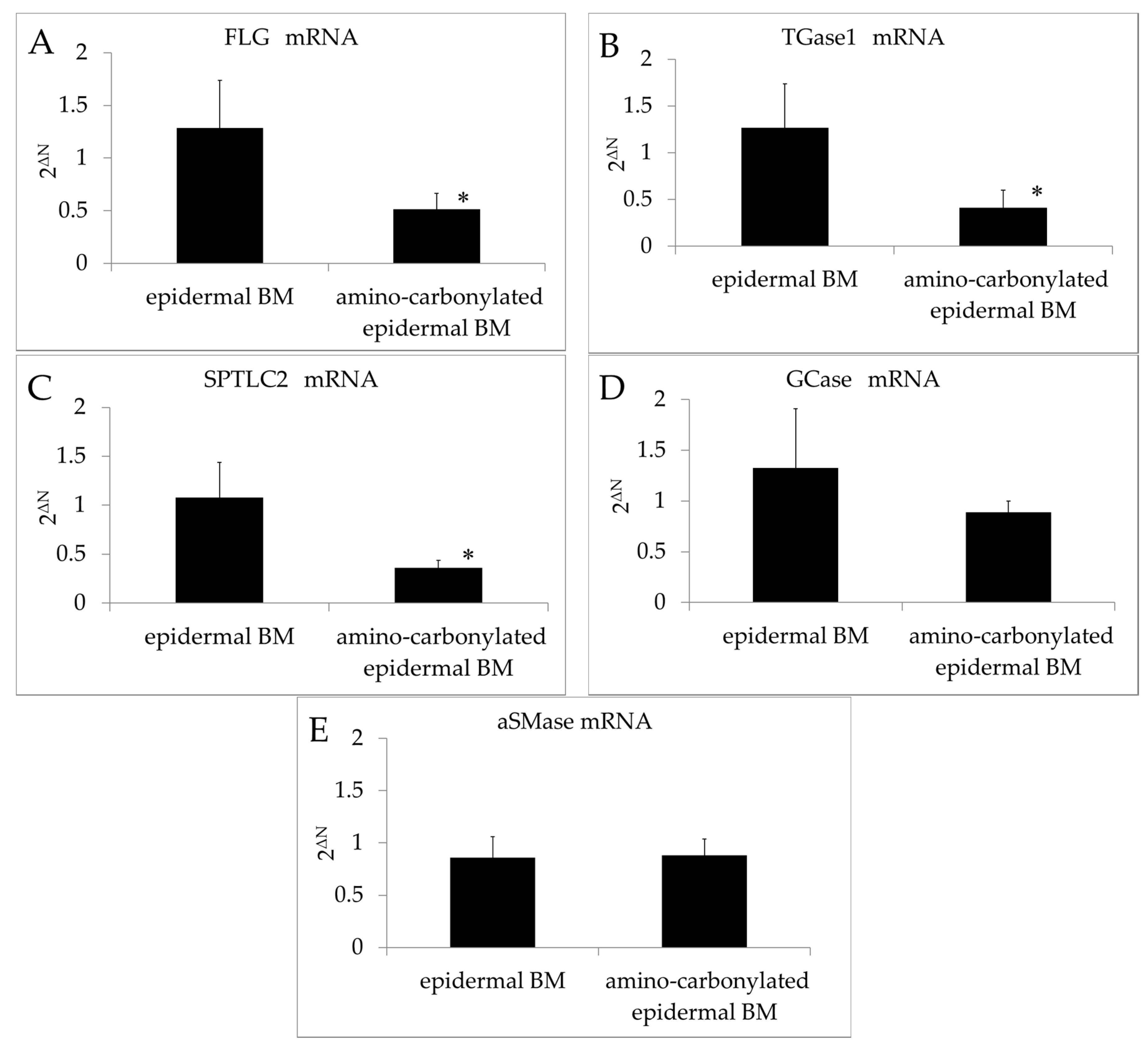
Figure 3.
Effects of amino carbonylation of the epidermal BM on FLG and TGase1 protein levels in a human epidermis model. The 3D models of human epidermis were cultured on epidermal BM (A) and BM treated with 5 mmol/L glyceraldehyde for 4 days (B). FLG (green) and TGase1 (red) localized to the cell membrane in the stratum corneum layer. Dotted lines show the boundary between the stratum corneum layer and air.
Figure 3.
Effects of amino carbonylation of the epidermal BM on FLG and TGase1 protein levels in a human epidermis model. The 3D models of human epidermis were cultured on epidermal BM (A) and BM treated with 5 mmol/L glyceraldehyde for 4 days (B). FLG (green) and TGase1 (red) localized to the cell membrane in the stratum corneum layer. Dotted lines show the boundary between the stratum corneum layer and air.
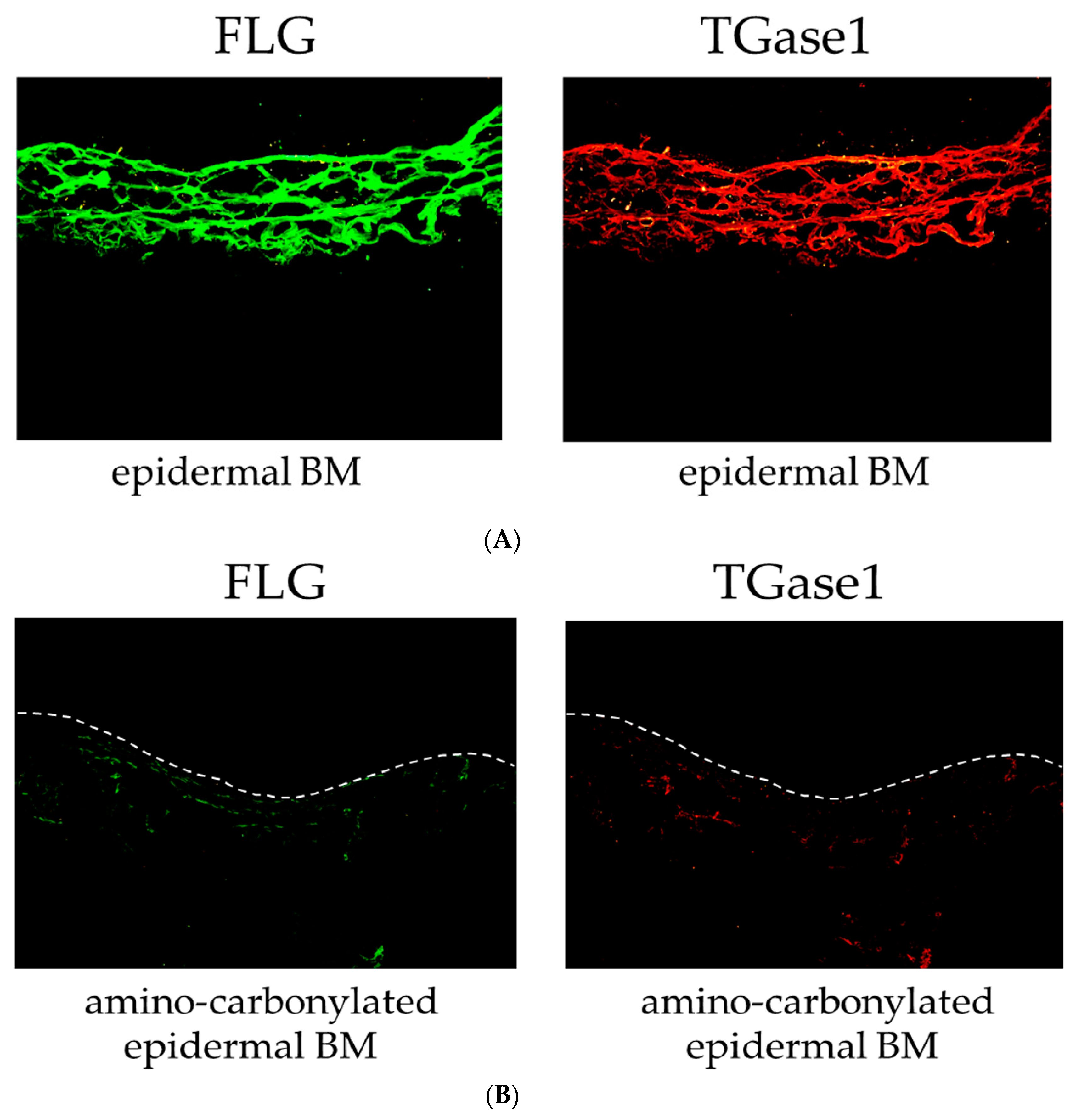
Figure 4.
Effects of glyceraldehyde on the relative fluorescence intensity of amino-carbonylated collagen gel. Relative fluorescence intensity (excitation/emission, 360/460 nm) was compared between collagen gel treated with glyceraldehyde (0.625, 1.25, 2.5, 5, 10, 25, and 50 mmol/L for 4 days) and untreated collagen gel (control). Results are expressed as mean ± standard deviation of three experiments. * p < 0.05 vs. control, *** p < 0.001 vs. control.
Figure 4.
Effects of glyceraldehyde on the relative fluorescence intensity of amino-carbonylated collagen gel. Relative fluorescence intensity (excitation/emission, 360/460 nm) was compared between collagen gel treated with glyceraldehyde (0.625, 1.25, 2.5, 5, 10, 25, and 50 mmol/L for 4 days) and untreated collagen gel (control). Results are expressed as mean ± standard deviation of three experiments. * p < 0.05 vs. control, *** p < 0.001 vs. control.
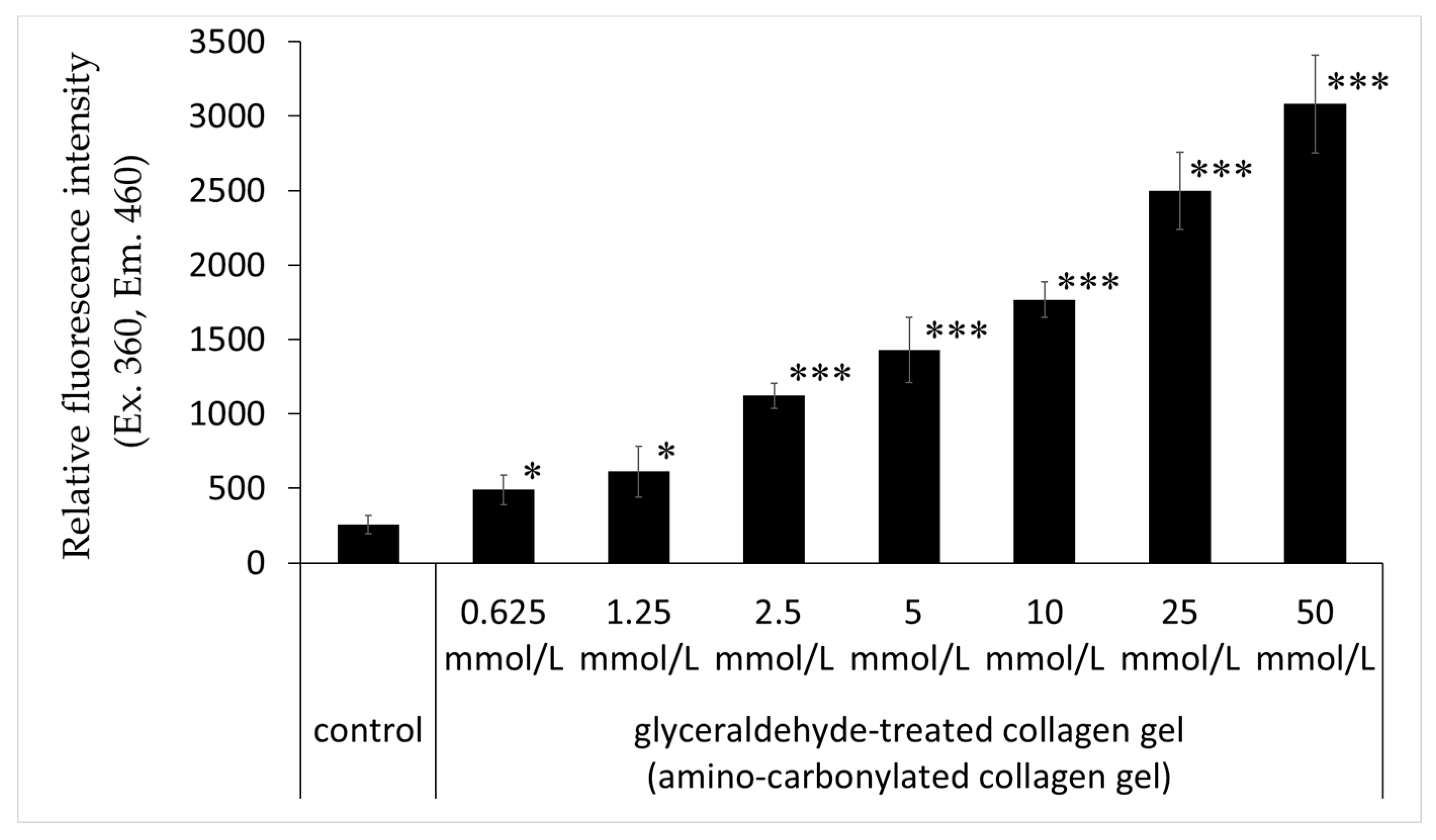
Figure 5.
Effect of methylparaben on the decreases in ATP and filaggrin mRNA levels in HaCaT cultured on amino-carbonylated collagen gel. (A) ATP levels in amino-carbonylated collagen gel in the left figure and control in right figure are the same data; (B) Filaggrin mRNA. Results are expressed as mean ± standard deviation of three experiments. ### p < 0.001 vs. untreated collagen gel; * p < 0.05, ** p < 0.01 vs. control on amino-carbonylated collagen gel.
Figure 5.
Effect of methylparaben on the decreases in ATP and filaggrin mRNA levels in HaCaT cultured on amino-carbonylated collagen gel. (A) ATP levels in amino-carbonylated collagen gel in the left figure and control in right figure are the same data; (B) Filaggrin mRNA. Results are expressed as mean ± standard deviation of three experiments. ### p < 0.001 vs. untreated collagen gel; * p < 0.05, ** p < 0.01 vs. control on amino-carbonylated collagen gel.

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Morimoto, H.; Gu, L.; Zeng, H.; Maeda, K. Amino Carbonylation of Epidermal Basement Membrane Inhibits Epidermal Cell Function and Is Suppressed by Methylparaben. Cosmetics 2017, 4, 38. https://doi.org/10.3390/cosmetics4040038
AMA Style
Morimoto H, Gu L, Zeng H, Maeda K. Amino Carbonylation of Epidermal Basement Membrane Inhibits Epidermal Cell Function and Is Suppressed by Methylparaben. Cosmetics. 2017; 4(4):38. https://doi.org/10.3390/cosmetics4040038
Chicago/Turabian StyleMorimoto, Haruka, Lihao Gu, Haifeng Zeng, and Kazuhisa Maeda. 2017. "Amino Carbonylation of Epidermal Basement Membrane Inhibits Epidermal Cell Function and Is Suppressed by Methylparaben" Cosmetics 4, no. 4: 38. https://doi.org/10.3390/cosmetics4040038
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.