Impact of Prolonged Blood Incubation and Extended Serum Storage at Room Temperature on the Human Serum Metabolome

Abstract
:1. Introduction
2. Results
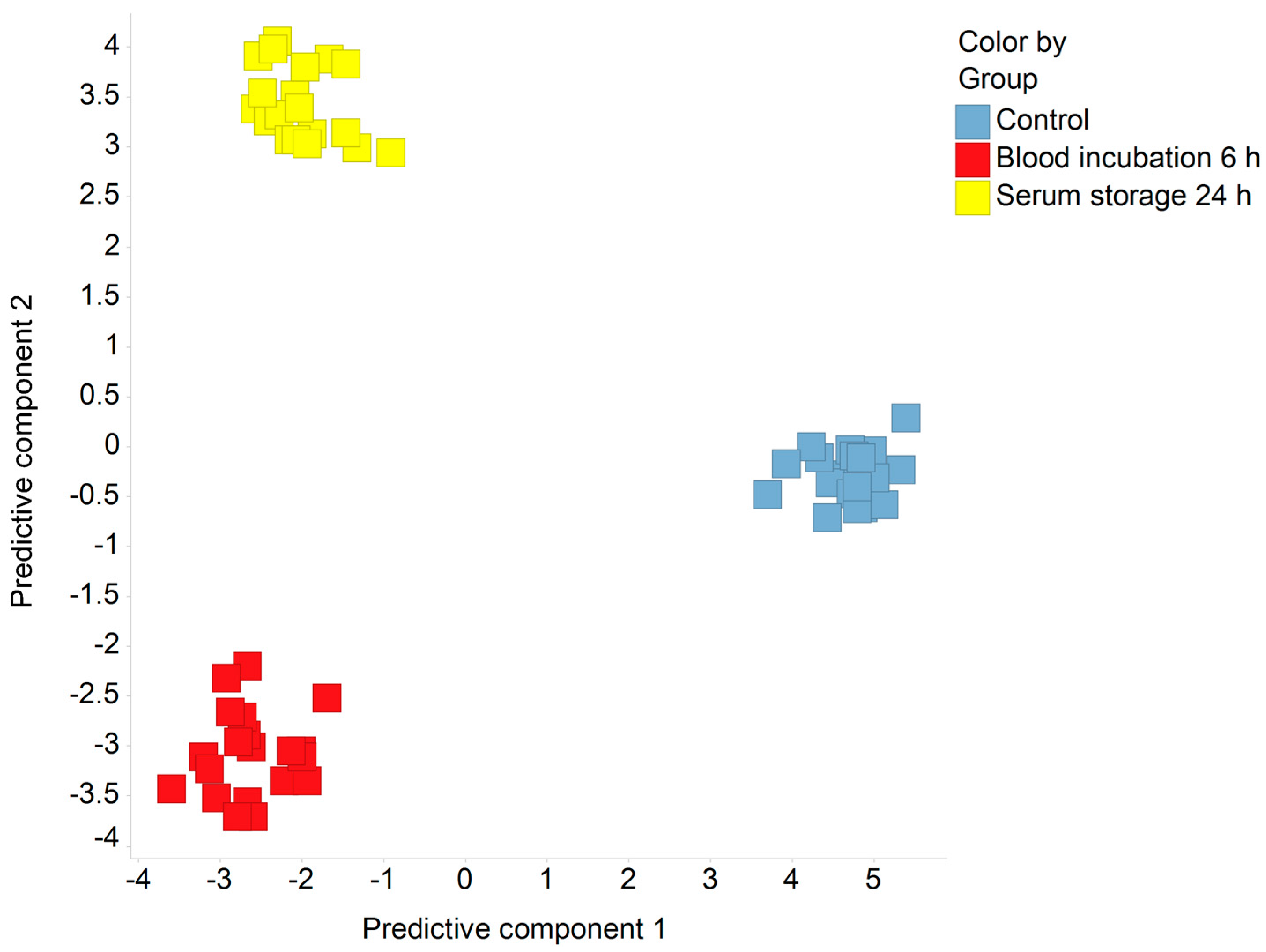
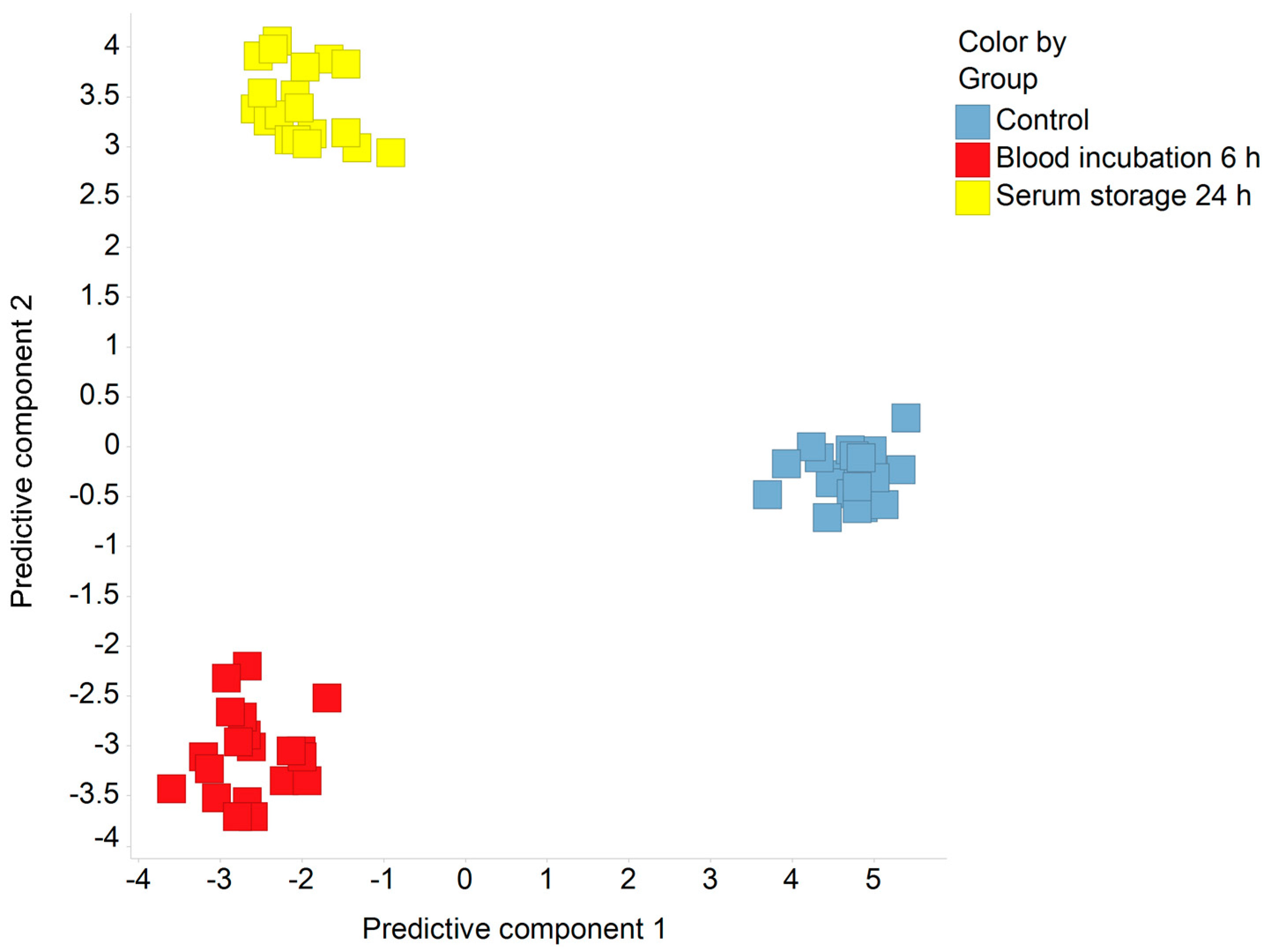
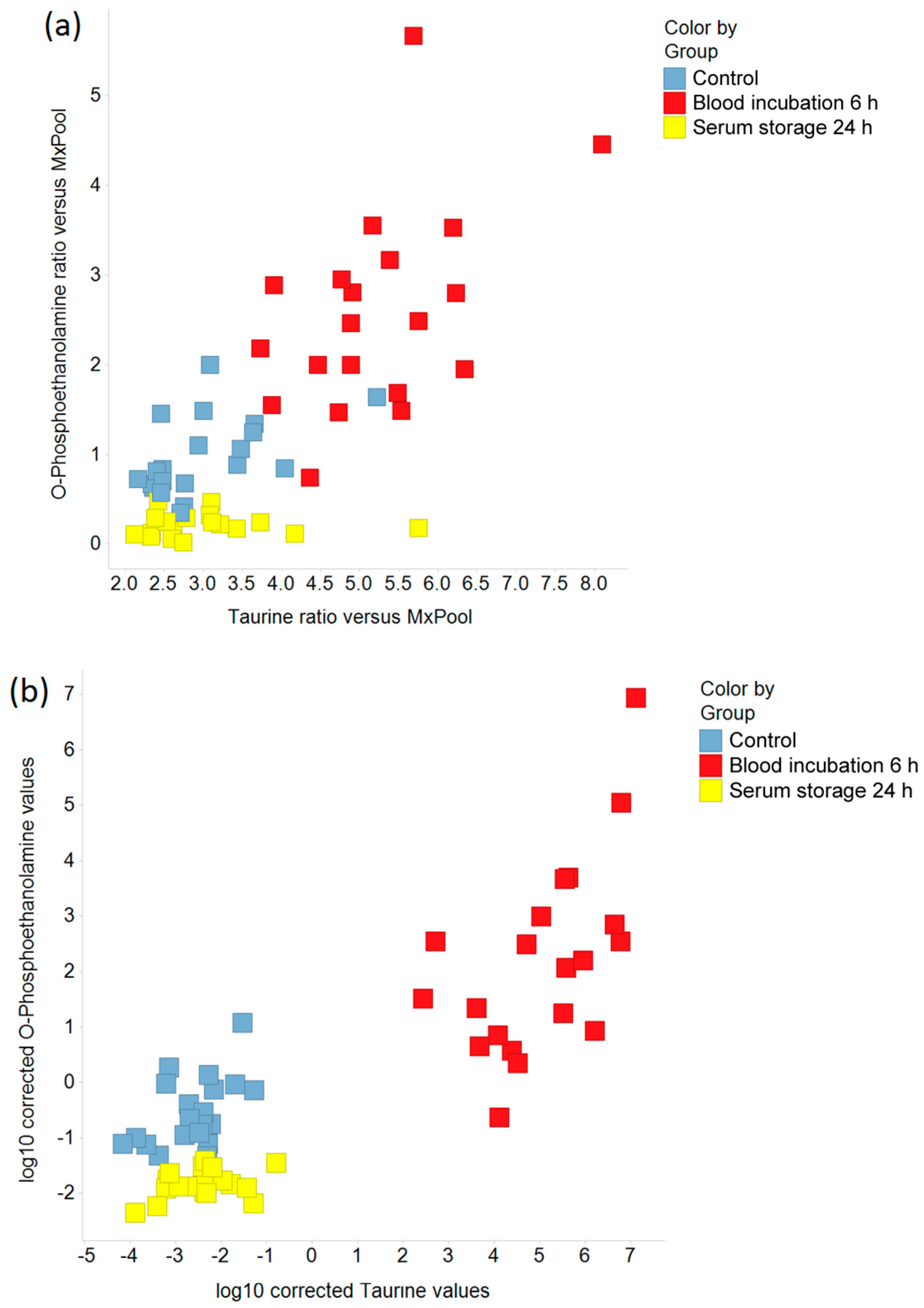
2.1. Discrimination of Pre-Analytical Confounded and Control Sample Groups
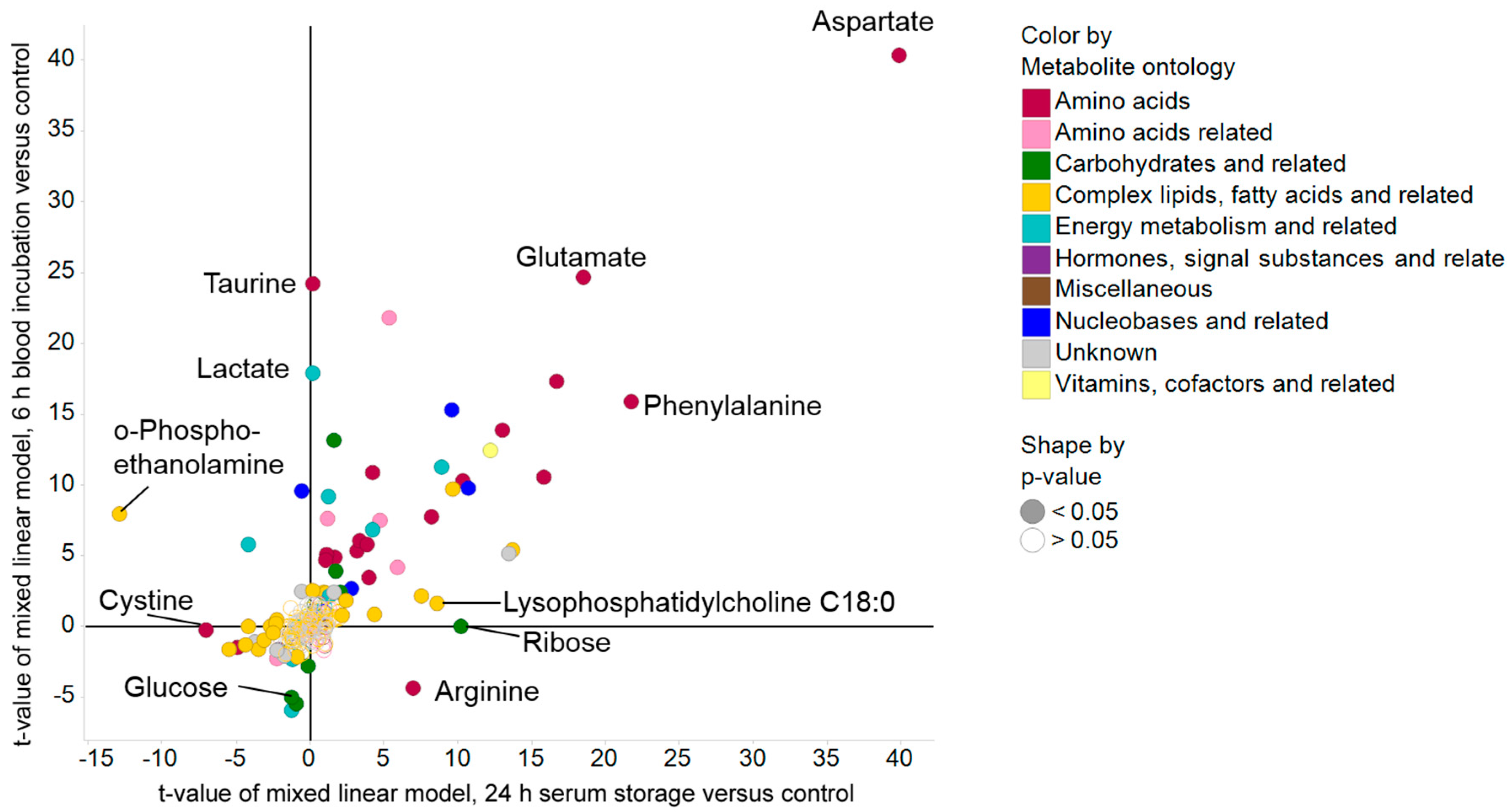
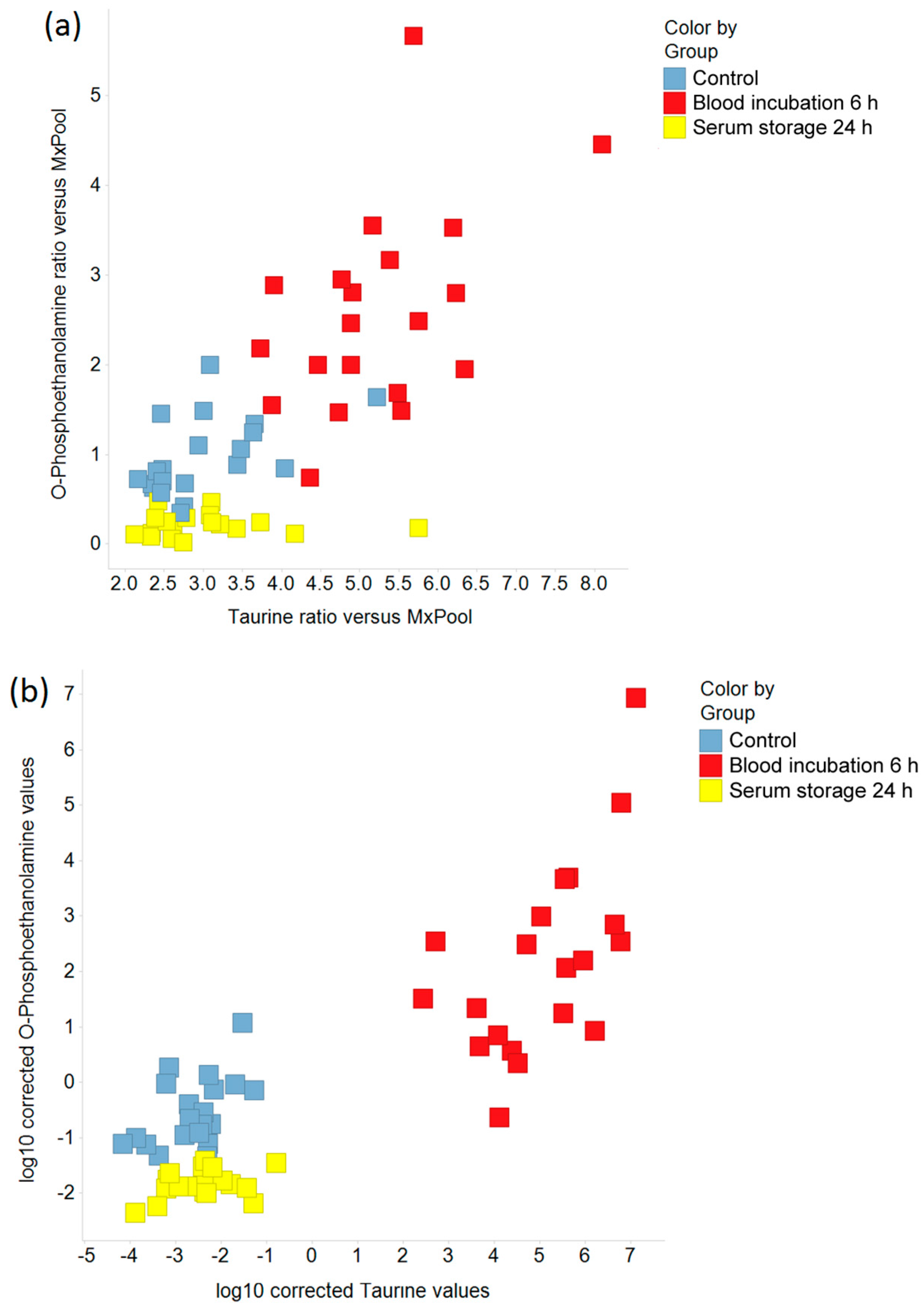
2.2. Pre-Analytical Variation Affects the Serum Metabolome
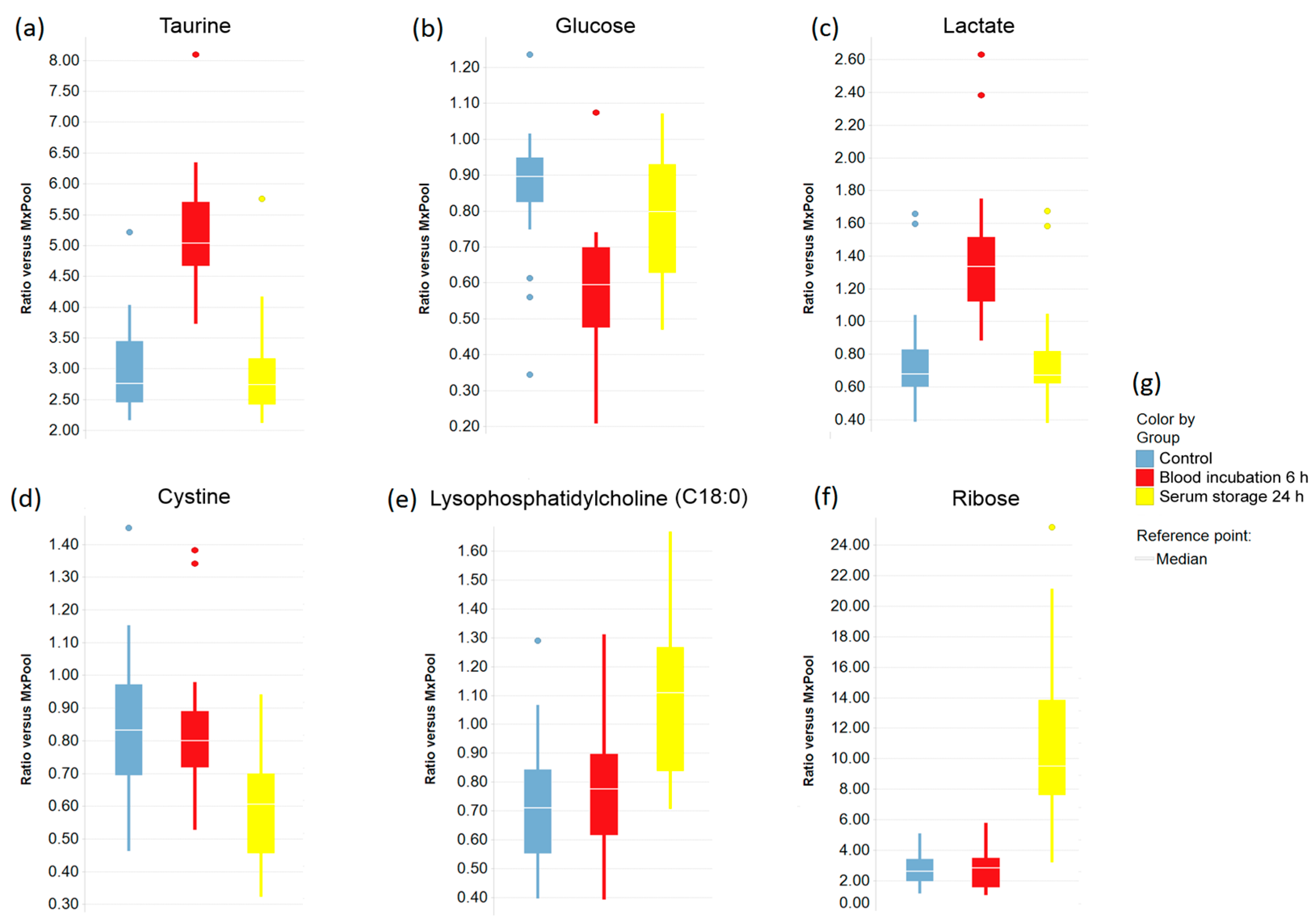
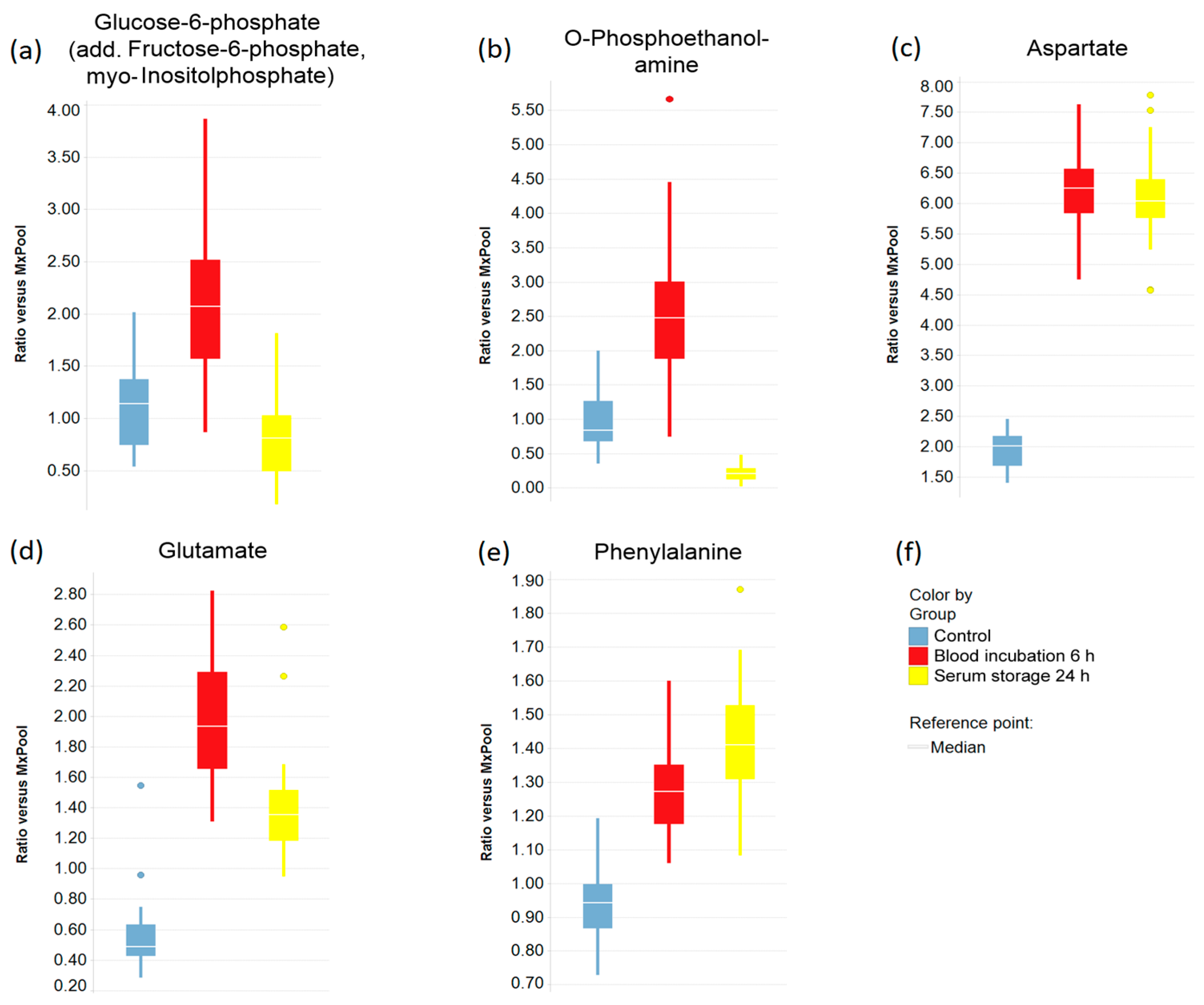
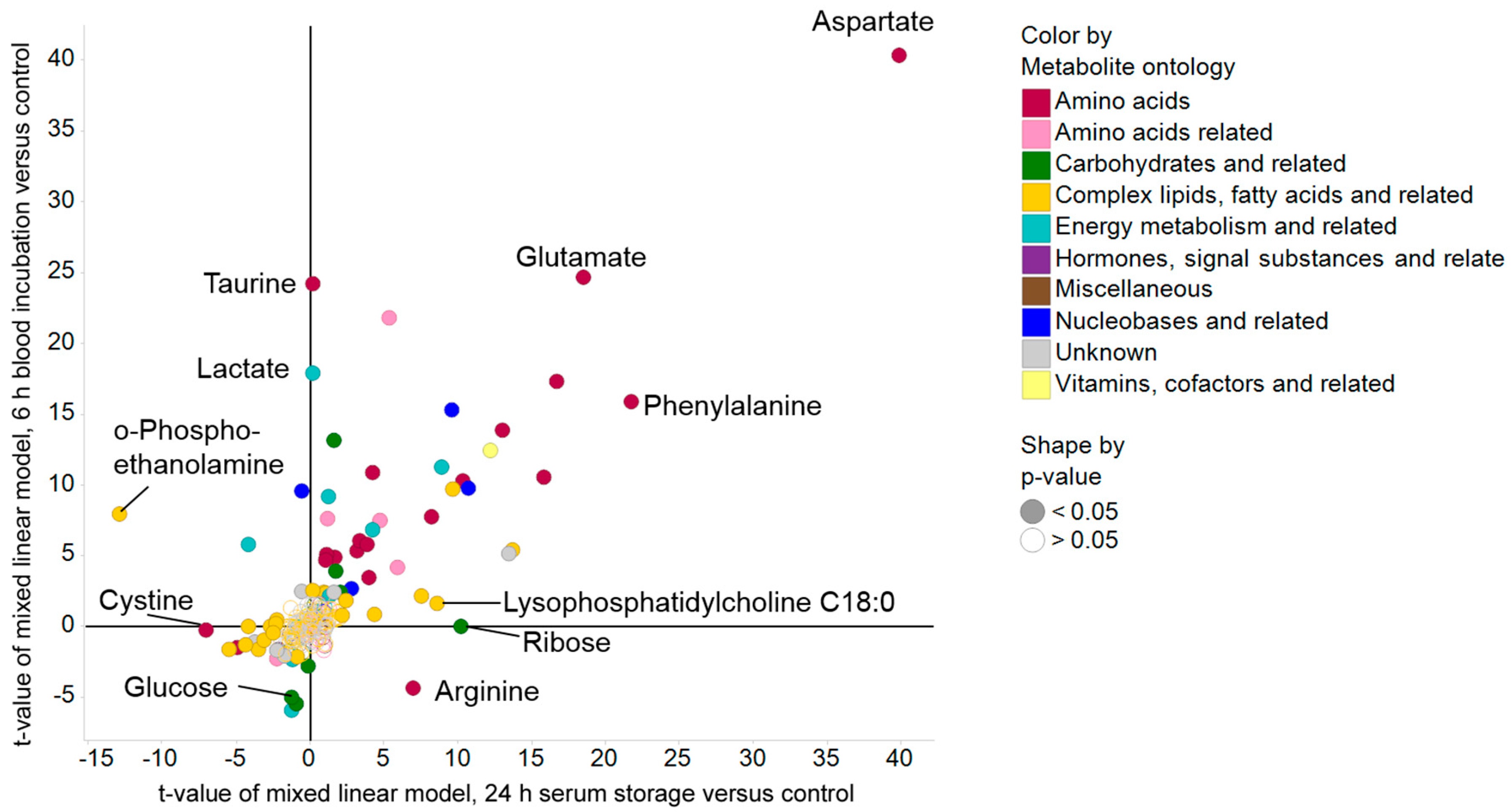
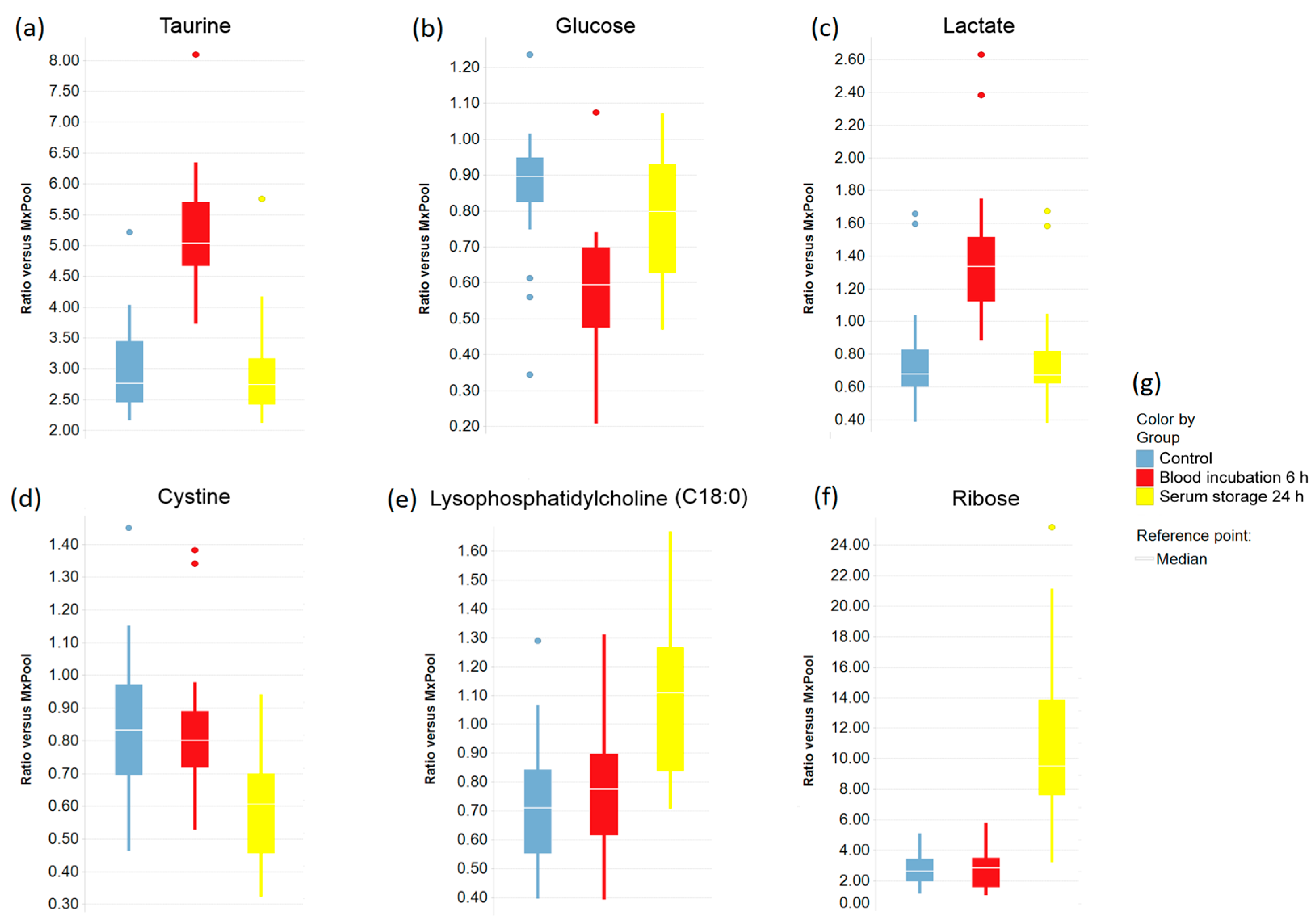
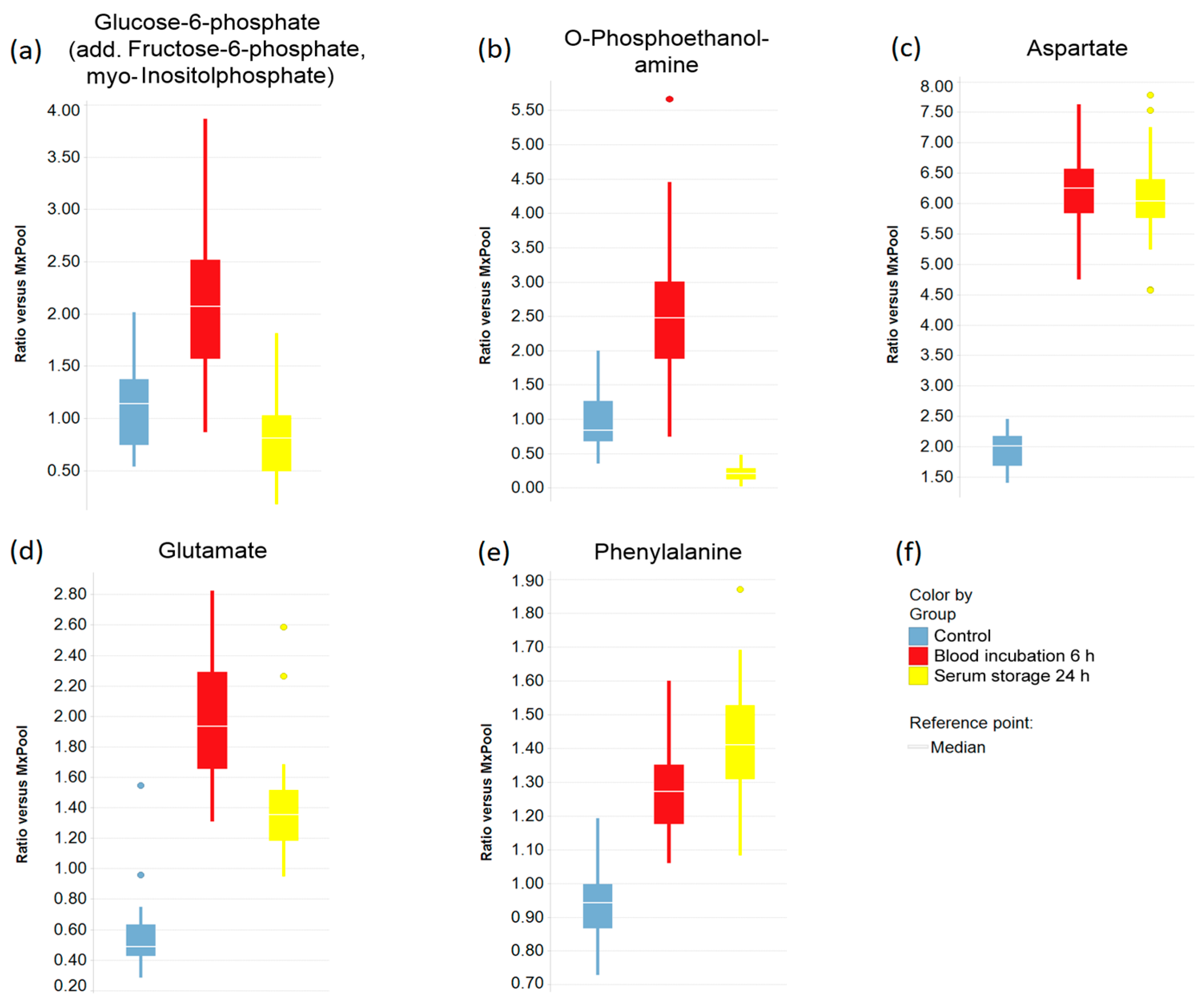
2.3. Impact of Pre-Analytical Variation on Selected Serum Metabolites
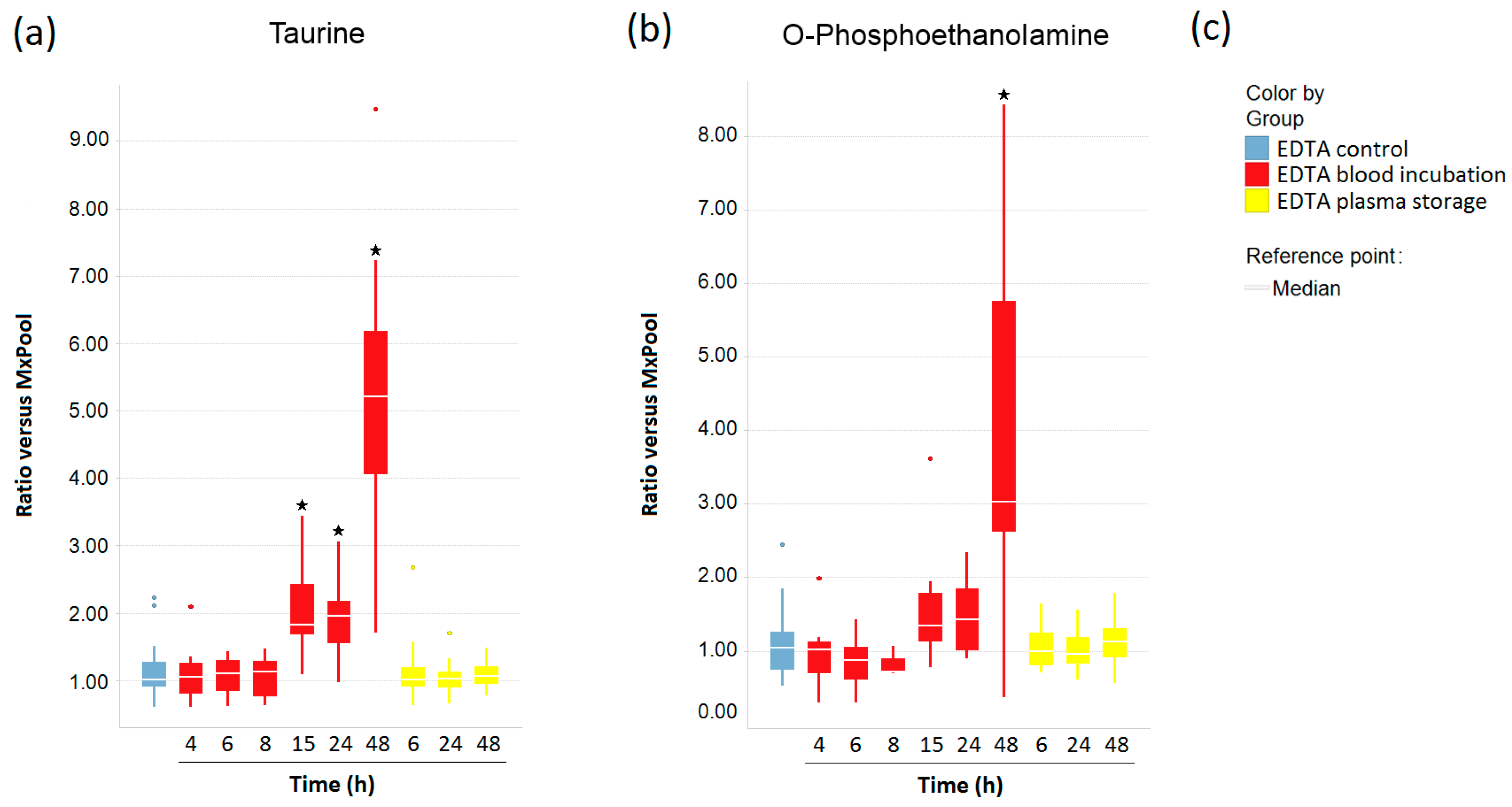
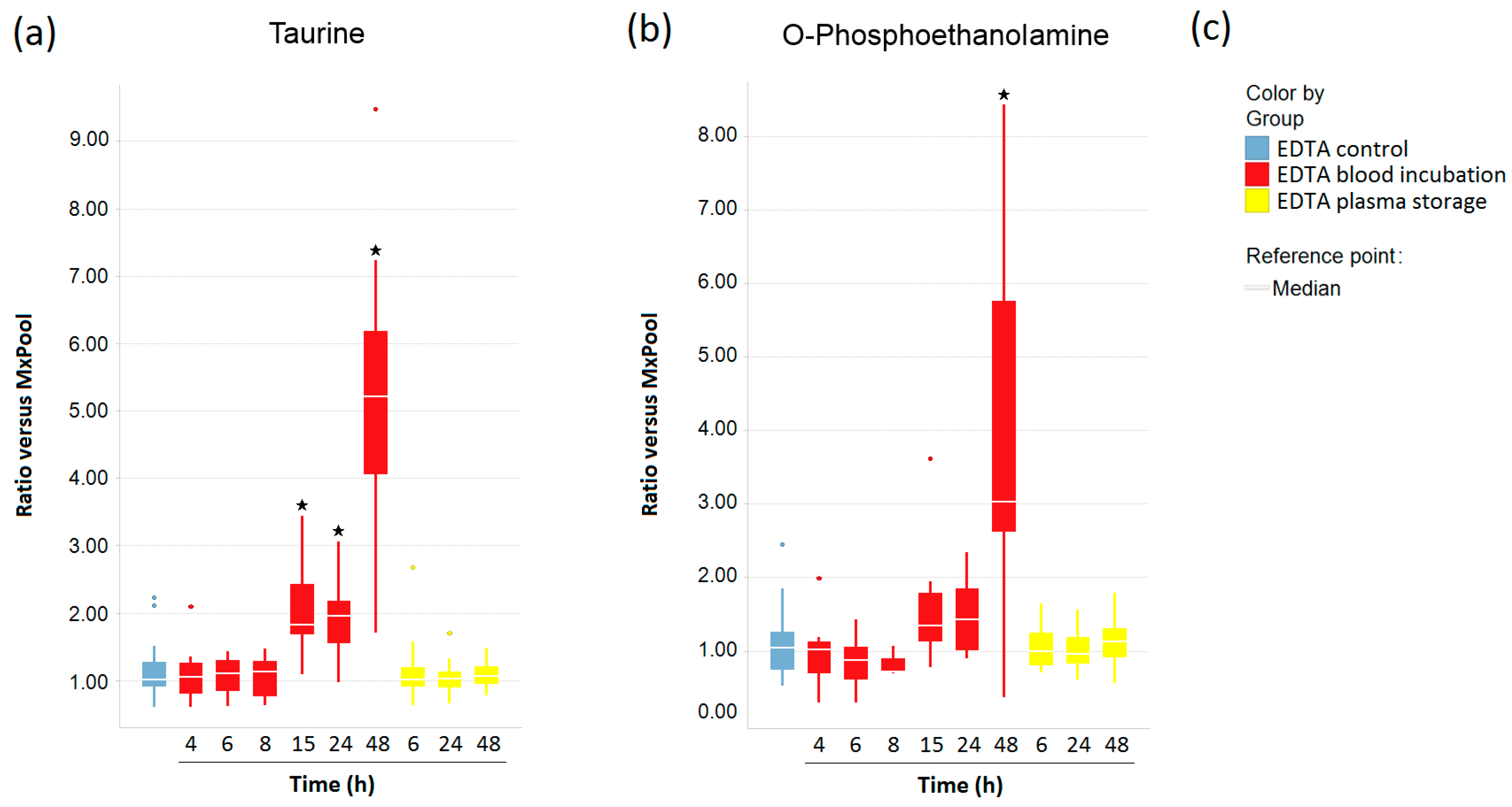
2.4. Impact of Pre-Analytical Variation on Taurine and O-Phosphoethanolamine in EDTA Blood and Plasma Samples
3. Discussion
4. Material and Methods
4.1. Serum and EDTA Plasma Samples
4.2. Metabolite Profiling Analysis
4.3. Data Normalization and Nomenclature
4.4. Statistical Analysis and Data Visualization
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Puchades-Carrasco, L.; Pineda-Lucena, A. Metabolomics in pharmaceutical research and development. Curr. Opin. Biotechnol. 2015, 35, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Chance, J.J.; Church, S.; Dazzi, P.; Fontana, R.; Giavarina, D.; Grankvist, K.; Huisman, W.; Kouri, T.; Palicka, V.; et al. Preanalytical quality improvement: From dream to reality. Clin. Chem. Lab. Med. 2011, 49, 1113–1126. [Google Scholar] [CrossRef] [PubMed]
- Fliniaux, O.; Gaillard, G.; Lion, A.; Cailleu, D.; Mesnard, F.; Betsou, F. Influence of common preanalytical variations on the metabolic profile of serum samples in biobanks. J. Biomol. NMR 2011, 51, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Peter, A.; Franken, H.; Zhao, X.; Neukamm, S.S.; Rosenbaum, L.; Lucio, M.; Zell, A.; Haring, H.U.; Xu, G.; et al. Preanalytical aspects and sample quality assessment in metabolomics studies of human blood. Clin. Chem. 2013, 59, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Chen, Y.; Xi, C.; Zhang, R.; Song, Y.; Zhan, Q.; Bi, X.; Abliz, Z. Liquid chromatography-tandem mass spectrometry-based plasma metabonomics delineate the effect of metabolites’ stability on reliability of potential biomarkers. Anal. Chem. 2013, 85, 2606–2610. [Google Scholar] [CrossRef] [PubMed]
- Barri, T.; Dragsted, L.O. Uplc-esi-qtof/ms and multivariate data analysis for blood plasma and serum metabolomics: Effect of experimental artefacts and anticoagulant. Anal. Chim. Acta 2013, 768, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Zivkovic, A.M.; Wiest, M.M.; Nguyen, U.T.; Davis, R.; Watkins, S.M.; German, J.B. Effects of sample handling and storage on quantitative lipid analysis in human serum. Metabolomics 2009, 5, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Anton, G.; Wilson, R.; Yu, Z.H.; Prehn, C.; Zukunft, S.; Adamski, J.; Heier, M.; Meisinger, C.; Romisch-Margl, W.; Wang-Sattler, R.; et al. Pre-analytical sample quality: Metabolite ratios as an intrinsic marker for prolonged room temperature exposure of serum samples. PLoS ONE 2015, 10, e0121495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamlage, B.; Maldonado, S.G.; Bethan, B.; Peter, E.; Schmitz, O.; Liebenberg, V.; Schatz, P. Quality markers addressing preanalytical variations of blood and plasma processing identified by broad and targeted metabolite profiling. Clin. Chem. 2014, 60, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, A.; Sugimoto, M.; Suzuki, A.; Hatakeyama, Y.; Enomoto, A.; Harada, S.; Soga, T.; Tomita, M.; Takebayashi, T. Effects of processing and storage conditions on charged metabolomic profiles in blood. Electrophoresis 2015, 36, 2148–2155. [Google Scholar] [CrossRef] [PubMed]
- Kalra, J. Medical errors: Impact on clinical laboratories and other critical areas. Clin. Biochem. 2004, 37, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Breier, M.; Wahl, S.; Prehn, C.; Fugmann, M.; Ferrari, U.; Weise, M.; Banning, F.; Seissler, J.; Grallert, H.; Adamski, J.; et al. Targeted metabolomics identifies reliable and stable metabolites in human serum and plasma samples. PLoS ONE 2014, 9, e89728. [Google Scholar] [CrossRef] [PubMed]
- Diamandis, E.P. Cancer biomarkers: Can we turn recent failures into success? J. Natl. Cancer Inst. 2010, 102, 1462–1467. [Google Scholar] [CrossRef] [PubMed]
- Gunther, K.; Malentacchi, F.; Verderio, P.; Pizzamiglio, S.; Ciniselli, C.M.; Tichopad, A.; Kubista, M.; Wyrich, R.; Pazzagli, M.; Gelmini, S. Implementation of a proficiency testing for the assessment of the preanalytical phase of blood samples used for rna based analysis. Clin. Chim. Acta 2012, 413, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.J.; Vitzthum, F. Effects of preanalytical variables on peptide and protein measurements in human serum and plasma: Implications for clinical proteomics. Expert Rev. Proteom. 2006, 3, 409–426. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Mahecha, A.; Kuzyk, M.A.; Domanski, D.; Borchers, C.H.; Basik, M. The effect of pre-analytical variability on the measurement of mrm-ms-based mid- to high-abundance plasma protein biomarkers and a panel of cytokines. PLoS ONE 2012, 7, e38290. [Google Scholar] [CrossRef] [PubMed]
- Gundisch, S.; Hauck, S.; Sarioglu, H.; Schott, C.; Viertler, C.; Kap, M.; Schuster, T.; Reischauer, B.; Rosenberg, R.; Verhoef, C.; et al. Variability of protein and phosphoprotein levels in clinical tissue specimens during the preanalytical phase. J. Proteome Res. 2012, 11, 5748–5762. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, R. Preanalytics: What can metabolomics learn from clinical chemistry? Bioanalysis 2015, 7, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Bearden, D.W.; Beger, R.D.; Broadhurst, D.; Dunn, W.B.; Edison, A.S.; Guillou, C.G.; Trengove, R.D.; Viant, M.R.; Wilson, I. The new data quality task group (dqtg): Ensuring high quality data today and in the future. Metabolomics 2014, 10, 539–540. [Google Scholar] [CrossRef]
- Yin, P.; Lehmann, R.; Xu, G. Effects of pre-analytical processes on blood samples used in metabolomics studies. Anal. Bioanal. Chem. 2015, 407, 4879–4892. [Google Scholar] [CrossRef] [PubMed]
- Simeon-Dubach, D.; Burt, A.D.; Hall, P.A. Quality really matters: The need to improve specimen quality in biomedical research. J. Pathol. 2012, 228, 431–433. [Google Scholar] [CrossRef] [PubMed]
- Bernini, P.; Bertini, I.; Luchinat, C.; Nincheri, P.; Staderini, S.; Turano, P. Standard operating procedures for pre-analytical handling of blood and urine for metabolomic studies and biobanks. J. Biomol. NMR 2011, 49, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Hubel, A.; Spindler, R.; Skubitz, A.P. Storage of human biospecimens: Selection of the optimal storage temperature. Biopreserv. Biobank. 2014, 12, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Hubel, A.; Aksan, A.; Skubitz, A.P.; Wendt, C.; Zhong, X. State of the art in preservation of fluid biospecimens. Biopreserv. Biobank. 2011, 9, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Elliott, P.; Peakman, T.C. The uk biobank sample handling and storage protocol for the collection, processing and archiving of human blood and urine. Int. J. Epidemiol. 2008, 37, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Gonon, A.T.; Sjoquist, P.O.; Lundberg, J.O.; Pernow, J. Arginase regulates red blood cell nitric oxide synthase and export of cardioprotective nitric oxide bioactivity. Proc. Natl. Acad. Sci. USA 2013, 110, 15049–15054. [Google Scholar] [CrossRef] [PubMed]
- Chap, H. Forty five years with membrane phospholipids, phospholipases and lipid mediators: A historical perspective. Biochimie 2016, 125, 234–249. [Google Scholar] [CrossRef] [PubMed]
- Hla, T.; Dannenberg, A.J. Sphingolipid signaling in metabolic disorders. Cell Metab. 2012, 16, 420–434. [Google Scholar] [CrossRef] [PubMed]
- Modica-Napolitano, J.S.; Renshaw, P.F. Ethanolamine and phosphoethanolamine inhibit mitochondrial function in vitro: Implications for mitochondrial dysfunction hypothesis in depression and bipolar disorder. Biol. Psychiatry 2004, 55, 273–277. [Google Scholar] [CrossRef]
- Schuller-Levis, G.B.; Park, E. Taurine: New implications for an old amino acid. FEMS Microbiol. Lett. 2003, 226, 195–202. [Google Scholar] [CrossRef]
- Ahtee, L.; Boullin, D.J.; Paasonen, M.K. Transport of taurine by normal human blood platelets. Br. J. Pharmacol. 1974, 52, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K.; Hirai, Y.; Yoshida, H.; Nakajima, T.; Usui, T. Free amino acid content of lymphocytes nd granulocytes compared. Clin. Chem. 1982, 28, 1758–1761. [Google Scholar] [PubMed]
- Hayes, K.C.; Pronczuk, A.; Addesa, A.E.; Stephan, Z.F. Taurine modulates platelet aggregation in cats and humans. Am. J. Clin. Nutr. 1989, 49, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Frendo, J.; Koj, A.; ZgliczyŃSki, J.M. Taurine in human blood platelets. Nature 1959, 183, 685. [Google Scholar] [CrossRef] [PubMed]
- Paasonen, M.K.; Solatunturi, E.; Nieminen, M.L. Blood platelets: Accumulation and release of taurine. Prog. Clin. Biol. Res. 1985, 179, 225–233. [Google Scholar] [PubMed]
- Kuc, S.; Koster, M.P.; Pennings, J.L.; Hankemeier, T.; Berger, R.; Harms, A.C.; Dane, A.D.; Schielen, P.C.; Visser, G.H.; Vreeken, R.J. Metabolomics profiling for identification of novel potential markers in early prediction of preeclampsia. PLoS ONE 2014, 9, e98540. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Aa, J.; Wang, G.; Yan, B.; Zhang, Y.; Wang, X.; Zhao, C.; Cao, B.; Shi, J.; Li, M.; et al. Differences in metabolite profile between blood plasma and serum. Anal. Biochem. 2010, 406, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Kastenmuller, G.; He, Y.; Belcredi, P.; Moller, G.; Prehn, C.; Mendes, J.; Wahl, S.; Roemisch-Margl, W.; Ceglarek, U.; et al. Differences between human plasma and serum metabolite profiles. PLoS ONE 2011, 6, e21230. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.D.; Macdonald, V.W.; Hess, J.R. Warm storage of whole blood for 72 hours. Transfusion 2007, 47, 2050–2056. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Gao, S.; Shu, C.; Wen, Y.; Yun, Y.; Tao, X.; Chen, W.; Zhang, F. Analysis of amino acids in human blood using uhplc-ms/ms: Potential interferences of storage time and vacutainer tube in pre-analytical procedure. Clin. Biochem. 2016, 49, 1372–1378. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Ichihara, K.; Hashiguchi, T.; Hidaka, Y.; Kang, D.; Maekawa, M.; Matsumoto, H.; Matsushita, K.; Okubo, S.; Tsuchiya, T.; et al. Evaluation of the short-term stability of specimens for clinical laboratory testing. Biopreserv. Biobank. 2015, 13, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Van Ravenzwaay, B.; Cunha, G.C.; Leibold, E.; Looser, R.; Mellert, W.; Prokoudine, A.; Walk, T.; Wiemer, J. The use of metabolomics for the discovery of new biomarkers of effect. Toxicol. Lett. 2007, 172, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Mutch, D.M.; Fuhrmann, J.C.; Rein, D.; Wiemer, J.C.; Bouillot, J.L.; Poitou, C.; Clement, K. Metabolite profiling identifies candidate markers reflecting the clinical adaptations associated with roux-en-y gastric bypass surgery. PLoS ONE 2009, 4, e7905. [Google Scholar] [CrossRef] [PubMed]
- Allaway, D.; Kamlage, B.; Gilham, M.S.; Hewson-Hughes, A.K.; Wiemer, J.C.; Colyer, A.; Rein, D. Effects of dietary glucose supplementation on the fasted plasma metabolome in cats and dogs. Metabolomics 2013, 9, 1096–1108. [Google Scholar] [CrossRef]
- Walk, T.B.; Dostler, M. Mass Spectrometry Method for Analysing Mixtures of Substances. PCT/EP2003/001274, 4 September 2003. [Google Scholar]
- Harville, D.A. Maximum likelihood approaches to variance component estimation and to related problems. J. Am. Stat. Assoc. 1977, 72, 320–338. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar]
- Trygg, J.; Wold, S. Orthogonal projections to latent structures (o-pls). J. Chemom. 2002, 16, 119–128. [Google Scholar] [CrossRef]
- Bylesjö, M.; Rantalainen, M.; Cloarec, O.; Nicholson, J.K.; Holmes, E.; Trygg, J. Opls discriminant analysis: Combining the strengths of pls-da and simca classification. J. Chemom. 2006, 20, 341–351. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Metabolite Ontology Class (Number) | Significantly Changed Metabolites versus the Control Group (Increase/Decrease) | |
|---|---|---|---|
| Number | Percent Change | ||
| Blood incubation (6 h) | All (225) | 54 (45/9) | 24 (20/4) |
| Amino acids (22) | 18 (17/1) | 82 (77/5) | |
| Amino acids related (15) | 5 (4/1) | 33 (27/7) | |
| Carbohydrates and related (10) | 6 (3/3) | 60 (30/30) | |
| Complex lipids, fatty acids and related (99) | 8 (7/1) | 8 (7/1) | |
| Energy metabolism and related (11) | 8 (6/2) | 73 (55/18) | |
| Hormones (2) | 0 (0/0) | 0 (0/0) | |
| Miscellaneous (9) | 0 (0/0) | 0 (0/0) | |
| Nucleobases and related (5) | 4 (4/0) | 80 (80/0) | |
| Vitamins, cofactors, and related (6) | 1 (1/0) | 17 (17/0) | |
| Unknowns (46) | 4 (3/1) | 9 (7/2) | |
| Serum storage (24 h) | All (225) | 48 (32/16) | 21 (14/7) |
| Amino acids (22) | 16 (14/2) | 73 (64/9) | |
| Amino acids related (15) | 4 (3/1) | 27 (20/7) | |
| Carbohydrates and related (10) | 1 (1/0) | 10 (10/0) | |
| Complex lipids, fatty acids, and related (99) | 17 (7/10) | 17 (7/10) | |
| Energy metabolism and related (11) | 3 (2/1) | 27 (18/9) | |
| Hormones (2) | 0 (0/0) | 0 (0/0) | |
| Miscellaneous (9) | 0 (0/0) | 0 (0/0) | |
| Nucleobases and related (5) | 3 (3/0) | 60 (60/0) | |
| Vitamins, cofactors, and related (6) | 1 (1/0) | 17 (17/0) | |
| Unknowns (46) | 3 (1/2) | 7 (2/4) | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamlage, B.; Neuber, S.; Bethan, B.; González Maldonado, S.; Wagner-Golbs, A.; Peter, E.; Schmitz, O.; Schatz, P. Impact of Prolonged Blood Incubation and Extended Serum Storage at Room Temperature on the Human Serum Metabolome. Metabolites 2018, 8, 6. https://doi.org/10.3390/metabo8010006
Kamlage B, Neuber S, Bethan B, González Maldonado S, Wagner-Golbs A, Peter E, Schmitz O, Schatz P. Impact of Prolonged Blood Incubation and Extended Serum Storage at Room Temperature on the Human Serum Metabolome. Metabolites. 2018; 8(1):6. https://doi.org/10.3390/metabo8010006
Chicago/Turabian StyleKamlage, Beate, Sebastian Neuber, Bianca Bethan, Sandra González Maldonado, Antje Wagner-Golbs, Erik Peter, Oliver Schmitz, and Philipp Schatz. 2018. "Impact of Prolonged Blood Incubation and Extended Serum Storage at Room Temperature on the Human Serum Metabolome" Metabolites 8, no. 1: 6. https://doi.org/10.3390/metabo8010006