The Brain–Intestinal Mucosa–Appendix– Microbiome–Brain Loop


1
Sydney Medical School, The University of Sydney, Sydney 2006, Australia
2
Medlab Clinical Ltd., Sydney 2015, Australia
*
Author to whom correspondence should be addressed.
Diseases 2018, 6(2), 23; https://doi.org/10.3390/diseases6020023
Submission received: 5 February 2018
/
Revised: 28 February 2018
/
Accepted: 29 March 2018
/
Published: 1 April 2018
(This article belongs to the Special Issue Gut Microbiome and Human Diseases)
{kind=link}
{kind=link}
Abstract
:The brain and the gut are connected from early fetal life. The mother’s exposure to microbial molecules is thought to exert in utero developmental effects on the fetus. These effects could importantly underpin the groundwork for subsequent pathophysiological mechanisms for achieving immunological tolerance and metabolic equilibrium post birth, events that continue through to 3–4 years of age. Furthermore, it is understood that the microbiome promotes cues that instruct the neonate’s mucosal tissues and skin in the language of molecular and cellular biology. Post birth mucosal lymphoid tissue formation and maturation (most probably including the vermiform appendix) is microbiota-encouraged co-establishing the intestinal microbiome with a developing immune system. Intestinal mucosal tissue maturation loops the brain-gut-brain and is postulated to influence mood dispositions via shifts in the intestinal microbiome phyla. A plausible appreciation is that dysregulated pro-inflammatory signals from intestinal resident macrophages could breach the loop by providing adverse mood signals via vagus nerve afferents to the brain. In this commentary, we further suggest that the intestinal resident macrophages act as an upstream traffic controller of translocated microbes and metabolites in order to maintain local neuro-endocrine-immunological equilibrium. When macrophages are overwhelmed through intestinal microbiome and intestinal epithelial cell dysbiosis, pro-inflammatory signals are sustained, which may then lead to mood disorders. The administration of probiotics as an adjunctive medicine co-administered with antidepressant medications in improving depressed mood may have biological and clinical standing.
1. Commentary
1.1. Macrophages and Intestinal Tissue
Both the intestines and the brain develop from the same cluster of embryonic tissue that can be traced back to the primitive streak in early vertebrate fetal growth. As such, this grooved structure that forms on day 15 of human development, along the caudal midline of the bilaminar embryonic disc is the first visible sign of gastrulation that will give rise to the ectoderm, mesoderm and endoderm [1]. When the tissue divides, a portion develops into the Central Nervous System (CNS) (i.e., brain and spinal cord) and the other into the Enteric Nervous System (ENS). Immune molecules are intimately related to the development of the CNS as they are for the intestinal tract. The intestinal tract is a complex multi-dimensional structure that is derived from a simple tubular structure [2]. Such is the complexity that the established intestinal epithelia are embraced by the crypt-villus elements that contain absorptive enterocytes and secretory tuft, goblet, Paneth, entero-endocrine cell types; while also accommodating resident intestinal stem cells and rapidly dividing progenitor cells. Adjacent to the intestinal epithelium is an additional complex structure comprising the lamina propria, submucosa, and muscular layers.
Immune molecules have been reported to participate in integral functions in the CNS throughout various stages of neural development, including affecting neurogenesis, neuronal migration, axon guidance, synapse formation, activity-dependent refinement of circuits, and synaptic plasticity [3]. For example, the innate-like, evolutionarily conserved MR1-restricted mucosa-associated invariant T cells (MAIT) from humans represent the most abundant T-cell subset that quickly respond to a wide variety of bacteria [4]. MAIT cells in humans are prevalent and are distributed throughout the blood and mucosal sites [5]. Studies of MAIT cells in human fetal development are important for the consideration of the fundamental immune-biological characteristics expressed, as well as for the role these cells play in the fetus and the newborn. In humans T cells are enriched in mucosal tissues [6].
In the intestines there is recognized a tripartite co-operation in order to maintain intestinal steady state homeostasis. This occurs between the intestinal epithelium barrier, the intestinal microbiome and gut mucosal immune cells such as macrophages [2,7,8].
Macrophages (mononuclear phagocytes) are distributed throughout the body in tissue sites that include for example the intestines, the lung, the liver, and the brain [9]. Macrophages are established from prenatal signals and from circulating monocytes post birth [9]. The appreciation that macrophages have multiple actions in order to maintain the overall immunological efficiency in the gut is an idea that reflects macrophage functional diversity in specific tissues, where the tissue site provides instructive signals for local macrophage differentiation [10].
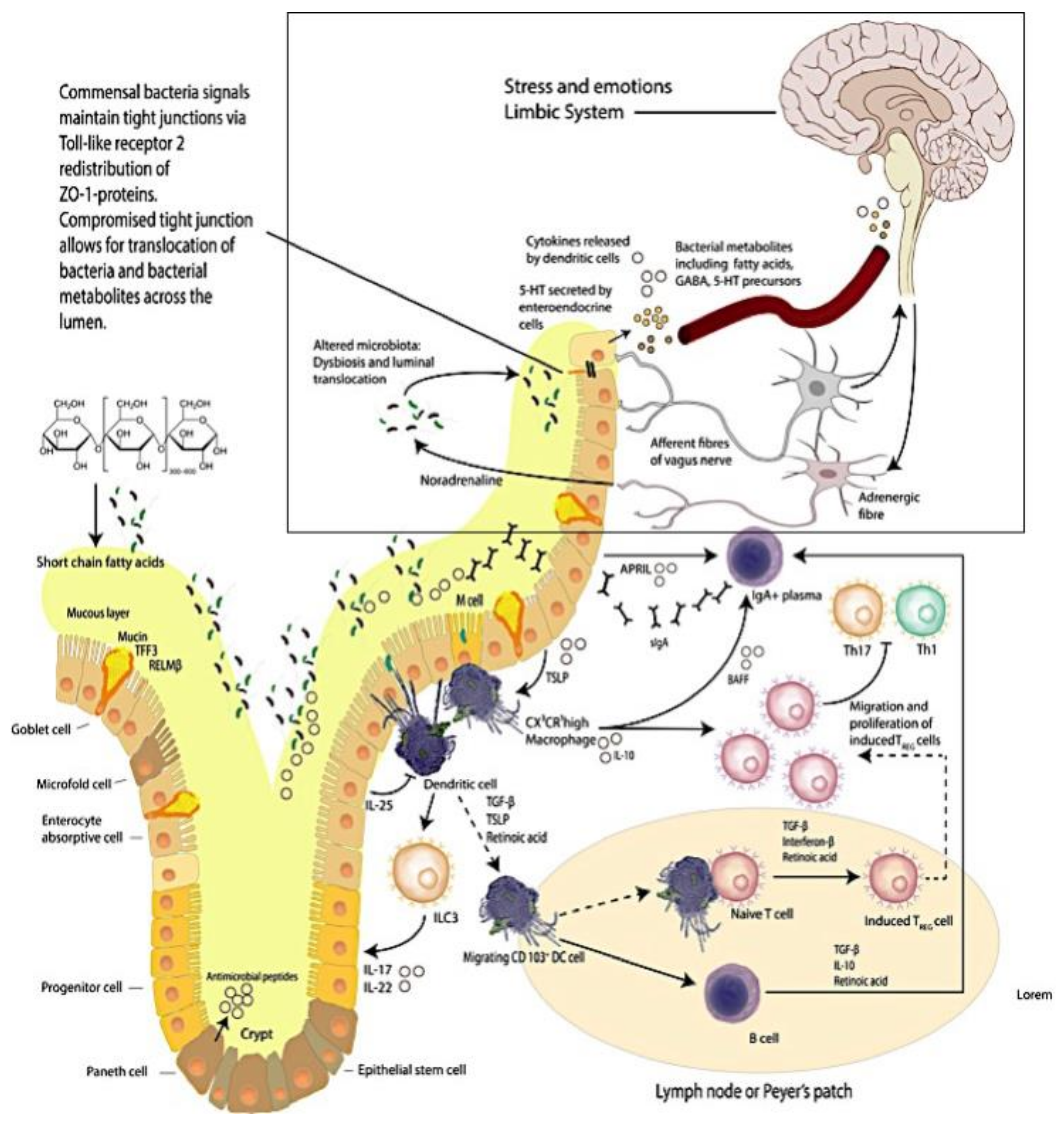
In the gut, intestinal resident macrophages (CX3CR1hi) are specialized cells that are involved in antigen presentation to T cells that in turn shape the T cell responses generated [11]. As such intestinal resident macrophages are important participants in contributing and maintaining the steady state equilibrium of mucosal immunity. Macrophages shape mucosal immune equilibrium through the action of phagocytosis eliciting protection from pathobiont translocations across the intestinal epithelium [12] (Figure 1).
The activated macrophage phenotype is known to present antigens to T lymphocytes, which initiate the controlled and appropriate immune response that is elicited by a recognition signal that responds to microbial proteins [13,14,15] (Figure 1). Studies report that antigen presentation triggers macrophage activities that activate T cells; the activity of macrophages is linked to up regulation of a sequence of cytokines that includes interleukin 1 (IL-1), interferon-alpha (IFN-a) and other cytotoxic proteins [13]. have an important role in maintaining immunological equilibrium [15].
Maintaining immunological equilibrium also involves the phagocytosis of exogenous antigens, cellular debris, insoluble particles and activated clotting factors [15]. It is also noted that intestinal resident macrophage populations sustain mucosal tolerance by contributing to the survival and expanding T lymphocytes already primed toward immunological defense of pathogens [8]. Further, the colitogenic T lymphocyte inflammatory response that occurs in the intestinal mucosa is suppressed by the anti-inflammatory cytokine IL-10 elaborated by intestinal resident macrophages; an activity promoting intestinal mucosa immunological tolerance. The macrophage can be hence envisaged to promote regulatory T cell activities [8].
The intestinal epithelia produce soluble protein factors (e.g., thymic stromal lymphopoietin, transforming growth factor-b, and retinoic acid) and also express the integrin ligand semaphorin 7A that undergoes contact-dependent interactions with intestinal macrophages. This activity induces the expression of IL-10 that in turn further promotes intestinal homeostasis. Therefore these cumulative actions combine components of the local innate immune system (i.e., macrophages and dendritic cells and others) and the intestinal epithelia in an interaction that preserves a tolerogenic functional steady state [13].
A recent review by Roman and colleagues [16] has progressed the view that macrophages can influence inflammatory disease outcomes and that sustained inflammatory responses can lead to depressive moods. Furthermore it was postulated that antidepressive medications can influence peripheral and brain macrophages skewing them toward anti-inflammatory activities that then can improve cognitive functions [16].
1.2. The Brain–Intestinal Microbiome/Epithelia–Mucosa–Brain Loop
There are multiple mechanisms in the gut including a mucus layer, antimicrobial peptides and a tight junction protein network that cooperate to continuously preserve local homeostasis and hence ensure that the intestinal epithelial barrier integrity is maintained. Goblet cells secrete mucin to provide a protective coating, provide structural integrity and regulate macrophage and adaptive T cell responses during inflammation [8,17,18,19,20,21,22,23,24,25,26,27,28,29] (Figure 1).
Mental health is very much intricately linked to physical health [31]. Gastrointestinal dysbiosis is associated with an intestinal epithelial cell barrier dysfunction that can be due to environmental and nutritional triggers, which can be further progressed by the intestinal pathobiont cohort that exacerbates and maintains intestinal barrier in a dysfunctional pro-inflammatory state. It has been recently reported that depression is linked to the exacerbation in gap junction integrity between intestinal epithelial cells (i.e., also termed a leaky gut) is a contentious posit. A recent study has reported that approximately 35 % of depressed individuals exhibited evidence of a leaky gut [32,33].
The importance exhibited by the brain–intestine–brain axis is that it provides a bidirectional flow of neuroendocrine-immunological equilibrium control. The intestinal microbiome is thought to exert effects on this axis that significantly impacts the biochemistry of the central and peripheral nervous systems and in turn behavior [34] (Figure 1 (see within the rectangular area)). Studies reporting that depression is accompanied by activation of immune–inflammatory pathways [35] with increased IgM/IgA responses to lipopolysaccharides (LPS) from gram-negative commensal bacteria, indicate that at least in part adverse mood is supported by commensal microbes and or metabolite translocations across the intestinal epithelial barrier [32,33]. We have reported [36] (as have others [37]) that resistant depression can be accompanied by systemic inflammatory states that are posited to originate from intestinal inflammation and the resultant intestinal dysbiosis.
1.3. The Vermiform Appendix
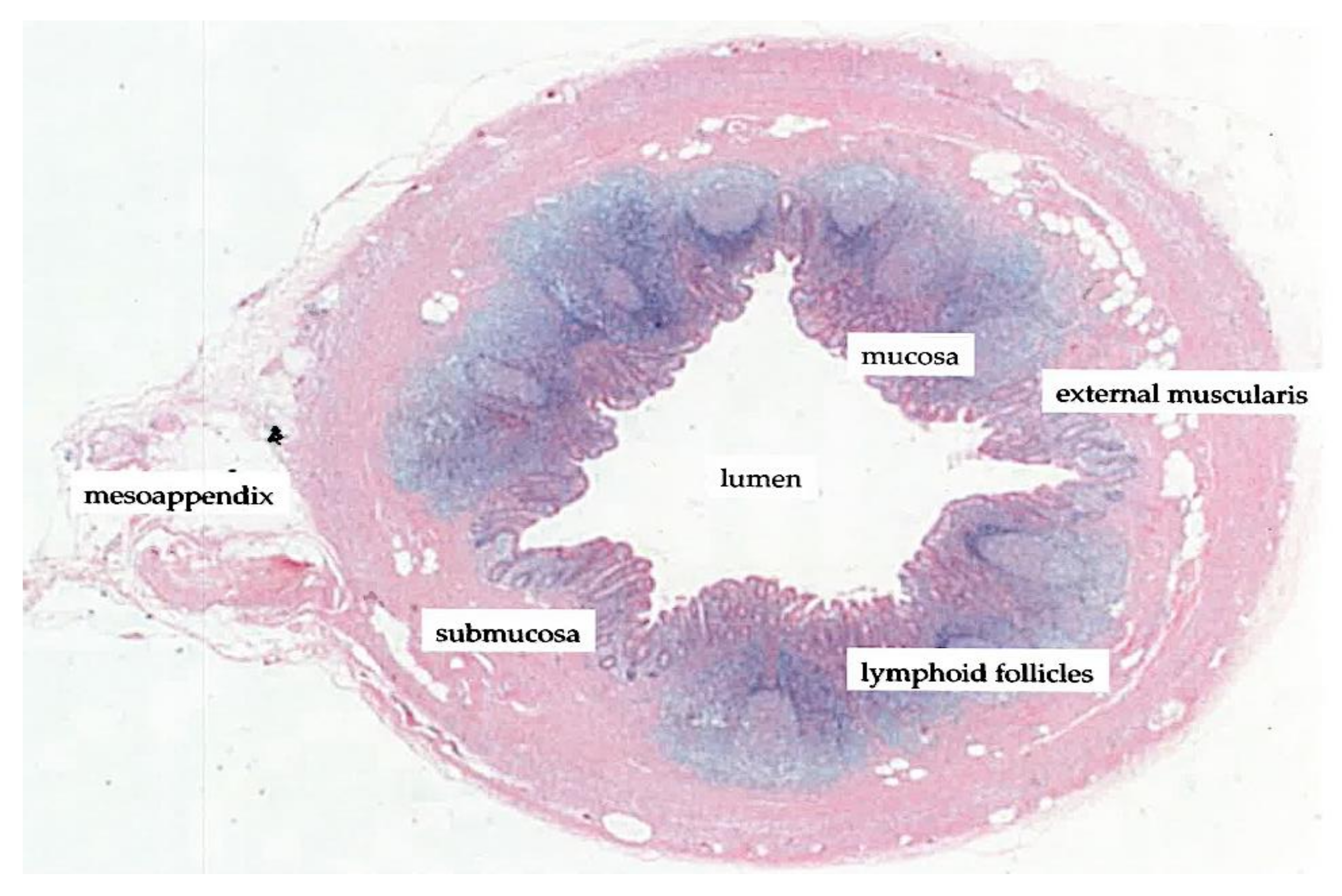
The vermiform appendix is characterized as a diverticulum of the cecum and delineates the beginning of the colon in the confluence of tanias [38]. The appendix is posterior-medially attached to the cecum, approximately 2 cm below the ileocecal junction. The histological structure of the appendix reflects that of the intestinal wall of the large bowel, in particular with appendiceal structures such as the mucosa, submucosa and lymphoid follicles (Figure 2).
It has only been recently recognized that the human vermiform appendix is not just a rudimentary part of the intestine, but rather as suggested by numerous studies an organ of immunological importance for the development and preservation of the intestinal immune system [39,40]. Furthermore the importance of the vermiform appendix has been demonstrated to have a direct functional interaction with the intestinal microbiome [41,42].
Reports present the vermiform appendix as an important participant in intestinal ecological microbial contributions and maintenance [41]. This is especially relevant to the ascending colon and the distal small bowel where much of the microbiome is posited to have high metabolic activities. Reports have documented key activities that through co-evolution the human host and the microbial cohort have shaped a tolerant relationship of benefits. These beneficial activities include (i) the intestinal based immune system sustains microbial biofilms in the intestines and is a key constituent of the mutualistic connection between mammals and bacteria [44,45] (ii) biofilms defined as mucilage layers in the gut have been reported to be safe bacterial zones [46], where bacteria and other entities (e.g., fungi) form communal relationships for safeguarding their activities and survival; (iii) the vermiform appendix is very much recognized as an immune tissue [47], with concentrated gut-associated lymphoid tissue; (iv) a recent reported concentrated biofilms contribute to the overall epithelial structure of the vermiform appendix [41,48]; (v) research also shows that biofilms in the intestines are subject to continual turnover an activity that helps to limit bacterial translocations across the intestinal epithelial cells and Peyer’s patches [48], where this mucin turnover activity is rapid (approximately 1–2 h) of any biofilms that adhere to the intestinal epithelia [49].
Early reports through culture dependent studies have demonstrated the presence of several Gram-negative bacilli such as Klebsiella, Enterobacter and Escherichia coli whereas Gram-positive cocci were less frequently observed [50]. In a more recent study with patients following an appendectomy showed a comprehensive view of the microbial population within the biofilm of the appendix, this can be determined by high-throughput DNA sequencing [51]. The human appendix was demonstrated to contain members of some fifteen bacterial phyla [51], including Firmicutes (the most dominant), Proteobacteria, Bacteroidetes, Actinobacteria, and Fusobacteria. Moreover, certain oral pathogens not associated with the intestines were also detected in the appendix samples (i.e., Gemella, Parvimonas, and Fusobacterium). This report presents an immunological organ with a significant associated microbial diversity which in part supports the posit that the appendix microbiota has important functions in human health; the biofilm in the appendix acts as a safe house for commensal bacteria, therefore, facilitating re-inoculation of the intestines post a gastrointestinal tract infection as the appendiceal lumen is spared from diarrhoeal clearance [40].
Health interventions that may disrupt the microbiome ecology of the large bowel may lead to disease. A recent study has highlighted that the vermiform appendix may have a significant association with chronic diseases in particular in the development of large bowel cancer following the surgical removal of the appendix [52]. Recently a study of patients clinically treated with antibiotics for appendicitis were reported to have a significant increased risk for large bowel cancer [53]. Moreover, although there is a paucity of studies on the relationship of appendicectomy to mood disorders, an early study has postulated that an appendicectomy may lead to psychological disturbances [54]. An interesting corollary from another study with subjects undergoing an appendicectomy showed that 80% of the participants with inflammatory problems of the appendix were designated as positive for depressive symptoms [55]. Moreover psychological depressive symptoms have been reported to continue post an appendicectomy [56]. These studies overall tend to support the notion that severe disruptions of the intestinal microbiome and the loss of the appendix biofilm may increase the risk of disease including adverse mood dispositions.
1.4. Probiotics
Current research continues to draw connections between the intestinal microbiome and environmental factors to sensitivities to the host’s emotional states [57]. In animal models, dysbiosis has been demonstrated to impair vagus signaling which results in reduced protein synthesis in the hippocampus, corrected by rescuing the intestinal microbiome with either specific strains of probiotics [58,59]. Our group has recently demonstrated in a small pilot study [60] with treatment resistant depression (while on SSRI medications) that the administration of a probiotic formulation improved depressive symptoms in a small cohort. An anti-inflammatory response was suggested. A meta-analysis by Ng and colleagues [61] investigating the administration of probiotics to alleviate depressive symptoms was largely inconclusive with an insignificant effect on mood.
An interesting recent study [62] in a healthy murine model demonstrated that oral administration of probiotic bacteria cell walls stimulated the immune system. This study although preliminary, showed that probiotic bacteria and their cell walls have important immunoregulatory effects on intestinal epithelial cells without an adverse effect on the local metabolic and environment equilibrium. The study by Lemme-Dumit and colleagues [62] showed that cell walls from probiotic bacteria increased immunoglobulin A secreting cells in the intestines and in innate immune cells as well as other tissues such as those of the spleen and peritoneum [62]. This study also demonstrated the capacity that probiotic bacteria provide to stimulate important cellular and immune elements such as the intestinal epithelia and intestinal resident macrophages in the gut [62].
2. Reprise
We have previously postulated that depressed mood [36] is linked to antagonistic immuno-endocrine control of intestinal homeostasis. The scientific rationale posited suggests that inflammation of the intestines relevant to depressed mood is an effect associated with the intestinal mucosa. Intestinal microbiome adverse shifts that maintain low-level pro-inflammatory activity and intestinal dysbiosis that overwhelm intestinal resident macrophages is posited to play a role in depressed mood. Clinical studies show that the vermiform appendix re-inoculates the proximal colon following excessive pro-inflammatory triggers (e.g., gastrointestinal inflammations, dysentery, antibiotic administration). Reports of a surgically removed chronically inflamed vermiform appendix have been linked to chronic disease developed including depression.
We postulate that intestinal inflammations may provoke an increased risk for adverse mood disorders in those without a vermiform appendix. In depression the loss of keystone intestinal bacterial species and the incapacity to restore microbial diversity and stability in the proximal colon (i.e., due to an appendectomy) could be an important factor that disrupts the steady state of neuro-endocrine-immunological equilibrium in the intestinal mucosa, especially following the over prescription of antibiotics. We advance the idea that an in situ normal functioning vermiform appendix continually contributes to diversity and stability to the intestinal bacterial cohort over a lifetime. Probiotics that can regulate the functioning of the immune system in the gut may have important plausible implications in improving depressive mood states; dedicated studies are warranted.
Acknowledgments
We thank Mr Michael Thomsen for his assistance with producing Figure 1. Luis Vitetta has received National Institute of Complementary Medicine and National Health and Medical Research Council of Australia competitive funding and Industry support for research into bio-compounds and probiotics.
Author Contributions
L.V. conception and design of commentary/limited review. L.V., G.V., S.H. read, amended and approved the final version of the manuscript.
Conflicts of Interest
L.V. and S.H. participate in research on probiotics in Medlab Clinical’s research laboratory facility in Sydney, Australia. The authors declare no other conflict of interest.
References
- Williams, M.L.; Solnica-Krezel, L. Regulation of gastrulation movements by emergent cell and tissue interactions. Curr. Opin. Cell Biol. 2017, 48, 33–39. [Google Scholar] [CrossRef] [PubMed]
- De Santa Barbara, P.; van den Brink, G.R.; Roberts, D.J. Development and differentiation of the intestinal epithelium. Cell. Mol. Life Sci. CMLS 2003, 60, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Garay, P.A.; McAllister, A.K. Novel roles for immune molecules in neural development: Implications for neurodevelopmental disorders. Front. Synaptic Neurosci. 2010, 2, 136. [Google Scholar] [CrossRef] [PubMed]
- Lantz, O.; Legoux, F. Mait cells: An historical and evolutionary perspective. Immunol. Cell Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Napier, R.J.; Adams, E.J.; Gold, M.C.; Lewinsohn, D.M. The role of mucosal associated invariant t cells in antimicrobial immunity. Front. Immunol. 2015, 6, 344. [Google Scholar] [CrossRef] [PubMed]
- Wong, E.B.; Ndung’u, T.; Kasprowicz, V.O. The role of mucosal-associated invariant t cells in infectious diseases. Immunology 2017, 150, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Caricilli, A.M.; Castoldi, A.; Camara, N.O. Intestinal barrier: A gentlemen’s agreement between microbiota and immunity. World J. Gastrointest. Pathophysiol. 2014, 5, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Epelman, S.; Lavine, K.J.; Randolph, G.J. Origin and functions of tissue macrophages. Immunity 2014, 41, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Gautier, E.L.; Shay, T.; Miller, J.; Greter, M.; Jakubzick, C.; Ivanov, S.; Helft, J.; Chow, A.; Elpek, K.G.; Gordonov, S.; et al. Gene-expression profiles and transcriptional regulatory pathways that underlie the identity and diversity of mouse tissue macrophages. Nat. Immunol. 2012, 13, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Huang, C.; Lin, Z.; Zhan, S.; Kong, L.; Fang, C.; Li, J. Macrophage polarization and function with emphasis on the evolving roles of coordinated regulation of cellular signaling pathways. Cell. Signal. 2014, 26, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Grainger, J.R.; Konkel, J.E.; Zangerle-Murray, T.; Shaw, T.N. Macrophages in gastrointestinal homeostasis and inflammation. Pflug. Arch. Eur. J. Physiol. 2017, 469, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Sansonetti, P. Host-pathogen interactions: The seduction of molecular cross talk. Gut 2002, 50 (Suppl. 3), iii2–iii8. [Google Scholar] [CrossRef] [PubMed]
- Weber, B.; Saurer, L.; Mueller, C. Intestinal macrophages: Differentiation and involvement in intestinal immunopathologies. Semin. Immunopathol. 2009, 31, 171–184. [Google Scholar] [CrossRef] [PubMed]
- De Schepper, S.; Stakenborg, N.; Matteoli, G.; Verheijden, S.; Boeckxstaens, G.E. Muscularis macrophages: Key players in intestinal homeostasis and disease. Cell. Immunol. 2017, in press. [Google Scholar] [CrossRef] [PubMed]
- Roman, A.; Kreiner, G.; Nalepa, I. Macrophages and depression—A misalliance or well-arranged marriage? Pharmacol. Rep. 2013, 65, 1663–1672. [Google Scholar] [CrossRef]
- Robinson, K.; Deng, Z.; Hou, Y.; Zhang, G. Regulation of the intestinal barrier function by host defense peptides. Front. Vet. Sci. 2015, 2, 57. [Google Scholar] [CrossRef] [PubMed]
- Ai, T.L.; Solomon, B.D.; Hsieh, C.S. T-cell selection and intestinal homeostasis. Immunol. Rev. 2014, 259, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Ginhoux, F.; Jakubzick, C.; Naik, S.H.; Onai, N.; Schraml, B.U.; Segura, E.; Tussiwand, R.; Yona, S. Dendritic cells, monocytes and macrophages: A unified nomenclature based on ontogeny. Nat. Rev. Immunol. 2014, 14, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, C.; Kurts, C. The debate about dendritic cells and macrophages in the kidney. Front. Immunol. 2015, 6, 435. [Google Scholar] [CrossRef] [PubMed]
- Hume, D.A. Macrophages as apc and the dendritic cell myth. J. Immunol. 2008, 181, 5829–5835. [Google Scholar] [CrossRef] [PubMed]
- Kayama, H.; Ueda, Y.; Sawa, Y.; Jeon, S.G.; Ma, J.S.; Okumura, R.; Kubo, A.; Ishii, M.; Okazaki, T.; Murakami, M.; et al. Intestinal CX3C chemokine receptor 1(high) (CX3CR1(high)) myeloid cells prevent t-cell-dependent colitis. Proc. Natl. Acad. Sci. USA 2012, 109, 5010–5015. [Google Scholar] [CrossRef] [PubMed]
- Kidd, P. Th1/Th2 balance: The hypothesis, its limitations, and implications for health and disease. Altern. Med. Rev. 2003, 8, 223–246. [Google Scholar] [PubMed]
- Zheng, S.G. Regulatory T cells vs Th17: Differentiation of Th17 versus treg, are the mutually exclusive? Am. J. Clin. Exp. Immunol. 2013, 2, 94–106. [Google Scholar] [PubMed]
- Wahl, S.M. Transforming growth factor-beta: Innately bipolar. Curr. Opin. Immunol. 2007, 19, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Nakahashi-Oda, C.; Udayanga, K.G.; Nakamura, Y.; Nakazawa, Y.; Totsuka, N.; Miki, H.; Iino, S.; Tahara-Hanaoka, S.; Honda, S.; Shibuya, K.; et al. Apoptotic epithelial cells control the abundance of treg cells at barrier surfaces. Nat. Immunol. 2016, 17, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Ogino, H.; Nakamura, K.; Ihara, E.; Akiho, H.; Takayanagi, R. CD4+CD25+ regulatory T cells suppress Th17-responses in an experimental colitis model. Dig. Dis. Sci. 2011, 56, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.K.; Benoist, C.; Bluestone, J.A.; Campbell, D.J.; Ghosh, S.; Hori, S.; Jiang, S.; Kuchroo, V.K.; Mathis, D.; Roncarolo, M.G.; et al. Regulatory T cells: Recommendations to simplify the nomenclature. Nat. Immunol. 2013, 14, 307–308. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.M.; Surette, M.; Bercik, P. The interplay between the intestinal microbiota and the brain. Nat. Rev. Microbiol. 2012, 10, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Essential exercise: Physical and mental health “inextricably” linked. Major disease prevention is shown to be an additional benefit. DukeMed. Healthnews 2010, 16, 4–5.
- Maes, M.; Kubera, M.; Leunis, J.C.; Berk, M.; Geffard, M.; Bosmans, E. In depression, bacterial translocation may drive inflammatory responses, oxidative and nitrosative stress (O&NS), and autoimmune responses directed against O&NS-damaged neoepitopes. Acta Psychiatr. Scand. 2013, 127, 344–354. [Google Scholar] [PubMed]
- Slyepchenko, A.; Maes, M.; Jacka, F.N.; Kohler, C.A.; Barichello, T.; McIntyre, R.S.; Berk, M.; Grande, I.; Foster, J.A.; Vieta, E.; et al. Gut microbiota, bacterial translocation, and interactions with diet: Pathophysiological links between major depressive disorder and non-communicable medical comorbidities. Psychother. Psychosom. 2017, 86, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Cryan, J.F. The microbiome-gut-brain axis in health and disease. Gastroenterol. Cl. N. Am. 2017, 46, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.; Reiche, E.M.V.; Murru, A.; Carvalho, A.F.; Maes, M.; Berk, M.; Puri, B.K. Multiple immune-inflammatory and oxidative and nitrosative stress pathways explain the frequent presence of depression in multiple sclerosis. Mol. Neurobiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Vitetta, L.; Bambling, M.; Alford, H. The gastrointestinal tract microbiome, probiotics, and mood. Inflammopharmacology 2014, 22, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Clapp, M.; Aurora, N.; Herrera, L.; Bhatia, M.; Wilen, E.; Wakefield, S. Gut microbiota's effect on mental health: The gut-brain axis. Clin. Pract. 2017, 7, 987. [Google Scholar] [CrossRef] [PubMed]
- Wakeley, C.P. The position of the vermiform appendix as ascertained by an analysis of 10,000 cases. J. Anat. 1933, 67, 277–283. [Google Scholar] [PubMed]
- Gebbers, J.O.; Laissue, J.A. Bacterial translocation in the normal human appendix parallels the development of the local immune system. Ann. N. Y. Acad. Sci. 2004, 1029, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Im, G.Y.; Modayil, R.J.; Lin, C.T.; Geier, S.J.; Katz, D.S.; Feuerman, M.; Grendell, J.H. The appendix may protect against clostridium difficile recurrence. Clin. Gastroenterol. Hepatol. 2011, 9, 1072–1077. [Google Scholar] [CrossRef] [PubMed]
- Randal Bollinger, R.; Barbas, A.S.; Bush, E.L.; Lin, S.S.; Parker, W. Biofilms in the large bowel suggest an apparent function of the human vermiform appendix. J. Theor. Biol. 2007, 249, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Van Wey, A.S.; Cookson, A.L.; Roy, N.C.; McNabb, W.C.; Soboleva, T.K.; Shorten, P.R. Bacterial biofilms associated with food particles in the human large bowel. Mol. Nutr. Food Res. 2011, 55, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Kooij, I.A.; Sahami, S.; Meijer, S.L.; Buskens, C.J.; Te Velde, A.A. The immunology of the vermiform appendix: A review of the literature. Clin. Exp. Immunol. 2016, 186, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, J.L.; Angenent, L.T.; Gordon, J.I. Getting a grip on things: How do communities of bacterial symbionts become established in our intestine? Nat. Immunol. 2004, 5, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Bollinger, R.R.; Everett, M.L.; Wahl, S.D.; Lee, Y.H.; Orndorff, P.E.; Parker, W. Secretory iga and mucin-mediated biofilm formation by environmental strains of escherichia coli: Role of type 1 pili. Mol. Immunol. 2006, 43, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nature reviews. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Bazar, K.A.; Lee, P.Y.; Joon Yun, A. An “eye” in the gut: The appendix as a sentinel sensory organ of the immune intelligence network. Med. Hypotheses 2004, 63, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Pelaseyed, T.; Bergstrom, J.H.; Gustafsson, J.K.; Ermund, A.; Birchenough, G.M.; Schutte, A.; van der Post, S.; Svensson, F.; Rodriguez-Pineiro, A.M.; Nystrom, E.E.; et al. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol. Rev. 2014, 260, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E. Fast renewal of the distal colonic mucus layers by the surface goblet cells as measured by in vivo labeling of mucin glycoproteins. PLoS ONE 2012, 7, e41009. [Google Scholar] [CrossRef] [PubMed]
- Leigh, D.A.; Simmons, K.; Norman, E. Bacterial flora of the appendix fossa in appendicitis and postoperative wound infection. J. Clin. Pathol. 1974, 27, 997–1000. [Google Scholar] [CrossRef] [PubMed]
- Guinane, C.M.; Tadrous, A.; Fouhy, F.; Ryan, C.A.; Dempsey, E.M.; Murphy, B.; Andrews, E.; Cotter, P.D.; Stanton, C.; Ross, R.P. Microbial composition of human appendices from patients following appendectomy. MBio 2013, 4, e00366-12. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.C.; Chen, W.T.; Muo, C.H.; Ke, T.W.; Fang, C.W.; Sung, F.C. Association between appendectomy and subsequent colorectal cancer development: An asian population study. PLoS ONE 2015, 10, e0118411. [Google Scholar] [CrossRef] [PubMed]
- Enblad, M.; Birgisson, H.; Ekbom, A.; Sandin, F.; Graf, W. Increased incidence of bowel cancer after non-surgical treatment of appendicitis. Eur. J. Surg. Oncol. 2017, 43, 2067–2075. [Google Scholar] [CrossRef] [PubMed]
- Canton, G.; Santonastaso, P.; Fraccon, I.G. Life events, abnormal illness behavior, and appendectomy. Gen. Hosp. Psychiatry 1984, 6, 191–195. [Google Scholar] [CrossRef]
- Beaurepaire, J.E.; Jones, M.; Eckstein, R.P.; Smith, R.C.; Piper, D.W.; Tennant, C. The acute appendicitis syndrome: Psychological aspects of the inflamed and non-inflamed appendix. J. Psychosom. Res. 1992, 36, 425–437. [Google Scholar] [CrossRef]
- Creed, F. Life events and appendicectomy. Lancet 1981, 1, 1381–1385. [Google Scholar] [CrossRef]
- Fond, G.; Boukouaci, W.; Chevalier, G.; Regnault, A.; Eberl, G.; Hamdani, N.; Dickerson, F.; Macgregor, A.; Boyer, L.; Dargel, A.; et al. The “psychomicrobiotic”: Targeting microbiota in major psychiatric disorders: A systematic review. Pathol. Biol. 2015, 63, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Cryan, J.F. Melancholic microbes: A link between gut microbiota and depression? Neurogastroenterol. Motil. 2013, 25, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Stanton, C.; Cryan, J.F. Psychobiotics: A novel class of psychotropic. Biol. Psychiatry 2013, 74, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Bambling, M.; Edwards, S.C.; Hall, S.; Vitetta, L. A combination of probiotics and magnesium orotate attenuate depression in a small ssri resistant cohort: An intestinal anti-inflammatory response is suggested. Inflammopharmacology 2017, 25, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Ng, Q.X.; Peters, C.; Ho, C.Y.X.; Lim, D.Y.; Yeo, W.S. A meta-analysis of the use of probiotics to alleviate depressive symptoms. J. Affect. Disord. 2018, 228, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Lemme-Dumit, J.M.; Polti, M.A.; Perdigon, G.; Galdeano, C.M. Probiotic bacteria cell walls stimulate the activity of the intestinal epithelial cells and macrophage functionality. Benefic. Microbes 2018, 9, 153–164. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The Brain–Intestinal Microbiome–Intestinal Epithelia–Neuroendocrine-immune–Vagus Nerve–Brain Loop. Mucus production and immune activities delineate the complexity of the intestinal mucosa site. (This figure was adapted from selected reviews [8,30]).
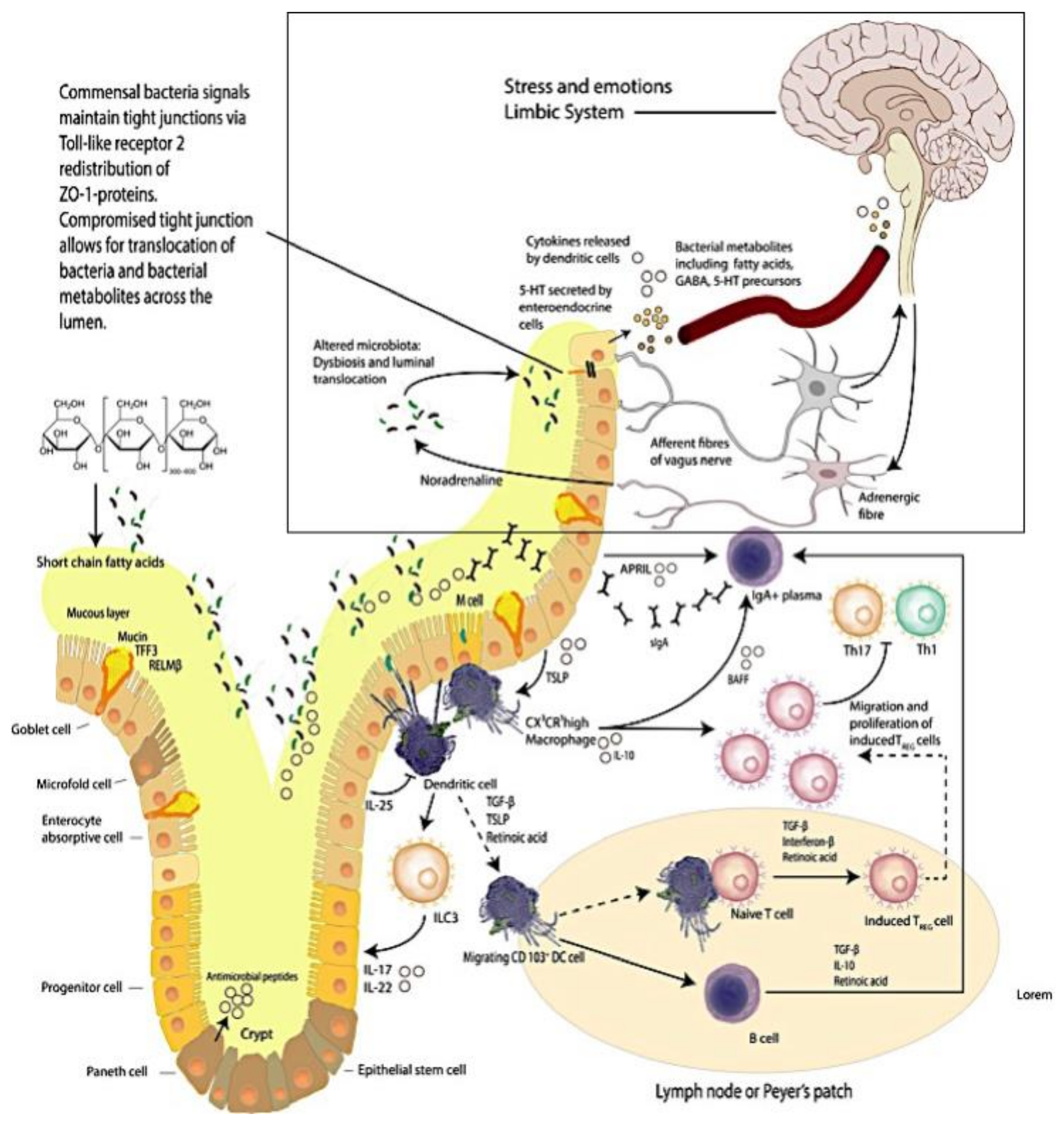
Figure 2.
Histological transverse section of the vermiform appendix (adapted/modified from Kooij et al., 2016 [43]).
Figure 2.
Histological transverse section of the vermiform appendix (adapted/modified from Kooij et al., 2016 [43]).

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vitetta, L.; Vitetta, G.; Hall, S. The Brain–Intestinal Mucosa–Appendix– Microbiome–Brain Loop. Diseases 2018, 6, 23. https://doi.org/10.3390/diseases6020023
AMA Style
Vitetta L, Vitetta G, Hall S. The Brain–Intestinal Mucosa–Appendix– Microbiome–Brain Loop. Diseases. 2018; 6(2):23. https://doi.org/10.3390/diseases6020023
Chicago/Turabian StyleVitetta, Luis, Gemma Vitetta, and Sean Hall. 2018. "The Brain–Intestinal Mucosa–Appendix– Microbiome–Brain Loop" Diseases 6, no. 2: 23. https://doi.org/10.3390/diseases6020023
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.