Exercise-Induced Alterations in Skeletal Muscle, Heart, Liver, and Serum Metabolome Identified by Non-Targeted Metabolomics Analysis

 , ,
, ,
Abstract
:1. Introduction
2. Results
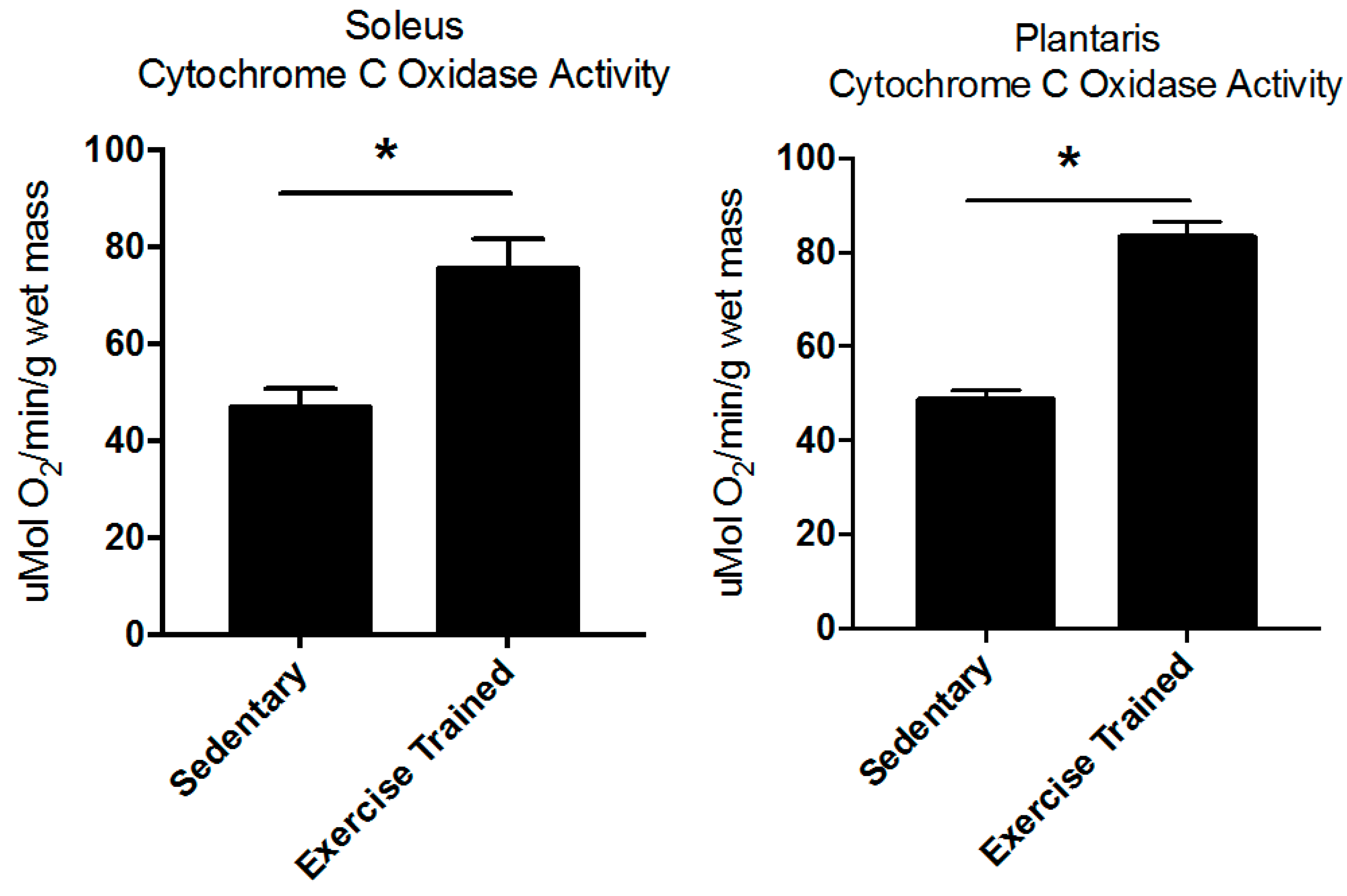
2.1. Body and Muscle Weights
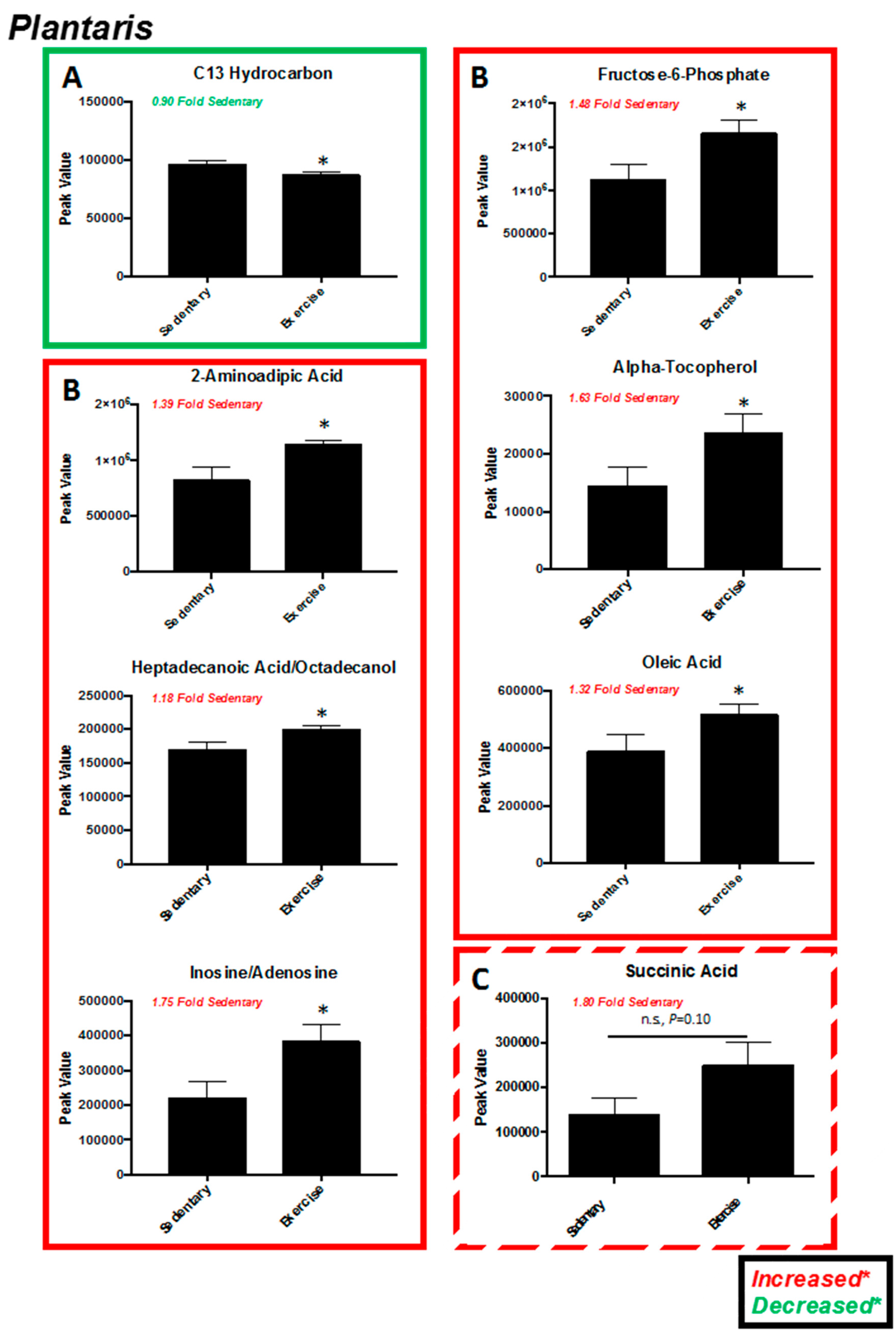
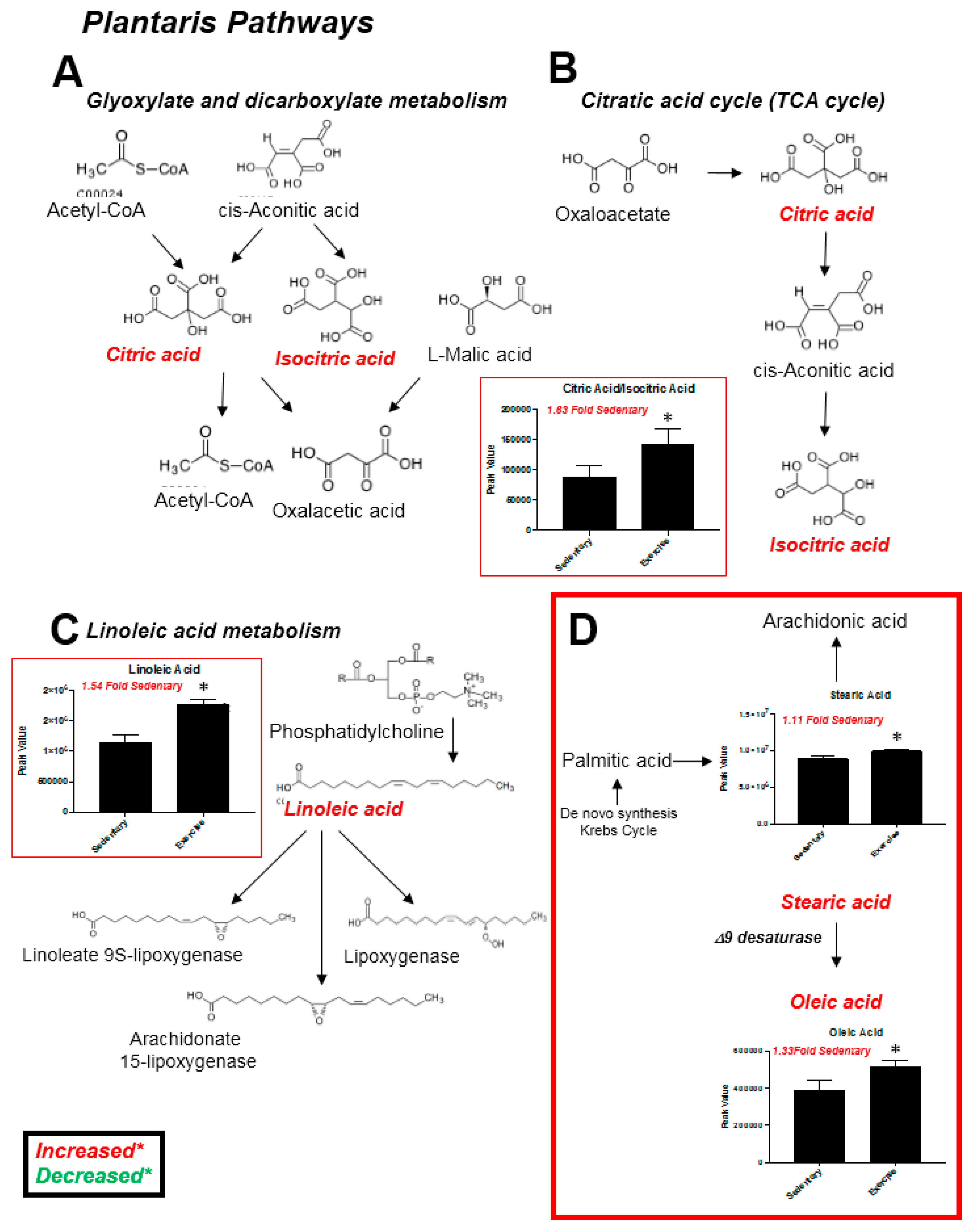
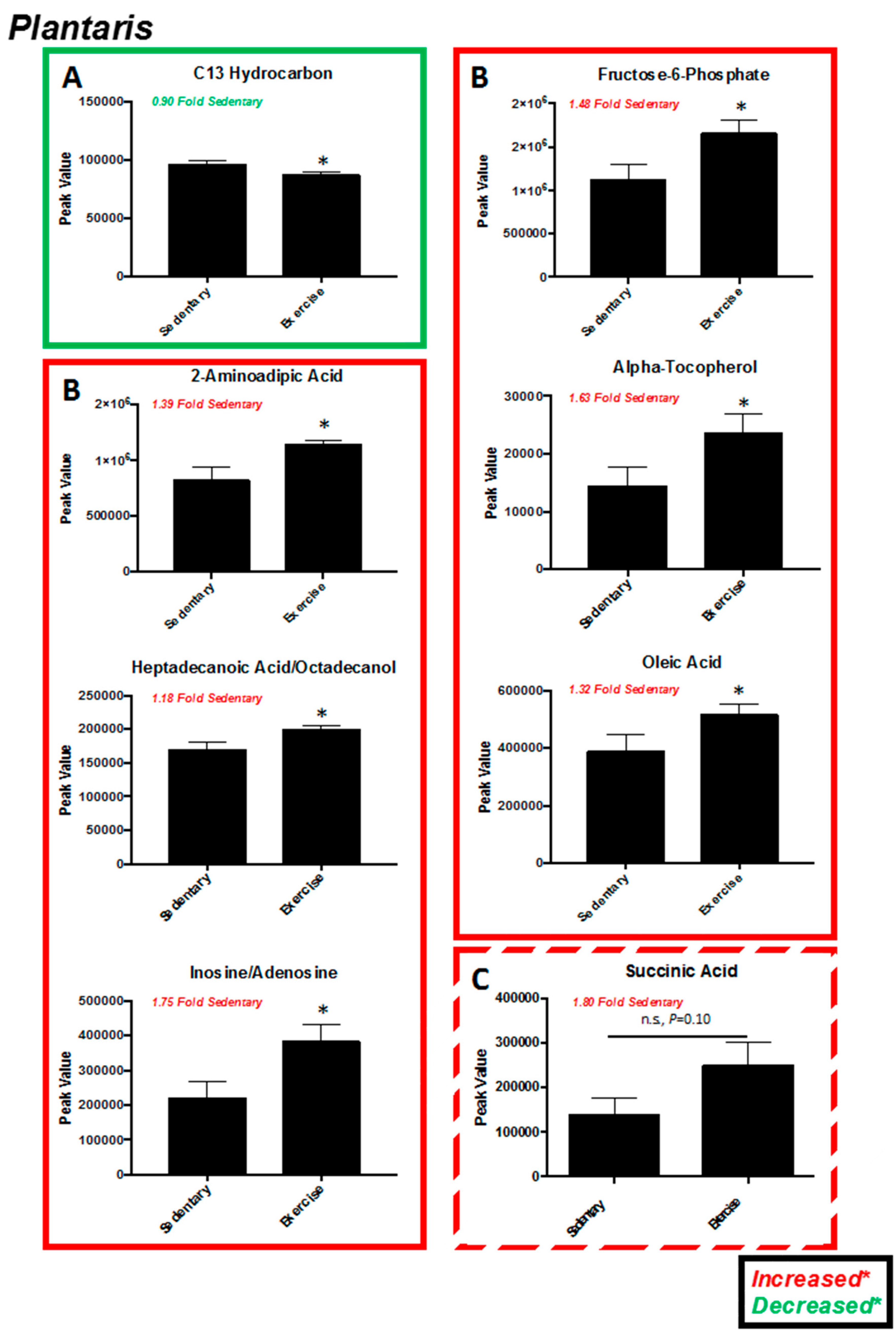
2.2. Plantaris Muscle
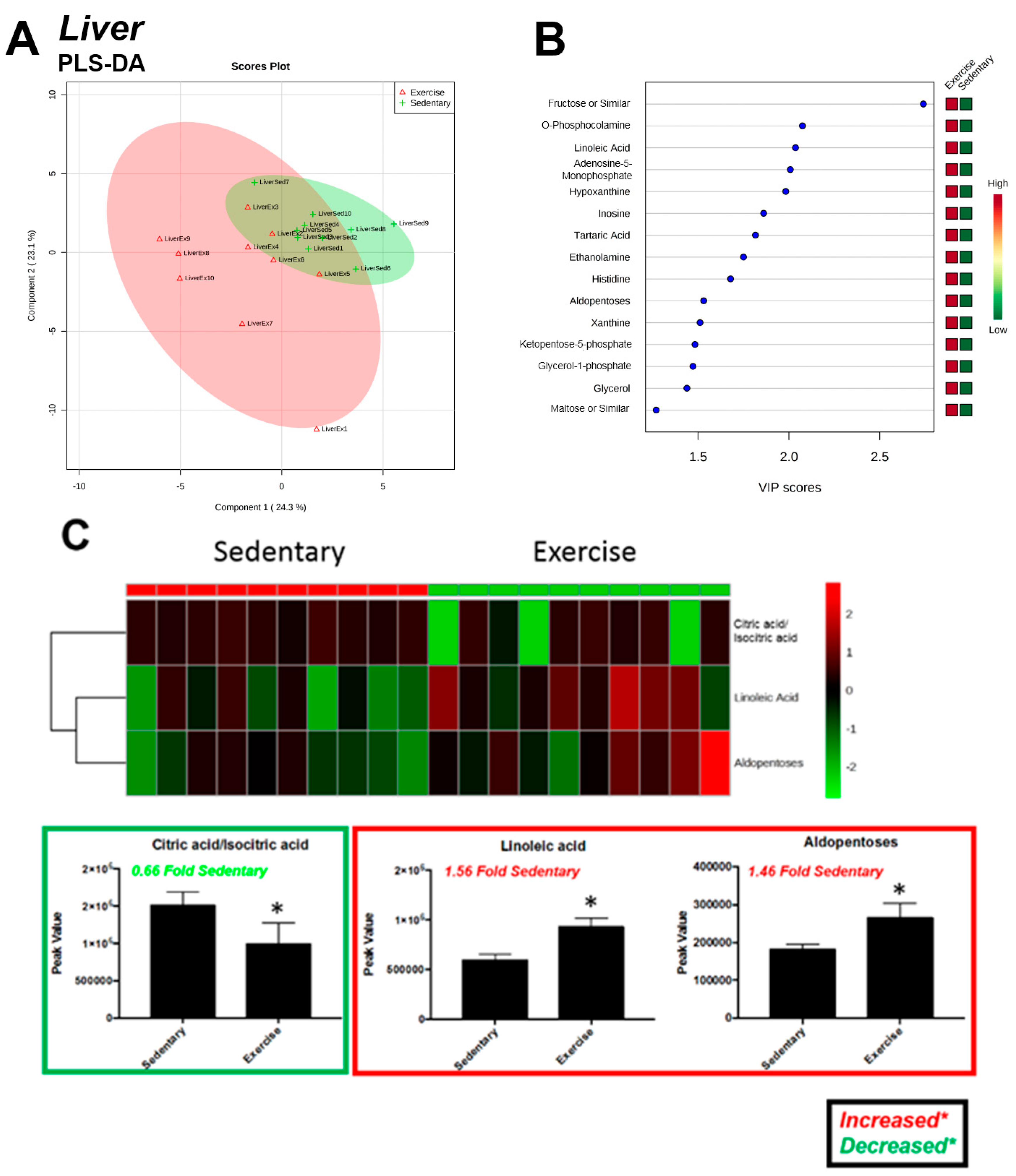
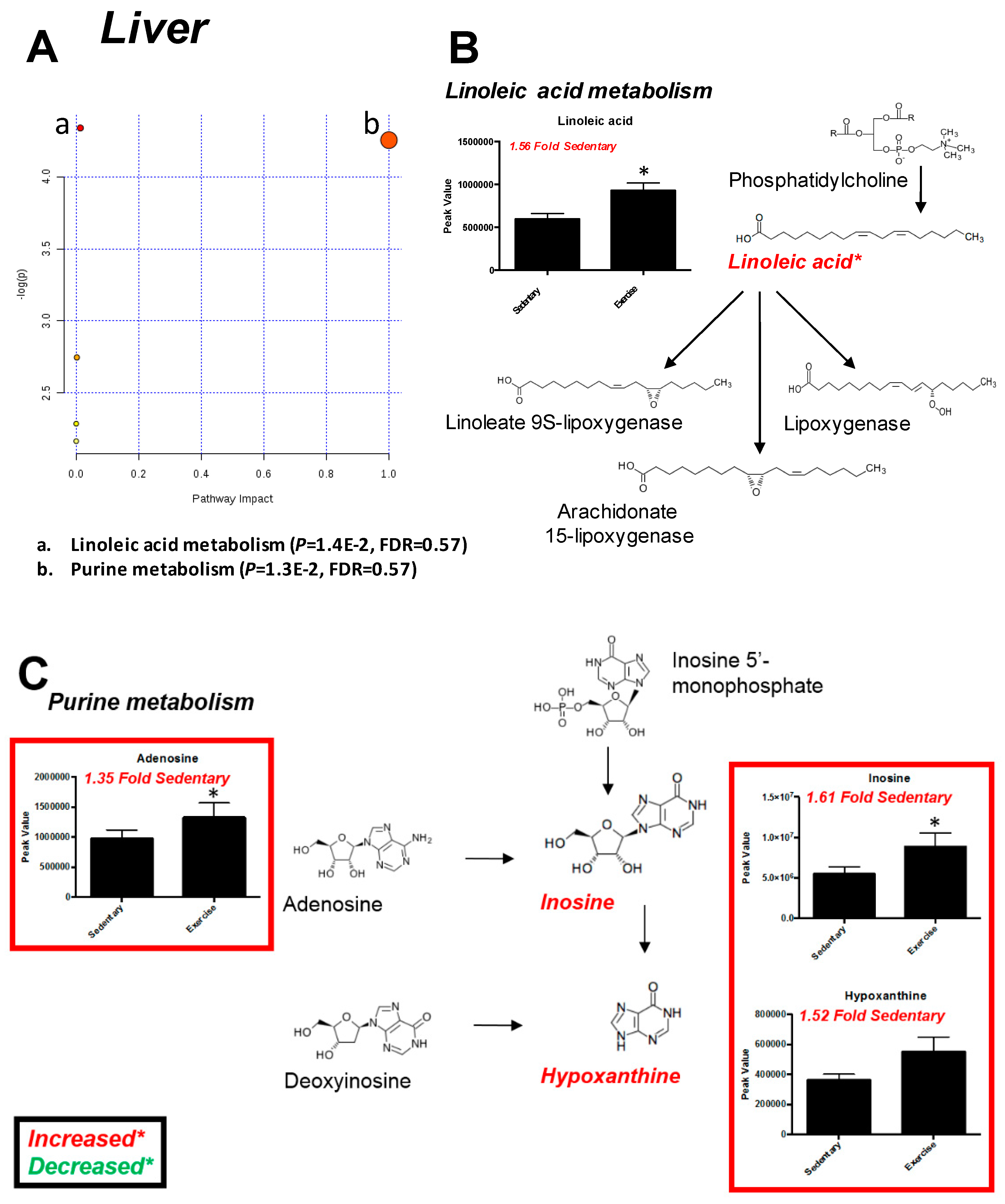
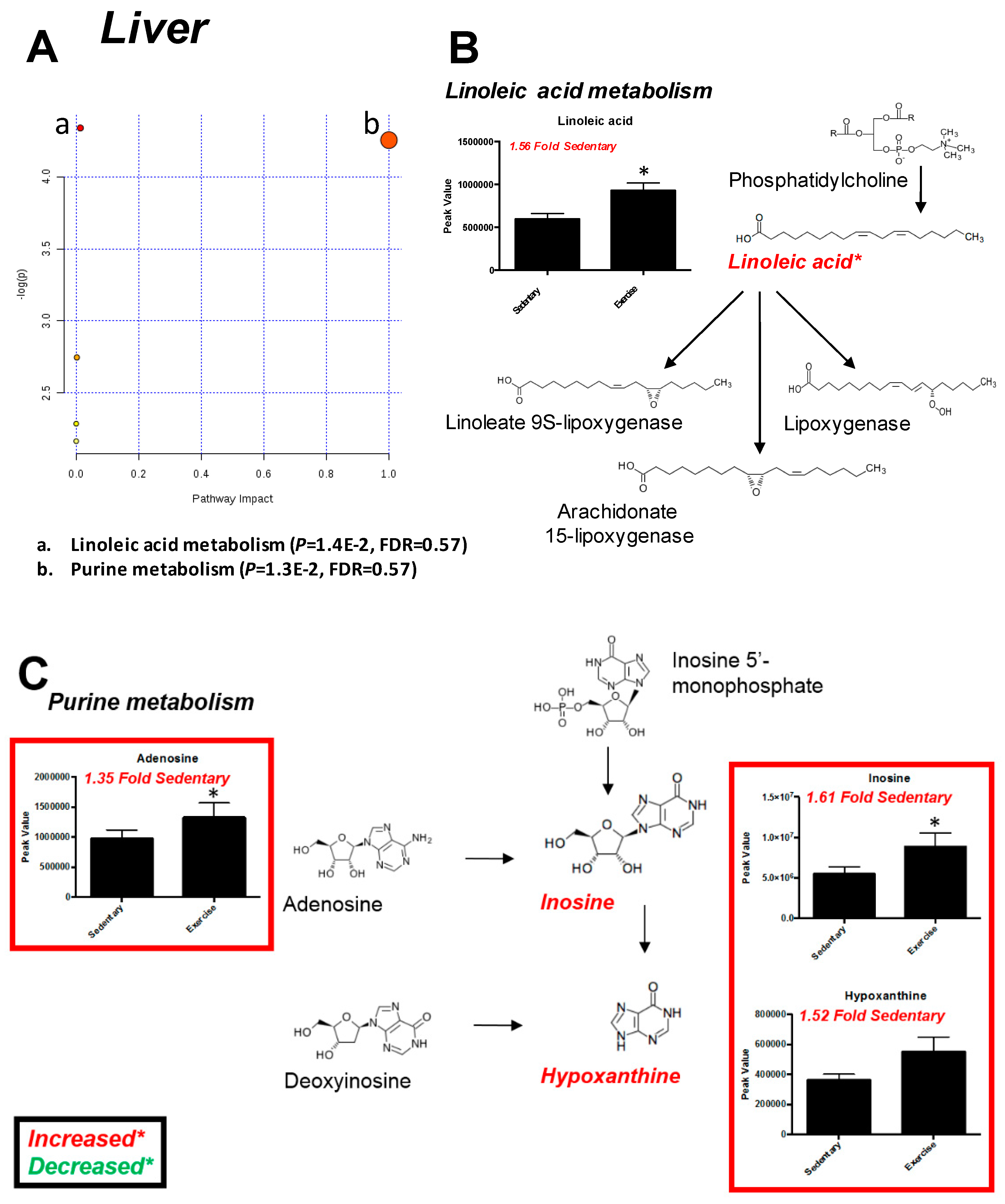
2.3. Liver
2.4. Soleus and Serum
2.5. Heart
2.6. Exercise-Induced Changes across Plantaris, Liver, Soleus, Serum, and Heart
3. Discussion
4. Materials and Methods
4.1. Animals and Exercise Treatment
4.2. Tissue and Serum Collection
4.3. Training Status
4.4. Non-Targeted Metabolomics Determination by GC–MS Instrumentation
4.5. Metabolomic Statistical Analyses
4.6. Other Statistical Analyses
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yang, X.; Li, Y.; Ren, X.; Xiong, X.; Wu, L.; Li, J.; Wang, J.; Gao, Y.; Shang, H.; Xing, Y. Effects of exercise-based cardiac rehabilitation in patients after percutaneous coronary intervention: A meta-analysis of randomized controlled trials. Sci. Rep. 2017, 7, 44789. [Google Scholar] [CrossRef] [PubMed]
- Gordon, B.; Chen, S.; Durstine, J.L. The effects of exercise training on the traditional lipid profile and beyond. Curr. Sports Med. Rep. 2014, 13, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Chorell, E.; Moritz, T.; Branth, S.; Antti, H.; Svensson, M.B. Predictive metabolomics evaluation of nutrition-modulated metabolic stress responses in human blood serum during the early recovery phase of strenuous physical exercise. J. Proteom. Res. 2009, 8, 2966–2977. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.M.; Hardman, A.E. Exercise and postprandial lipid metabolism: An update on potential mechanisms and interactions with high-carbohydrate diets. J. Nutr. Biochem. 2003, 14, 122–132. [Google Scholar] [CrossRef]
- Gill, J.M.; Cooper, A.R. Physical activity and prevention of type 2 diabetes mellitus. Sports Med. 2008, 38, 807–824. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Cabrera, M.C.; Domenech, E.; Vina, J. Moderate exercise is an antioxidant: Upregulation of antioxidant genes by training. Free Radic. Biol. Med. 2008, 44, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.D.; Franklin, B.A.; Balady, G.J.; Blair, S.N.; Corrado, D.; Estes, N.A.; Fulton, J.E.; Gordon, N.F.; Haskell, W.L.; Link, M.S.; et al. Exercise and acute cardiovascular events placing the risks into perspective: A scientific statement from the american heart association council on nutrition, physical activity, and metabolism and the council on clinical cardiology. Circulation 2007, 115, 2358–2368. [Google Scholar] [PubMed]
- Lew, J.K.; Pearson, J.T.; Schwenke, D.O.; Katare, R. Exercise mediated protection of diabetic heart through modulation of microrna mediated molecular pathways. Cardiovasc. Diabetol. 2017, 16, 10. [Google Scholar] [CrossRef] [PubMed]
- Boccatonda, A.; Tripaldi, R.; Davi, G.; Santilli, F. Oxidative stress modulation through habitual physical activity. Curr. Pharm. Des. 2016, 22, 3648–3680. [Google Scholar] [CrossRef] [PubMed]
- Idorn, M.; Thor Straten, P. Exercise and cancer: From “healthy” to “therapeutic”? Cancer Immunol. Immunother. 2017, 66, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Idorn, M.; Hojman, P. Exercise-dependent regulation of NK cells in cancer protection. Trends Mol. Med. 2016, 22, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wei, S.; Shi, Y.; Pang, S.; Qin, Q.; Yin, J.; Deng, Y.; Chen, Q.; Wei, S.; Nie, S.; et al. The dose-response effect of physical activity on cancer mortality: Findings from 71 prospective cohort studies. Br. J. Sports Med. 2016, 50, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Mielcarek, M.; Isalan, M. A shared mechanism of muscle wasting in cancer and huntington’s disease. Clin. Transl. Med. 2015, 4, 34. [Google Scholar] [CrossRef] [PubMed]
- Pescatello, L.S.; Franklin, B.A.; Fagard, R.; Farquhar, W.B.; Kelley, G.A.; Ray, C.A.; American College of Sports Medicine position stand. Progression models in resistance training for healthy adults. Med. Sci. Sports Exerc. 2004, 36, 533–553. [Google Scholar] [CrossRef] [PubMed]
- Bray, M.S.; Hagberg, J.M.; Perusse, L.; Rankinen, T.; Roth, S.M.; Wolfarth, B.; Bouchard, C. The human gene map for performance and health-related fitness phenotypes: The 2006–2007 update. Med. Sci. Sports Exerc. 2009, 41, 35–73. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; A, J.; Wang, G.; Lu, H.; Huang, X.; Liu, Y.; Zha, W.; Hao, H.; Zhang, Y.; Liu, L.; et al. Metabolomic investigation into variation of endogenous metabolites in professional athletes subject to strength-endurance training. J. Appl. Physiol. 2009, 106, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Burniston, J.G. Changes in the rat skeletal muscle proteome induced by moderate-intensity endurance exercise. Biochim. Biophys. Acta 2008, 1784, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Fazelzadeh, P.; Hangelbroek, R.W.; Tieland, M.; de Groot, L.C.; Verdijk, L.B.; van Loon, L.J.; Smilde, A.K.; Alves, R.D.; Vervoort, J.; Muller, M.; et al. The muscle metabolome differs between healthy and frail older adults. J. Proteom. Res. 2016, 15, 499–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.C.; Lin, W.T.; Hsu, F.L.; Tsai, P.W.; Hou, C.C. Metabolomics investigation of exercise-modulated changes in metabolism in rat liver after exhaustive and endurance exercises. Eur. J. Appl. Physiol. 2010, 108, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Feger, B.J.; Starnes, J.W. Exercise alters the regulation of myocardial Na+/H+ exchanger-1 activity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R1182–R1189. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.J.; Harris, M.B.; Boluyt, M.O.; Hwang, H.S.; Starnes, J.W. Effect of N-2-mercaptopropionyl glycine on exercise-induced cardiac adaptations. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R993–R1000. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.R.; Harris, M.B.; Cordaro, A.R.; Starnes, J.W. Effect of body temperature during exercise on skeletal muscle cytochrome c oxidase content. J. Appl. Physiol. 2002, 93, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Huffman, K.M.; Koves, T.R.; Hubal, M.J.; Abouassi, H.; Beri, N.; Bateman, L.A.; Stevens, R.D.; Ilkayeva, O.R.; Hoffman, E.P.; Muoio, D.M.; et al. Metabolite signatures of exercise training in human skeletal muscle relate to mitochondrial remodelling and cardiometabolic fitness. Diabetologia 2014, 57, 2282–2295. [Google Scholar] [CrossRef] [PubMed]
- Bentley, D.J.; Ackerman, J.; Clifford, T.; Slattery, K.S. Acute and chronic effects of antioxidant supplementation on exercise performance. In Antioxidants in Sport Nutrition; Lamprecht, M., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 141–154. [Google Scholar] [CrossRef]
- Reid, M.B.; Stokic, D.S.; Koch, S.M.; Khawli, F.A.; Leis, A.A. N-acetylcysteine inhibits muscle fatigue in humans. J. Clin. Investig. 1994, 94, 2468–2474. [Google Scholar] [CrossRef] [PubMed]
- Watson, T.A.; MacDonald-Wicks, L.K.; Garg, M.L. Oxidative stress and antioxidants in athletes undertaking regular exercise training. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.B. Invited review: Redox modulation of skeletal muscle contraction: What we know and what we don’t. J. Appl. Physiol. 2001, 90, 724–731. [Google Scholar] [PubMed]
- Labazi, M.; McNeil, A.K.; Kurtz, T.; Lee, T.C.; Pegg, R.B.; Angeli, J.P.; Conrad, M.; McNeil, P.L. The antioxidant requirement for plasma membrane repair in skeletal muscle. Free Radic. Biol. Med. 2015, 84, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Howard, A.C.; McNeil, A.K.; McNeil, P.L. Promotion of plasma membrane repair by vitamin E. Nat. Commun. 2011, 2, 597. [Google Scholar] [CrossRef] [PubMed]
- Dudley, G.A.; Abraham, W.M.; Terjung, R.L. Influence of exercise intensity and duration on biochemical adaptations in skeletal muscle. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1982, 53, 844–850. [Google Scholar] [PubMed]
- Bowles, D.K.; Farrar, R.P.; Starnes, J.W. Exercise training improves cardiac function after ischemia in the isolated, working rat heart. Am. J. Physiol. 1992, 263, H804–H809. [Google Scholar] [PubMed]
- Ariano, M.A.; Armstrong, R.B.; Edgerton, V.R. Hindlimb muscle fiber populations of five mammals. J. Histochem. Cytochem. 1973, 21, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Bultman, S.J.; Holley, D.; Hillhouse, C.; Bain, J.R.; Newgard, C.B.; Muehlbauer, M.J.; Willis, M.S. Non-targeted metabolomics of Brg1/Brm double-mutant cardiomyocytes reveals a novel role for swi/snf complexes in metabolic homeostasis. Metabolomics 2015, 11, 1287–1301. [Google Scholar] [CrossRef] [PubMed]
- Guler, A.T.; Waaijer, C.J.; Palmblad, M. Scientific workflows for bibliometrics. Scientometrics 2016, 107, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Rocca-Serra, P.; Salek, R.M.; Arita, M.; Correa, E.; Dayalan, S.; Gonzalez-Beltran, A.; Ebbels, T.; Goodacre, R.; Hastings, J.; Haug, K.; et al. Data standards can boost metabolomics research, and if there is a will, there is a way. Metabolomics 2016, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Sud, M.; Fahy, E.; Cotter, D.; Azam, K.; Vadivelu, I.; Burant, C.; Edison, A.; Fiehn, O.; Higashi, R.; Nair, K.S.; et al. Metabolomics workbench: An international repository for metabolomics data and metadata, metabolite standards, protocols, tutorials and training, and analysis tools. Nucleic Acids Res. 2016, 44, D463–D470. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. Metaboanalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weights | Group | |
|---|---|---|
| - | Sedentary | Exercise Trained |
| Body, (g) | 392.5 ± 13.0 | 361.8 ± 8.1 |
| Heart, (mg) | 1205 ± 25 | 1208 ± 24 |
| Soleus, (mg) | 149.9 ± 7.2 | 151.9 ± 8.8 |
| Plantaris, (mg) | 426.6 ± 15.4 | 396.9 ± 16.7 |
| Heart/Body, (mg/g) | 3.10 ± 0.09 | 3.34 ± 0.05 * |
| Soleus/Body, (mg/g) | 0.38 ± 0.01 | 0.42 ± 0.02 |
| Plantaris/Body, (mg/g) | 1.09 ± 0.05 | 1.10 ± 0.04 |
| Significantly Altered Plantaris Metabolites with Exercise (t-Test, Figure 2C) | Significantly Altered Liver Metabolites with Exercise (t-Test, Figure 5C) | Significantly Altered Soleus Metabolites with Exercise (t-Test, Figure S3C,S5A) | Significantly Altered Serum Metabolites with Exercise (t-Test, Figure S4C,S5B) | Significantly Altered Heart Metabolites with Exercise (t-Test, Figure S7C) |
|---|---|---|---|---|
| C13 Hydrocarbon (↓) | - | - | - | - |
| Inosine/Adenosine (↑) | - | - | - | - |
| Fructose-6-Phosphate (↑) | - | - | - | |
| Glucose-6-Phosphate or Similar (↑) | - | - | - | - |
| Citric Acid/Isocitric Acid (↑) | Citric Acid/Isocitric Acid (↓) | - | - | - |
| 2-Aminoadipic Acid (↑) | - | - | - | - |
| Heptadecanoic Acid/Octadecanol (↑) | - | - | - | - |
| Stearic acid (↑) | - | - | - | - |
| alpha-Tocopherol (↑) | - | - | alpha-Tocopherol (↓) | - |
| Linoleic Acid (↑) | Linoleic Acid (↑) | - | - | - |
| Oleic Acid (↑) | - | - | - | - |
| - | Aldopentoses (↑) | - | - | - |
| - | - | Glycine (↓) | - | - |
| - | - | - | - | Malic acid (↑) |
| - | - | - | - | Serine (↑) |
| - | - | - | - | Aspartic acid (↓) |
| - | - | - | - | Myoinositol (↑) |
| - | - | - | - | Glutamine (↓) |
| - | - | - | - | Gluconic acid-6-phosphate (↑) |
| - | - | - | - | Glutamic acid (↑) |
| - | - | - | - | Pyrophosphate (↑) |
| - | - | - | - | Campesterol (↑) |
| - | - | - | - | Phosphoric acid (↑) |
| - | - | - | - | Creatinine (↑) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starnes, J.W.; Parry, T.L.; O’Neal, S.K.; Bain, J.R.; Muehlbauer, M.J.; Honcoop, A.; Ilaiwy, A.; Christopher, P.M.; Patterson, C.; Willis, M.S. Exercise-Induced Alterations in Skeletal Muscle, Heart, Liver, and Serum Metabolome Identified by Non-Targeted Metabolomics Analysis. Metabolites 2017, 7, 40. https://doi.org/10.3390/metabo7030040
Starnes JW, Parry TL, O’Neal SK, Bain JR, Muehlbauer MJ, Honcoop A, Ilaiwy A, Christopher PM, Patterson C, Willis MS. Exercise-Induced Alterations in Skeletal Muscle, Heart, Liver, and Serum Metabolome Identified by Non-Targeted Metabolomics Analysis. Metabolites. 2017; 7(3):40. https://doi.org/10.3390/metabo7030040
Chicago/Turabian StyleStarnes, Joseph W., Traci L. Parry, Sara K. O’Neal, James R. Bain, Michael J. Muehlbauer, Aubree Honcoop, Amro Ilaiwy, Peter M. Christopher, Cam Patterson, and Monte S. Willis. 2017. "Exercise-Induced Alterations in Skeletal Muscle, Heart, Liver, and Serum Metabolome Identified by Non-Targeted Metabolomics Analysis" Metabolites 7, no. 3: 40. https://doi.org/10.3390/metabo7030040