Extracellular Microbial Metabolomics: The State of the Art


1
The New Zealand Institute for Plant & Food Research Limited, Private Bag 92169, Auckland 1142, New Zealand
2
School of Biological Sciences, University of Auckland, Private Bag 92019, Auckland 1010, New Zealand
*
Author to whom correspondence should be addressed.
Metabolites 2017, 7(3), 43; https://doi.org/10.3390/metabo7030043
Submission received: 31 July 2017
/
Revised: 21 August 2017
/
Accepted: 22 August 2017
/
Published: 22 August 2017
(This article belongs to the Special Issue Microbial Metabolomics Volume 2)
Abstract
:Microorganisms produce and secrete many primary and secondary metabolites to the surrounding environment during their growth. Therefore, extracellular metabolites provide important information about the changes in microbial metabolism due to different environmental cues. The determination of these metabolites is also comparatively easier than the extraction and analysis of intracellular metabolites as there is no need for cell rupture. Many analytical methods are already available and have been used for the analysis of extracellular metabolites from microorganisms over the last two decades. Here, we review the applications and benefits of extracellular metabolite analysis. We also discuss different sample preparation protocols available in the literature for both types (e.g., metabolites in solution and in gas) of extracellular microbial metabolites. Lastly, we evaluate the authenticity of using extracellular metabolomics data in the metabolic modelling of different industrially important microorganisms.
1. Introduction
Extracellular metabolomics is the study of low molecular weight extracellular metabolites that are secreted by microbial cells into their environment, namely the culture media. The entire complement of these metabolites is often referred to as the exometabolome. It includes all of the metabolites secreted outside the microbial cell, as well as those components of the microbial culture media that are found in the supernatant [1].
Analysis of the exometabolome yields information about microbial activities under different culture conditions which, coupled with the intracellular metabolic profile, provides a comprehensive overview of microbial metabolism. There are some advantages of extracellular metabolite analysis (metabolic footprinting) over the analysis of intracellular metabolites (metabolic fingerprinting). The separation of extracellular metabolites from microbial cells and their intracellular metabolites can be achieved by simple techniques such as centrifugation and filtration, while the extraction of intracellular metabolites from microbial cells is a complex process [2,3]. Sample preparation and the handling of extracellular samples are also comparatively easier than intracellular sample preparation. Necessary precautions do, however, need to be taken to avoid cross-contamination between intra- and extracellular metabolites [4].
Extracellular metabolites are mainly produced as by-products of the metabolic activities of microorganisms growing in a specific environment. There are various environmental factors which can affect the uptake and secretion of metabolites from and into their environment (i.e., culture medium) such as temperature, pH, the concentration of nutrients, and others. Consequently, the level of extracellular metabolites can be easily modified by changing any one, or more, of these factors [5]. Exometabolome analysis, or metabolic footprint analysis, is the global identification and quantification of the metabolites present in the spent culture medium of microbial cells using different analytical techniques. As the name suggests, the metabolic footprint is the microorganism’s footprint in the extracellular medium due to the cells' uptake of nutrients and secretion of other metabolites during growth [6]. This approach is both qualitative and semi-quantitative [5,7].
1.1. Applications and Implications of Metabolic Footprinting
Metabolic footprint analysis provides valuable information about the metabolism of different microorganisms that change in response to different environmental conditions. Therefore, this approach has been extensively applied in different areas, such as bioprocess monitoring, bioremediation, biomarker discovery, and functional genomics. Due to the rapid development of different technical platforms, the popularity of extracellular metabolite analysis has increased considerably in the last ten years. In the near future, analysis of extracellular metabolites will provide more substantial biological information because of the rapid advancement and accessibility of a wide range of different analytical approaches.
The availability of different analytical tools and high-throughput methods has made extracellular metabolite analysis an invaluable technique for monitoring bioprocesses. These technologies have been shown to be useful for the quality control of fermentation processes since they enable the monitoring of changes in metabolite levels during fermentation [5,8]. Metabolic footprinting has been used, for example, to study heterologous protein production [9,10], identify fermentation biomarkers [11], monitor wine aroma development [12,13], and to study metabolic interactions among different yeast strains [14].
In addition to monitoring bioprocesses, metabolic footprint analysis has also been used to monitor and assess the mineralization process of xenobiotic compounds (e.g., petroleum and pesticides), both in the environment [15] and in the laboratory [16]. It is desirable, in this process, to determine whether these compounds are undergoing complete degradation (to CO2 and H2O) or producing hazardous and more recalcitrant compounds [3]. Potentially, metabolic footprint analysis can be used to identify microorganisms which are able to mineralize xenobiotic compounds. Furthermore, if those microorganisms are pathogenic to humans and plants, it might be possible to employ metabolic engineering techniques to reconstruct new species or to transfer the required genes to non-pathogenic microorganisms [5,17].
Besides the usefulness of extracellular metabolomics in important areas like bioprocess and bioremediation, the integration of metabolic footprinting with other “omics” (i.e., proteomics and genomics) has also been undertaken successfully. These integrated approaches were applied efficiently for the identification and characterization of unknown genes responsible for the production of important enzymes involved in expedient and novel pathways. For instance, there is currently great interest in the industrial production of ethanol (biofuel) from agricultural waste [18,19,20]. This conversion is achieved by completely degrading plant lignocelluloses into fermentable sugars. Panagiotou et al. [21] studied the simultaneous saccharification and fermentation of cellulose to ethanol in a one-step process by using Fusarium oxysporum. They identified extracellular organic and amino acids, and found that the activation of the GABA shunt caused the inhibition of the fungus’ Kreb’s cycle. Using both metabolic footprinting and fingerprinting, they described a novel fungal metabolic pathway (Phosphoketolase pathway) involved in xylose metabolism [22,23]. There are a number of other studies that show the value of metabolic footprinting in unraveling the degradation processes of lignocelluloses and monitoring the fermentation behavior of some industrially important microorganisms [24,25,26].
Metabolic footprinting has also gained popularity in microbial functional genomics because of its utility in discriminating between microbial strains. It has been used, for instance, to discriminate between yeast strains by identifying extracellular metabolites related to insertion and deletion mutations [27,28]. In addition, mutant bacterial strains (Escherichia coli tryptophan mutants) have also been differentiated after analyzing the exometabolome [4]. This approach, combined with genomics data, is also a powerful and low-cost tool for characterizing and phenotyping single deletion mutants. For example, the phenotypic characterization of transposon inserted mutants of xylanolytic and proteolytic rumen bacteria Clostridium proteoclasticum B316T has been shown by analyzing extracellular metabolites [29].
Furthermore, analysis of extracellular metabolites that are secreted by the microorganisms into the culture media allows the identification of quorum sensing (QS) molecules, (i.e., compounds used in cell-to-cell communication in microbial communities) [30,31]. These metabolites need to be analyzed during the different growth phases of the microbial community to determine when the QS molecules are produced. Pseudomonas aeruginosa can cause a chronic lung infection by producing QS molecules that enable this microorganism to form biofilm and develop a resistance to antibiotics [32]. Microbial footprinting is a potential approach to developing drugs which inhibit the production of QS molecules in such pathogenic microorganisms [5].
Given that metabolic footprinting has already been successfully applied to various types of biotechnological research, this approach has already proved to be very powerful in unraveling innovative and novel information [9,29,33,34,35,36]. However, the analysis of extracellular metabolites cannot provide evidence about what is happening inside the cell [37], although recently a few software packages have become available to predict the intracellular metabolic statuses of cells using concentrations of extracellular metabolites [38,39]. However, to generate a holistic picture of cell metabolism, the application of both intracellular and extracellular metabolomics is highly recommended.
1.2. Dynamic or Time-Resolved Metabolic Footprinting
Recently, dynamic or time-resolved metabolic footprinting has gained considerable interest from the scientific community because this approach allows a better understanding of metabolites secreted by different microbial cells at different time intervals due to the changes in environmental conditions [40,41]. Traditional metabolite footprinting mostly relies on single time point data, thus the analysis of metabolites depends on the microbial growth rate in addition to other factors (e.g., inoculum size). On the other hand, dynamic metabolite footprinting gathers data on microbial growth at different time points [41]. Therefore, this approach is more data-rich and informative than the single point extracellular metabolite analysis [40], thus being an important tool for in-depth phenotyping.
The changes in growth media due to the presence of contaminants can also be easily determined by analyzing samples at different time intervals rather than at a single point. For example, Sue et al. [42] reported a proof of concept study where they used time series extracellular metabolomics data to detect microbial contamination in microalgal fermentation broths (Figure 1). These broths were deliberately contaminated with common contaminating microorganisms and monitored for nine hour periods at different time intervals to determine the changes in extracellular metabolite profiles. While traditional microbial detection and identification techniques can take two days, this type of metabolite footprinting provides a rapid and conclusive determination of microbial contaminants within a few hours in a batch fermentation process (Figure 1) [42]. Therefore, interventions can be undertaken sooner to avoid the further loss of product quality.
The main benefit of the dynamic or time-resolved metabolic footprinting approach is the ability to capture the changes in the metabolism of microorganisms with different growth rates (especially mutants). Therefore, it is possible to discriminate between mutants and also among the same microbial species. Behrends et al. [43] successfully used this approach to determine the differences between two closely related species of the Burkholderia cepacia complex. They also employed a similar approach to determine the differences between Pseudomonas aeruginosa wild type and its mutant (mucA22), and identified metabolites associated with osmotic tolerance (e.g., glutamate, trehalose, and glycine-betaine) that caused the main difference between wild-type and mutant strains [43].
The data analysis platform for handling large datasets obtained from dynamic or time-resolved metabolic footprinting has also been updated in recent years [41,44,45]. For instance, Chumnanpuen et al. [41] reported a novel data analysis method (cherry picking and correlation analysis) of MS data based on the identification of significantly correlated metabolites over time, which corresponds to flux ratios of different metabolites. This was followed by a reconstruction of interaction maps to provide more information on metabolic pathways associated with correlated metabolites, thus showing the importance of this approach in generating novel information relating to microbial metabolism [41].
2. Sample Preparation for the Analysis of Extracellular Metabolites
Sample preparation is a crucial part of any experimental procedure. In metabolomic analyses the first, and perhaps most critical, step is quenching to halt any biochemical reactions beyond the time of sampling. The microbial culture is then separated from the cells by centrifugation or filtration (Figure 2). The next step is the addition of internal standards that allow the detection and compensation of the technical variability during the sample preparation [46,47]. Moreover, to deactivate the enzymatic activity and concentrate the extracellular metabolites, the culture supernatant must be freeze-dried under vacuum before chemical derivatization. Any remaining culture medium can be stored in the dark at a low temperature (−20 °C or less).
Prior to performing experiments, several factors need to be considered carefully. Especially, the numbers of inoculated cells relative to culture medium volume is particularly of interest so that metabolites are secreted into the surrounding environment in a detectable amount. Microbial cells in a concentration of 105–106 C.F.U per mL are usually recommended for inoculation during extracellular metabolite analysis [42,48]. Moreover, it is also crucial to analyze the unspent culture medium as the control to determine the non-biological changes that occur during the experiments [49].
Extracellular metabolites can be present in solution or in the gas-phase [6]. The sample preparation varies considerably based on the state of the metabolites and the major considerations during the extraction and analysis of metabolites in a liquid or gaseous state will be outlined below:
2.1. Metabolites in Solution
Extracellular metabolites are present in the microbial culture media. The composition of liquid media and the possible presence of interferences (matrix), such as a high amount of salts, sugars, proteins, lipids, and sometimes even water, can cause problems with the operation of analytical equipment. As a result, metabolites that are present in low concentrations may not be detected in the biological samples. Therefore, problems caused by the sample matrix need to be identified and resolved to ensure the acquisition of informative and accurate extracellular metabolite profiles [50]. There are several options for overcoming the difficulties instigated by the matrix of a biological sample, e.g.,
- (a)
- liquid-liquid separation where metabolites of interest are separated into an immiscible solvent,
- (b)
- using a column or solid-phase matrix to trap the metabolites, and
- (c)
- selective solubilization, which is the complete evaporation of the solvent to concentrate the sample and the metabolites are then dissolved with suitable solvents.
The use of solvents in the liquid-liquid separation of metabolites from extracellular media is very laborious and time consuming, making it unpopular for metabolite profiling. This approach is also known to result in incomplete phase separations and lower recovery rates. In addition, this type of separation also requires the use of large amounts of glassware and organic solvents. A better separation of metabolites, however, is achieved via solid-phase extraction to trap metabolites of interest [6] and selective solubilization is also a widely used technique for the analysis of extracellular metabolites in solution. However, there are two commonly used methods for the separation of targeted metabolites that are present in solution, namely solid-phase extraction (SPE) and solid-phase microextraction (SPME) (Figure 3). The technical details of these two separation procedures will be discussed in the next subsections.
2.1.1. Solid-Phase Extraction
Solid-phase extraction (SPE) is a sample separation and extraction method, which causes the exhaustive removal of metabolites from a liquid sample resulting in targeted analytes being retained on a solid sorbent [51]. SPE is usually used to prepare liquid samples and for the extraction of semi-volatile and non-volatile metabolites. It is more efficient than liquid-liquid separation and is easier to perform. Both solid phase and liquid phase are used to isolate one type of analyte from a solution. SPE is widely used to concentrate the metabolites of interest, mainly for targeted analysis, when they are in very low concentrations [52]. Moreover, it is very useful to remove impurities and interfering matrix components from samples or to prepare the sample in solution for subsequent analysis [6].
Different types of SPE sorbent materials, such as silica, alkylated silica (e.g., C-18), carbon-based sorbents, ion exchange materials, polymer materials, and restricted access materials (RAM) are commercially available for different types of metabolite extraction. Mixed-mode sorbents are a newer development and are based on multiple retention mechanisms due to incorporating diverse ligands in the same sorbent [53].
The SPE cartridge, or stationary phase, is contained in a glass or plastic column above a frit or glass wool. The column has a frit on the top and there is a stopcock that controls the flow of solvent in the column. The cartridges are placed on a vacuum manifold which is used to control the solvent flow rate through the cartridge. Collection tubes are placed under the cartridge to collect the waste liquid after washing the column. During elution, clean tubes replace those waste tubes to collect the final sample containing the metabolites of interest [6]. The trapped metabolites in the SPE cartridge can then be eluted by using an appropriate extraction solvent of sufficient strength [53], selected on the basis of its polarity, pH, and mobile phase ionic strength [6].
Three different SPE stationary phases, such as normal, reversed, and ion-exchange, are available with SPE cartridges and disks. These stationary phases are mainly silica-based and commonly used in the preparation of samples for metabolomics analysis, allowing the separation of metabolites according to their chemical properties (Table 1).
● Normal Phase
A non-polar matrix and polar stationary phase are used to extract non-polar analytes. The retention of analytes depends on chemical interactions (e.g., hydrogen bonding, pi-pi interactions, dipole-dipole interactions, etc.) between functional groups of analytes and sorbents. The trapped analytes are eluted by a more polar solvent which is able to break up the binding mechanism.
● Reversed Phase
Reversed phase SPE has a mobile phase with a polar or moderately polar matrix and a non-polar stationary phase. This non-polar stationary phase is derivatized with hydrocarbon chains that allow the retention of non-polar metabolites because of the hydrophobic effect. Non-polar forces involved in this phase are known as van der Wals or dispersion forces. A non-polar solvent is also used to elute the analytes.
● Ion-Exchange Phase
Metabolites which are charged while in solution can be extracted by using an ion-exchange phase. The retention of analytes is due to the electrostatic attraction of the charged functional group of the metabolite to the charged group of the matrix. A solvent with a pH level that neutralizes these charged groups is used to elute the analytes.
SPE is a sensitive method that enables the simultaneous analysis of a number of metabolites by limiting possible ion suppression effects [54]. The main limitation of SPE in metabolomics is its selectivity. Thus, it is suitable for targeted analyses, but not for global metabolite profiling. While the use of SPE for extracellular metabolite extraction is not very common, it has been successfully applied to purify and identify antifungal compounds (phenyllacticacid, 4-hydroxyphenyllactic acid, benzoic acid, methylhydantoin, mevalonolactone, etc.) and some other small peptides from lactic acid bacteria [55,56]. This sample preparation technique has great potential for the extraction and purification of antibacterial compounds (small analytes) secreted into the extracellular media.
2.1.2. Solid-Phase Microextraction
Solid-phase microextraction (SPME) is a portable small sample preparation technique invented by Pawliszyn and colleagues which is both fast and simple [57,58,59]. The development of an automated SPME method made this technique suitable for the high-throughput analysis of samples [60,61]. In SPME, a small amount of sample (<100 µL) is exposed for a specific time period to a small amount of extracting phase dispersed on a solid support allowing metabolite extraction. Two approaches are available. In one, a partitioning equilibrium occurs and it causes convection between the sample matrix and the extraction phase. The number of metabolites extracted is independent of convection conditions. The other approach uses a short-time pre-equilibrium where convection is constant and, as a result, the amount of extracted metabolite is time dependent [62].
The first SPME device consisted of coated fibers in a micro-syringe referred to as a fiber SPME. A polymer-coated fused-silica fiber was exposed to a sample for a predetermined time until equilibrium was reached. The fiber was then removed from the solution to desorb the analytes into the injector of an analytical instrument [63]. Different types of SPME coating are commercially available today, which are able to extract a wide range of metabolites [64].
Even though the use of SPME allows the separation of many metabolites, this technique has both advantages and disadvantages. For instance, it is resource efficient in that it does not require solvent for the elution of analytes and the fibers are reusable. Furthermore, SPME is portable so that it can be used in the field, as well as in the laboratory. However, the coated fibers are expensive, fragile, and have a limited extraction capacity, resulting in poor sensitivity [6,59]. Although SPME is a popular technique because of its simplicity, sensitivity, and reproducibility, the optimization of sampling conditions and set up of suitable experiments are required to obtain good quality data [65].
2.2. Metabolites in Gas Phase
Volatile compounds, or gaseous metabolites, present in extracellular samples provide precious information on different properties of microorganisms and/or their phenotypes and their metabolic pathways. Metabolites already in gas phase are easy to analyze by GC-MS and require no further derivatization (Figure 2). Thus, their analysis takes less time than non-volatile compounds. However, they are very difficult to collect due to their high diffusion rates and also because of their very low concentrations (sometimes near or below the detection limit) [6]. Thus, to improve their detection by analytical instruments, the metabolites must be captured and concentrated before analysis.
Several methods have been developed in recent years to trap and concentrate volatile compounds in extracellular samples. SPME methods using adsorptive materials, such as porous carbon, or sorptive polymers, are often used to analyze volatile compounds. Headspace analysis coupled with SPME is another alternative technique employed to analyze these compounds.
SPME and Headspace Analysis of Volatile Compounds
SPME is considered a promising sample preparation technique for the extraction of volatile compounds from microorganisms because of its ability to separate metabolites with a wide range of properties (both in liquid and solid samples) at a very low concentration (ppt to ppm) [66]. SPME and headspace analysis are often used in combination with GC-MS to analyze the metabolites in gas phase. Volatile compounds from microbial liquid cultures and fermentation flasks can easily be captured from the amount of headspace gas above the culture and they can then be detected and identified by GC-MS. This can be done in two ways:
- Direct headspace analysis where volatile compounds are collected into a syringe and analyzed by GC.
- Headspace SPME (HS-SPME) which is a coupled technique in which the gas sample in the headspace is trapped on the SPME fiber [6].
In headspace analysis, a liquid sample is collected and heated to vaporize in the headspace. Both manual and automated systems can be applied; however, automated systems show better reproducibility [67]. The headspace analysis of a microbial culture provides qualitative results about volatile compounds [68]. The microbial contamination of food can also be determined by collecting the volatile metabolites in the headspace gas and then analyzing those using GC-MS or an electronic nose [69]. HS-SPME is a popular solvent-free method employed for analyzing microbial samples. Volatile compounds are isolated and concentrated without any derivatization [70]. This fast, simple, and sensitive method yields highly reproducible results. Moreover, HS-SPME is more advantageous compared to direct SPME. For instance, it requires less time to extract the targeted metabolites due to the faster diffusion rate of the analytes in the gas phase. Moreover, matrix effects can also be reduced by using HS-SPME. Overall, the use of SPME along with HS provides highly reproducible results, which is obviously very useful for the analysis of metabolites in the gas phase.
3. Concentration of Extracellular Samples to Improve Detection
Sample concentration is an important aspect of metabolome analysis because metabolites are diluted in both intracellular and extracellular samples. Concentration of the samples is necessary before analysis by analytical tools to improve the limit of detections [47,71]. If the extracellular samples are concentrated, they can be derivatized or analyzed directly [47]. Lyophilization, or freeze drying, is commonly used to concentrate extracellular samples by the removal of water. Conversely, organic solvent evaporating systems or vacuum drying methods are also used to remove water from samples, although the thermal degradation of metabolites needs to be considered when these techniques are used. Non-aqueous extracellular samples are, however, concentrated using solvent evaporation techniques.
3.1. Freeze-Drying
Freeze-drying is a widely used sample concentration method in metabolomics where samples are immediately frozen and then dried by sublimation to remove all of the frozen solvents. Freeze-drying is considered a non-destructive sample concentration and dehydration process that avoids the thermal degradation of compounds. A variety of manual and completely automated freeze-dryers are commercially available these days, including facilities to dry samples under a very low temperature (–65 °C). Some “smart” freeze-drying instruments, for example, a manometric temperature measurement (MTM), measure the metabolite temperature by rapidly isolating the freeze-drying chamber from the condenser while also monitoring the pressure [72]. Recent developments in freeze-drying techniques are very useful and efficient for concentrating samples for both intracellular and extracellular metabolite analysis [73]. It is, however, a slow and complex procedure that may be not be suitable for metabolites that are susceptible to oxidation. To overcome the oxidative degradation of metabolites, inert gas (e.g., nitrogen) must be used to break the vacuum [47].
The nature of the metabolites and the sample matrix need to be considered carefully while concentrating the metabolomics samples. Moreover, appropriate precautions also need to be followed to overcome problems, e.g., the thermal and oxidative degradation of labile metabolites. Factors such as the solution pH and sample matrix are very important and these may cause problems while freeze-drying the samples. For instance, extracellular samples with a high sugar concentration are almost impossible to dry completely, leaving only sticky undried samples. To avoid this, such samples must be diluted prior to freeze-drying. Moreover, samples containing a high volume of organic solvent cannot be frozen and must have water added to increase the water:solvent ratio. This then enables the sample to be frozen but also increases the drying time. After freeze-drying, another issue is the resuspension of samples because metabolite losses may occur due to the incomplete recovery of the dried samples [74,75]. However, besides having some minor disadvantages, freeze-drying is still a method of choice for many laboratories because of its efficiency in terms of the sample concentration.
3.2. Vacuum-Drying
The vaccum-drying procedure using a speed-vac instrument makes use of a combination of the centrifugal force, heat, and vacuum to remove moisture from a given sample. This process dries a sample through evaporation, mainly by converting the liquid to vapor. It is mostly used to concentrate a small amount of sample to complete dryness. Therefore, vacuum-drying, being faster and comparatively less aggressive than freeze-drying, is preferable in some cases [6]. For instance, the reagents for trimethyl silyl derivatization are extremely sensitive to moisture, and thus, the vacuum-drying of samples is often preferable [26,36]. However, vacuum-drying often requires the use of heat; therefore, thermos-labile metabolites might be degraded during the process. Sometimes, the addition of extra solvent (e.g., methanol) is also required, which assists in the fast evaporation of samples [76].
4. Storage of Extracellular Microbial Samples
Once extracellular samples have been extracted and concentrated, the next step is to store them under appropriate conditions prior to analysis. To avoid any unwanted changes during the storage, necessary precautions need to be undertaken. Storage conditions and time are, however, dependent on the types and stability of metabolites. Even though metabolomic samples are quenched before the separation and extraction of extracellular metabolites, it is better to ensure that no chemical reaction is taking place during the storage. Therefore, it is highly recommended that the extracellular samples must be stored in the dark and at a very low temperatures (e.g., −20 °C, −80 °C) to avoid the degradation caused by light and heat. This way, it is possible to retain the original properties of metabolites.
Some metabolites can lose their functionality during storage at low temperature and can be oxidized easily, and this may completely alter their characteristics. To protect them, they must be kept under vacuum [6]. Stable metabolites (i.e., do not oxidize and are unaffected by exposure to normal temperature) can be stored at room temperature and mostly, the samples concentrated to powder that do not contain any water are suitable for storage under vacuum at room temperature. However, this is not a common practice in metabolomic laboratories. Setting up appropriate storage conditions for all types of metabolomic samples is a primary responsibility of the laboratories in order to generate reproducible and high-quality data.
5. Integration of Extracellular Metabolomics Data to Genome Scale Metabolic Models
The genome-scale reconstruction of entire organisms and modelling of their physiology in a predictive manner is a comparatively new approach and has been successfully used in systems metabolic engineering to design microbial cell factories [77,78,79,80,81,82]. This is an emerging interdisciplinary field that makes use of a systems-level view of an organism to determine the genotype-phenotype relationship, without the need for kinetic parameters [83,84]. The network is explored by imposing physico-chemical constraints (e.g., omics data, cell growth) and by testing the cell efficiency under optimality criteria [85]. The high amount of data generated from different omics approaches, especially from metabolomic studies, can now be integrated into a genome-scale model to gain novel insights on microbial metabolism [86].
Recent developments in profiling extracellular metabolites are now allowing us to gather a large amount of information on microbial systems. These extracellular metabolites are either present in the culture media or secreted by the microorganisms during growth due to experimental perturbations. Therefore, these data contain valuable information about the cell metabolism and how the environment can affect the utilization and production of various metabolites, indicating the alteration of intracellular status. The generation of intracellular metabolites data is comparatively troublesome, mainly because of the need to rupture the cell in order to release the metabolites [9,26,86]. Therefore, the integration of extracellular metabolomics data into metabolic models is clearly an advantageous approach.
The integration of extracellular metabolomics data into genome-scale models is now allowing the prediction of the intracellular status of microorganisms. For instance, Cakir et al. (2007) incorporated small-scale extracellular metabolomics data with a genome-scale metabolic model of yeast to predict the oxygen consumption and ethanol production ability of mutant strains with respiratory deficiencies [87]. A few years later, Mo et al. (2009) published a study that integrated a large extracellular metabolite profiling data into a highly curated genome-scale metabolic model of Saccharomyces cerevisiae iMM904 to determine how the variation in extracellular metabolite levels is associated with changes in intracellular metabolites fluxes [86]. They also successfully validated the predictions generated from the model.
A comprehensive and detailed protocol entitled “MetaboTools” is now available for integrating extracellular metabolomics data obtained from microorganisms, plants, and animals [39]. This toolbox integrates the extracellular metabolite profiles into the network context and generates contextualized models that contain a subset of metabolic models. These metabolic models are designed to predict the intracellular pathways based on the extracellular metabolites by determining the utilization and production of different metabolites under different environmental conditions [38,39]. Although this approach of integrating extracellular metabolomics data into metabolic models and the prediction of intracellular status is gaining popularity, a comprehensive status of microbial metabolism can only be obtained by combining both intra- and extracellular metabolomic datasets [37].
6. Conclusions
The analysis of extracellular metabolites from a microbial culture provides crucial information about the intracellular status of a microbial system that occurs in response to the surrounding environment. Therefore, this approach has already been used widely in both fundamental and applied research [88,89,90,91]. However, there is still scope to improve the identification and quantification of metabolites in a wide range of biological samples and it would be useful for the analysis of both extracellular and intracellular samples. The sets of information gained from both intracellular and extracellular microbial samples are complementary to each other, which will allow an understanding of the complete overview of microbial metabolism.
Acknowledgments
F.R.P. received funding from The Agricultural and Marketing Research and Development Trust (AGMARDT; contract number A16017).
Author Contributions
F.R.P. and S.V.B. conceived the idea, and F.R.P. wrote and revised the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mashego, M.R.; Van Gulik, W.M.; Heijnen, J.J. Metabolome dynamic responses of Saccharomyces cerevisiae to simultaneous rapid perturbations in external electron acceptor and electron donor. FEMS Yeast Res. 2007, 7, 48–66. [Google Scholar] [CrossRef] [PubMed]
- Tredwell, G.D.; Edwards-Jones, B.; Leak, D.J.; Bundy, J.G. The development of metabolomic sampling procedures for pichia pastoris, and baseline metabolome data. PLoS ONE 2011, 6, e16286. [Google Scholar] [CrossRef] [PubMed]
- Villas-Bôas, S.G.; Bruheim, P. The potential of metabolomics tools in bioremediation studies. OMICS A J. Integr. Biol. 2007, 11, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Kaderbhai, N.N.; Broadhurst, D.I.; Ellis, D.I.; Goodacre, R.; Kell, D.B. Functional genomics via metabolic footprinting: Monitoring metabolite secretion by Escherichia coli tryptophan metabolism mutants using FT-IR and direct injection electrospray mass spectrometry. Comp. Funct. Genom. 2003, 4, 376–391. [Google Scholar] [CrossRef] [PubMed]
- Mapelli, V.; Olsson, L.; Nielsen, J. Metabolic footprinting in microbiology: Methods and applications in functional genomics and biotechnology. Trends Biotechnol. 2008, 26, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Villas-Bôas, S.G.; Roessner, U.; Hansen, M.E.; Smedsgaard, J.; Nielsen, J. Metabolome Analsis An Introduction; Wiley: Hoboken, NJ, USA, 2007. [Google Scholar]
- Oldiges, M.; Lütz, S.; Pflug, S.; Schroer, K.; Stein, N.; Wiendahl, C. Metabolomics: Current state and evolving methodologies and tools. Appl. Microbiol. Biotechnol. 2007, 76, 495–511. [Google Scholar] [CrossRef] [PubMed]
- McGovern, A.C.; Broadhurst, D.; Taylor, J.; Kaderbhai, N.; Winson, M.K.; Small, D.A.; Rowland, J.J.; Kell, D.B.; Goodacre, R. Monitoring of complex industrial bioprocesses for metabolite concentrations using modern spectroscopies and machine learning: Application to gibberellic acid production. Biotechnol. Bioeng. 2002, 78, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, S.; Villas-Bôas, S.G.; Ferreira, E.C.; Rocha, I. Metabolic footprint analysis of recombinant Escherichia coli strains during fed-batch fermentations. Mol. Biosyst. 2011, 7, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Graf, A.; Dragosits, M.; Gasser, B.; Mattanovich, D. Yeast systems biotechnology for the production of heterologous proteins. FEMS Yeast Res. 2009, 9, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Yang, H.J.; Kim, M.J.; Han, E.S.; Kim, H.J.; Kwon, D.Y. Metabolomic analysis of meju during fermentation by ultra performance liquid chromatography-quadrupole-time of flight mass spectrometry (UPLC-Q-TOF MS). Food Chem. 2011, 127, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Hong, Y.S.; Lee, C.H. Characterization of fermentative behaviors of lactic acid bacteria in grape wines through 1H NMR- and GC-based metabolic profiling. J. Agric. Food Chem. 2009, 57, 4810–4817. [Google Scholar] [CrossRef] [PubMed]
- Rossouw, D.; Næs, T.; Bauer, F.F. Linking gene regulation and the exo-metabolome: A comparative transcriptomics approach to identify genes that impact on the production of volatile aroma compounds in yeast. BMC Genom. 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Howell, K.S.; Cozzolino, D.; Bartowsky, E.J.; Fleet, G.H.; Henschke, P.A. Metabolic profiling as a tool for revealing saccharomyces interactions during wine fermentation. FEMS Yeast Res. 2006, 6, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Han, Z.; Bai, Z.; Zhuang, G.; Shim, H. Progress in decontamination by halophilic microorganisms in saline wastewater and soil. Environ. Pollut. 2010, 158, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.R.; Luo, Z.H.; Kwok-Kei Chow, R.; Vrijmoed, L.L.P. Purification and characterization of an extracellular laccase from the anthracene-degrading fungus fusarium solani MAS2. Bioresour. Technol. 2010, 101, 9772–9777. [Google Scholar] [CrossRef] [PubMed]
- Parrilli, E.; Papa, R.; Tutino, M.L.; Sannia, G. Engineering of a psychrophilic bacterium for the bioremediation of aromatic compounds. Bioeng. Bugs 2010, 1, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Plácido, J.; Imam, T.; Capareda, S. Evaluation of ligninolytic enzymes, ultrasonication and liquid hot water as pretreatments for bioethanol production from cotton gin trash. Bioresour. Technol. 2013, 139, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.P.; Trivedi, R.K. Ethanol, an Economical & Environmentally Feasible Way of Biofuel from Cellulosic Materials: Process and Discussion. Available online: https://www.ripublication.com/Volume/ijaerv7n11_spl..htm (accessed on 22 August 2017).
- Zain, N.A.M.; Shen, N.S.; Suhaimi, M.S.; Hasan, N.B.; Aziman, S.N. Potential use of liquid pinapple waste for bioethanol production by immobilized bakers‘ yeast. Jurnal Teknologi 2012, 59, 43–47. [Google Scholar]
- Panagiotou, G.; Christakopoulos, P.; Olsson, L. The influence of different cultivation conditions on the metabolome of Fusarium oxysporum. J. Biotechnol. 2005, 118, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Panagiotou, G.; Anderson, M.R.; Grotkjær, T.; Regueira, T.B.; Hofmann, G.; Nielsen, J.; Olsson, L. Systems analysis unfolds the relationship between the phosphoketolase pathway and growth in aspergillus nidulans. PLoS ONE 2008, 3, e3847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panagiotou, G.; Kouskoumvekaki, I.; Jónsdóttir, S.O.; Olsson, L. Monitoring novel metabolic pathways using metabolomics and machine learning: Induction of the phosphoketolase pathway in aspergillus nidulans cultivations. Metabolomics 2007, 3, 503–516. [Google Scholar] [CrossRef]
- Meijer, S.; Panagiotou, G.; Olsson, L.; Nielsen, J. Physiological characterization of xylose metabolism in aspergillus niger under oxygen-limited conditions. Biotechnol. Bioeng. 2007, 98, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Panagiotou, G.; Pachidou, F.; Petroutsos, D.; Olsson, L.; Christakopoulos, P. Fermentation characteristics of Fusarium oxysporum grown on acetate. Bioresour. Technol. 2008, 99, 7397–7401. [Google Scholar] [CrossRef] [PubMed]
- Villas-Bôas, S.G.; Noel, S.; Lane, G.A.; Attwood, G.; Cookson, A. Extracellular metabolomics: A metabolic footprinting approach to assess fiber degradation in complex media. Anal. Biochem. 2006, 349, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.; Davey, H.M.; Broadhurst, D.; Heald, J.K.; Rowland, J.J.; Oliver, S.G.; Kell, D.B. High-throughput classification of yeast mutants for functional genomics using metabolic footprinting. Nat. Biotechnol. 2003, 21, 692–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mas, S.; Villas-Bôas, S.G.; Hansen, M.E.; Åkesson, M.; Nielsen, J. A comparison of direct infusion ms and GC-MS for metabolic footprinting of yeast mutants. Biotechnol. Bioeng. 2007, 96, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Villas-Bôas, S.G.; Moon, C.D.; Noel, S.; Hussein, H.; Kelly, W.J.; Cao, M.; Lane, G.A.; Cookson, A.L.; Attwood, G.T. Phenotypic characterization of transposon-inserted mutants of Clostridium proteoclasticum B316T using extracellular metabolomics. J. Biotechnol. 2008, 134, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, S.; Williams, P. Quorum sensing and social networking in the microbial world. J. R. Soc. Interface 2009, 6, 959–978. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, E.P. Sociomicrobiology: A personal perspective on an emerging research area. Microbe 2010, 5, 206–211. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Jensen, P.Ø.; Jakobsen, T.H.; Phipps, R.; Nielsen, A.K.; Rybtke, M.T.; Tolker-Nielsen, T.; Givskov, M.; Høiby, N.; Ciofu, O. Quorum sensing and virulence of pseudomonas aeruginosa during lung infection of cystic fibrosis patients. PLoS ONE 2010, 5, e10115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pope, G.A.; MacKenzie, D.A.; Defernez, M.; Aroso, M.A.M.M.; Fuller, L.J.; Mellon, F.A.; Dunn, W.B.; Brown, M.; Goodacre, R.; Kell, D.B.; et al. Metabolic footprinting as a tool for discriminating between brewing yeasts. Yeast 2007, 24, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Villas-Bôas, S.G.; Moxley, J.F.; Åkesson, M.; Stephanopoulos, G.; Nielsen, J. High-throughput metabolic state analysis: The missing link in integrated functional genomics of yeasts. Biochem. J. 2005, 388, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.W.M.; Lorkiewicz, P.K.; Sellers, K.; Moseley, H.N.B.; Higashi, R.M.; Lane, A.N. Stable isotope-resolved metabolomics and applications for drug development. Pharm. Ther. 2012, 133, 366–391. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.; Edwards, P.B.; Jouanneau, S.; Kilmartin, P.; Gardner, R.; Villas-Boas, S. Sauvignon blanc metabolomics: Grape juice metabolites affecting the development of varietal thiols and other aroma compounds in wines. Metabolomics 2013, 556–573. [Google Scholar] [CrossRef]
- Granucci, N.; Pinu, F.R.; Han, T.L.; Villas-Boas, S.G. Can we predict the intracellular metabolic state of a cell based on extracellular metabolite data? Mol. Biosyst. 2015, 11, 3297–3304. [Google Scholar] [CrossRef] [PubMed]
- Aurich, M.K.; Paglia, G.; Rolfsson, O.; Hrafnsdottir, S.; Magnusdottir, M.; Stefaniak, M.M.; Palsson, B.O.; Fleming, R.M.T.; Thiele, I. Prediction of intracellular metabolic states from extracellular metabolomic data. Metabolomics 2015, 11, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aurich, M.K.; Fleming, R.M.T.; Thiele, I. Metabotools: A comprehensive toolbox for analysis of genome-scale metabolic models. Front. Physiol. 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Behrends, V.; Ebbels, T.M.D.; Williams, H.D.; Bundy, J.G. Time-resolved metabolic footprinting for nonlinear modeling of bacterial substrate utilization. Appl. Environ. Microbiol. 2009, 75, 2453–2463. [Google Scholar] [CrossRef] [PubMed]
- Chumnanpuen, P.; Hansen, M.A.E.; Smedsgaard, J.; Nielsen, J. Dynamic metabolic footprinting reveals the key components of metabolic network in yeast saccharomyces cerevisiae. Int. J. Genom. 2014. Available online: https://www.hindawi.com/journals/ijg/2014/894296/abs/ (accessed on 19 August 2017).
- Sue, T.; Obolonkin, V.; Griffiths, H.; Villas-Bôas, S.G. An exometabolomics approach to monitoring microbial contamination in microalgal fermentation processes by using metabolic footprint analysis. Appl. Environ. Microbiol. 2011, 77, 7605–7610. [Google Scholar] [CrossRef] [PubMed]
- Behrends, V.; Ryall, B.; Wang, X.; Bundy, J.G.; Williams, H.D. Metabolic profiling of pseudomonas aeruginosa demonstrates that the anti-sigma factor muca modulates osmotic stress tolerance. Mol. Biosyst. 2010, 6, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zeng, J.; Zhou, L.N.; Hu, C.X.; Yin, P.Y.; Lin, X.H. A new strategy for analyzing time-series data using dynamic networks: Identifying prospective biomarkers of hepatocellular carcinoma. Sci. Rep. 2016, 6, 32448. [Google Scholar] [CrossRef] [PubMed]
- Smilde, A.K.; Westerhuis, J.A.; Hoefsloot, H.C.J.; Bijlsma, S.; Rubingh, C.M.; Vis, D.J.; Jellema, R.H.; Pijl, H.; Roelfsema, F.; van der Greef, J. Dynamic metabolomic data analysis: A tutorial review. Metabolomics 2010, 6, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Villas-Bôas, S.G.; Koulman, A.; Lane, G.A. Analytical methods from the perspective of method standardization. In Metabolomics; Springer: Berlin/Heidelberg, Germany, 2007; Volume 18, pp. 11–52. [Google Scholar]
- Smart, K.F.; Aggio, R.B.M.; Van Houtte, J.R.; Villas-Boas, S.G. Analytical platform for metabolome analysis of microbial cells using methyl chloroformate derivatization followed by gas chromatography-mass spectrometry. Nat. Protoc. 2010, 5, 1709–1729. [Google Scholar] [CrossRef] [PubMed]
- Japelt, K.B.; Christensen, J.H.; Villas-Boas, S.G. Metabolic fingerprinting of lactobacillus paracasei: The optimal quenching strategy. Microb. Cell Fact. 2015, 14, 10. [Google Scholar] [CrossRef] [PubMed]
- Duportet, X.; Aggio, R.B.M.; Carneiro, S.; Villas-Bôas, S.G. The biological interpretation of metabolomic data can be misled by the extraction method used. Metabolomics 2012, 8, 410–421. [Google Scholar] [CrossRef]
- Álvarez-Sánchez, B.; Priego-Capote, F.; Castro, M.D.L.D. Metabolomics analysis II. Preparation of biological samples prior to detection. Trends Anal. Chem. 2010, 29, 120–127. [Google Scholar] [CrossRef]
- Dettmer, K.; Aronov, P.A.; Hammock, B.D. Mass spectrometry-based metabolomics. Mass Spectrom. Rev. 2007, 26, 51–78. [Google Scholar] [CrossRef] [PubMed]
- Kraly, J.R.; Holcomb, R.E.; Guan, Q.; Henry, C.S. Review: Microfluidic applications in metabolomics and metabolic profiling. Anal. Chim. Acta 2009, 653, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S. Sample Preparation Techniques in Analytical Chemsitry; Whiley: Hoboken, NJ, USA, 2003. [Google Scholar]
- Theodoridis, G.; Gika, H.; Franceschi, P.; Caputi, L.; Arapitsas, P.; Scholz, M.; Masuero, D.; Wehrens, R.; Vrhovsek, U.; Mattivi, F. LC-MS based global metabolite profiling of grapes: Solvent extraction protocol optimisation. Metabolomics 2012, 8, 175–185. [Google Scholar] [CrossRef]
- Ström, K.; Sjögren, J.; Broberg, A.; Schnürer, J. Lactobacillus plantarum milab 393 produces the antifungal cyclic dipeptides Cyclo(L-Phe-L-Pro) and Cyclo(L-Phe-Trans-4-OH-L-Pro) and 3-phenyllactic acid. Appl. Environ. Microbiol. 2002, 68, 4322–4327. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.J.; Chang, H.C. Purification of a new antifungal compound produced by lactobacillus plantarum af1 isolated from kimchi. Int. J. Food Microbiol. 2010, 139, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Pawliszyn, J. New directions in sample preparation for analysis of organic compounds. Trends Anal. Chem. 1995, 14, 113–122. [Google Scholar] [CrossRef]
- Pawliszyn, J. Chapter 13 Solid Phase Microextraction; Wiley: Hoboken, NJ, USA, 2002; Volume 37, pp. 389–477. [Google Scholar]
- Risticevic, S.; Chen, Y.; Kudlejova, L.; Vatinno, R.; Baltensperger, B.; Stuff, J.R.; Hein, D.; Pawliszyn, J. Protocol for the development of automated high-throughput SPME-GC methods for the analysis of volatile and semivolatile constituents in wine samples. Nat. Protoc. 2010, 5, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, G.; Zhao, W.; Pawliszyn, J. Kinetic calibration for automated headspace liquid-phase microextraction. Anal. Chem. 2005, 77, 8122–8128. [Google Scholar] [CrossRef] [PubMed]
- Vuckovic, D.; Zhang, X.; Cudjoe, E.; Pawliszyn, J. Solid-phase microextraction in bioanalysis: New devices and directions. J. Chromatogr. A 2010, 1217, 4041–4060. [Google Scholar] [CrossRef] [PubMed]
- Lord, H.; Pawliszyn, J. Evolution of solid-phase microextraction technology. J. Chromatogr. A 2000, 885, 153–193. [Google Scholar] [CrossRef]
- Pawliszyn, J. New developments and applications of solvent-free sampling and sample preparation technologies for the investigation of living systems. Aust. J. Chem. 2003, 56, 155–158. [Google Scholar] [CrossRef]
- Ouyang, G.; Pawliszyn, J. Recent developments in SPME for on-site analysis and monitoring. Trends Anal. Chem. 2006, 25, 692–703. [Google Scholar] [CrossRef]
- Jeleń, H.H. Use of solid phase microextraction (SPME) for profiling fungal volatile metabolites. Lett. Appl. Microbiol. 2003, 36, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Pawliszyn, J. Water analysis by solid phase microextraction based on physical chemical properties of the coating. Anal. Chem. 1997, 69, 1992–1998. [Google Scholar]
- Stansbridge, E.M.; Mills, G.A.; Walker, V. Automated headspace gas chromatographic analysis of faecal short-chain fatty acids. J. Chromatogr.-Biomed. Appl. 1993, 621, 7–13. [Google Scholar] [CrossRef]
- Demyttenaere, J.C.R.; Moriña, R.M.; Sandra, P. Monitoring and fast detection of mycotoxin-producing fungi based on headspace solid-phase microextraction and headspace sorptive extraction of the volatile metabolites. J. Chromatogr. A 2003, 985, 127–135. [Google Scholar] [CrossRef]
- Siripatrawan, U.; Linz, J.E.; Harte, B.R. Detection of Escherichia coli in packaged alfalfa sprouts with an electronic nose and an artificial neural network. J. Food Prot. 2006, 69, 1844–1850. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.A.; Yu, S.J.; Zhang, L.; Chen, X.D. The effects of ac electric field on wine maturation. Innov. Food Sci. Emerg. Technol. 2008, 9, 463–468. [Google Scholar] [CrossRef]
- Villas-Bôas, S.G.; Højer-Pedersen, J.; Åkesson, M.; Smedsgaard, J.; Nielsen, J. Global metabolite analysis of yeast: Evaluation of sample preparation methods. Yeast 2005, 22, 1155–1169. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Nail, S.L.; Pikal, M.J. Freeze-drying process design by manometric temperature measurement: Design of a smart freeze-dryer. Pharm. Res. 2005, 22, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T. Lyophilizer qualification: Some practical advice. Drugs Pharm. Sci. 2004, 135, 517–534. [Google Scholar]
- Oikawa, A.; Otsuka, T.; Jikumaru, Y.; Yamaguchi, S.; Matsuda, F.; Nakabayashi, R.; Takashina, T.; Isuzugawa, K.; Saito, K.; Shiratake, K. Effects of freeze-drying of samples on metabolite levels in metabolome analyses. J. Sep. Sci. 2011, 34, 3561–3567. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.H.; Yu, H.L.; Ba, Z.Y.; Chen, J.Y.; Sun, H.G.; Han, B.Z. Sampling methods for NMR-based metabolomics of staphylococcus aureus. Biotechnol. J. 2010, 5, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.R.; De Carvalho-Silva, S.; Uetanabaro, A.P.T.; Villas-Boas, S.G. Vinegar metabolomics: An explorative study of commercial balsamic vinegars using gas chromatography-mass spectrometry. Metabolites 2016, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Forster, J.; Famili, I.; Fu, P.; Palsson, B.O.; Nielsen, J. Genome-scale reconstruction of the Saccharomyces cerevisiae metabolic network. Genom. Res. 2003, 13, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Heavner, B.D.; Price, N.D. Comparative analysis of yeast metabolic network models highlights progress, opportunities for metabolic reconstruction. PLoS Comput. Biol. 2015, 11, e1004530. [Google Scholar] [CrossRef] [PubMed]
- Otero, J.M.; Cimini, D.; Patil, K.R.; Poulsen, S.G.; Olsson, L.; Nielsen, J. Industrial systems biology of Saccharomyces cerevisiae enables novel succinic acid cell factory. PLoS ONE 2013, 8, e54144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvas, M. Genome scale metabolic model reconstruction for the needs of industrial biotechnology. New Biotechnol. 2016, 33. [Google Scholar] [CrossRef]
- Becker, S.A.; Feist, A.M.; Mo, M.L.; Hannum, G.; Palsson, B.O.; Herrgard, M.J. Quantitative prediction of cellular metabolism with constraint-based models: The cobra toolbox. Nat. Protoc. 2007, 2, 727–738. [Google Scholar] [CrossRef] [PubMed]
- King, Z.A.; Lloyd, C.J.; Feist, A.M.; Palsson, B.O. Next-generation genome-scale models for metabolic engineering. Curr. Opin. Biotechnol. 2015, 35, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Famili, I.; Forster, J.; Nielson, J.; Palsson, B.O. Saccharomyces cerevisiae phenotypes can be predicted by using constraint-based analysis of a genome-scale reconstructed metabolic network. Proc. Natl. Acad. Sci. USA 2003, 100, 13134–13139. [Google Scholar] [CrossRef] [PubMed]
- Lewis, N.E.; Nagarajan, H.; Palsson, B.O. Constraining the metabolic genotype-phenotype relationship using a phylogeny of in silico methods. Nat. Rev. Microbiol. 2012, 10, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Thiele, I.; Palsson, B.O. A protocol for generating a high-quality genome-scale metabolic reconstruction. Nat. Protoc. 2010, 5, 93–121. [Google Scholar] [CrossRef] [PubMed]
- Mo, M.L.; Palsson, B.O.; Herrgard, M.J. Connecting extracellular metabolomic measurements to intracellular flux states in yeast. BMC Syst. Biol. 2009, 3, 17. [Google Scholar] [CrossRef] [PubMed]
- Cakir, T.; Efe, C.; Dikicioglu, D.; Hortacsu, A.; Kirdar, B.; Oliver, S.G. Flux balance analysis of a genome-scale yeast model constrained by exometabolomic data allows metabolic system identification of genetically different strains. Biotechnol. Prog. 2007, 23, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Beale, D.J.; Barratt, R.; Marlow, D.R.; Dunn, M.S.; Palombo, E.A.; Morrison, P.D.; Key, C. Application of metabolomics to understanding biofilms in water distribution systems: A pilot study. Biofouling 2013, 29, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Creek, D.J.; Nijagal, B.; Kim, D.H.; Rojas, F.; Matthews, K.R.; Barrett, M.P. Metabolomics guides rational development of a simplified cell culture medium for drug screening against trypanosoma brucei. Antimicrob. Agent. Chemother. 2013, 57, 2768–2779. [Google Scholar] [CrossRef] [PubMed]
- Courant, F.; Martzolff, A.; Rabin, G.; Antignac, J.P.; Le Bizec, B.; Giraudeau, P.; Tea, I.; Akoka, S.; Couzinet, A.; Cogne, G.; et al. How metabolomics can contribute to bio-processes: A proof of concept study for biomarkers discovery in the context of nitrogen-starved microalgae grown in photobioreactors. Metabolomics 2013, 9, 1286–1300. [Google Scholar] [CrossRef]
- Mohmad-Saberi, S.E.; Hashim, Y.Z.H.Y.; Mel, M.; Amid, A.; Ahmad-Raus, R.; Packeer-Mohamed, V. Metabolomics profiling of extracellular metabolites in CHO-K1 cells cultured in different types of growth media. Cytotechnology 2012, 65, 577–586. [Google Scholar] [CrossRef] [PubMed]
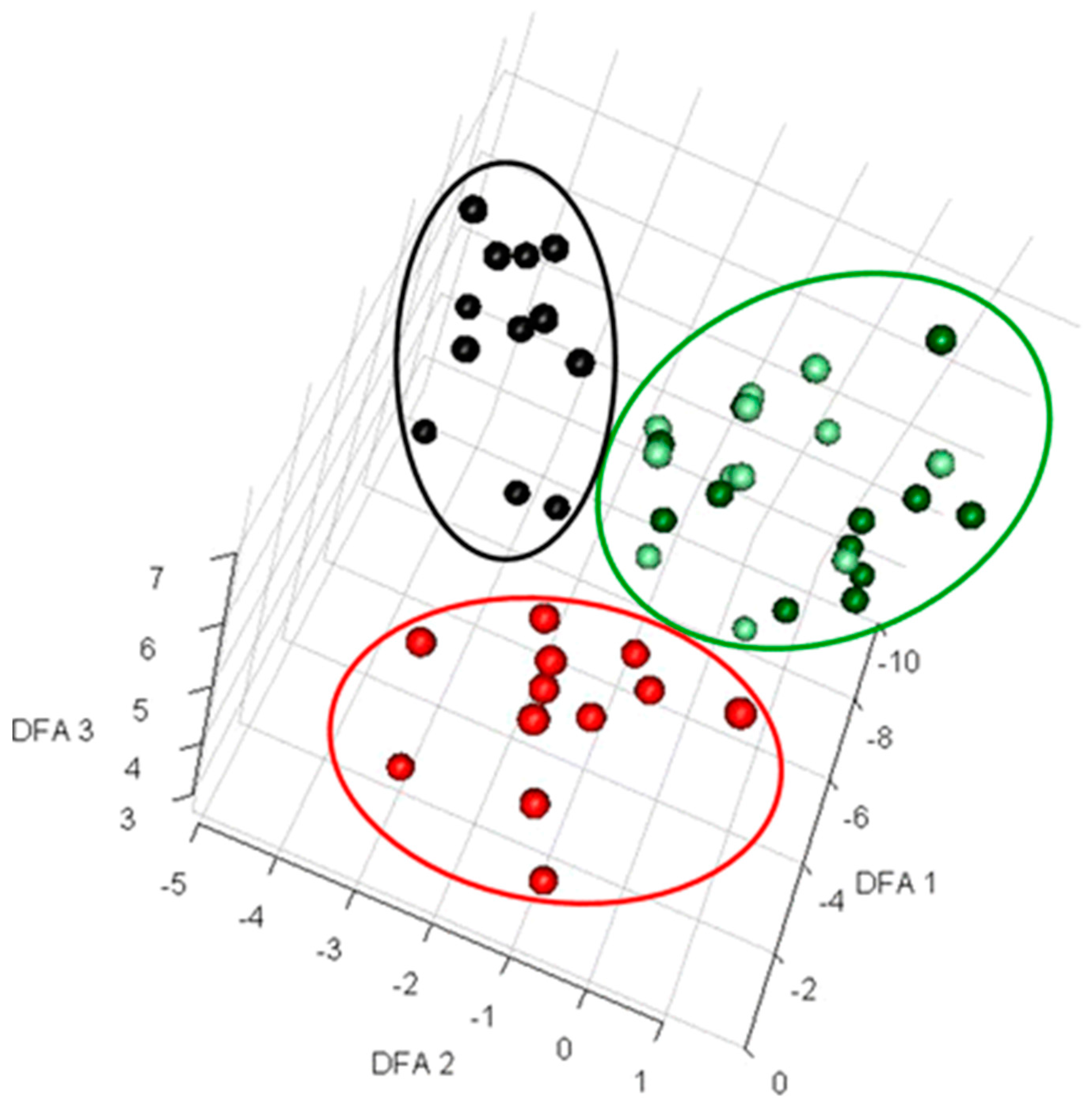
Figure 1.
Discriminant function analysis (DFA) to visualize exometabolomics data. Extracellular metabolites from deliberately contaminated and control samples from microalgal fermentation were analyzed using Gas-chromatography and mass spectrometry (GC-MS) [42]. The data from 56 samples were log transformed prior to performing DFA and three distinct clusters were observed. Here, black circles represent the samples from flasks contaminated with Pseudomonas aeruginosa, red circles show the samples contaminated by Bacillus subtilis, and light green and dark green circles present the samples collected from contaminated flasks at time 0 and non-contaminated flasks, respectively. This figure was reproduced from Sue et al. with the authors’ permission [42].
Figure 1.
Discriminant function analysis (DFA) to visualize exometabolomics data. Extracellular metabolites from deliberately contaminated and control samples from microalgal fermentation were analyzed using Gas-chromatography and mass spectrometry (GC-MS) [42]. The data from 56 samples were log transformed prior to performing DFA and three distinct clusters were observed. Here, black circles represent the samples from flasks contaminated with Pseudomonas aeruginosa, red circles show the samples contaminated by Bacillus subtilis, and light green and dark green circles present the samples collected from contaminated flasks at time 0 and non-contaminated flasks, respectively. This figure was reproduced from Sue et al. with the authors’ permission [42].
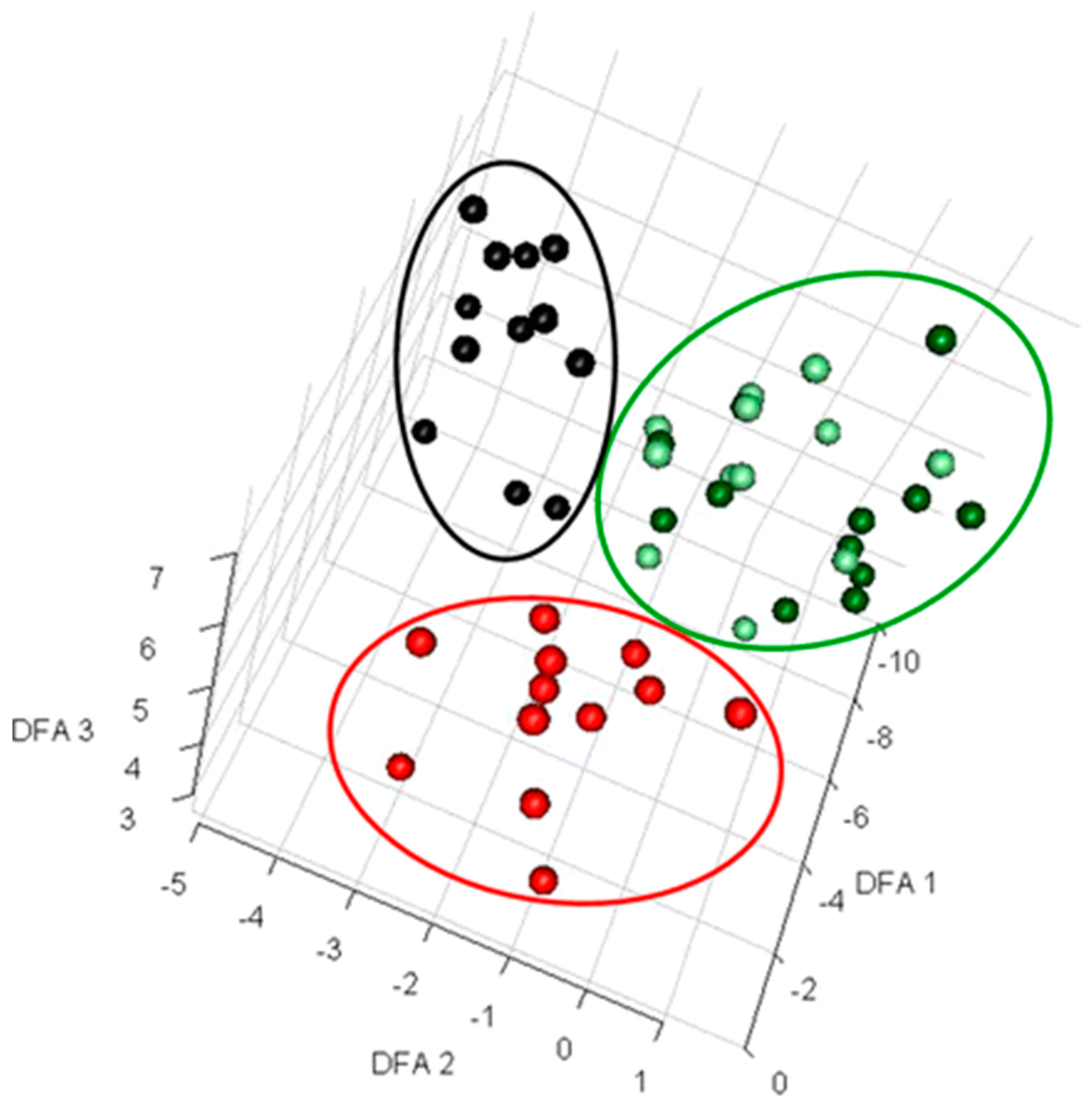
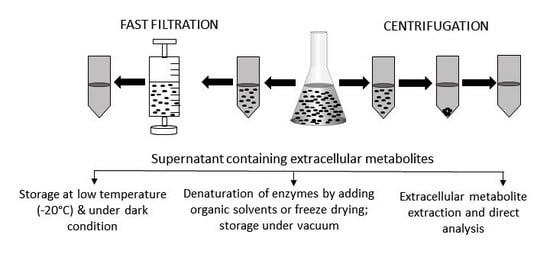
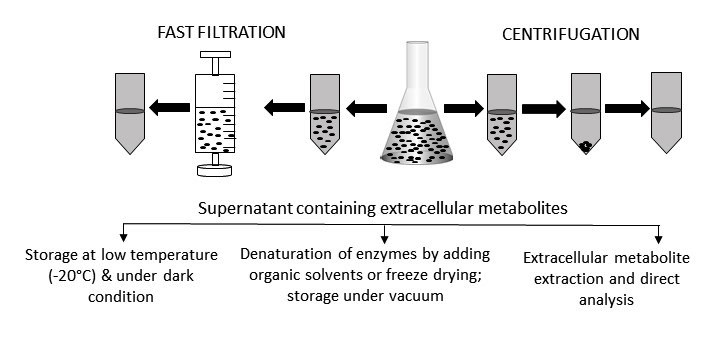
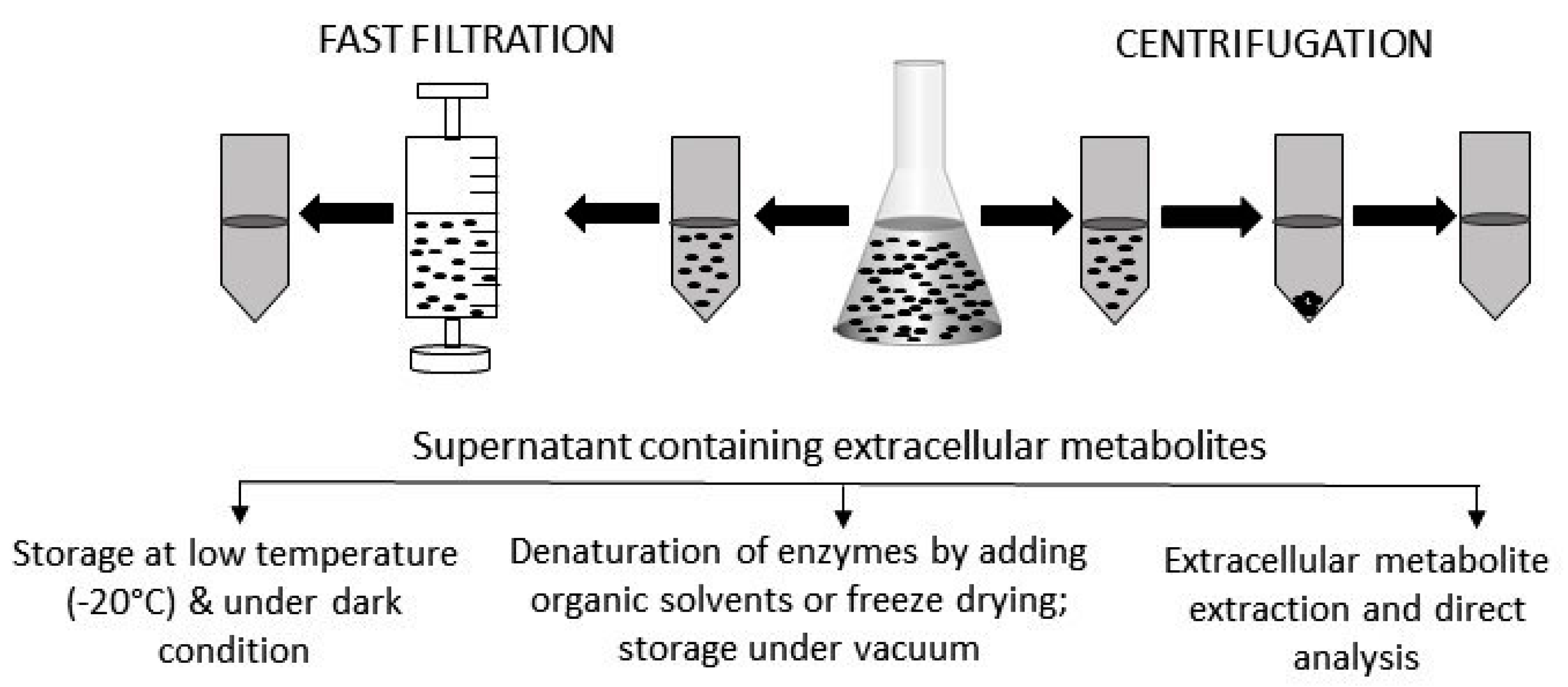
Figure 2.
Extracellular sample preparation, handling, and storage. After centrifugation or fast filtration, culture media containing extracellular metabolites are usually stored at a low temperature and under dark conditions. Sometime, organic solvents are added to denature active enzymes. For some metabolites, specific extraction procedures need to be followed before being analyzed by appropriate instruments. Prior to analysis, extracellular samples are often freeze-dried to concentrate the level of metabolites.
Figure 2.
Extracellular sample preparation, handling, and storage. After centrifugation or fast filtration, culture media containing extracellular metabolites are usually stored at a low temperature and under dark conditions. Sometime, organic solvents are added to denature active enzymes. For some metabolites, specific extraction procedures need to be followed before being analyzed by appropriate instruments. Prior to analysis, extracellular samples are often freeze-dried to concentrate the level of metabolites.
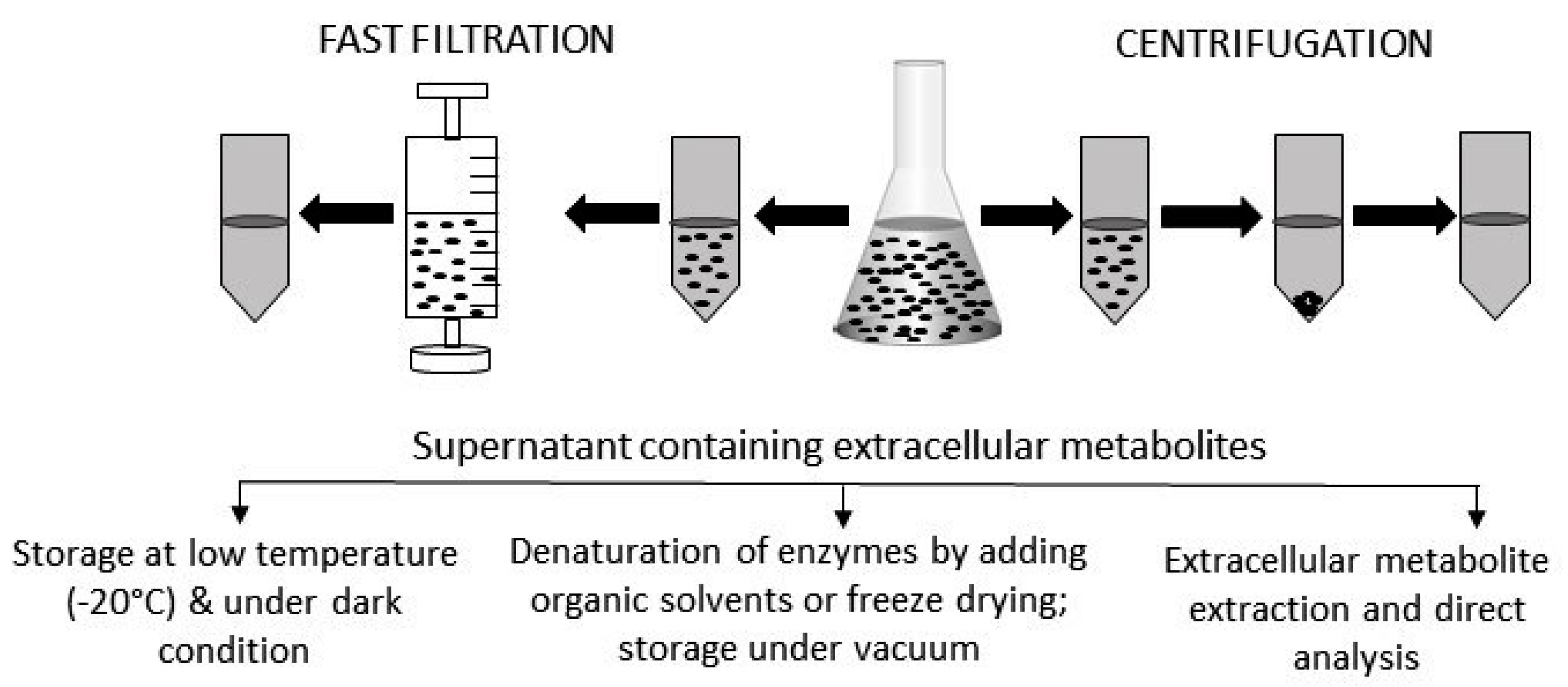
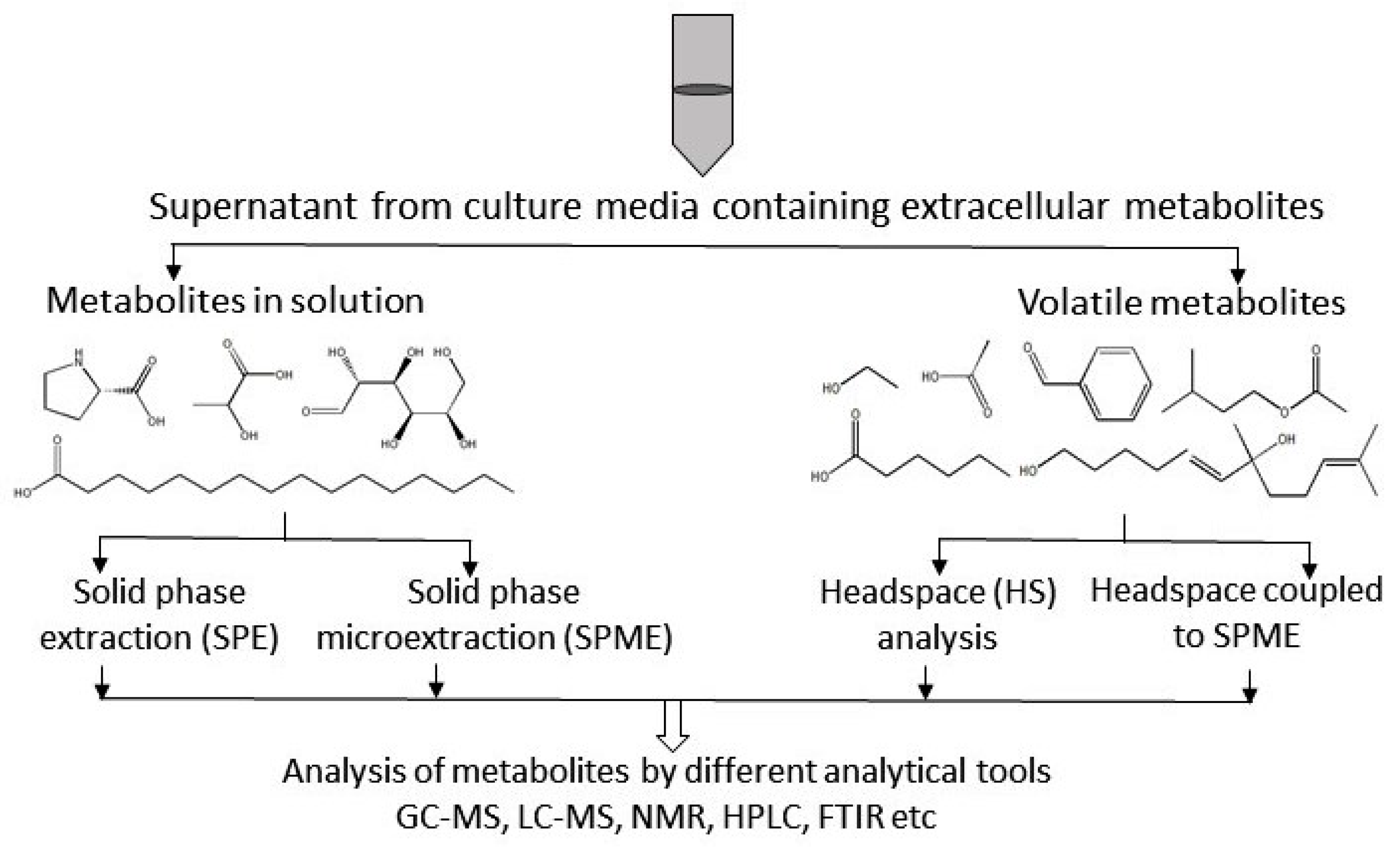
Figure 3.
Overview of extracellular metabolite analysis. Different technical methods, such as solid-phase extraction (SPE), solid-phase micro extraction (SPME), head space (HS) analysis, and HS-SPME, are used for the preparation of extracellular samples. Sample preparation protocols depend on the type of metabolite. Here, GC-MS = gas-chromatography coupled to mass spectrometry; LC-MS = liquid-chromatography coupled to mass spectrometry; NMR = nuclear magnetic resonance spectroscopy; HPLC = high pressure liquid chromatography; FTIR = Fourier transform infra-red spectroscopy.
Figure 3.
Overview of extracellular metabolite analysis. Different technical methods, such as solid-phase extraction (SPE), solid-phase micro extraction (SPME), head space (HS) analysis, and HS-SPME, are used for the preparation of extracellular samples. Sample preparation protocols depend on the type of metabolite. Here, GC-MS = gas-chromatography coupled to mass spectrometry; LC-MS = liquid-chromatography coupled to mass spectrometry; NMR = nuclear magnetic resonance spectroscopy; HPLC = high pressure liquid chromatography; FTIR = Fourier transform infra-red spectroscopy.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison among different solid-phase extraction phases.
| Phase | |||
|---|---|---|---|
| Parameters | Normal | Reversed | Ion-Exchange |
| Solvent polarity | High | Low | High |
| Range of solvent polarity | Low to medium | High to medium | High |
| Solvents for elution | Acetone, ethyl acetate | Water/methanol/acetonitrile solution | Salts and buffers |
| Loading solvents | Toluene, hexane | Water and buffers | Water and buffers |
| Eluted sample | Less polar | Most polar | Weakly ionized |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pinu, F.R.; Villas-Boas, S.G. Extracellular Microbial Metabolomics: The State of the Art. Metabolites 2017, 7, 43. https://doi.org/10.3390/metabo7030043
AMA Style
Pinu FR, Villas-Boas SG. Extracellular Microbial Metabolomics: The State of the Art. Metabolites. 2017; 7(3):43. https://doi.org/10.3390/metabo7030043
Chicago/Turabian StylePinu, Farhana R., and Silas G. Villas-Boas. 2017. "Extracellular Microbial Metabolomics: The State of the Art" Metabolites 7, no. 3: 43. https://doi.org/10.3390/metabo7030043
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.