Identification of an Epoxide Metabolite of Lycopene in Human Plasma Using 13C-Labeling and QTOF-MS
by
 , ,
, ,
Morgan J. Cichon
1,*, 
Nancy E. Moran
2,†,
Ken M. Riedl
1,2,
Steven J. Schwartz
1,2 and
Steven K. Clinton
2,3 1
Department of Food Science & Technology, The Ohio State University, Columbus, OH 43210, USA
2
The Ohio State Comprehensive Cancer Center, Columbus, OH 43210, USA
3
Division of Medical Oncology, Department of Internal Medicine, The Ohio State University Comprehensive Cancer Center, Columbus, OH 43210, USA
*
Author to whom correspondence should be addressed.
†
Current address: The USDA/ARS Children’s Nutrition Research Center, Department of Pediatrics, Baylor College of Medicine, Houston, TX 77030, USA.
Metabolites 2018, 8(1), 24; https://doi.org/10.3390/metabo8010024
Submission received: 31 January 2018
/
Revised: 3 March 2018
/
Accepted: 19 March 2018
/
Published: 20 March 2018
(This article belongs to the Special Issue Selected Papers from The Inaugural Conference on Food and Nutritional Metabolomics and 14th Annual Ohio Mass Spectrometry Symposium)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The carotenoid lycopene is a bioactive component of tomatoes and is hypothesized to reduce risk of several chronic diseases, such as prostate cancer. The metabolism of lycopene is only beginning to be understood and some studies suggest that metabolites of lycopene may be partially responsible for bioactivity associated with the parent compound. The detection and characterization of these compounds in vivo is an important step in understanding lycopene bioactivity. The metabolism of lycopene likely involves both chemical and enzymatic oxidation. While numerous lycopene metabolites have been proposed, few have actually been identified in vivo following lycopene intake. Here, LC-QTOF-MS was used along with 13C-labeling to investigate the post-prandial oxidative metabolism of lycopene in human plasma. Previously reported aldehyde cleavage products were not detected, but a lycopene 1,2-epoxide was identified as a new candidate oxidative metabolite.
1. Introduction
Epidemiological studies have demonstrated an association between the consumption of tomato products and the carotenoid lycopene and a decreased risk of prostate cancer [1,2], a relationship supported by experimental models [3,4]. Lycopene is the pigment responsible for the red color of tomatoes and is one of the predominant phytochemicals in the tomato [5]. Lycopene metabolism is believed to progress through chemical or enzymatic oxidative cleavage of the hydrocarbon chain. Provitamin A carotenoids are centrally and oxidatively cleaved to generate vitamin A metabolites that interact with specific retinoid receptors to impact cell biology, while eccentric cleavage is likely an initial step in degradation pathways [6]. The mammalian genes coding for central and eccentric carotenoid cleavage have been characterized, and it is proposed that oxidative cleavage metabolites of lycopene may be responsible for some of the biological effects associated with lycopene [7], perhaps acting as partial agonists or antagonists to receptors in the steroid receptor superfamily [3,8,9]. Potential oxidation products (apo-lycopenoids) have been identified in vitro and include aldehydes, ketones, and acids [10,11]. Our understanding of lycopene metabolism is far more limited than for provitamin A carotenoids, and only a few potential lycopene metabolites have been reported in vivo. Our group has previously observed several apo-lycopenals (apo-6′-, apo-8′-, apo-10′-, apo-12′, and apo-14′-lycopenal) in human plasma [12]. These compounds are also found in small quantities in lycopene-containing foods, including raw tomatoes, tomato sauce, and tomato juice. Thus, it is unclear whether they are absorbed from the diet, are products of in vivo chemical or enzymatic metabolism, or perhaps are the result of both mechanisms.
Isotopic labeling has been used as a strategy for investigating carotenoid absorption, metabolism, and excretion in vivo. 14C-labeling was used to study the biodistribution of lycopene in rodents [13,14] and to investigate the long-term bioavailability of lycopene in human plasma from a single oral dose [15]. Radiolabeling is a highly sensitive technique, but as 14C is radioactive, the amount of 14C-lycopene that can be safely administered is limited and cannot be used to assess carotenoid metabolism at concentrations found in the diet. For this reason, it is difficult to obtain concrete structural information for lycopene metabolites using 14C-labeling. Stable isotope labeling provides an alternative low-risk approach, where 13C-labeled carotenoids can be consumed at concentrations found in the diet. This strategy has been used to study β-carotene absorption and conversion to vitamin A [16,17], as well as lycopene [18] and phytoene [19] pharmacokinetics.
The objective of this study is to investigate the oxidative metabolism of lycopene in humans using a 13C-label. Plasma samples for this analysis were obtained from a recent study conducted by our group [18]. We expect labeling with a stable isotope will offer enhanced sensitivity to identify biological metabolites using liquid chromatography-quadrupole time-of-flight mass spectrometry (LC-QTOF-MS).
2. Results
2.1. Targeted Search for Apo-Lycopenals
Given our group’s previous identification of apo-lycopenals in human plasma, we expected to observe 13C-labeled aldehyde cleavage products of lycopene in the plasma of subjects from this study. The LC-MS method used for the plasma analysis was, therefore, optimized to detect the various apo-lycopenals expected (apo-6′-, apo-8′-, apo-10′-, apo-12′-, and apo-14′-lycopenal). This study provided a dose of 10.2 mg 13C-lycopene, which is physiologically relevant. However, masses corresponding to the 13C-labeled apo-lycopenals were not detected in the plasma of the eight subjects at the any of the time points examined. Additionally, native, unlabeled apo-lycopenals were also not detected.
2.2. Identification of New Lycopene Metabolites
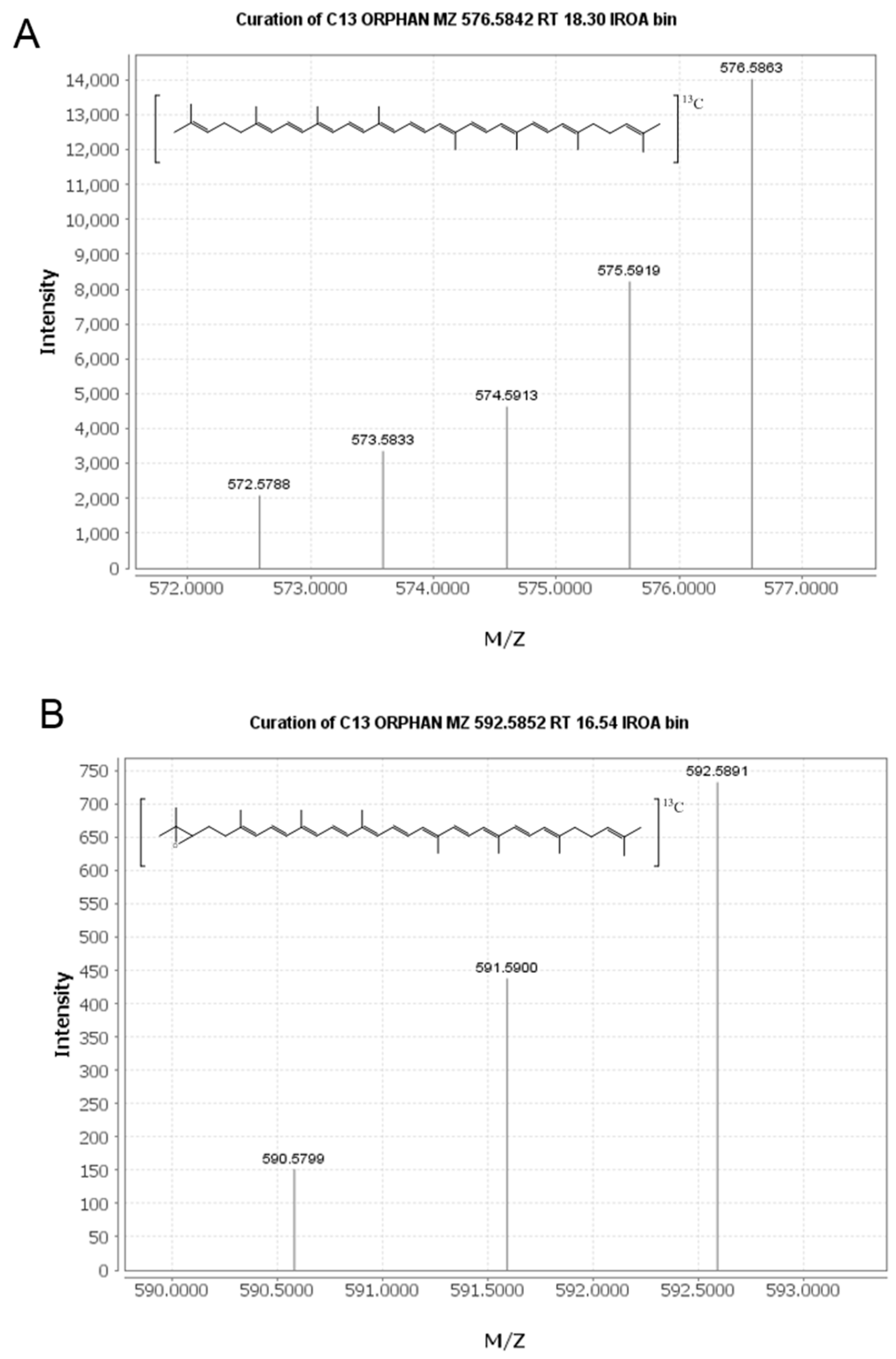
An untargeted approach was taken to mine the full-scan LC-MS data collected and identify new plasma metabolites of lycopene. The IROA ClusterFinder software (version 1, build 81) was used to search the data and extract mass spectral features with isotopic distributions characteristic of the 13C enrichment provided in the tracer dose. This approach was validated with the detection of 13C-lycopene in our plasma samples (Figure 1A). All features detected by the software were manually reviewed to eliminate false positives resulting from noise in the spectra. For a feature to be considered a labeled metabolite, it needed to be clearly present post-dose, but not at baseline.
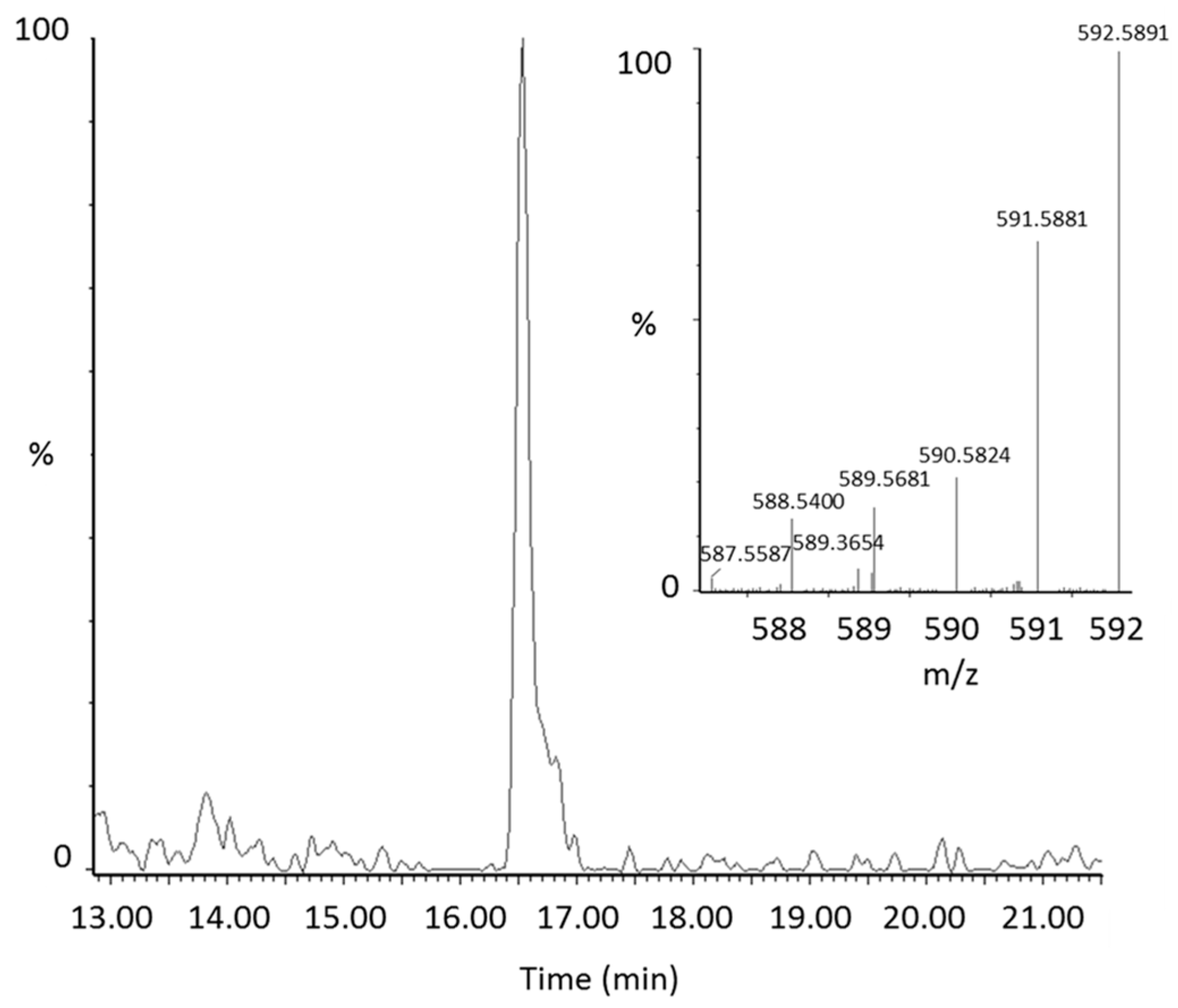
From manual review of the ClusterFinder data, we discovered another labeled compound besides 13C-lycopene that was present post-dose, but not at baseline and had the same characteristic reverse isotope distribution as the labeled lycopene (Figure 1B). With an m/z of 592.58, this compound was determined to be a 13C-labeled lycopene epoxide as the mass corresponds to the addition of oxygen to the parent molecule. The compound had a retention time of 16.5 min (Figure 2) and eluted in the same region of the chromatogram as all-trans-lycopene (18.2 min), 5-cis-lycopene (18.4 min), and other lycopene cis isomers (14.8–16.8 min), demonstrating structural similarity.
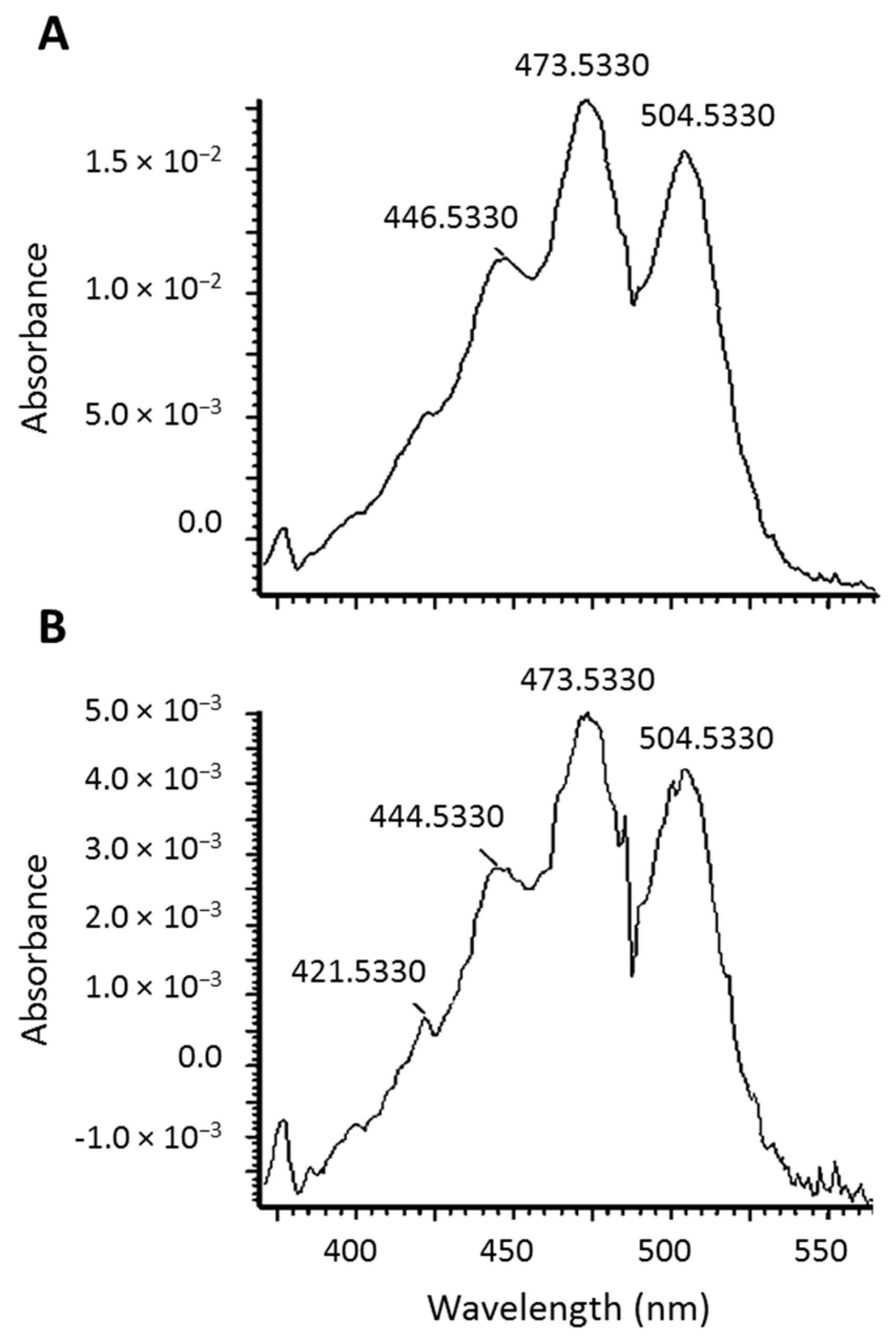
Lycopene mono- and di-epoxides have been reported in tomato products, along with NMR and UV-Vis confirmation [20]. Carotenoids have characteristic absorption spectra and the epoxide observed here was found to have a similar absorption spectrum to all-trans-lycopene (Figure 3). Therefore, we hypothesize that epoxidation is occurring at the 1,2 position (see structure in Figure 1B) where the chromophore and UV-Vis spectrum would be unaltered [21]. This is in agreement with the UV-Vis data reported previously for lycopene 1,2-epoxide [20]. Epoxidation of lycopene at the 5,6 position has also been reported, but this alteration causes a shortening of the chromophore and a hypsochromic shift of approximately 20 nm [22,23]. The terminal double bond on the lycopene structure is the only position where epoxidation would not alter the absorption spectrum.
2.3. Quantitation of 13C-lycopene 1,2-Epoxide in Plasma
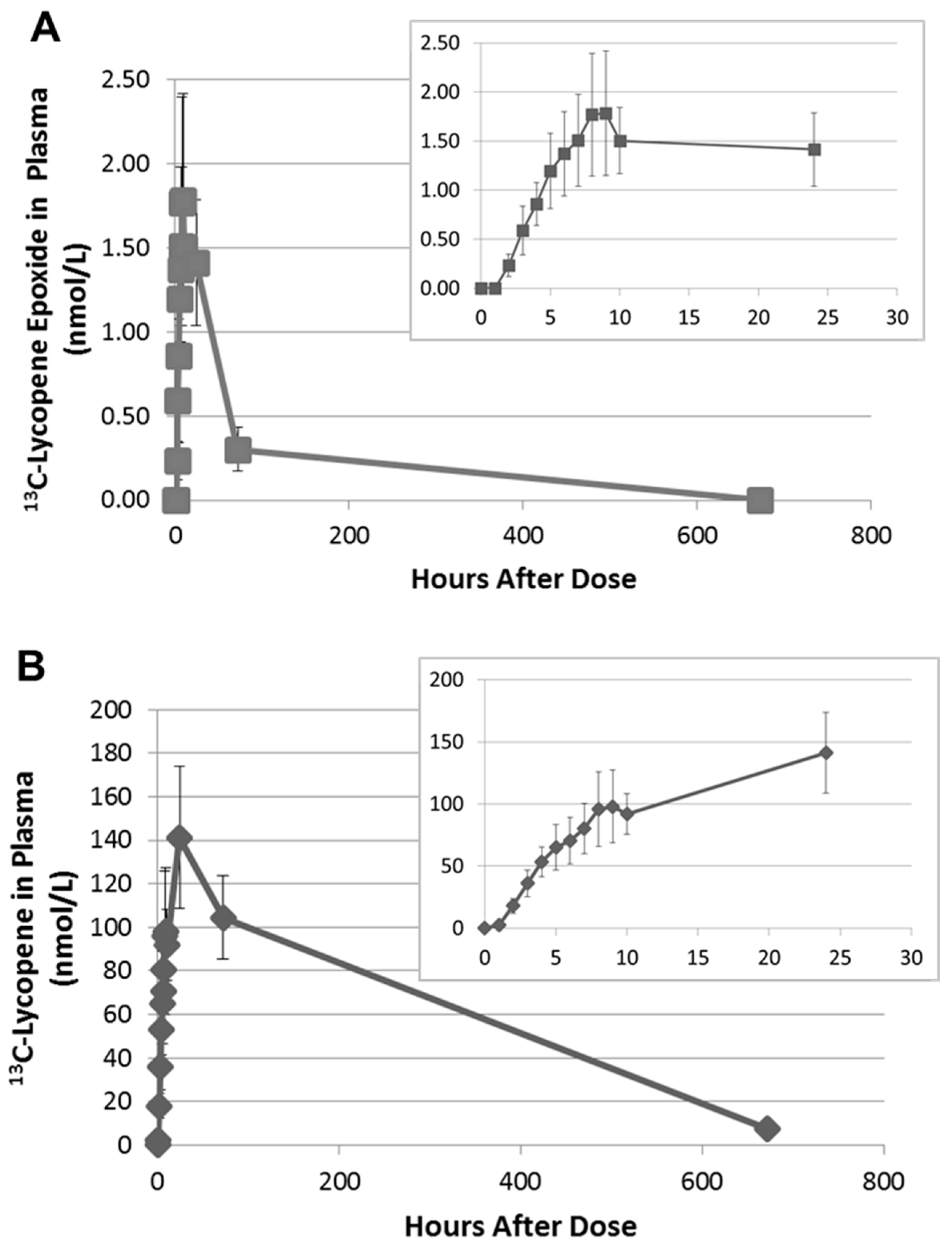
Upon further inspection of the data, the 13C-labeled lycopene 1,2-epoxide was found to be present in the plasma of all subjects and was first detected between 2 and 4 h. The PDA was used to calculate the relative MS response of the epoxide compared to lycopene. As the 1,2-epoxide and lycopene have the same chromophore and UV-Vis spectra, they will have equivalent extinction coefficients. The MS response factor was then used to quantitate the 13C-labeled lycopene 1,2-epoxide in plasma based on the external calibration curve for 13C-labeled lycopene. The 13C-lycopene 1,2-epoxide was found to increase linearly over the first 8 h after administration of the 13C-lycopene (Figure 4A). Most subjects reached maximum plasma levels between 8 and 9 h after dosing, with concentrations calculated to be 0.82–4.65 nmol/L. This corresponds to 1.71 ± 0.11% of the 13C-lycopene detected at those time points.
Interestingly, on average, there was little change in the 13C-epoxide in plasma between 10 and 24 h. However, by 72 h the 13C-epoxide had decreased by 70–100% of the maximum concentration and after 28 days was non-detectable in any of the subjects. This time course differs from the 13C-lycopene, which continued to increase significantly in plasma between 10 and 24 h and was still quantifiable after 28 days (Figure 4B).
A metabolite corresponding to the unlabeled form of the lycopene 1,2-epoxide was also detected at the same retention time in the plasma of all subjects. The concentrations are low (<1.0 nmol/L), but quantifiable, on the day of dosing (0–10 h) and increased significantly once subjects resumed their moderate lycopene diet (10–20 mg lycopene/d). At 28 days, the unlabeled lycopene 1,2-epoxide was found to be between 1.63 and 11.18 nmol/L in plasma. We have ruled out the lycopene epoxide being an artifact of sample handling based on the changing ratios of the native (unlabeled) epoxide to native lycopene in plasma over the course of the study. The native lycopene epoxide is present in plasma at 0.089 ± 0.027% the concentration of native circulating lycopene on the day of dosing (0–10 h) when unlabeled lycopene was not consumed. However, the native epoxide is present at 0.52 ± 0.16% and 0.50 ± 0.12% the concentration of native lycopene at 72 h and 28 days, respectively, when subjects returned to a moderate lycopene diet.
3. Discussion
In this study, we utilized stable isotope technology to detect and characterize the presence of lycopene epoxides in human biosamples. This effort extends the work of Khachik et al. (1997), reporting the presence of lycopene diols in human serum and hypothesizing that epoxidation is an initial step in the oxidative metabolism of lycopene, followed by enzymatic or acid hydrolysis to generate the diol forms [24,25]. They did not observe the epoxide but invoked it as a metabolic precursor to explain the presence of lycopene diols. Here, we did detect lycopene epoxides consistently in plasma samples from this study using a combination of accurate mass, UV-Vis, and chromatographic behavior, but were unable to detect lycopene diols. This could be an issue of ionization, limit of detection, or stability of these forms. Targeted LC-MS methods should be developed to confirm the presence of lycopene diols along with the epoxide in vivo.
Lycopene 1,2-epoxide and 5,6-epoxide are known derivatives of lycopene detected in tomatoes and processed tomato products [22,26,27,28,29]. Lycopene epoxides have also been chemically generated with m-chloroperbenzoic acid [20,28] and via autooxidation in low moisture and aqueous model systems [28]. Upon further investigation of our 13C-labeled lycopene dose, we found that one of the peaks in the LC chromatogram originally thought to be a minor cis-isomer of lycopene, was actually the 1,2 epoxide. The 13C-labeled lycopene 1,2-epoxide was present in the dose at approximately 1% the concentration of lycopene. Additionally, the plasma appearance curve of the 13C-labeled lycopene epoxide mimics that of the 13C-labeled lycopene over the first 10 h (Figure 4), suggesting that it may be similarly absorbed as 13C-labeled lycopene and cleared or metabolized more rapidly. However, alternative explanations may exist. For example, the 13C-labeled lycopene 1,2-epoxide in the dose may be very reactive and degrade quickly by hydrolysis during ingestion and digestion, as proposed for many epoxides [24,30]. In that case, the 1,2-epoxide in blood may be part of a dynamic process of epoxidation and diol formation in vivo. The presence of the unlabeled lycopene 1,2 epoxide in plasma the day of dosing, when native lycopene was not consumed, further points to continued metabolism of circulating lycopene through epoxidation. We have ruled out artifactual formation from sample handling as the ratio of the lycopene epoxide to lycopene changes over the course of the study and tracks with lycopene (native and labeled) intake. If the epoxide were being formed during the extraction, we would expect the ratio of the epoxide to lycopene to fluctuate randomly or remain constant.
Future studies with the pure compound are needed to determine whether the lycopene 1,2-epoxide is absorbed from the diet or an early oxidative metabolite of lycopene. Epoxycarotenoids are widely distributed in nature, but xanthophyll epoxides do not appear to be significantly absorbed by humans [31]. The presence of lycopene diols in plasma and the absence of the epoxide forms led Khachick et al. (1998) to hypothesize that the epoxides are not absorbed intact [25]. On the other hand, orally administered dietary and synthetic forms of 5,6-epoxy-β-carotene were found to be bioavailable in humans [32] and have been shown to be biologically active in inducing differentiation of NB4 leukemia cells in vitro [33]. Other epoxy lipids have also been reported to have important biological activity. For example, fatty acid monoepoxides have been reported to have anti-inflammatory properties [34], antinociceptive effects [35], protect cardiovascular function [36], and inhibit angiogenesis and tumorigenesis [37]. However, reactive epoxides may also lead to non-specific macromolecule covalent binding that is deleterious to cells. Therefore, regardless of whether the lycopene epoxide is absorbed or formed in vivo, it is present in human plasma at biologically relevant concentrations and may contribute to some of the bioactivity associated with lycopene and tomatoes.
In the current study, we were unable to detect 13C-labeled or unlabeled apo-lycopenals in plasma at any of the time points. Our group previously reported apo-lycopenals in the plasma of subjects who had been on a high tomato juice dietary intervention for 8 weeks [12]. Despite significant tomato consumption, the concentrations of these oxidation products were quite low in plasma (0.12–0.73 nmol/L). Apo-lycopenals are present in tomato juice, but not our 13C-labeled lycopene dose, and therefore, may only accumulate at circulating levels after chronic intake of tomato products. Alternatively, apo-lycopenals may be transient metabolites in vivo, making them challenging to detect from a single dose of 13C-lycopene.
4. Materials and Methods
4.1. Clinical Study Design
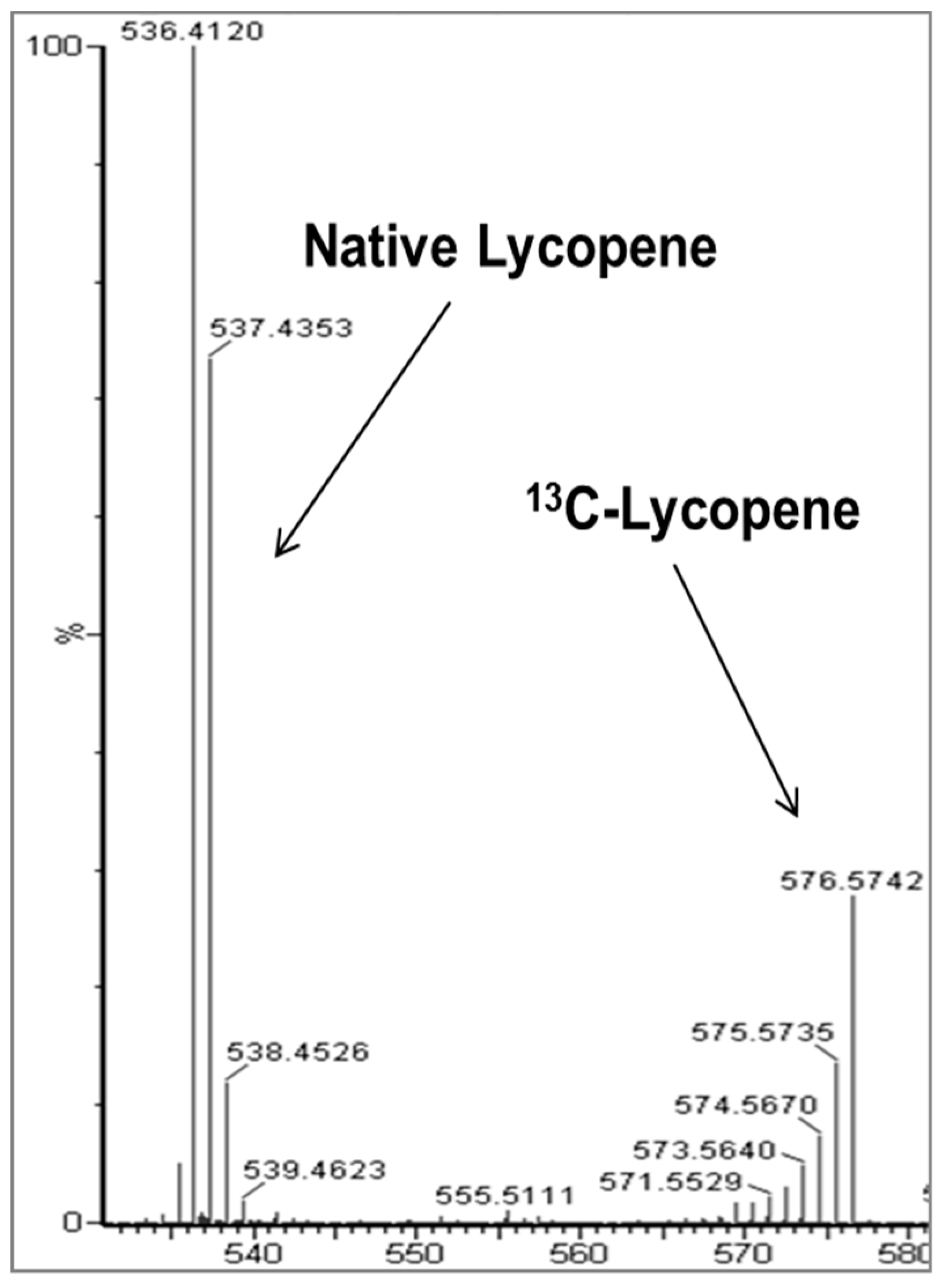
Full details of the 13C-lycopene dose and pharmacokinetic study, including MS chromatograms and spectra, have been published [18,38]. The 13C-lycopene was biosynthesized via tomato cell suspensions cultured with 13C-glucose and isolated for human consumption. The lycopene was purified using a crystallization method [38] and the spectroscopic purity was 99% as determined by UV-Vis and no native lycopene was detected in the dose by MS. The resulting 13C-lycopene dose was over 91% 13C atomic purity and approximately 30% uniformly labeled, giving a unique reverse isotope distribution when analyzed by MS (Figure 5). A total of 8 healthy adults (4 males and 4 females) were administered approximately 10.2 mg of 13C-labeled lycopene in olive oil. The oil was added to solubilize the lycopene, which is lipophilic, and because a number of studies have shown that carotenoid bioavailability is much improved with the co-consumption of lipid [39,40,41]. The study was conducted in compliance with the ethical standards of and was approved by The Ohio State University Institutional Review Board (#2009C0104), and written informed consent was obtained from all subjects. Blood was drawn hourly from 0–10 h after dosing and at 1, 3, and 28 days after dosing. Subjects abstained from lycopene containing foods the day of dosing and were instructed to consume a controlled lycopene diet two weeks prior to and four weeks following their visit. Our physiologic compartmental modeling of this study indicated that 23 ± 6% of the 13C-lycopene administered was absorbed from the dose [18].
4.2. Plasma Preparation and LC-MS Analysis
The sample preparation and HPLC-QTOF-MS analysis of the plasma were conducted as previously described [18]. Briefly, plasma was extracted using a mixture of hexane/ethanol/acetone/toluene (10:6:7:7) following protein precipitation with ethanol containing 0.1% butylated hydroxytoluene to prevent oxidation. Extracts were dried under nitrogen and reconstituted in methyl tert-butyl ether (MtBE)/methanol (1:1) prior to analysis. All samples were analyzed by HPLC-QTOF-MS the same day they were extracted to minimize potential degradation and oxidation of lycopene and its metabolites. A model 2695 HPLC with a 996 PDA (Waters Corp., Milford, MA, USA) was coupled to a quadrupole time-of-flight (Q-TOF) Premier hybrid mass spectrometer (Micromass UK Ltd., Manchester, UK) via an atmospheric pressure chemical ionization probe operated in negative ion mode. Compounds were separated on a YMC C30 column (3 μm; 4.6 × 250 mm) (Waters Corp.) with a mobile phase gradient consisting of solvent mixtures A (60% methanol, 35% MtBE, and 5% water) and B (78% MtBE, 20% methanol, and 2% water). MS parameters were as follows: corona current, 30 µA; collision energy, 8 eV; cone voltage, 40 V; source block temperature, 110 °C; probe temperature, 600 °C; and desolvation gas flow, 400 L·h−1. All MS experiments were run in Enhanced Duty Cycle mode for increased sensitivity with V-optics enabled (7500 mass resolution). MS data were acquired using MassLynx software version 4.1 (Waters Corp.).
4.3. Data Mining
The HPLC-QTOF-MS plasma data acquired from this study were mined for new 13C-labeled metabolites of lycopene using an untargeted approach. Waters format data files (.raw) were converted to an open data format (.mzXML) using ProteoWizard (version 3.0.10875) [42]. A subset of the data files were then analyzed using Isotope Ratio Outlier Analysis (IROA) ClusterFinder software (IROA Technologies, Bolton, MA, USA), which is a feature finding program designed to search for 13C isotope patterns within raw mass spectral data. The resulting features were manually assessed and potential 13C-labeled metabolites were evaluated in the full dataset.
5. Conclusions
We have demonstrated that stable isotope labeling is an effective strategy for studying phytochemical metabolites in humans. A lycopene epoxide has been observed in human plasma at biologically relevant concentrations for the first time. As other lipid epoxides have been reported to have biological activity, the lycopene epoxide may contribute to the biological properties associated with lycopene and tomatoes. Follow-up experiments are needed to determine the source of the lycopene epoxide in plasma and to investigate the bioactivity of this oxidation product.
Acknowledgments
We thank Chris Beecher, (IROA Technologies) for providing the ClusterFinder software and assisting in its use. The analytical portion of this study was performed within the Nutrient and Phytochemical Analytics Shared Resource under The Ohio State University Comprehensive Cancer Center (NIH P30 CA016058). This work was supported by the National Center for Complementary and Integrative Health (NCCIH) (5R21AT005166 to S.K.C.), NCCIH and the Office of Dietary Supplements (ODS) (K99/R00 AT008576 to N.E.M.), The USDA Agricultural Research Service (CRIS 3092-51000-056-03S to N.E.M.), and the Lisa and Dan Wampler Endowed Fellowship for Foods and Health Research (M.J.C.). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health or the United States Department of Agriculture.
Author Contributions
S.K.C., N.E.M. and M.J.C. conducted the study; M.J.C. and K.M.R. performed the mass spectrometry analysis and data interpretation; S.J.S and S.K.C. supervised the experimentation, and all authors have contributed to and approved the final version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Key, T.J.; Appleby, P.N.; Travis, R.C.; Albanes, D.; Alberg, A.J.; Barricarte, A.; Black, A.; Boeing, H.; Bueno-De-Mesquita, H.B.; Chan, J.M.; et al. Carotenoids, retinol, tocopherols, and prostate cancer risk: Pooled analysis of 15 studies. Am. J. Clin. Nutr. 2015, 102, 1142–1157. [Google Scholar] [CrossRef] [PubMed]
- Rowles, J.L.; Ranard, K.M.; Applegate, C.C.; Jeon, S.; An, R.; Erdman, J.W. Processed and raw tomato consumption and risk of prostate cancer: A systematic review and dose-response meta-analysis. Prostate Cancer Prostatic Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.-L.; Thomas-Ahner, J.M.; Moran, N.E.; Cooperstone, J.L.; Erdman, J.W.; Young, G.S.; Clinton, S.K. β-carotene 9′,10′ oxygenase modulates the anticancer activity of dietary tomato or lycopene on prostate carcinogenesis in the TRAMP model. Cancer Prev. Res. 2017, 10, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Boileau, T.W.-M.; Liao, Z.; Kim, S.; Lemeshow, S.; Erdman, J.W.; Clinton, S.K. Prostate carcinogenesis in N-methyl-N-nitrosourea (NMU)-testosterone-treated rats fed tomato powder, lycopene, or energy-restricted diets. J. Natl. Cancer Inst. 2003, 95, 1578–1586. [Google Scholar] [CrossRef] [PubMed]
- Clinton, S.K. Lycopene: Chemistry, biology, and implications for human health and disease. Nutr. Rev. 1998, 56, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, A.; Harrison, E.H. Carotenoid metabolism in mammals, including man: Formation, occurrence, and function of apocarotenoids. J. Lipid Res. 2013, 54, 1719–1730. [Google Scholar] [CrossRef] [PubMed]
- Lindshield, B.L.; Canene-Adams, K.; Erdman, J.W. Lycopenoids: Are lycopene metabolites bioactive? Arch. Biochem. Biophys. 2007, 458, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, A.; Hruszkewycz, D.P.; dela Sena, C.; Narayanasamy, S.; Riedl, K.M.; Kopec, R.E.; Schwartz, S.J.; Curley, R.W.; Harrison, E.H. Naturally occurring eccentric cleavage products of provitamin A β-carotene function as antagonists of retinoic acid receptors. J. Biol. Chem. 2012, 287, 15886–15895. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, A.; Hruszkewycz, D.P.; Curley, R.W.; Harrison, E.H. The eccentric cleavage product of β-carotene, β-apo-13-carotenone, functions as an antagonist of RXRα. Arch. Biochem. Biophys. 2010, 504, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Caris-Veyrat, C.; Schmid, A.; Carail, M.; Böhm, V. Cleavage products of lycopene produced by in vitro oxidations: Characterization and mechanisms of formation. J. Agric. Food Chem. 2003, 51, 7318–7325. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Nara, E.; Kobayashi, H.; Terao, J.; Nagao, A. Formation of cleavage products by autoxidation of lycopene. Lipids 2001, 36, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Kopec, R.E.; Riedl, K.M.; Harrison, E.H.; Curley, R.W.; Hruszkewycz, D.P.; Clinton, S.K.; Schwartz, S.J. Identification and quantification of apo-lycopenals in fruits, vegetables, and human plasma. J. Agric. Food Chem. 2010, 58, 3290–3296. [Google Scholar] [CrossRef] [PubMed]
- Zaripheh, S.; Boileau, T.W.-M.; Lila, M.A.; Erdman, J.W. [14C]-lycopene and [14C]-labeled polar products are differentially distributed in tissues of F344 rats prefed lycopene. J. Nutr. 2003, 133, 4189–4195. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.E.; Clinton, S.K.; Erdman, J.W. Differential bioavailability, clearance, and tissue distribution of the acyclic tomato carotenoids lycopene and phytoene in mongolian gerbils. J. Nutr. 2013, 143, 1920–1926. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.B.; Vuong, L.T.; Ruckle, J.; Synal, H.A.; Schulze-Konig, T.; Wertz, K.; Rumbeli, R.; Liberman, R.G.; Skipper, P.L.; Tannenbaum, S.R.; et al. Lycopene bioavailability and metabolism in humans: An accelerator mass spectrometry study. Am. J. Clin. Nutr. 2011, 93, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Fleshman, M.K.; Riedl, K.M.; Novotny, J.A.; Schwartz, S.J.; Harrison, E.H. An LC/MS method for d8-β-carotene and d4-retinyl esters: β-carotene absorption and its conversion to vitamin A in humans. J. Lipid Res. 2012, 53, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Kurilich, A.C.; Britz, S.J.; Clevidence, B.A.; Novotny, J.A. Isotopic labeling and LC-APCI-MS quantification for investigating absorption of carotenoids and phylloquinone from kale (Brassica oleracea). J. Agric. Food Chem. 2003, 51, 4877–4883. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.E.; Cichon, M.J.; Riedl, K.M.; Grainger, E.M.; Schwartz, S.J.; Novotny, J.A.; Erdman, J.W.; Clinton, S.K. Compartmental and noncompartmental modeling of 13C-lycopene absorption, isomerization, and distribution kinetics in healthy adults. Am. J. Clin. Nutr. 2015, 102, 1436–1449. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.E.; Novotny, J.A.; Cichon, M.J.; Riedl, K.M.; Rogers, R.B.; Grainger, E.M.; Schwartz, S.J.; Erdman, J.W.; Clinton, S.K. Absorption and distribution kinetics of the 13C-labeled tomato carotenoid phytoene in healthy adults. J. Nutr. 2016, 146, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Khachik, F.; Steck, A.; Niggli, U.A.; Pfander, H. Partial synthesis and structural elucidation of the oxidative metabolites of lycopene identified in tomato paste, tomato juice, and human serum. J. Agric. Food Chem. 1998, 46, 4874–4884. [Google Scholar] [CrossRef]
- Mercadante, A.Z.; Egeland, E.S. Carotenoids Handbook; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhauser Verlag: Basel, Switzerland, 2004. [Google Scholar]
- Khachik, F.; Goli, M.B.; Beecher, G.R.; Holden, J.; Lusby, W.R.; Tenorio, M.D.; Barrera, M.R. Effect of food preparation on qualitative and quantitative distribution of major carotenoid constituents of tomatoes and several green vegetables. J. Agric. Food Chem. 1992, 40, 390–398. [Google Scholar] [CrossRef]
- Eugster, C.H. Chemical Derivatization: Microscale Tests for the Presence of Common Functional Groups in Carotenoids. In Carotenoids Volume 1A: Isolation and Analysis; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhauser Verlag: Basel, Switzerland, 1995; pp. 71–80. [Google Scholar]
- Khachik, F.; Spangler, C.J.; Smith, J.C.; Canfield, L.M.; Steck, A.; Pfander, H. Identification, quantification, and relative concentrations of carotenoids and their metabolites in human milk and serum. Anal. Chem. 1997, 69, 1873–1881. [Google Scholar] [CrossRef] [PubMed]
- Khachik, F.; Pfander, H.; Traber, B. Proposed mechanisms for the formation of synthetic and naturally occurring metabolites of lycopene in tomato products and human serum. J. Agric. Food Chem. 1998, 46, 4885–4890. [Google Scholar] [CrossRef]
- Britton, G.; Goodwin, T.W. Carotene epoxides from the Delta tomato mutant. Phytochemistry 1975, 14, 2530–2532. [Google Scholar] [CrossRef]
- Tonucci, L.H.; Holden, J.M.; Beecher, G.R.; Khachik, F.; Davis, C.S.; Mulokozi, G. Carotenoid content of thermally processed tomato-based food products. J. Agric. Food Chem. 1995, 43, 579–586. [Google Scholar] [CrossRef]
- Rodriguez, E.B.; Rodriguez-Amaya, D.B. Lycopene epoxides and apo-lycopenals formed by chemical reactions and autoxidation in model systems and processed foods. J. Food Sci. 2009, 74, C674–C682. [Google Scholar] [CrossRef] [PubMed]
- Cichon, M.J.; Riedl, K.M.; Schwartz, S.J. A metabolomic evaluation of the phytochemical composition of tomato juices being used in human clinical trials. Food Chem. 2017, 228, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.W.; Morisseau, C.; Hammock, B.D. Epoxide hydrolases: Their roles and interactions with lipid metabolism. Prog. Lipid Res. 2005, 44, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Barua, A.B.; Olson, J.A. Xanthophyll epoxides, unlike beta-carotene monoepoxides, are not detectibly absorbed by humans. J. Nutr. 2001, 131, 3212–3215. [Google Scholar] [CrossRef] [PubMed]
- Barua, A.B. Intestinal absorption of epoxy-beta-carotenes by humans. Biochem. J. 1999, 339, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Duitsman, P.K.; Barua, A.B.; Becker, B.; Olson, J.A. Effects of epoxycarotenoids, beta-carotene, and retinoic acid on the differentiation and viability of the leukemia cell line NB4 in vitro. Int. J. Vitam. Nutr. Res. 1999, 69, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Node, K.; Huo, Y.; Ruan, X.; Yang, B.; Spiecker, M.; Ley, K.; Zeldin, D.C.; Liao, J.K. Anti-inflammatory properties of cytochrome P450 epoxygenase-derived eicosanoids. Science 1999, 285, 1276–1279. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.; Vito, S.; Inceoglu, B.; Hammock, B.D. The role of long chain fatty acids and their epoxide metabolites in nociceptive signaling. Prostaglandins Other Lipid Mediat. 2014, 113–115, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, K.M.; Yang, W.; Gross, G.J.; Campbell, W.B. Roles of epoxyeicosatrienoic acids in vascular regulation and cardiac preconditioning. J. Cardiovasc. Pharmacol. 2007, 50, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Panigrahy, D.; Mahakian, L.M.; Yang, J.; Liu, J.-Y.; Stephen Lee, K.S.; Wettersten, H.I.; Ulu, A.; Hu, X.; Tam, S.; et al. Epoxy metabolites of docosahexaenoic acid (DHA) inhibit angiogenesis, tumor growth, and metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 6530–6535. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.E.; Rogers, R.B.; Lu, C.-H.; Conlon, L.E.; Lila, M.A.; Clinton, S.K.; Erdman, J.W. Biosynthesis of highly enriched 13C-lycopene for human metabolic studies using repeated batch tomato cell culturing with 13C-glucose. Food Chem. 2013, 139, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Goltz, S.R.; Campbell, W.W.; Chitchumroonchokchai, C.; Failla, M.L.; Ferruzzi, M.G. Meal triacylglycerol profile modulates postprandial absorption of carotenoids in humans. Mol. Nutr. Food Res. 2012, 56, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Unlu, N.Z.; Bohn, T.; Clinton, S.K.; Schwartz, S.J. Carotenoid absorption from salad and salsa by humans is enhanced by the addition of avocado or avocado oil. J. Nutr. 2005, 135, 431–436. [Google Scholar] [CrossRef] [PubMed]
- White, W.S.; Zhou, Y.; Crane, A.; Dixon, P.; Quadt, F.; Flendrig, L.M. Modeling the dose effects of soybean oil in salad dressing on carotenoid and fat-soluble vitamin bioavailability in salad vegetables. Am. J. Clin. Nutr. 2017, 106, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mass spectra of the 13C-labeled lycopene (A) and lycopene epoxide (B) detected with the IROA ClusterFinder software.
Figure 1.
Mass spectra of the 13C-labeled lycopene (A) and lycopene epoxide (B) detected with the IROA ClusterFinder software.

Figure 2.
Representative extracted ion chromatogram of the lycopene epoxide (m/z 592.58) in plasma with the corresponding mass spectrum of the peak.
Figure 2.
Representative extracted ion chromatogram of the lycopene epoxide (m/z 592.58) in plasma with the corresponding mass spectrum of the peak.
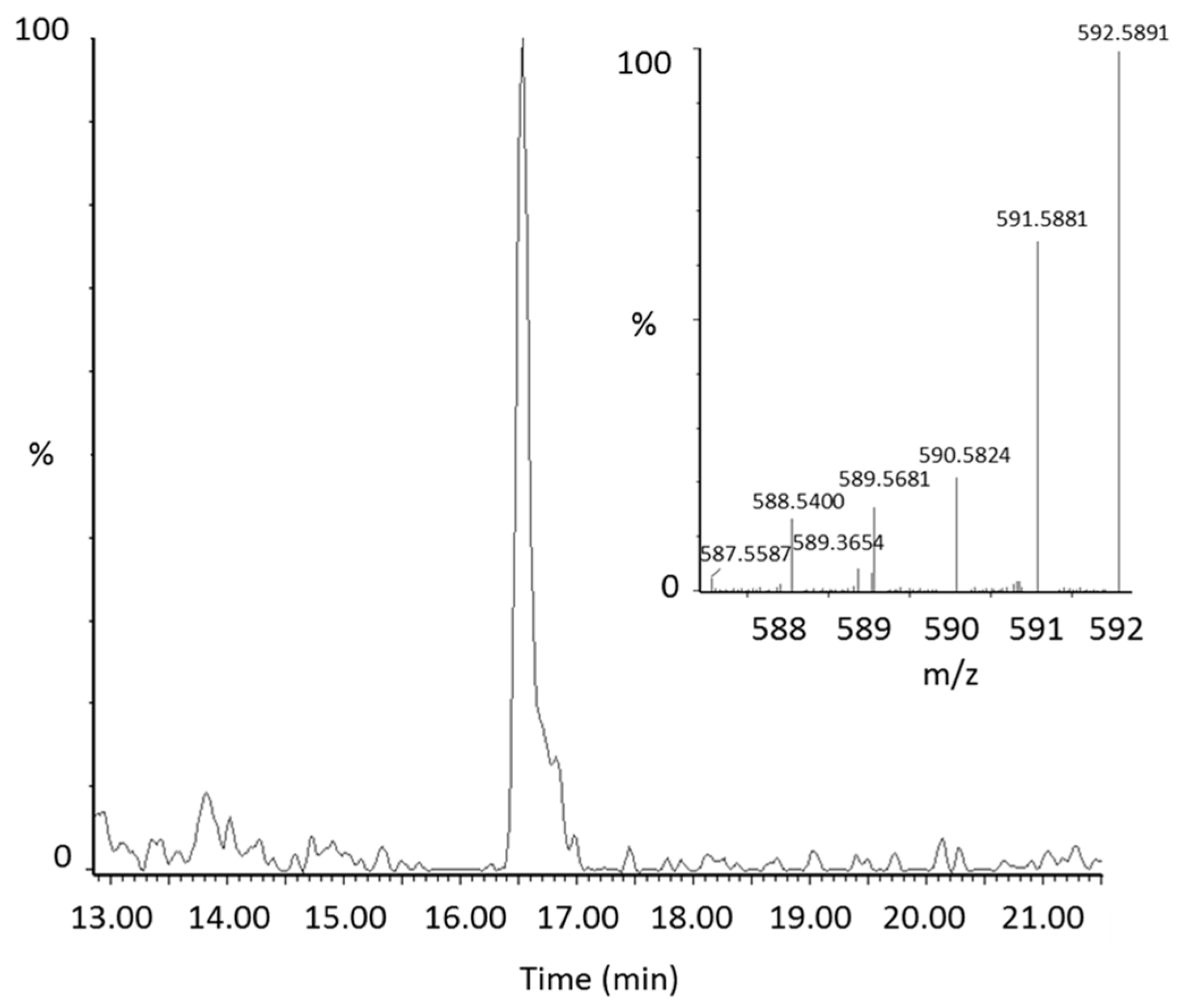
Figure 3.
UV-Vis spectra of all-trans-lycopene (A) and the lycopene epoxide (B).
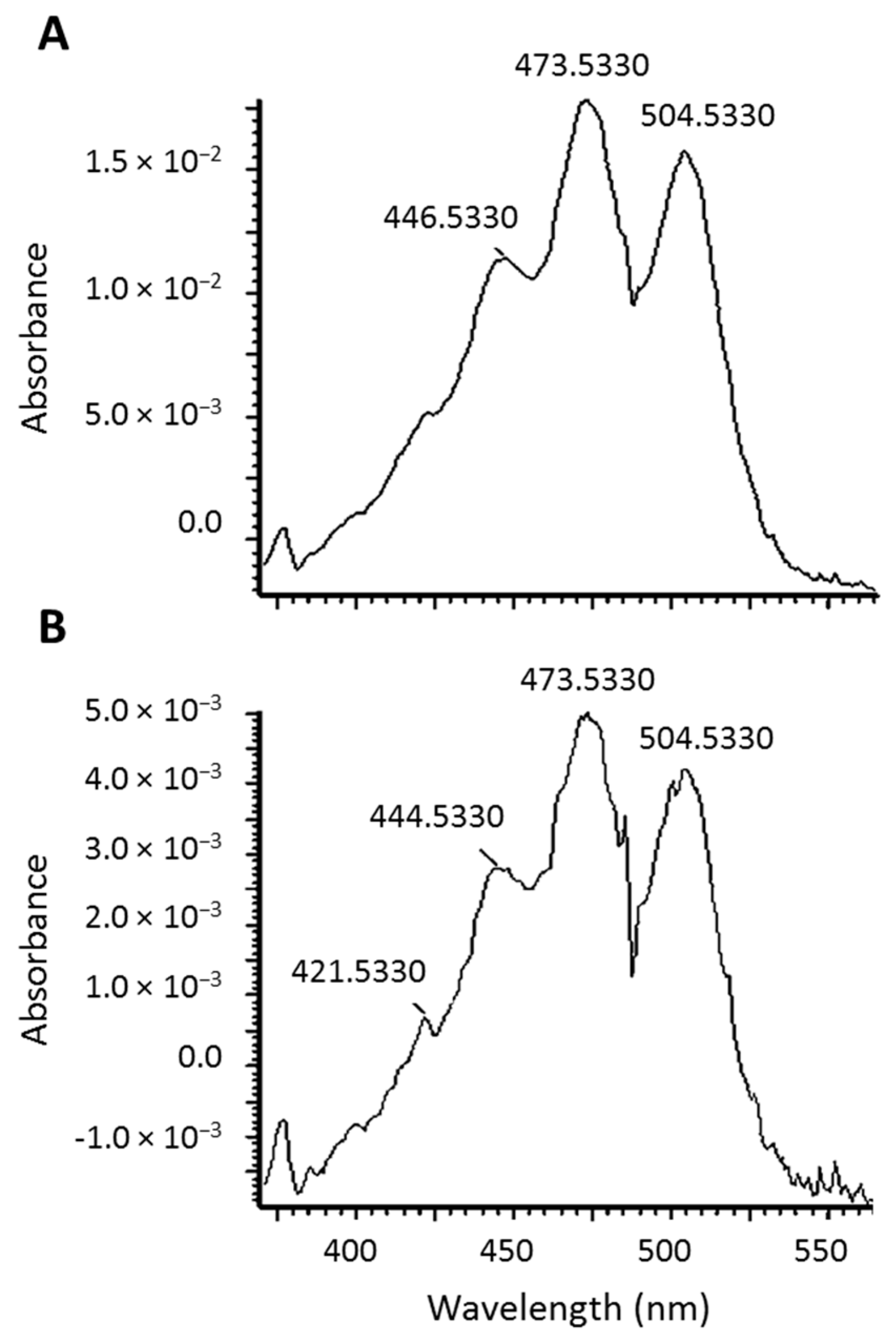
Figure 4.
Average appearance of the 13C-lycopene epoxide metabolite (A) and the 13C-lycopene parent (B) in the plasma of subjects (±SEM) with insets zoomed to the first 24 h. (13C-lycopene data have been previously published [18] and are visualized here for comparison purposes.).
Figure 4.
Average appearance of the 13C-lycopene epoxide metabolite (A) and the 13C-lycopene parent (B) in the plasma of subjects (±SEM) with insets zoomed to the first 24 h. (13C-lycopene data have been previously published [18] and are visualized here for comparison purposes.).
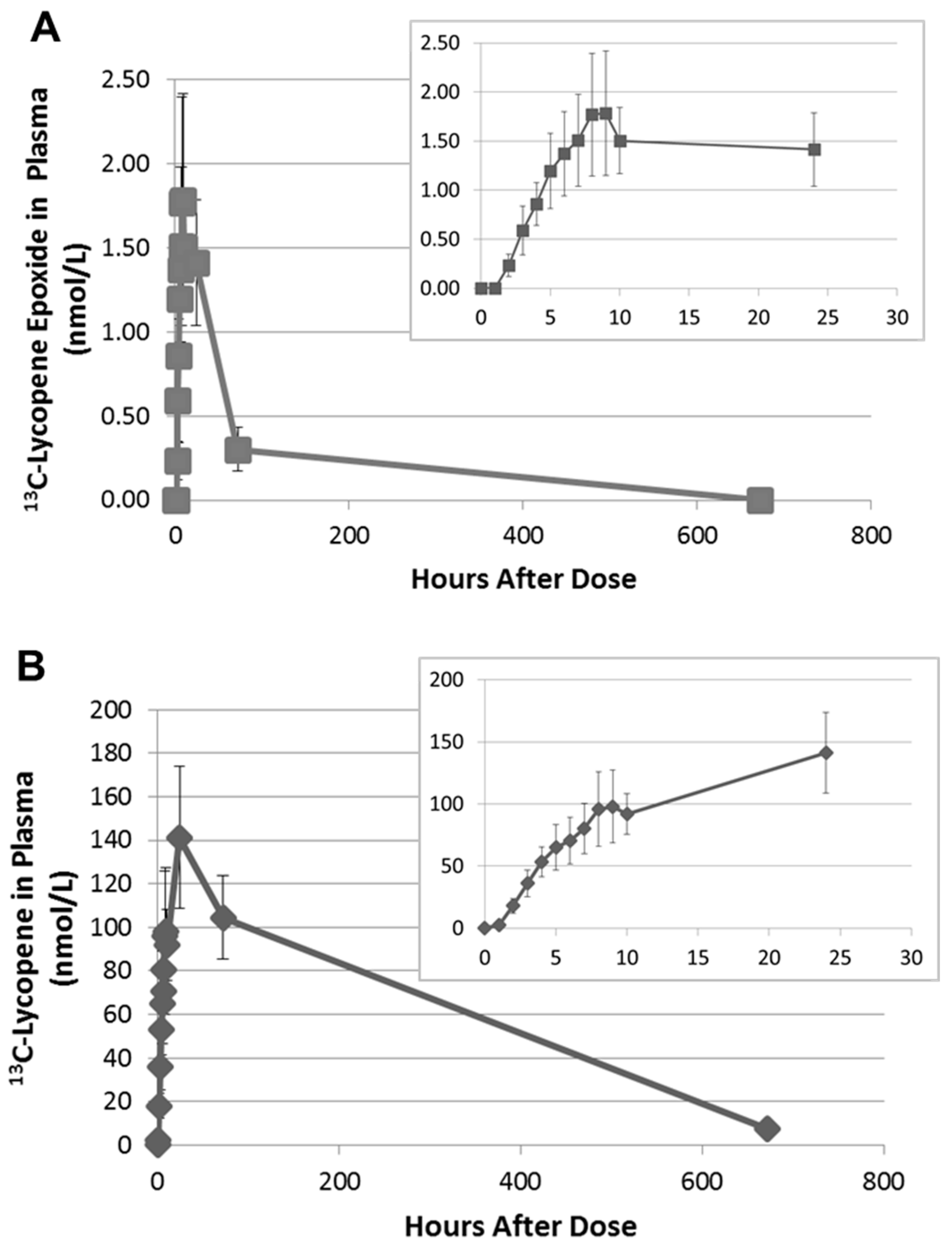
Figure 5.
Mass spectrum comparing the isotope distributions for native (unlabeled) lycopene and the 13C-labeled lycopene in a representative plasma sample.
Figure 5.
Mass spectrum comparing the isotope distributions for native (unlabeled) lycopene and the 13C-labeled lycopene in a representative plasma sample.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cichon, M.J.; Moran, N.E.; Riedl, K.M.; Schwartz, S.J.; Clinton, S.K. Identification of an Epoxide Metabolite of Lycopene in Human Plasma Using 13C-Labeling and QTOF-MS. Metabolites 2018, 8, 24. https://doi.org/10.3390/metabo8010024
AMA Style
Cichon MJ, Moran NE, Riedl KM, Schwartz SJ, Clinton SK. Identification of an Epoxide Metabolite of Lycopene in Human Plasma Using 13C-Labeling and QTOF-MS. Metabolites. 2018; 8(1):24. https://doi.org/10.3390/metabo8010024
Chicago/Turabian StyleCichon, Morgan J., Nancy E. Moran, Ken M. Riedl, Steven J. Schwartz, and Steven K. Clinton. 2018. "Identification of an Epoxide Metabolite of Lycopene in Human Plasma Using 13C-Labeling and QTOF-MS" Metabolites 8, no. 1: 24. https://doi.org/10.3390/metabo8010024
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.