Functional Aspects of PARP1 in DNA Repair and Transcription

1
Singapore Immunology Network, A*STAR, 8A Biomedical Grove, #03-06 Immunos, Singapore 138648, Singapore
2
Department of Microbiology, Yong Loo Lin School of Medicine, National University of Singapore, 5 Science Drive 2, MD4, Level 5, Singapore 117597, Singapore
*
Author to whom correspondence should be addressed.
Biomolecules 2012, 2(4), 524-548; https://doi.org/10.3390/biom2040524
Submission received: 18 September 2012
/
Revised: 24 October 2012
/
Accepted: 31 October 2012
/
Published: 12 November 2012
(This article belongs to the Special Issue DNA Damage Response)
Abstract
:Poly (ADP-ribose) polymerase 1 (PARP1) is an ADP-ribosylating enzyme essential for initiating various forms of DNA repair. Inhibiting its enzyme activity with small molecules thus achieves synthetic lethality by preventing unwanted DNA repair in the treatment of cancers. Through enzyme-dependent chromatin remodeling and enzyme-independent motif recognition, PARP1 also plays important roles in regulating gene expression. Besides presenting current findings on how each process is individually controlled by PARP1, we shall discuss how transcription and DNA repair are so intricately linked that disturbance by PARP1 enzymatic inhibition, enzyme hyperactivation in diseases, and viral replication can favor one function while suppressing the other.
Keywords:
ADP-ribosylation; DNA repair; transcription; PARP inhibitors; cancer; inflammation; oncogenic virus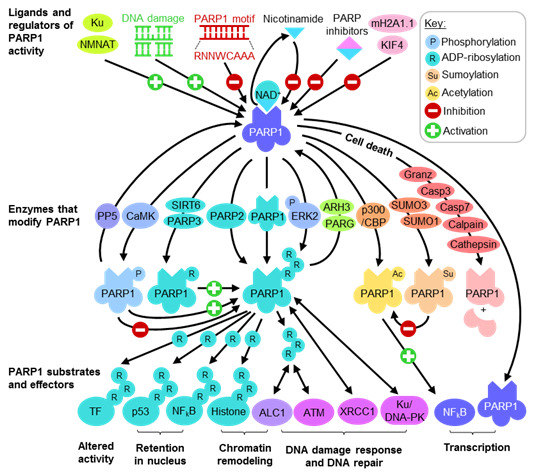
1. Introduction
Poly (ADP-ribose) polymerase 1 (PARP1) is well known as an ADP-ribosylating enzyme, which becomes activated upon binding to DNA single-strand and double-strand breaks (ssDB and dsDB respectively) [1,2,3,4,5,6]. The interaction is important for DNA repair, as auto-ribosylation is necessary to assemble and activate multiprotein complexes to carry out the process [6,7]. The critical role of PARP1 in DNA repair is reflected by its frequent upregulation in cancer [8,9], as well as the hypersensitivity of PARP1 null animals towards the mutagenic effects of DNA damaging agents [10]. Since PARP1 is involved in the repair of modified bases, ssDB and dsDB [7], blocking the ADP-ribosylation activity with small molecules, can achieve synthetic lethality with DNA damaging agents in the treatment of cancer [8,9,11,12,13,14,15,16,17]. Besides DNA repair, the importance of PARP1 as a transcriptional regulator is also well established. As an enzyme, PARP1 acts on chromatin remodeling complexes to control DNA accessibility for RNA polymerase [18,19,20,21,22]. PARP1 also functions as a transcription factor by binding an octamer motif in promoter elements to regulate gene expression [23,24,25,26,27,28,29,30]. Interestingly, despite the roles of PARP1 in DNA repair and transcription, little is known about how one process affects the other. In this review, we will summarize the roles of PARP1 in DNA repair and transcriptional regulation. Through the motif–PARP1 interaction, we will also discuss how transcription and DNA repair affect one another in normal cell functions and diseases states.
2. PARP1 Function and Regulation
PARP1 is the first to be identified among a family of 17 proteins that cleaves NAD+ for the ADP-ribosylation of protein acceptors, generating nicotinamide as a by-product [16,31]. The large 113kDa nuclear protein usually has low intrinsic enzymatic activity [32] which may be significantly enhanced by binding both ssDB and dsDB via either of its N-terminal zinc fingers [33], bringing about conformational changes through its third zinc finger to increase catalytic activity at the C-terminal [3,34,35]. As large amounts of negative charges are conferred by adding extensive polymers of ADP-ribose (PAR), PARP1 modulates the activity of its substrates, including itself, to control several important cellular functions (Figure 1) such as DNA damage repair, transcriptional regulation and cell death [6,16,19,20,21,22,31,36,37,38,39,40,41,42]. However, PAR is short-lived and as soon as its purpose is served, it is rapidly degraded within minutes of synthesis by the exoglycosidic and endoglycosidic activities of poly (ADP-ribose) glycohydrase (PARG) or PAR hydrolase (ARH) [6].
2.1. Regulating PARP1 ADP-Ribosylation Activity
PARP1 enzymatic activation accounts for the bulk of cellular ADP-ribosylation reactions [43,44] and consumes large amounts of NAD+. The accumulation of PAR is a cytotoxic signal as targeted disruption of PARG is shown to be embryonically lethal in mice and associated with apoptotic cell death in blastocysts [45]. Not surprisingly, PARP1 enzymatic activity is thus regulated at several levels (Figure 1). Acting directly at the catalytic domain, the by-product nicotinamide exerts mild inhibitory effects on PARP1 ADP-ribosylation activity [8,9,14,15,17]. The molecular basis for interference with NAD+ binding is well studied, and often imitated or improved upon when designing high affinity small molecule inhibitors targeting PARP1 functions. The end product PAR also helps to limit NAD+ consumption by mildly inhibiting PARP1 and, when becoming highly branched, confers excessive negative charges for repulsion from DNA, switching off ADP-ribosylation. Interestingly, while the best characterized ligands for the PARP1 enzyme are ssDB and dsDB, PARP1 has a higher affinity for intact DNA and specifically recognizes the octamer motif “RNNWCAAA” found in various gene promoters [23]. This interaction suppresses PARP1 ADP-ribosylation activity and interferes with its enzyme-dependent functions.
Figure 1.
PARP1 function and regulation. Abbreviations: Granz: Granzyme; Casp:Caspase; TF: Transcription factor.
Figure 1.
PARP1 function and regulation. Abbreviations: Granz: Granzyme; Casp:Caspase; TF: Transcription factor.
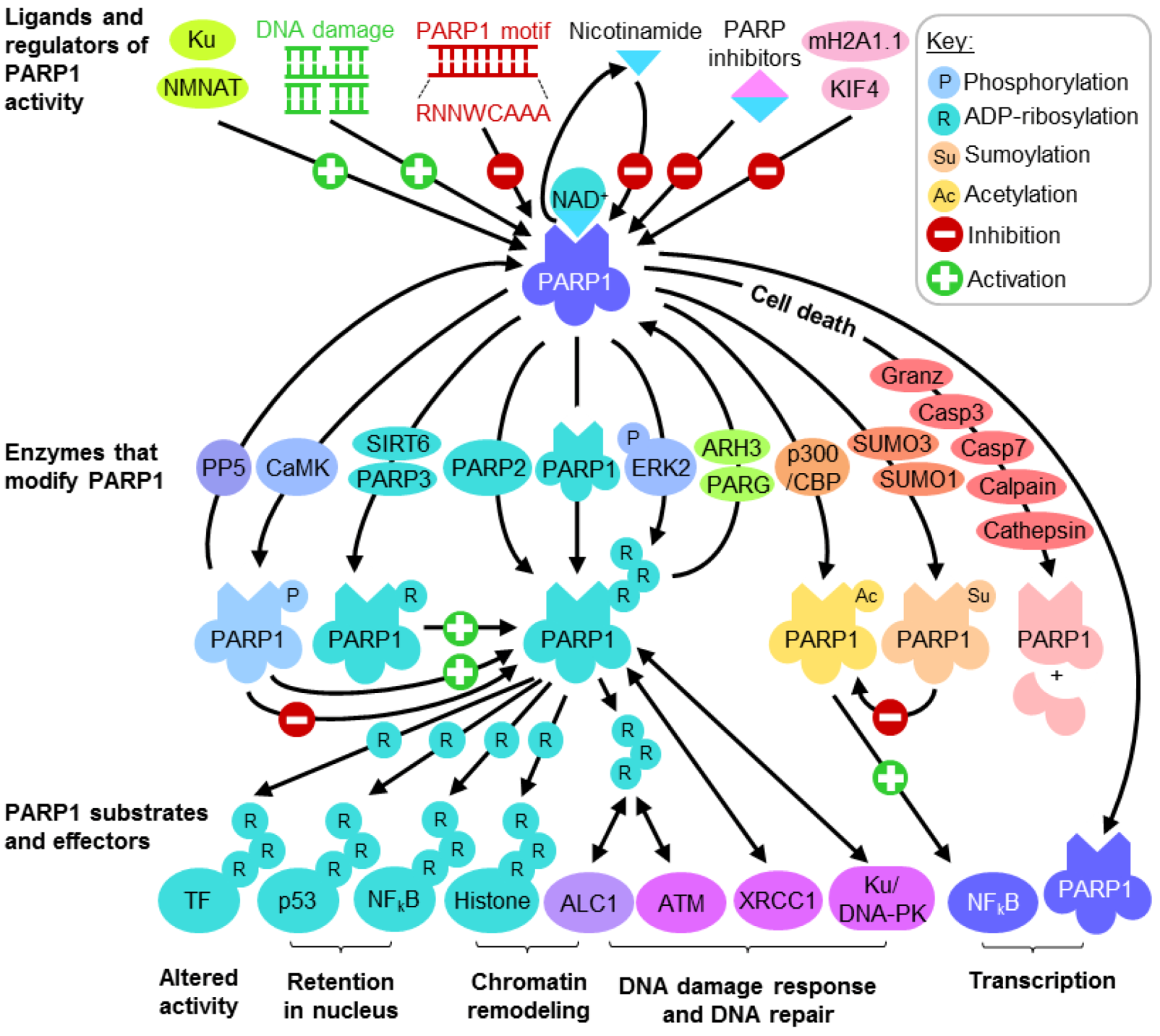
Various posttranslational modifications are known to regulate PARP1 enzyme activity. PARP1 may be acted upon by other members of the PARP family such as PARP2, and mono-ADP-ribosylation by PARP3 has been shown to enhance PARP1 activity and automodification [46]. Other enzymes possessing ADP-ribosylating activity, such as SIRT6, also act on PARP1, promoting dsDB repair in oxidative stress [47]. Depending on which residue is modified on the large protein, PARP1 phosphorylation seems to exert differential effects on its ADP-ribosylation activity. In the presence and absence of damaged DNA, its catalytic activity may be significantly enhanced and maximized by action of phosphorylated ERK2 [48,49,50]. Similarly, phosphorylation by activated calcium-dependent protein kinase (CaMKII) during neuronal development activates PARP1 enzyme and promotes the nuclear export of its negative regulator KIF4 [51]. However, overexpression of protein phosphatase 5 (PP5) increases PARP1 enzymatic activity towards dsDB [52], suggesting that the kinases and PP5 act on different residues to bring about contrasting effects on PARP1 enzymatic activity. Intricate cross-talks between PARP1 acetylation and sumoylation have also been observed, where modification by SUMO1 and SUMO3 prevents p300-mediated acetylation of PARP1 [53]. Given the number and types of posttranslational modifications PARP1 is subjected to, it would not be surprising to find more of such cross-talks.
Besides direct interference with catalysis and molecular switches by posttranslational modification, several proteins are known to bind and modulate PARP1 activity. Perhaps acting as a convenient source of substrate, nuclear NMNAT catalyzes the final step of NAD+ synthesis and associates with PAR to enhance PARP1 enzyme activity [54]. Other proteins regulating PARP1 activity include Ku [52], histone variant mH2A1.1 [55] and KIF4 [56]. Another important strategy often used in cell death pathways to control PARP1 activity is cleavage by proteases. In early apoptosis, PARP1 is one of the first substrates cleaved by caspase 3 and caspase 7 between the second and third zinc-binding domains [57,58], preventing DNA strand-break binding from inducing NAD+ catalysis [40,59,60]. In immune responses, cytotoxic lymphocytes and NK cells release of granzyme A and granzyme B that also target PARP1 [61]. In response to intracellular calcium perturbations, PARP1 may also be cleaved by calpain 1 [61,62]. Furthermore, PARP1 cleavage is also observed in autophagic and necrotic cell death pathways by action of various cathepsins [61,63]. The cleavage of PARP1 by a myriad of proteases at different loci seems to be a common mechanism in cell death, however, the significance of this is not yet fully understood.
2.2. PARP1 Substrates and Effectors Play Important Roles in Transcription and DNA Damage Response
PARP1 is itself the major acceptor of PAR [43]. The extensive branching network of PAR on PARP1 acts as the cue to attract and assist in assembling multiprotein complexes involved in chromatin remodeling, DNA repair and damage checkpoint signaling (Figure 1) [50,64,65,66,67,68,69]. Histones such as H1 and H2B are important substrates of PARP1 [22,65,70,71], which, when displaced by ADP-ribosylation, enables enhanced accessibility of large protein complexes assembled during DNA repair and transcription. After strand-break dependent activation, DNA repair scaffold proteins such as XRCC1 may be directly recruited by automodified PARP1 [72], while PAR provides the localization signal for directing the nucleosome repositioning enzyme ALC1 (Amplified in Liver Cancer 1) [73] in response to DNA damage. PAR also recruits the DNA damage checkpoint protein ATM (Ataxia Telangiectasia Mutated) [74], activating the signaling cascade for DNA damage and cell cycle arrest. Besides recruiting and activating nuclear complexes, PARP1 also exerts its effects by directly modifying protein activity and localization. Transcription factors and transcription coregulators such as SP1, Oct-1 and hnRNP K (heterogeneous nuclear ribonucleoprotein K) are known targets of PARP1, when ADP-ribosylated are repelled from DNA hence resulting in altered transcript expression profiles [31,75,76,77]. The bulky posttranslational modification prevents the association of transcription factors p53 and NF-κB to nuclear export factors such as Crm1, enabling nuclear retention [78,79]. Thus, through altering transcription factor function and localization, as well as remodeling chromatin structure and recruiting DNA processing complexes, PARP1 plays pivotal roles in both transcriptional regulation and DNA damage response.
3. PARP1 ADP-Ribosylation Activity Is Important for Mediating DNA Repair
Though not directly involved in any of the processes per se, PARP1 initiates and modulates multiple DNA repair pathways (Table 1) and is thus important for maintaining genomic integrity. Indeed, PARP1 knockout mice are highly susceptible to DNA damaging agents such as γ-irradiation and DNA alkylating agents, accounting for DNA strand break accumulation, increased sister chromatid exchange, and high genomic instability in them [80]. While these mice are viable and phenotypically normal [80], PARP1 knockout mice haploinsufficient for DNA repair enzymes such as Ku80 have increased spontaneous mutations and present higher liver and brain tumor incidence with age [81,82]. Female PARP1 knockout mice also develop mammary carcinomas with age, which is accelerated with the loss of p53 [83]. The importance of PARP1 for DNA repair is further demonstrated by embryonic lethality in knockout mice models doubly-deficient for PARP1 and DNA repair proteins Ku80 [82], BRCA1 [84], ATM [38] or DNA polymerase β [85]. The requirement for PARP1 in DNA damage repair is dependent on its ADP-ribosylation activity, as all male rats fed on a diet to deprive liver NAD+ spontaneously developed hepatocellular carcinoma with age [86].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DNA Repair Mechanism | PARP1 Function | References |
|---|---|---|
| Base excision repair (BER) | Binds AP site | [87] |
| Auto-modified PARP1 recruits BER complex | [88] | |
| Nucleotide excision repair (NER) | ADP-ribosylates XPA | [89,90] |
| Mismatch repair (MMR) | ADP-ribosylates MSH6 | [89,90] |
| Single-strand break repair (SSBR) | Auto-modified PARP1 recruits BER complex | [89,91,92] |
| Double-strand break repair by nonhomologous end joining (NHEJ) | Ku enhances PARP1 ADP-ribosylation activity | [89,90] |
| ADP-ribosylates and activates DNA-PKcs | ||
| Double-strand break repair by homologous recombination (HR) | Auto-modified PARP1 recruits Mre11 | [66] |
| PAR activates ATM signalling | [74] |
3.1. PARP1 in the Repair of Modified DNA
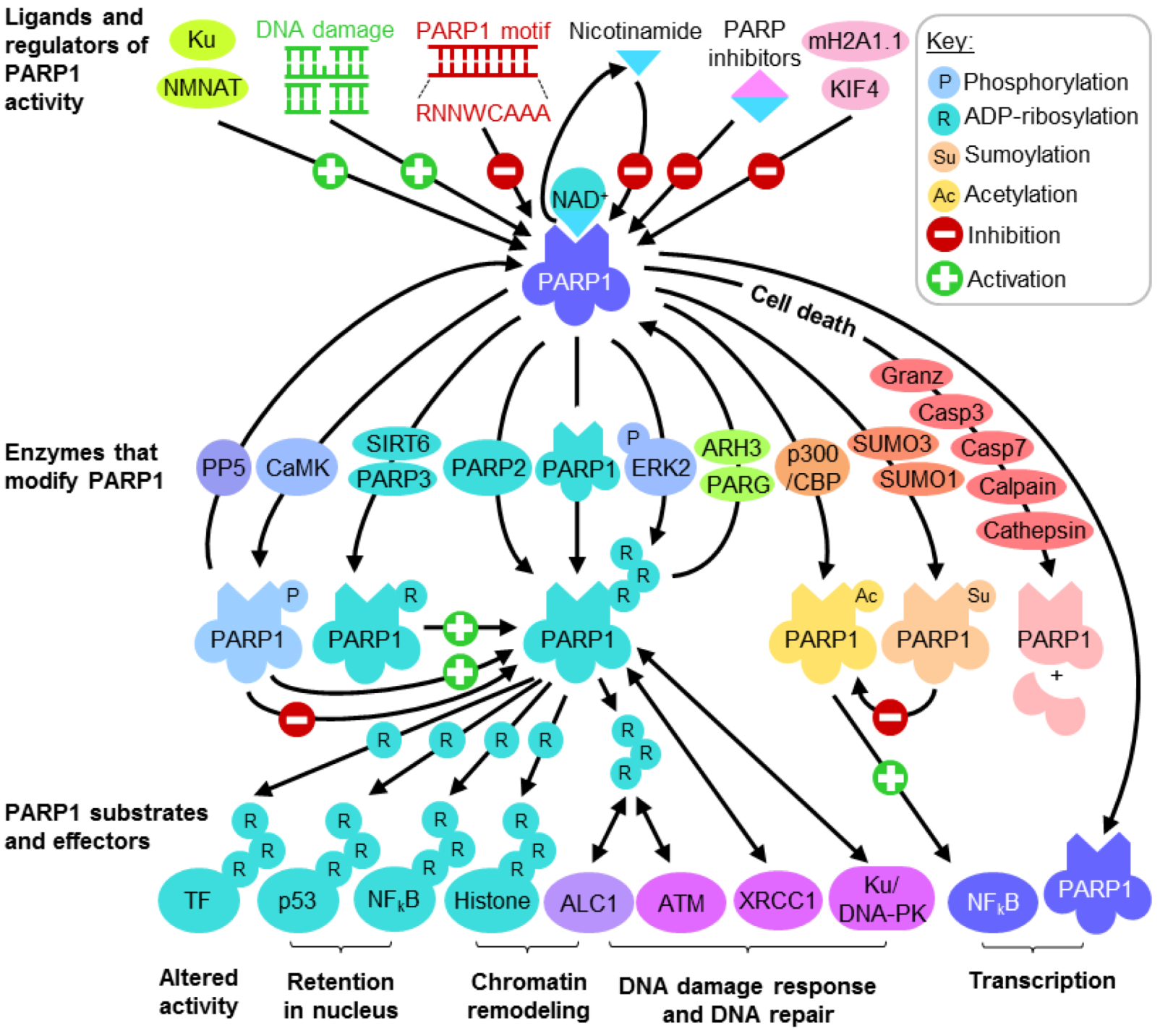
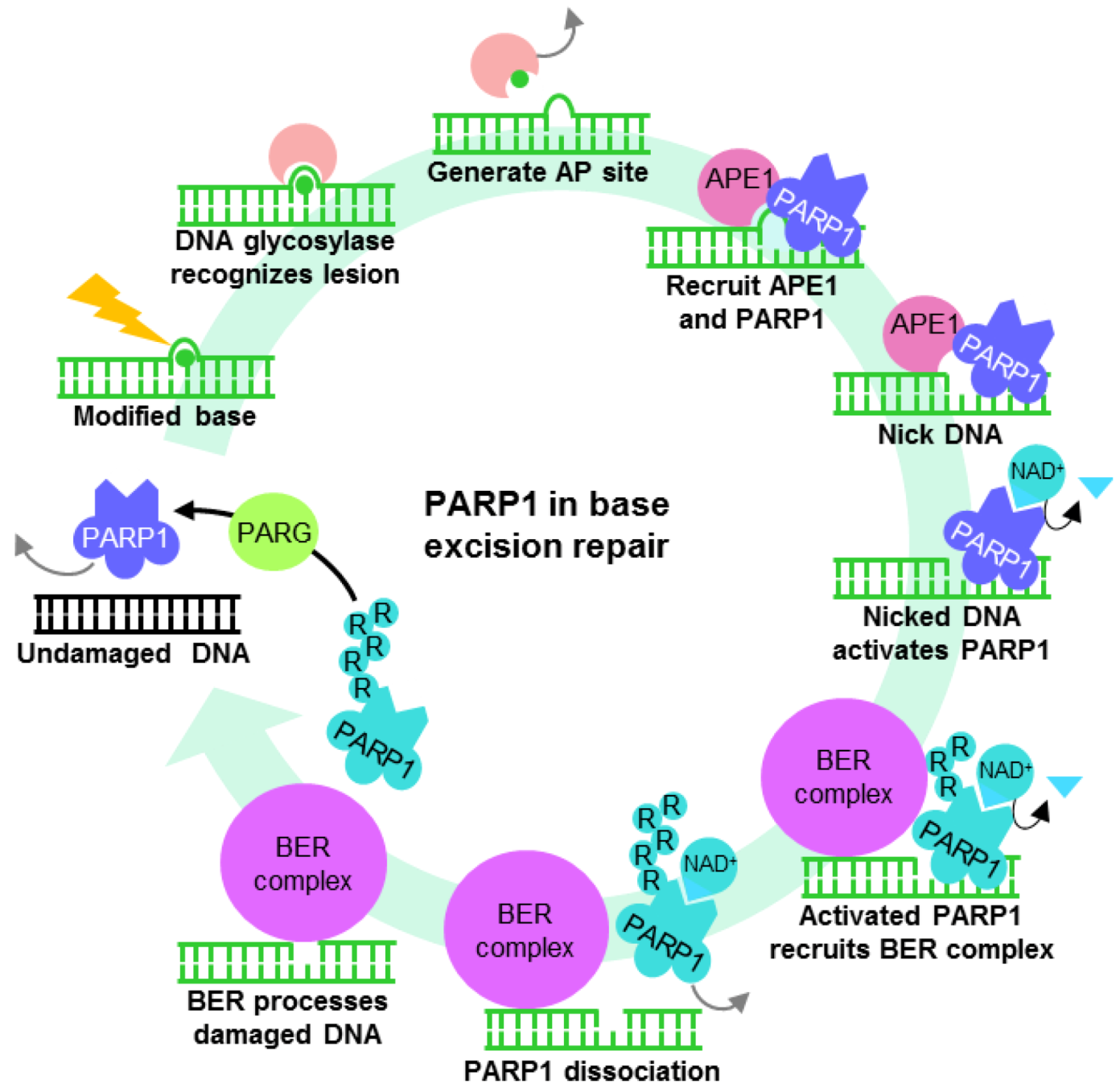
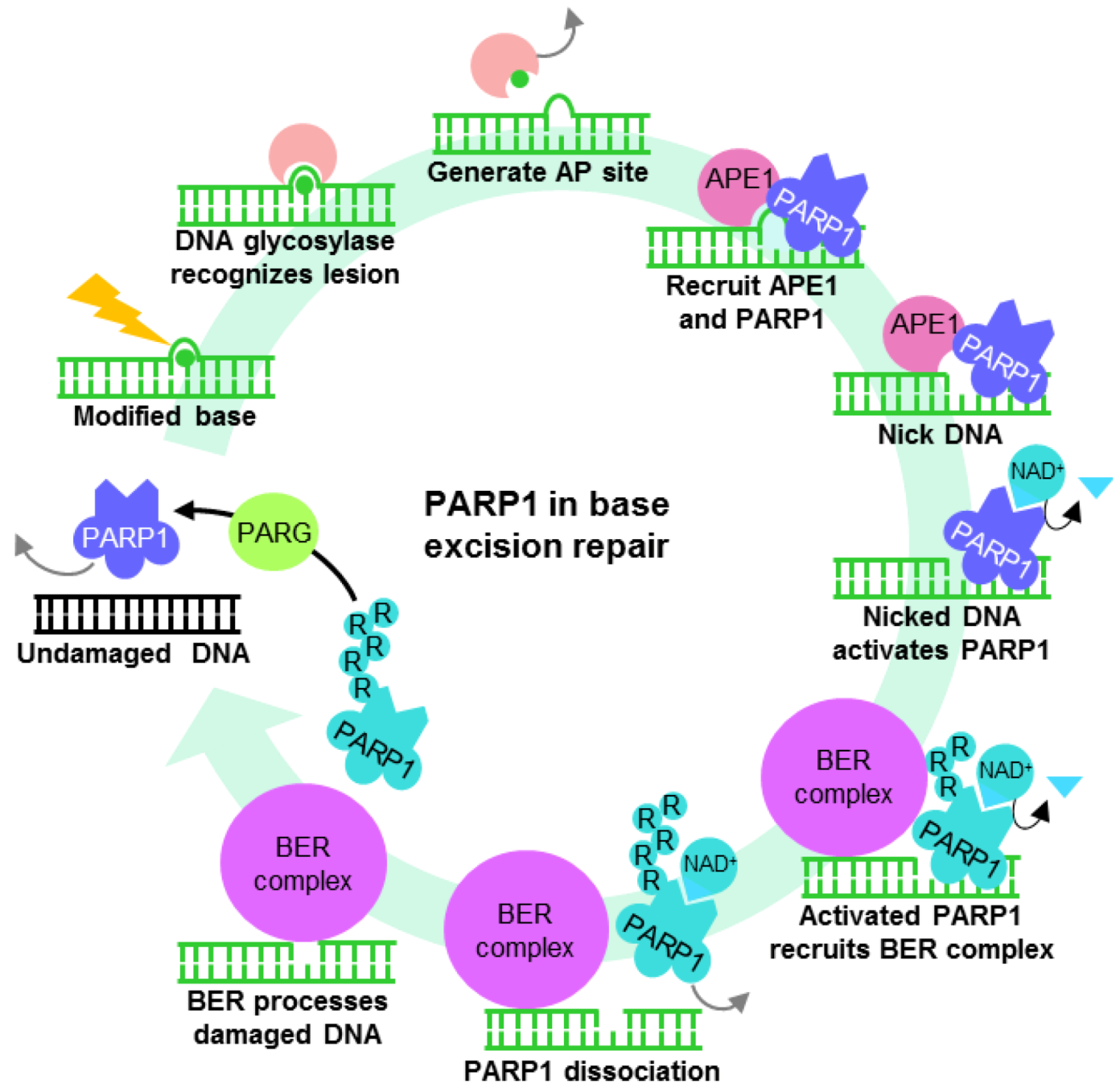
Several DNA repair pathways are in place to tackle a variety of genotoxic lesions. These pathways are activated depending on the type of DNA insult and phase within the cell cycle. Minor damage to bases such as methylation may be chemically reversed by specific glycosylases such as MGMT in an energy inefficient “suicide” reaction, restoring the base but rendering the enzyme unusable for subsequent reactions. To overcome this, base excision repair (BER) complexes targeting nonbulky modified bases may be recruited by PARP1 (Figure 2). Base modification by methylation, deamination and oxidation are recognized and removed by specific DNA glycosylases, generating an apurinic/apyrimidinic site (AP site) which, through a mechanism that is not completely understood, recruits PARP1 and APE1 [87]. APE1 removes the deoxyribose phosphate backbone at the site of lesion, generating nicked DNA which significantly enhances PARP1 ADP-ribosylation activity. The highly charged PAR produced keeps the DNA structure open [1] while various components of the BER complex are recruited. These components include the scaffold protein XRCC1 (X-ray Repair Cross-Complementing Protein 1), the DNA end-processing kinase/phosphatase PNK (Bifunctional polynucleotide phosphatase/kinase), the gap-filling polymerase DNA polymerase β and DNA ligase III [91,92] (Figure 2). By the time the BER complex is assembled, PARP1 accumulates enough negative charges for repulsion from the DNA lesion, enabling the BER and subsequent ligation to restore DNA.
Figure 2.
The role of PARP1 in base excision repair (BER). PARP1 may also mediate DNA single-strand break repair by recruiting the BER complex.
Figure 2.
The role of PARP1 in base excision repair (BER). PARP1 may also mediate DNA single-strand break repair by recruiting the BER complex.
Large adducts that distort DNA structure such as thymine dimers formed by UV irradiation require nucleotide excision repair (NER) for resolution [91,93]. In transcriptionally quiescent cells, global genomic NER (GG-NER) is initiated by the recruitment of XPC/HHRAD23B complex whereas in cells undergoing active transcription, the stalled RNA polymerase II is displaced with the aid of CSA and CSB proteins in transcription coupled NER (TC-NER). The two sub-pathways converge with the unwinding of DNA at the site of damage by transcription factor IIH, and the recruitment of DNA lesion recognition factors XPA and RPA. The endonucleases XPG and ERCC1/XPF then cut one strand of the unwound DNA at either ends of the damage to produce a 23–30 nt fragment containing the DNA lesion. While the resultant gap in DNA is not known to bind PARP1, XPA interestingly has been shown to be associated with PAR [90,91] (Table 1). Prior to DNA ligation the gap is repaired by DNA polymerase δ or ε, together with factors including PCNA, RPA and RFC.
When faced with singly or doubly mismatched bases and small loops arising from insertions or deletions, lesion recognition by MUTSα (MSH2-MHS6 heterodimer) initiates DNA mismatch repair (MMR) [92] and the recruitment of MUTLα (PMS2-MLH1 heterodimer). Conformation changes allow MUTSα-MUTLα to move along and nick DNA near the mismatch, enabling EXO1 (Exonuclease 1) to cleave and remove the damaged section of DNA. It is unclear whether PARP1 is activated at the site of DNA strand break, but MSH6 has been shown in independent studies [89,90] to be an acceptor of PAR (Table 1). Subsequently, gap filling by DNA polymerase δ or ε followed by ligation restores DNA.
3.2. PARP1 in the Repair of DNA Strand Breaks
DNA strand breaks may be directly induced by γ-irradiation or X-rays, as well as drugs such as bleomycin [51]. Under physiological conditions, they can be purposefully induced to enhance genetic diversity for meiotic recombination and antibody class switching [94]. Stalled replication forks may also require dsDB for resolution [66]. Single-strand DNA breaks are readily repaired by the BER complex, as PARP1 readily recognizing nicked DNA to organize its recruitment. The repair of dsDB is trickier as the involvement of both DNA strands prevents the use of neither as template. Thus, two pathways have evolved to repair such DNA lesions—nonhomologous end joining (NHEJ) and homologous recombination (HR). Even though their mechanistic details have not been fully elucidated, recent data indicates that PARP1 is not only involved in them (Table 1) but possibly also controls the choice of pathway utilized [95,96,97,98].
HR repairs DNA with high fidelity, using either homologous chromosomes in G1 phase or the sister chromatid after DNA replication as the blueprint for repair [99]. Following end recognition, the MRN complex comprising Mre11 nuclease, Rad50 and Nbs1 is recruited along with CtIP complex bearing BRCA1 to resect DNA, generating 3’ single-stranded overhangs which are stabilized by RPA1. PARP1 may contribute to this process at stalled replication forks by binding short single-stranded overhangs and recruiting Mre11 [66]. MRN also activates ATM signaling, initiating the DNA damage response including cell cycle arrest. BRCA2 then mediates the exchange of RPA1 for RAD5, and directs the presynaptic filament in its search for homologous DNA template. Strand invasion and exchange ensues, allowing DNA polymerase to extend the 3’ end of the invading strand. Repaired heteroduplexed DNA is generated after DNA ligase I joins the DNA ends and the resultant holiday junctions resolved by resolvases. When the damaged DNA may not be rapidly repaired, ssDNA-RPA1 may activate ATR signaling via DNA resection to sustain the DNA damage response [100,101,102,103,104]. This involves RPA1 interaction with ATRIP, RFC-mediated loading of the 9-1-1 clamp, and subsequent recruitment of TOPBP1 (topoisomerase binding protein 1) necessary for ATR activation.
NHEJ is an error-prone mechanism that ligates DNA ends together, often occurring in G1 phase when a suitable repair template is not available [105]. The ring-structured Ku heterodimer comprising Ku70/Ku80 slips and binds onto the broken ends of DNA, recruiting other factors such as DNA-PK (DNA-dependent protein kinase), XRCC4 and DNA ligase IV. DNA strand-break binding activates the catalytic subunit of DNA-PK (DNA-PKcs), initiating DNA damage signaling cascade by autophosphorylating substrates including ATM, p53 and itself, while DNA end-processing enzymes such as Artemis prepares the damaged DNA for ligation. Recent data points to the recruitment of MRN for end processing, as well. As V(D)J recombination for antibody class switching utilizes the NHEJ machinery, the unusual antibody profiles of PARP1 knockout mice point to its involvement in the process [94]. Instead of directly binding dsDB, PARP1 ADP-ribosylation activity is strongly enhanced by interaction with Ku, forming a functional complex with DNA-PK [52]. The kinase activity of DNA-PK is significantly increased by ADP-ribosylation [106]. However, the precise role of PARP1 in NHEJ is still unclear.
3.3. Uncontrolled PARP1 ADP-Ribosylation Activity during DNA Repair Results in Cell Death
When DNA damage is minimal, the recruitment of PARP1 to sites of DNA lesions activates the DNA damage response. Depending on the type of lesion encountered, signaling mediated by molecules such as p53 and ATM or ATR to promote cell cycle arrest, buying time for DNA repair enzymes to work. However, when DNA damage is extensive and irreparable, PARP1 is rapidly cleaved within minutes of DNA damage by effector caspases [107,108], presumably to prevent futile DNA repair when DNA is eventually cleaved later in the process. The dissociated N-terminal fragment is believed to remain bound to DNA strand breaks while the C-terminal is rapidly shuttled out of the nucleus. While the reason for PARP1 cleavage is not clear, their different subcellular localizations is thought to conserve energy required for apoptosis by preventing unnecessary NAD+ consumption. In support of this, energy failure by making large amounts of PAR when DNA is extensively damaged results in cell death by necrosis instead [8,16,40,41,59,109]. PAR can also mediate PARP1-dependent cell death (parthanatos) through the release of Apoptosis-inducing factor (AIF) from mitochondria in a caspase-independent manner [42,110].
4. PARP1 as a Transcriptional Regulator Controlling Expression of DNA Damage Response Genes
PARP1 is an important regulator of transcription, as can be seen at PAR-rich Drosophila chromosomal “puffs” undergoing active transcription [111]. The importance of PARP1 in controlling transcription was further supported by global alterations in gene expression [112,113,114], most notably of those involved in cell cycle, DNA repair and metabolism. Transcriptional regulation by PARP1 involves both ADP-ribosylation-dependent and independent mechanisms (Figure 3). Furthermore, PARP1 exerts its effects on transcription both in a DNA sequence-dependent and independent manner, through motif recognition at specific gene promoters [24,25,26,27,30,115,116,117,118,119,120,121,122,123] and chromatin remodeling [20,22], respectively.
4.1. PARP1 ADP-Ribosylation Activity Controls Transcription States
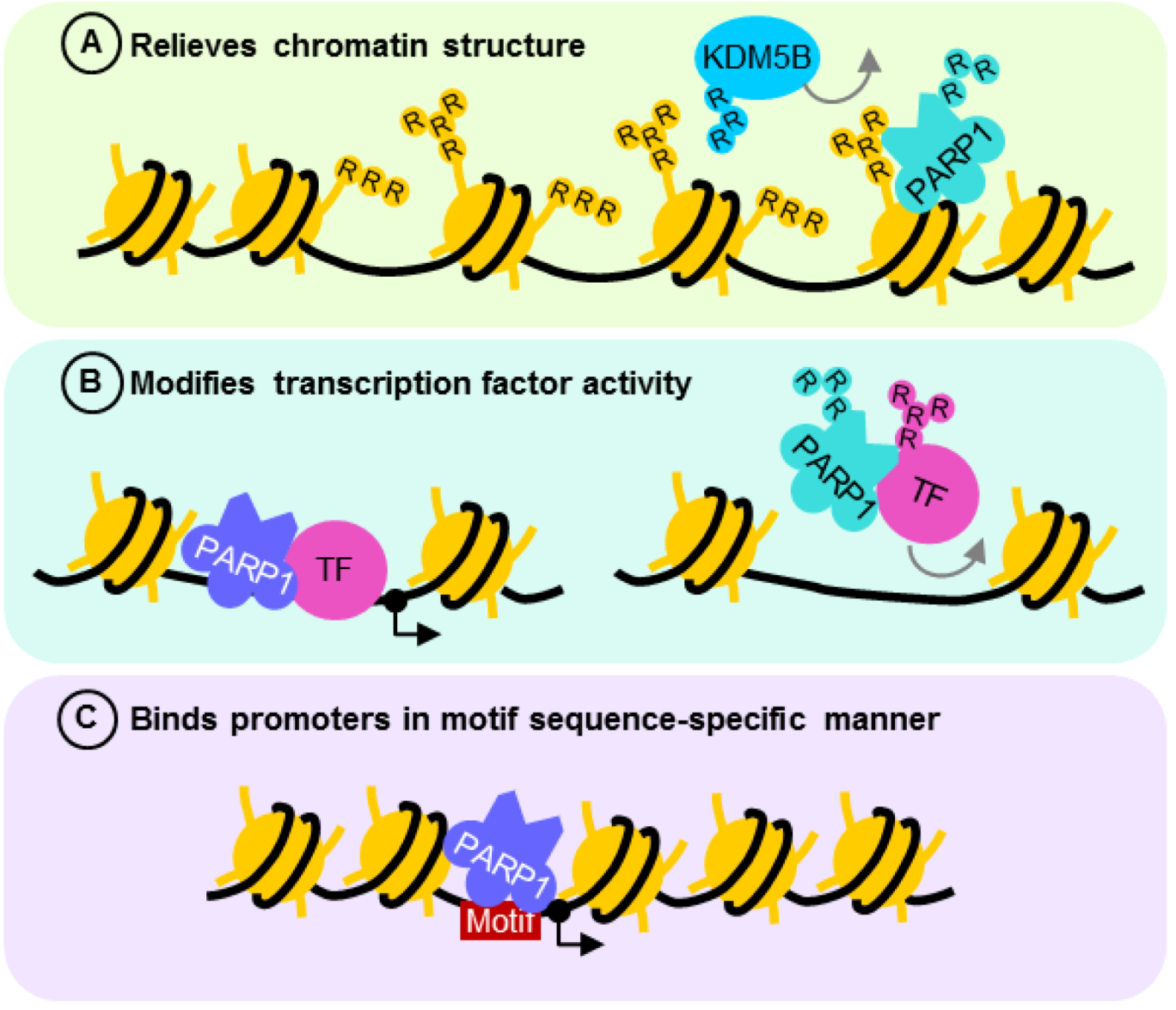
The “opening” of chromatin for active transcription often requires PARP1 ADP-ribosylation activity (Figure 3A) [18,22]. In the absence of NAD+, minimally automodified PARP1 acts as a transcriptional repressor by bridging neighboring nucleosomes to compact chromatin [18]. However, when enzymatically activated, the extensive negative charges conferred by automodified PARP1 loosens chromatin structure, thereby enabling transcription factors to bind. Upon recognition of its response element, transcription activators such as estrogen receptor α recruit a complex containing topoisomerase II-β and PARP1 [65]. While the topoisomerase resolves DNA secondary structures by creating transient dsDB, the DNA lesion activates PARP1 to ADP-ribosylate histones H1 and H2B. The negative charges on ADP-ribosylated histones repel DNA, loosening chromatin for increased DNA accessibility to the transcriptional machinery [18,22]. PARP1 is thus found in place of histone H1 in most transcriptionally active genes [21,69,70,113]. Modified histone H1 may also then be exchanged for histone H1-HMGB (histone H1 high mobility group B) favorable for transcription [65]. To maintain chromatin in its transcriptionally active state, PARP1 also prevents the histone demethylase KDM5B from approaching trimethylated histone H3K4 by repelling it from DNA through ADP-ribosylation [69]. The action of PARP1 on chromatin and chromatin remodeling complexes thus enables RNA polymerase II to load readily onto transcriptionally active promoter regions [69].
Figure 3.
The role of PARP1 in transcriptional regulation. (A) PARP1 relieves and maintains an “open” chromatin structure by ADP-ribosylation of histones or preventing the action of histone demethylase KDM5B. (B) PARP1 forms functional complexes with transcription factors such as NFκB, altering their activity depending on its state of posttranslational modification. This interaction need not activate or require PARP1 enzyme activity, although when stimulated, ADP-ribosylation usually reduces the affinity of the complex for DNA cis elements. The effect of PARP1 on transcription in both cases is dependent on the type of binding partner and nature of promoter element recognized. (C) PARP1 acts as a transcription activator or repressor by binding its recognition motif. Grey arrows are repulsion from DNA or chromatin. R—ADP-ribosylation; TF—transcription factor.
Figure 3.
The role of PARP1 in transcriptional regulation. (A) PARP1 relieves and maintains an “open” chromatin structure by ADP-ribosylation of histones or preventing the action of histone demethylase KDM5B. (B) PARP1 forms functional complexes with transcription factors such as NFκB, altering their activity depending on its state of posttranslational modification. This interaction need not activate or require PARP1 enzyme activity, although when stimulated, ADP-ribosylation usually reduces the affinity of the complex for DNA cis elements. The effect of PARP1 on transcription in both cases is dependent on the type of binding partner and nature of promoter element recognized. (C) PARP1 acts as a transcription activator or repressor by binding its recognition motif. Grey arrows are repulsion from DNA or chromatin. R—ADP-ribosylation; TF—transcription factor.
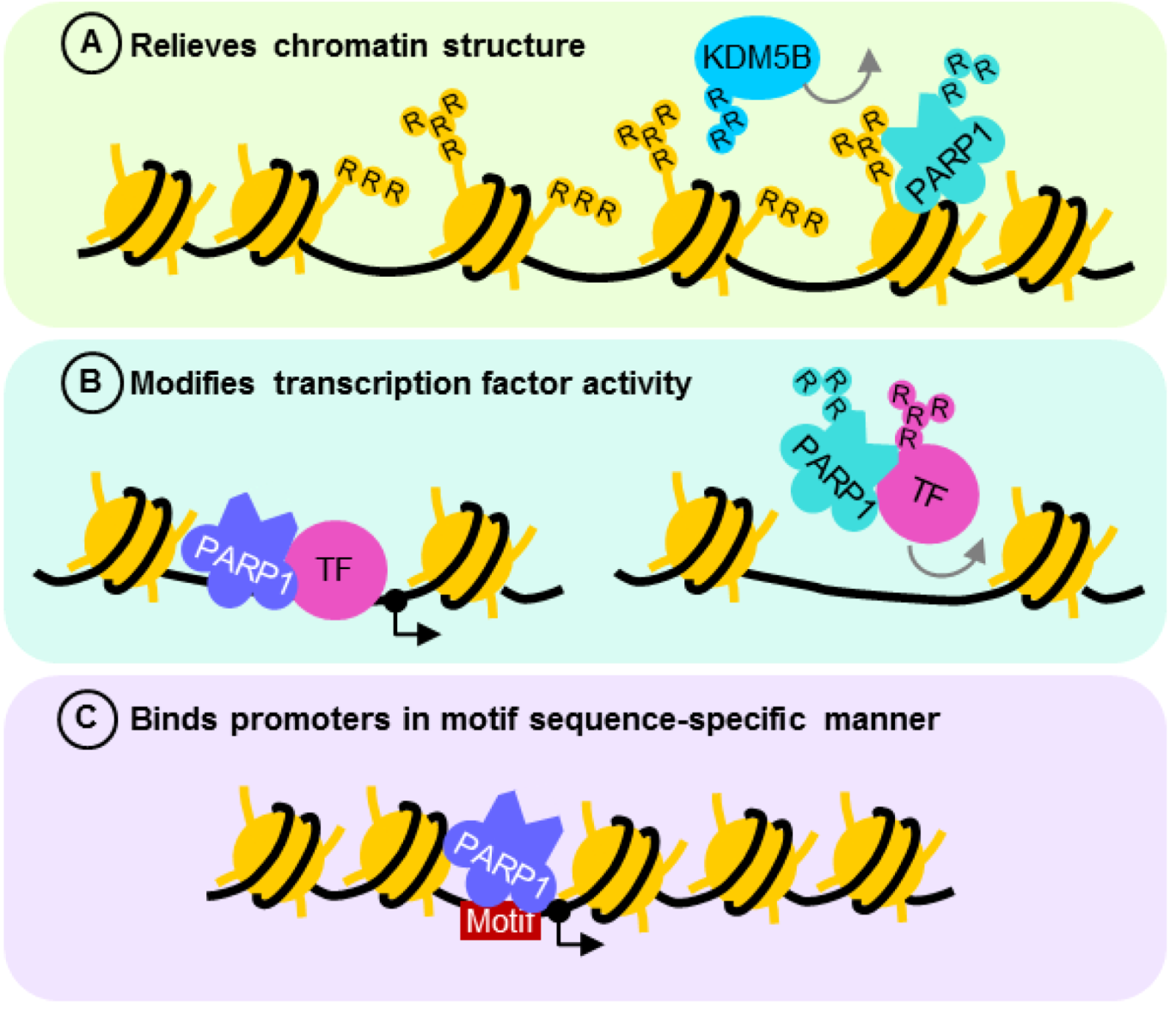
By modulating the affinity of transcription factors for their response elements and interacting partners, ADP-ribosylation acts as a molecular switch to control transcription [76,124,125,126,127] (Figure 3B). Through direct protein–protein interaction, PARP1 behaves like a coactivator or corepressor by forming a stable complex with transcription factors and its associated DNA cis element. Upon stimulation, the active PARP1 enzyme acts on its binding partner and results in complex dissociation from DNA. Transcription factors whose function is regulated by PARP1 in this manner include Oct-1 [76], SP1 [124], PPARγ [125], Smad3/Smad4 [126] and Sox2 [127], accounting for the dysregulation of multiple genes, hence the perturbation of several cellular processes in the absence of PARP1. For instance, transcription initiated by functional interaction between Smad3/Smad4 and PARP1 is disrupted when TGFβ1 signaling activates the PARP1 enzyme, attenuating Smad-dependent gene transcription for epithelial-mesenchymal transition [126]. Likewise, FGF/ERK signaling regulating embryonic stem cell differentiation relieves Sox2 interaction with its DNA responsive element by enhancing PARP1-dependent ADP-ribosylation [127].
PARP1 can also exert its effects as an inactive enzyme by directly interacting with and altering transcription factor function (Figure 3B). While ADP-ribosylated NFκB is retained in the nucleus [78], PARP1 controls its transcriptional activity [53,64,68,128,129] in an enzyme-independent manner. This is seen when acetylated PARP1 association with NFκB results in transcription of its downstream targets [64]. Sumoylation of PARP1, however, prevents p300/CBP from acetylating PARP1, hence the loss of coactivator function [53]. Surprisingly, in response to inflammatory stimulation by lipopolysaccharide, PARP1 loses its repressive function on NFκB at different sets of gene promoters when cleaved by inflammasome-dependent caspase 7, enabling transcription of pro-inflammatory genes [130]. Taken together, PARP1 is shown to be important for regulating transcription at multiple levels—from macroregulation of chromatin structure to the complex fine-tuning of gene expression which is dependent on its state of posttranslational modification.
4.2. PARP1 is a Motif-Dependent Transcription Factor
Evidence for the sequence requirement of PARP1 transcription factor function (Figure 3C) came from studies of single nucleotide polymorphisms (SNP) within promoters that alter the affinity of PARP1 for undamaged DNA. For example, a single C➔T SNP within the IFNAR1 (Interferon-α/β receptor 1) promoter reduces PARP1-dependent transcription hence confers increased susceptibility towards chronic infection from the hepatitis B virus (HBV) [27]. Likewise, increased affinity of PARP1 for the SMARCB1 promoter by a single G➔T SNP enhanced the SWI/SNF chromatin remodeling complex transcript and protein expression [26]. In agreement with phenotypes from promoter SNP variant analysis, single base substitutions within the HBV PARP1 binding motif was sufficient to abrogate transcriptional activation at the viral core promoter [23]. The effect of single base substitutions on promoter transcriptional activity concurred with the ability of PARP1 to bind the mutant DNA sequence [24], indicating that the motif through which PARP1 exerts transcriptional effects is “RNNWCAAA,” where “R” is “A” or “G,” and “W” is “A” or “T,” and “N” may be any nucleotide. Motif recognition is heavily reliant on the 3’ half of the sequence, especially at nucleotide positions 5 and 6, as their mutation abrogated transcription and PARP1 binding. This recognition motif may be readily identified in other gene promoters whose activities are also regulated by PARP1, including immune regulators interferon-γ [119] and IL-10 [123], response elements of viruses such as the human T-cell leukemia virus (HTLV) [24], as well as BRCA2 [120]. Since BRCA2 is crucial for dsDB repair by HR, the finding that its expression is regulated by PARP1 suggests another means through which PARP1 controls DNA repair. Indeed, promoters of DNA repair genes often contain PARP1 binding motifs within 3kb upstream of the transcription start site (Table 2), many of which have important functions in HR.
| DNA repair mechanism | Gene | Gene function [References] | PARP1 motif |
|---|---|---|---|
| Double-strand break repair by homologous recombination (HR) | BRCA1 | E3 ubiquitin ligase with multiple roles including controlling DNA damage signaling [131] | GAAACAAA |
| BRCA2# | Mediates recombination [132,133] | GGTACAAA | |
| BRIP1 | Interacts with BRCA1 [131] | AGTTCAAA | |
| OBFC2B | SOSS complex component; ATM signaling [134] | GCGACAAA | |
| SSBIP1 | SOSS complex component; ATM signaling [134] | GAGACAAA | |
| TOPBP1 | Stalled replication forks; ATR signaling [135] | ATTTCAAA | |
| NSMCE2 | E3 SUMO ligase of SMC5-SMC6 complex [136] | GGATCAAA | |
| SLX1B | SLX1-SLX4 resolvase catalytic subunit [137,138] | AGGACAAA | |
| DMC1 | Meiosis-specific recombinase; Interacts with BRCA2 [132,139] | AGAACAAA | |
| Base excision repair (BER) | NEIL3 | DNA glycosylase [140] | AGCTCAAA |
| MBD4^ | DNA glycosylase specific for G:T or G:U mismatches within CpG islands [141,142] | ACAACAAA | |
| Nucleotide excision repair (NER) | CETN2 | Component of XPC complex [143] | GAGACAAA |
| Mismatch repair (MMR) | MSH6 | Component of MMR [91] | GGGTCAAA |
| Direct base reversal | ALKBH3 | Oxidative demethylation of alkylated DNA [144,145] | GCCACAAA |
| Interstrand crosslink repair (ICL) | FANCG | Component of FA core complex [146] | ACTACAAA |
| DNA repair accessory proteins | RPA1 | Stabilize single-strand DNA intermediates | GTGACAAA |
| DNA polymerases | POLA2 | Subunit of primase complex | GCTACAAA |
| POLD3 | DNA polymerase δ subunit | ACTTCAAA |
5. Dysregulated PARP1 ADP-Ribosylation and Transcription Activities
Although PARP1 is heavily involved in DNA repair and transcriptional regulation, PARP1 activation at sites of DNA damage favors repair by shutting down transcription, recruiting polycomb and NuRD complexes that convert chromatin to its transcriptionally repressed state [147]. Conversely, as opposed to binding DNA strand breaks, sequence-specific motif binding suppresses the nuclear enzymatic activity of PARP1, reducing ADP-ribosylation on histone H1 and compromising cellular DNA repair [23]. However, the mechanism discriminating between intact binding motifs and damaged DNA remains unknown. Under physiological conditions, PARP1 ADP-ribosylation activity curiously follows the rhythmic circadian cycle [148]. The mechanism governing oscillating PARP1 enzymatic activity is not known, although in mice, autoregulatory loops with PARP1 acting as a transcriptional repressor at its own promoter may help to achieve this [149]. Rhythmic cycling of PARP1 ADP-ribosylation activity predicts for temporally compartmentalized DNA repair and transcription factor functions, where the efficacies of each is conversely related and differs throughout the day in a circadian rhythm-dependent manner (Figure 4). This intricate balance of PARP1 functions, however, may be perturbed in disease states such as cancer and inflammation, or by external agents such as small molecule PARP inhibitors and viruses, favoring one function over the other. The hepatitis B virus (HBV) provides a model to demonstrate this, whereby the utilization of motif–PARP1 interaction to drive viral replication suppresses PARP1 ADP-ribosylation activity hence compromises cellular DNA repair [23].
Figure 4.
PARP1 DNA repair and motif-dependent transcription is intricately regulated and possibly temporally compartmentalized by the circadian rhythm. Disturbances to this may favor one function over the other, and arise from diseases such as cancer and inflammatory disorders, as well as the addition of small molecule inhibitors and utilization of PARP1 for viral replication.
Figure 4.
PARP1 DNA repair and motif-dependent transcription is intricately regulated and possibly temporally compartmentalized by the circadian rhythm. Disturbances to this may favor one function over the other, and arise from diseases such as cancer and inflammatory disorders, as well as the addition of small molecule inhibitors and utilization of PARP1 for viral replication.

5.1. PARP1 Inhibition, Enzymatic Hyper-Activation and Disease
As the major contributor of ADP-ribosylation activities in the cell, PARP1 receives lots of attention for its enzymatic hyperactivation in various diseases. In particular, the reliance on it for initiating multiple DNA repair pathways, especially BER, HR and NHEJ, is exploited by cancer cells to safeguard against cell death induced by the accumulation of cytotoxic DNA lesions. As such, PARP1 expression is often elevated and its ADP-ribosylation activity increased in cancerous tissues [150,151], rendering it a good candidate for sensitizing cancer cells to the cytotoxic effects of DNA damaging agents [8,9,11,12,13,14,15,16,17]. Synthetic lethality may be achieved with PARP inhibitors designed to compete with NAD+ for the PARP1 catalytic site, and these small molecules have produced promising results in clinical trials for the treatment of several cancers. Further evidence for synthetic lethality was obtained in mice bearing BRCA1-deficient ovarian cancer cells, where life expectancy was significantly extended by delivery of nanoparticles containing siRNA targeting PARP1 [152]. PARP1 is also frequently implicated in inflammatory disorders such as sepsis, diabetes, myocardial infarction and stroke [8,9,11,12,15,16,109], as considerable by-stander DNA damage resulting from the generation of reactive oxygen species hyperactivates PARP1. Large amounts of NAD+ consumed in the process can also result in necrosis, aggravating the inflammatory condition. Under such circumstances, treatment with PARP1 inhibitors can provide symptomatic relief in animal models for such diseases [8,9,11,12,13,14,15,16,17,59,109], raising hope for clinical efficacy in the near future.
The ability of PARP1 to function without its enzymatic activity is often underappreciated. Indeed, many diseases involving PARP1 manifest independently of its ADP-ribosylation activity, and arise from dysregulated expression of molecules because of altered PARP1 affinity for its recognition motif in susceptibility genetic loci. Sequence-dependent PARP1 binding for SNP variants within promoter elements of the cytokine IL-10 [123], chemokine CCL2 [30], interferon α/β receptor 1 (IFNAR1) [27] and SMARCB1 [26], have all been associated with systemic lupus erythematosus (SLE), carotid atherosclerosis, chronic infection with HBV and acute lymphoblastic lymphoma (ALL), respectively. The effect of PARP1 inhibition on the outcome of such diseases, however, has not been evaluated. Importantly, because both PARP1 DNA repair and transcription factor functions may be implicated, the outcome of enzymatic inhibition in certain diseases need not be immediately conclusive. In diabetes, however, PARP inhibitors may be therapeutically beneficial as inhibiting ADP-ribosylation suppresses inflammation and concurrently enhances PARP1 transcription activator function at the Reg promoter for β-cell regeneration [29]. Interestingly, PARP1 inhibitors have gender-specific effects on animal disease models [153]. Whether this may also be true in the clinical setting remains to be seen.
5.2. Oncogenic Viruses—Inhibiting the PARP1 Enzyme to Enhance Viral Replication
The balance of PARP1 functions between enzyme-independent transcription and enzyme-dependent DNA repair is easily tipped by external stimuli. HBV is one external agent which deprives the infected host cell of PARP1 activity for its efficient replication whilst preventing its function in DNA repair, as the PARP1 binding motif “ACTTCAAA” carried within its genome is readily recognizable for transcriptional activation [23]. This may contribute to the oncogenic properties of HBV, especially in carriers of the virus with high HBV DNA loads [154,155,156]. By utilizing PARP1 to increase replication efficiency, the large amounts of viral DNA produced act as template to support further viral replication while concurrently inhibiting PARP1 ADP-ribosylation, reducing DNA repair capacity of the infected host cell. Accumulation of sublethal DNA lesions with prolonged infection [157] thus increases the risk of developing hepatocellular carcinoma (HCC). Several oncogenic viruses also possess high affinity PARP1 binding motifs in their genome (Table 3), suggesting that they may act on PARP1 in a similar manner to increase host risk for developing cancer. Viral DNA-PARP1 interaction has already been shown to be important for enhancing replication of the human T-cell leukemia virus (HTLV) [24] and the Kaposi’s sarcoma-associated virus (KSHV) [158,159]. Since these studies also show that PARP1 enzymatic inhibitors positively regulate viral replication, the use of small molecules targeting only the PARP1 catalytic domain may be contraindicated in many patients with such viral infections. Given the potential benefits of PARP inhibitors for the treatment of multiple diseases, it is perhaps worthwhile to invest in novel strategies that can overcome the pitfalls of aggravating viral replication and altered transcription factor function of current PARP inhibitors.
| Oncogenic virus | Gene or DNA element | Motif |
|---|---|---|
| Human herpesvirus 4 (EBV) | OriLyt replication origin | ACTTCAAA |
| Hepatitis B Virus (HBV) | Core promoter | ACTTCAAA |
| Human T-cell leukemia virus (HTLV) | Tax responsive element | ACGACAAC |
| Human herpesvirus 8 (KSHV) | ORF4 complement control protein | GCTACAAA |
| Primase | ACGTCAAA | |
| Merkel cell polyomavirus | VP3 capsid protein | ACTTCAAA |
6. Conclusions
PARP1 plays important roles in both DNA repair and transcription, and the interplay of these processes in relation to cellular function and diseases states have not been well defined. As PARP1 binding motifs may be readily found in promoter elements of DNA repair genes, the expanding role of PARP1 in DNA repair need not be independent of transcription. Moreover, since PARP1 ADP-ribosylation is very important for DNA repair and transcription, yet the PAR-independent mechanism of transcriptional regulation through specific binding of PARP1 at its recognition motif also exists, rhythmic cycling of PARP1 enzyme activity suggests that these processes are unlikely to occur optimally together. The suppression of DNA repair by motif-dependent replication of oncogenic viruses illustrates this possibility. Thus, there is a need to better understand the effect of PARP1 inhibition in the therapeutic context and its effect on cellular transcription.
Conflict of Interest
The authors declare no conflict of interest.
Acknowledgments
This work was funded by the Agency for Science, Technology and Research (A*STAR), Singapore.
References
- Satoh, M.S.; Poirier, G.G.; Lindahl, T. Dual function for poly(ADP-ribose) synthesis in response to DNA strand breakage. Biochemistry 1994, 33, 7099–7106. [Google Scholar]
- Langelier, M.F.; Planck, J.L.; Roy, S.; Pascal, J.M. Crystal structures of poly(ADP-ribose) polymerase-1 (PARP-1) zinc fingers bound to DNA: Structural and functional insights into DNA-dependent PARP-1 activity. J. Biol. Chem. 2011, 286, 10690–10701. [Google Scholar]
- Langelier, M.F.; Planck, J.L.; Roy, S.; Pascal, J.M. Structural basis for DNA damage-dependent poly(ADP-ribosyl)ation by human PARP-1. Science 2012, 336, 728–732. [Google Scholar] [CrossRef]
- Eustermann, S.; Videler, H.; Yang, J.C.; Cole, P.T.; Gruszka, D.; Veprintsev, D.; Neuhaus, D. The DNA-binding domain of human PARP-1 interacts with DNA single-strand breaks as a monomer through its second zinc finger. J. Mol. Biol. 2011, 407, 149–170. [Google Scholar] [CrossRef]
- Lindahl, T.; Satoh, M.S.; Poirier, G.G.; Klungland, A. Post-translational modification of poly(adp-ribose) polymerase induced by DNA strand breaks. Trends Biochem. Sci. 1995, 20, 405–411. [Google Scholar] [CrossRef]
- D'Amours, D.; Desnoyers, S.; D'Silva, I.; Poirier, G.G. Poly(adp-ribosyl)ation reactions in the regulation of nuclear functions. Biochem. J. 1999, 342 (Pt 2), 249–268. [Google Scholar]
- De Vos, M.; Schreiber, V.; Dantzer, F. The diverse roles and clinical relevance of PARPs in DNA damage repair: Current state of the art. Biochem. Pharmacol. 2012, 84, 137–146. [Google Scholar]
- Jagtap, P.; Szabo, C. Poly(ADP-ribose) polymerase and the therapeutic effects of its inhibitors. Nat. Rev. Drug Discov. 2005, 4, 421–440. [Google Scholar] [CrossRef]
- Rouleau, M.; Patel, A.; Hendzel, M.J.; Kaufmann, S.H.; Poirier, G.G. PARP inhibition: PARP1 and beyond. Nat. Rev. Cancer 2010, 10, 293–301. [Google Scholar] [CrossRef]
- Shall, S.; de Murcia, G. Poly(ADP-ribose) polymerase-1: What have we learned from the deficient mouse model? Mutat. Res. 2000, 460, 1–15. [Google Scholar] [CrossRef]
- Masutani, M.; Nakagama, H.; Sugimura, T. Poly(ADP-ribosyl)ation in relation to cancer and autoimmune disease. Cell. Mol. Life Sci. 2005, 62, 769–783. [Google Scholar] [CrossRef]
- Szabo, C.; Pacher, P.; Swanson, R.A. Novel modulators of poly(ADP-ribose) polymerase. Trends Pharmacol. Sci. 2006, 27, 626–630. [Google Scholar] [CrossRef]
- Miwa, M.; Masutani, M. PolyADP-ribosylation and cancer. Cancer Sci. 2007, 98, 1528–1535. [Google Scholar] [CrossRef]
- Kling, J. PARP inhibitors blaze a trail in difficult-to-treat cancers. Nat. Biotechnol. 2009, 27, 784–786. [Google Scholar] [CrossRef]
- Peralta-Leal, A.; Rodriguez-Vargas, J.M.; Aguilar-Quesada, R.; Rodriguez, M.I.; Linares, J.L.; de Almodovar, M.R.; Oliver, F.J. PARP inhibitors: New partners in the therapy of cancer and inflammatory diseases. Free Radic. Biol. Med. 2009, 47, 13–26. [Google Scholar] [CrossRef]
- Schreiber, V.; Dantzer, F.; Ame, J.C.; de Murcia, G. Poly(ADP-ribose): Novel functions for an old molecule. Nat. Rev. Mol. Cell Biol. 2006, 7, 517–528. [Google Scholar] [CrossRef]
- Plummer, E.R. Inhibition of poly(ADP-ribose) polymerase in cancer. Curr. Opin. Pharmacol. 2006, 6, 364–368. [Google Scholar] [CrossRef]
- Kim, M.Y.; Mauro, S.; Gevry, N.; Lis, J.T.; Kraus, W.L. NAD+-dependent modulation of chromatin structure and transcription by nucleosome binding properties of PARP-1. Cell 2004, 119, 803–814. [Google Scholar] [CrossRef]
- Kraus, W.L.; Lis, J.T. PARP goes transcription. Cell 2003, 113, 677–683. [Google Scholar] [CrossRef]
- Kraus, W.L. Transcriptional control by PARP-1: Chromatin modulation, enhancer-binding, coregulation, and insulation. Curr. Opin. Cell Biol. 2008, 20, 294–302. [Google Scholar] [CrossRef]
- Krishnakumar, R.; Kraus, W.L. The PARP side of the nucleus: Molecular actions, physiological outcomes, and clinical targets. Mol. Cell 2010, 39, 8–24. [Google Scholar] [CrossRef]
- Rouleau, M.; Aubin, R.A.; Poirier, G.G. Poly(ADP-ribosyl)ated chromatin domains: Access granted. J. Cell Sci. 2004, 117, 815–825. [Google Scholar] [CrossRef]
- Ko, H.L.; Ren, E.C. Novel poly (ADP-ribose) polymerase 1 binding motif in hepatitis B virus core promoter impairs DNA damage repair. Hepatology 2011, 54, 1190–1198. [Google Scholar] [CrossRef]
- Zhang, Z.; Hildebrandt, E.F.; Simbulan-Rosenthal, C.M.; Anderson, M.G. Sequence-specific binding of poly(ADP-ribose) polymerase-1 to the human T cell leukemia virus type-I tax responsive element. Virology 2002, 296, 107–116. [Google Scholar] [CrossRef]
- Ambrose, H.E.; Papadopoulou, V.; Beswick, R.W.; Wagner, S.D. Poly-(ADP-ribose) polymerase-1 (PARP-1) binds in a sequence-specific manner at the bcl-6 locus and contributes to the regulation of bcl-6 transcription. Oncogene 2007, 26, 6244–6252. [Google Scholar] [CrossRef]
- Pottier, N.; Cheok, M.H.; Yang, W.; Assem, M.; Tracey, L.; Obenauer, J.C.; Panetta, J.C.; Relling, M.V.; Evans, W.E. Expression of SMARCB1 modulates steroid sensitivity in human lymphoblastoid cells: Identification of a promoter SNP that alters PARP1 binding and SMARCB1 expression. Hum. Mol. Genet. 2007, 16, 2261–2271. [Google Scholar] [CrossRef]
- Zhou, J.; Huang, J.D.; Poon, V.K.; Chen, D.Q.; Chan, C.C.; Ng, F.; Guan, X.Y.; Watt, R.M.; Lu, L.; Yuen, K.Y., et al. Functional dissection of an IFN-alpha/beta receptor 1 promoter variant that confers higher risk to chronic hepatitis b virus infection. J. Hepatol. 2009, 51, 322–332. [Google Scholar]
- Chung, E.Y.; Liu, J.; Zhang, Y.; Ma, X. Differential expression in lupus-associated Il-10 promoter single-nucleotide polymorphisms is mediated by poly(ADP-ribose) polymerase-1. Genes Immun. 2007, 8, 577–589. [Google Scholar] [CrossRef]
- Akiyama, T.; Takasawa, S.; Nata, K.; Kobayashi, S.; Abe, M.; Shervani, N.J.; Ikeda, T.; Nakagawa, K.; Unno, M.; Matsuno, S., et al. Activation of Reg gene, a gene for insulin-producing beta-cell regeneration: Poly(ADP-ribose) polymerase binds Reg promoter and regulates the transcription by auto poly(ADP-ribosyl)ation. Proc. Natl. Acad. Sci. USA 2001, 98, 48–53. [Google Scholar]
- Nyquist, P.; Zhang, J.; De Graba, T.J. The -928 G/C and -362 G/C single-nucleotide polymorphisms in the promoter of MCP-1: Increased transcriptional activity and novel binding sites. Cerebrovasc. Dis. 2009, 29, 242–247. [Google Scholar]
- Hassa, P.O.; Haenni, S.S.; Elser, M.; Hottiger, M.O. Nuclear ADP-ribosylation reactions in mammalian cells: Where are we today and where are we going? Microbiol. Mol. Biol. Rev. 2006, 70, 789–829. [Google Scholar] [CrossRef]
- Pion, E.; Ullmann, G.M.; Ame, J.C.; Gerard, D.; de Murcia, G.; Bombarda, E. DNA-induced dimerization of poly(ADP-ribose) polymerase-1 triggers its activation. Biochemistry 2005, 44, 14670–14681. [Google Scholar]
- Petrucco, S.; Percudani, R. Structural recognition of DNA by poly(ADP-ribose)polymerase-like zinc finger families. FEBS J. 2008, 275, 883–893. [Google Scholar] [CrossRef]
- Langelier, M.F.; Ruhl, D.D.; Planck, J.L.; Kraus, W.L.; Pascal, J.M. The Zn3 domain of human poly(ADP-ribose) polymerase-1 (PARP-1) functions in both DNA-dependent poly(ADP-ribose) synthesis activity and chromatin compaction. J. Biol. Chem. 2010, 285, 18877–18887. [Google Scholar]
- Gagne, J.P.; Rouleau, M.; Poirier, G.G. Structural biology. PARP-1 activation--bringing the pieces together. Science 2010, 336, 678–679. [Google Scholar]
- Luo, X.; Kraus, W.L. On par with parp: Cellular stress signaling through poly(ADP-ribose) and PARP-1. Genes Dev. 2012, 26, 417–432. [Google Scholar] [CrossRef]
- Hong, S.J.; Dawson, T.M.; Dawson, V.L. Nuclear and mitochondrial conversations in cell death: PARP-1 and AIF signaling. Trends Pharmacol. Sci. 2004, 25, 259–264. [Google Scholar] [CrossRef]
- Huber, A.; Bai, P.; de Murcia, J.M.; de Murcia, G. PARP-1, PARP-2 and ATM in the DNA damage response: Functional synergy in mouse development. DNA Repair (Amst) 2004, 3, 1103–1108. [Google Scholar]
- Kim, M.Y.; Zhang, T.; Kraus, W.L. Poly(ADP-ribosyl)ation by PARP-1: 'Par-laying' NAD+ into a nuclear signal. Genes Dev. 2005, 19, 1951–1967. [Google Scholar] [CrossRef]
- Koh, D.W.; Dawson, T.M.; Dawson, V.L. Mediation of cell death by poly(ADP-ribose) polymerase-1. Pharmacol. Res. 2005, 52, 5–14. [Google Scholar] [CrossRef]
- Heeres, J.T.; Hergenrother, P.J. Poly(ADP-ribose) makes a date with death. Curr. Opin. Chem. Biol. 2007, 11, 644–653. [Google Scholar] [CrossRef]
- Hakme, A.; Wong, H.K.; Dantzer, F.; Schreiber, V. The expanding field of poly(ADP-ribosyl)ation reactions. 'Protein modifications: Beyond the usual suspects' review series. EMBO Rep. 2008, 9, 1094–1100. [Google Scholar]
- Ogata, N.; Ueda, K.; Kawaichi, M.; Hayaishi, O. Poly(ADP-ribose) synthetase, a main acceptor of poly(ADP-ribose) in isolated nuclei. J. Biol. Chem. 1981, 256, 4135–4137. [Google Scholar]
- Ame, J.C.; Rolli, V.; Schreiber, V.; Niedergang, C.; Apiou, F.; Decker, P.; Muller, S.; Hoger, T.; Menissier-de Murcia, J.; de Murcia, G. Parp-2, a novel mammalian DNA damage-dependent poly(ADP-ribose) polymerase. J. Biol. Chem. 1999, 274, 17860–17868. [Google Scholar]
- Koh, D.W.; Lawler, A.M.; Poitras, M.F.; Sasaki, M.; Wattler, S.; Nehls, M.C.; Stoger, T.; Poirier, G.G.; Dawson, V.L.; Dawson, T.M. Failure to degrade poly(ADP-ribose) causes increased sensitivity to cytotoxicity and early embryonic lethality. Proc. Natl. Acad. Sci. USA 2004, 101, 17699–17704. [Google Scholar]
- Loseva, O.; Jemth, A.S.; Bryant, H.E.; Schuler, H.; Lehtio, L.; Karlberg, T.; Helleday, T. PARP-3 is a mono-ADP-ribosylase that activates PARP-1 in the absence of DNA. J. Biol. Chem. 2010, 285, 8054–8060. [Google Scholar]
- Mao, Z.; Hine, C.; Tian, X.; Meter, M. V.; Au, M.; Vaidya, A.; Seluanov, A.; Gorbunova, V. SIRT6 promotes DNA repair under stress by activating PARP1. Science 2011, 332, 1443–1446. [Google Scholar] [CrossRef]
- Kauppinen, T.M.; Chan, W.Y.; Suh, S.W.; Wiggins, A.K.; Huang, E.J.; Swanson, R.A. Direct phosphorylation and regulation of poly(ADP-ribose) polymerase-1 by extracellular signal-regulated kinases 1/2. Proc. Natl. Acad. Sci. USA 2006, 103, 7136–7141. [Google Scholar]
- Cohen-Armon, M.; Visochek, L.; Rozensal, D.; Kalal, A.; Geistrikh, I.; Klein, R.; Bendetz-Nezer, S.; Yao, Z.; Seger, R. DNA-independent PARP-1 activation by phosphorylated ERK2 increases ELK1 activity: A link to histone acetylation. Mol. Cell 2007, 25, 297–308. [Google Scholar] [CrossRef]
- Cohen-Armon, M. PARP-1 activation in the ERK signaling pathway. Trends Pharmacol. Sci. 2007, 28, 556–560. [Google Scholar] [CrossRef]
- Ju, B.G.; Solum, D.; Song, E.J.; Lee, K.J.; Rose, D.W.; Glass, C.K.; Rosenfeld, M.G. Activating the PARP-1 sensor component of the groucho/ TLE1 corepressor complex mediates a camkinase II delta-dependent neurogenic gene activation pathway. Cell 2004, 119, 815–829. [Google Scholar] [CrossRef]
- Dong, F.; Soubeyrand, S.; Hache, R.J. Activation of PARP-1 in response to bleomycin depends on the Ku antigen and protein phosphatase 5. Oncogene 2010, 29, 2093–2103. [Google Scholar] [CrossRef]
- Messner, S.; Schuermann, D.; Altmeyer, M.; Kassner, I.; Schmidt, D.; Schar, P.; Muller, S.; Hottiger, M.O. Sumoylation of poly(ADP-ribose) polymerase 1 inhibits its acetylation and restrains transcriptional coactivator function. FASEB J. 2009, 23, 3978–3989. [Google Scholar] [CrossRef] [Green Version]
- Berger, F.; Lau, C.; Ziegler, M. Regulation of poly(ADP-ribose) polymerase 1 activity by the phosphorylation state of the nuclear nad biosynthetic enzyme NMN adenylyl transferase 1. Proc. Natl. Acad. Sci. USA 2007, 104, 3765–3770. [Google Scholar]
- Ouararhni, K.; Hadj-Slimane, R.; Ait-Si-Ali, S.; Robin, P.; Mietton, F.; Harel-Bellan, A.; Dimitrov, S.; Hamiche, A. The histone variant mH2A1.1 interferes with transcription by down-regulating PARP-1 enzymatic activity. Genes Dev. 2006, 20, 3324–3336. [Google Scholar]
- Midorikawa, R.; Takei, Y.; Hirokawa, N. KIF4 motor regulates activity-dependent neuronal survival by suppressing PARP-1 enzymatic activity. Cell 2006, 125, 371–383. [Google Scholar] [CrossRef]
- Kaufmann, S.H.; Desnoyers, S.; Ottaviano, Y.; Davidson, N.E.; Poirier, G.G. Specific proteolytic cleavage of poly(ADP-ribose) polymerase: An early marker of chemotherapy-induced apoptosis. Cancer Res. 1993, 53, 3976–3985. [Google Scholar]
- Nicholson, D.W.; Ali, A.; Thornberry, N.A.; Vaillancourt, J.P.; Ding, C.K.; Gallant, M.; Gareau, Y.; Griffin, P.R.; Labelle, M.; Lazebnik, Y.A., et al. Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature 1995, 376, 37–43. [Google Scholar]
- Erdelyi, K.; Bakondi, E.; Gergely, P.; Szabo, C.; Virag, L. Pathophysiologic role of oxidative stress-induced poly(ADP-ribose) polymerase-1 activation: Focus on cell death and transcriptional regulation. Cell. Mol. Life Sci. 2005, 62, 751–759. [Google Scholar] [CrossRef]
- Chevanne, M.; Calia, C.; Zampieri, M.; Cecchinelli, B.; Caldini, R.; Monti, D.; Bucci, L.; Franceschi, C.; Caiafa, P. Oxidative DNA damage repair and PARP 1 and PARP 2 expression in epstein-barr virus-immortalized b lymphocyte cells from young subjects, old subjects, and centenarians. Rejuvenation Res. 2007, 10, 191–204. [Google Scholar] [CrossRef]
- Chaitanya, G.V.; Babu, P.P. Differential parp cleavage: An indication of heterogeneous forms of cell death and involvement of multiple proteases in the infarct of focal cerebral ischemia in rat. Cell. Mol. Neurobiol. 2009, 29, 563–573. [Google Scholar] [CrossRef]
- McGinnis, K.M.; Gnegy, M.E.; Park, Y.H.; Mukerjee, N.; Wang, K.K. Procaspase-3 and poly(ADP)ribose polymerase (PARP) are calpain substrates. Biochem. Biophys. Res. Commun. 1999, 263, 94–99. [Google Scholar] [CrossRef]
- Gobeil, S.; Boucher, C.C.; Nadeau, D.; Poirier, G.G. Characterization of the necrotic cleavage of poly(ADP-ribose) polymerase (PARP-1): Implication of lysosomal proteases. Cell Death Differ. 2001, 8, 588–594. [Google Scholar] [CrossRef]
- Hassa, P.O.; Haenni, S.S.; Buerki, C.; Meier, N.I.; Lane, W.S.; Owen, H.; Gersbach, M.; Imhof, R.; Hottiger, M.O. Acetylation of poly(ADP-ribose) polymerase-1 by p300/creb-binding protein regulates coactivation of NF-kappaB-dependent transcription. J. Biol. Chem. 2005, 280, 40450–40464. [Google Scholar]
- Ju, B.G.; Lunyak, V.V.; Perissi, V.; Garcia-Bassets, I.; Rose, D.W.; Glass, C.K.; Rosenfeld, M.G. A topoisomerase II beta-mediated dsdna break required for regulated transcription. Science 2006, 312, 1798–1802. [Google Scholar] [CrossRef]
- Bryant, H.E.; Petermann, E.; Schultz, N.; Jemth, A.S.; Loseva, O.; Issaeva, N.; Johansson, F.; Fernandez, S.; McGlynn, P.; Helleday, T. PARP is activated at stalled forks to mediate Mre11-dependent replication restart and recombination. EMBO J. 2009, 28, 2601–2615. [Google Scholar] [CrossRef]
- Gottschalk, A.J.; Timinszky, G.; Kong, S.E.; Jin, J.; Cai, Y.; Swanson, S.K.; Washburn, M.P.; Florens, L.; Ladurner, A.G.; Conaway, J.W., et al. Poly(ADP-ribosyl)ation directs recruitment and activation of an ATP-dependent chromatin remodeler. Proc. Natl. Acad. Sci. USA 2009, 106, 13770–13774. [Google Scholar]
- Stilmann, M.; Hinz, M.; Arslan, S.C.; Zimmer, A.; Schreiber, V.; Scheidereit, C. A nuclear poly(ADP-ribose)-dependent signalosome confers DNA damage-induced Ikappab kinase activation. Mol. Cell 2009, 36, 365–378. [Google Scholar] [CrossRef]
- Krishnakumar, R.; Kraus, W.L. PARP-1 regulates chromatin structure and transcription through a KDM5B-dependent pathway. Mol. Cell 2010, 39, 736–749. [Google Scholar] [CrossRef]
- Krishnakumar, R.; Gamble, M.J.; Frizzell, K.M.; Berrocal, J.G.; Kininis, M.; Kraus, W.L. Reciprocal binding of PARP-1 and histone H1 at promoters specifies transcriptional outcomes. Science 2008, 319, 819–821. [Google Scholar]
- Timinszky, G.; Till, S.; Hassa, P.O.; Hothorn, M.; Kustatscher, G.; Nijmeijer, B.; Colombelli, J.; Altmeyer, M.; Stelzer, E.H.; Scheffzek, K., et al. A macrodomain-containing histone rearranges chromatin upon sensing PARP1 activation. Nat. Struct. Mol. Biol. 2009, 16, 923–929. [Google Scholar]
- Masson, M.; Niedergang, C.; Schreiber, V.; Muller, S.; Menissier-de Murcia, J.; de Murcia, G. XRCC1 is specifically associated with poly(ADP-ribose) polymerase and negatively regulates its activity following DNA damage. Mol. Cell. Biol. 1998, 18, 3563–3571. [Google Scholar]
- Ahel, D.; Horejsi, Z.; Wiechens, N.; Polo, S.E.; Garcia-Wilson, E.; Ahel, I.; Flynn, H.; Skehel, M.; West, S.C.; Jackson, S.P., et al. Poly(ADP-ribose)-dependent regulation of DNA repair by the chromatin remodeling enzyme ALC1. Science 2009, 325, 1240–1243. [Google Scholar] [CrossRef]
- Haince, J.F.; Kozlov, S.; Dawson, V.L.; Dawson, T.M.; Hendzel, M.J.; Lavin, M.F.; Poirier, G.G. Ataxia telangiectasia mutated (ATM) signaling network is modulated by a novel poly(ADP-ribose)-dependent pathway in the early response to DNA-damaging agents. J. Biol. Chem. 2007, 282, 16441–16453. [Google Scholar]
- Smith, S. The world according to PARP. Trends Biochem. Sci. 2001, 26, 174–179. [Google Scholar] [CrossRef]
- Nie, J.; Sakamoto, S.; Song, D.; Qu, Z.; Ota, K.; Taniguchi, T. Interaction of oct-1 and automodification domain of poly(ADP-ribose) synthetase. FEBS Lett. 1998, 424, 27–32. [Google Scholar] [CrossRef]
- Gagne, J.P.; Hunter, J.M.; Labrecque, B.; Chabot, B.; Poirier, G.G. A proteomic approach to the identification of heterogeneous nuclear ribonucleoproteins as a new family of poly(ADP-ribose)-binding proteins. Biochem. J. 2003, 371, 331–340. [Google Scholar] [CrossRef]
- Zerfaoui, M.; Errami, Y.; Naura, A.S.; Suzuki, Y.; Kim, H.; Ju, J.; Liu, T.; Hans, C.P.; Kim, J.G.; Abd Elmageed, Z.Y., et al. Poly(ADP-ribose) polymerase-1 is a determining factor in crm1-mediated nuclear export and retention of p65 NF-kappa B upon TLR4 stimulation. J. Immunol. 2010, 185, 1894–1902. [Google Scholar]
- Kanai, M.; Hanashiro, K.; Kim, S.H.; Hanai, S.; Boulares, A.H.; Miwa, M.; Fukasawa, K. Inhibition of Crm1-p53 interaction and nuclear export of p53 by poly(ADP-ribosyl)ation. Nat. Cell. Biol. 2007, 9, 1175–1183. [Google Scholar] [CrossRef]
- Shibata, A.; Kamada, N.; Masumura, K.; Nohmi, T.; Kobayashi, S.; Teraoka, H.; Nakagama, H.; Sugimura, T.; Suzuki, H.; Masutani, M. PARP-1 deficiency causes an increase of deletion mutations and insertions/rearrangements in vivo after treatment with an alkylating agent. Oncogene 2005, 24, 1328–1337. [Google Scholar] [CrossRef]
- Shibata, A.; Maeda, D.; Ogino, H.; Tsutsumi, M.; Nohmi, T.; Nakagama, H.; Sugimura, T.; Teraoka, H.; Masutani, M. Role of PARP-1 in suppressing spontaneous deletion mutation in the liver and brain of mice at adolescence and advanced age. Mutat. Res. 2009, 664, 20–27. [Google Scholar] [CrossRef]
- Tong, W.M.; Cortes, U.; Hande, M.P.; Ohgaki, H.; Cavalli, L.R.; Lansdorp, P.M.; Haddad, B.R.; Wang, Z.Q. Synergistic role of Ku80 and poly(ADP-ribose) polymerase in suppressing chromosomal aberrations and liver cancer formation. Cancer Res. 2002, 62, 6990–6996. [Google Scholar]
- Tong, W.M.; Yang, Y.G.; Cao, W.H.; Galendo, D.; Frappart, L.; Shen, Y.; Wang, Z.Q. Poly(ADP-ribose) polymerase-1 plays a role in suppressing mammary tumourigenesis in mice. Oncogene 2007, 26, 3857–3867. [Google Scholar] [CrossRef]
- Wang, X.; Liu, L.; Montagna, C.; Ried, T.; Deng, C.X. Haploinsufficiency of PARP1 accelerates BRCA1-associated centrosome amplification, telomere shortening, genetic instability, apoptosis, and embryonic lethality. Cell Death Differ. 2007, 14, 924–931. [Google Scholar]
- Sugo, N.; Niimi, N.; Aratani, Y.; Masutani, M.; Suzuki, H.; Koyama, H. Decreased PARP-1 levels accelerate embryonic lethality but attenuate neuronal apoptosis in DNA polymerase beta-deficient mice. Biochem. Biophys. Res. Commun. 2007, 354, 656–661. [Google Scholar] [CrossRef]
- Henning, S.M.; Swendseid, M.E.; Coulson, W.F. Male rats fed methyl- and folate-deficient diets with or without niacin develop hepatic carcinomas associated with decreased tissue nad concentrations and altered poly(ADP-ribose) polymerase activity. J. Nutr. 1997, 127, 30–36. [Google Scholar]
- Khodyreva, S.N.; Prasad, R.; Ilina, E.S.; Sukhanova, M.V.; Kutuzov, M.M.; Liu, Y.; Hou, E.W.; Wilson, S.H.; Lavrik, O.I. Apurinic/apyrimidinic (AP) site recognition by the 5'-dRP/AP lyase in poly(ADP-ribose) polymerase-1 (PARP-1). Proc. Natl. Acad. Sci. USA 2010, 107, 22090–22095. [Google Scholar]
- Hegde, M.L.; Hazra, T.K.; Mitra, S. Early steps in the DNA base excision/single-strand interruption repair pathway in mammalian cells. Cell Res. 2008, 18, 27–47. [Google Scholar] [CrossRef]
- Pleschke, J.M.; Kleczkowska, H.E.; Strohm, M.; Althaus, F.R. Poly(ADP-ribose) binds to specific domains in DNA damage checkpoint proteins. J. Biol. Chem. 2000, 275, 40974–40980. [Google Scholar]
- Gagne, J.P.; Isabelle, M.; Lo, K.S.; Bourassa, S.; Hendzel, M.J.; Dawson, V.L.; Dawson, T.M.; Poirier, G.G. Proteome-wide identification of poly(ADP-ribose) binding proteins and poly(ADP-ribose)-associated protein complexes. Nucleic Acids Res. 2008, 36, 6959–6976. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.; Calvo, J.A.; Samson, L.D. Balancing repair and tolerance of DNA damage caused by alkylating agents. Nat. Rev. Cancer 2012, 12, 104–120. [Google Scholar]
- Dantzer, F.; de La Rubia, G.; Menissier-De Murcia, J.; Hostomsky, Z.; de Murcia, G.; Schreiber, V. Base excision repair is impaired in mammalian cells lacking poly(ADP-ribose) polymerase-1. Biochemistry 2000, 39, 7559–7569. [Google Scholar]
- Friedberg, E.C. How nucleotide excision repair protects against cancer. Nat. Rev. Cancer 2001, 1, 22–33. [Google Scholar] [CrossRef]
- Ambrose, H.E.; Willimott, S.; Beswick, R.W.; Dantzer, F.; de Murcia, J.M.; Yelamos, J.; Wagner, S.D. Poly(ADP-ribose) polymerase-1 (PARP-1)-deficient mice demonstrate abnormal antibody responses. Immunology 2009, 127, 178–186. [Google Scholar] [CrossRef]
- Shrivastav, M.; De Haro, L.P.; Nickoloff, J.A. Regulation of DNA double-strand break repair pathway choice. Cell Res. 2008, 18, 134–147. [Google Scholar] [CrossRef]
- Paddock, M.N.; Bauman, A.T.; Higdon, R.; Kolker, E.; Takeda, S.; Scharenberg, A.M. Competition between PARP-1 and Ku70 control the decision between high-fidelity and mutagenic DNA repair. DNA Repair (Amst) 2011, 10, 338–343. [Google Scholar]
- Mansour, W.Y.; Rhein, T.; Dahm-Daphi, J. The alternative end-joining pathway for repair of DNA double-strand breaks requires PARP1 but is not dependent upon microhomologies. Nucleic Acids Res. 2010, 38, 6065–6077. [Google Scholar] [CrossRef]
- Hochegger, H.; Dejsuphong, D.; Fukushima, T.; Morrison, C.; Sonoda, E.; Schreiber, V.; Zhao, G.Y.; Saberi, A.; Masutani, M.; Adachi, N., et al. PARP-1 protects homologous recombination from interference by ku and ligase iv in vertebrate cells. EMBO J. 2006, 25, 1305–1314. [Google Scholar] [CrossRef]
- San Filippo, J.; Sung, P.; Klein, H. Mechanism of eukaryotic homologous recombination. Annu. Rev. Biochem. 2008, 77, 229–257. [Google Scholar] [CrossRef]
- Zhou, B.B.; Bartek, J. Targeting the checkpoint kinases: Chemosensitization versus chemoprotection. Nat. Rev. Cancer 2004, 4, 216–225. [Google Scholar] [CrossRef]
- Shiloh, Y. ATM and related protein kinases: Safeguarding genome integrity. Nat. Rev. Cancer 2003, 3, 155–168. [Google Scholar] [CrossRef]
- Lobrich, M.; Jeggo, P.A. The impact of a negligent G2/M checkpoint on genomic instability and cancer induction. Nat. Rev. Cancer 2007, 7, 861–869. [Google Scholar] [CrossRef]
- Cimprich, K.A.; Cortez, D. ATR: An essential regulator of genome integrity. Nat. Rev. Mol. Cell Biol. 2008, 9, 616–627. [Google Scholar]
- Nam, E.A.; Cortez, D. ATR signalling: More than meeting at the fork. Biochem. J 2011, 436, 527–536. [Google Scholar] [CrossRef]
- Lieber, M.R.; Wilson, T.E. Snapshot: Nonhomologous DNA end joining (NHEJ). Cell 2010, 142. 496–496 e491. [Google Scholar]
- Ruscetti, T.; Lehnert, B.E.; Halbrook, J.; Le Trong, H.; Hoekstra, M.F.; Chen, D.J.; Peterson, S.R. Stimulation of the DNA-dependent protein kinase by poly(ADP-ribose) polymerase. J. Biol. Chem. 1998, 273, 14461–14467. [Google Scholar]
- Alvarez-Gonzalez, R.; Spring, H.; Muller, M.; Burkle, A. Selective loss of poly(ADP-ribose) and the 85-kDa fragment of poly(ADP-ribose) polymerase in nucleoli during alkylation-induced apoptosis of hela cells. J. Biol. Chem. 1999, 274, 32122–32126. [Google Scholar] [CrossRef]
- Soldani, C.; Lazze, M.C.; Bottone, M.G.; Tognon, G.; Biggiogera, M.; Pellicciari, C.E.; Scovassi, A.I. Poly(ADP-ribose) polymerase cleavage during apoptosis: When and where? Exp. Cell Res. 2001, 269, 193–201. [Google Scholar] [CrossRef]
- Beneke, S. Poly(ADP-ribose) polymerase activity in different pathologies--the link to inflammation and infarction. Exp. Gerontol. 2008, 43, 605–614. [Google Scholar] [CrossRef]
- Yu, S.W.; Wang, H.; Poitras, M.F.; Coombs, C.; Bowers, W.J.; Federoff, H.J.; Poirier, G.G.; Dawson, T.M.; Dawson, V.L. Mediation of poly(ADP-ribose) polymerase-1-dependent cell death by apoptosis-inducing factor. Science 2002, 297, 259–263. [Google Scholar]
- Tulin, A.; Spradling, A. Chromatin loosening by poly(ADP)-ribose polymerase (PARP) at drosophila puff loci. Science 2003, 299, 560–562. [Google Scholar] [CrossRef]
- Simbulan-Rosenthal, C.M.; Ly, D.H.; Rosenthal, D.S.; Konopka, G.; Luo, R.; Wang, Z.Q.; Schultz, P.G.; Smulson, M.E. Misregulation of gene expression in primary fibroblasts lacking poly(ADP-ribose) polymerase. Proc. Natl. Acad. Sci. USA 2000, 97, 11274–11279. [Google Scholar]
- Frizzell, K.M.; Gamble, M.J.; Berrocal, J.G.; Zhang, T.; Krishnakumar, R.; Cen, Y.; Sauve, A.A.; Kraus, W.L. Global analysis of transcriptional regulation by poly(ADP-ribose) polymerase-1 and poly(ADP-ribose) glycohydrolase in MCF-7 human breast cancer cells. J. Biol. Chem. 2009, 284, 33926–33938. [Google Scholar]
- Ogino, H.; Nozaki, T.; Gunji, A.; Maeda, M.; Suzuki, H.; Ohta, T.; Murakami, Y.; Nakagama, H.; Sugimura, T.; Masutani, M. Loss of PARP-1 affects gene expression profile in a genome-wide manner in es cells and liver cells. BMC Genomics 2007, 8, 41. [Google Scholar] [CrossRef]
- Butler, A.J.; Ordahl, C.P. Poly(ADP-ribose) polymerase binds with transcription enhancer factor 1 to MCAT1 elements to regulate muscle-specific transcription. Mol. Cell Biol. 1999, 19, 296–306. [Google Scholar]
- Plaza, S.; Aumercier, M.; Bailly, M.; Dozier, C.; Saule, S. Involvement of poly (ADP-ribose)-polymerase in the Pax-6 gene regulation in neuroretina. Oncogene 1999, 18, 1041–1051. [Google Scholar] [CrossRef]
- Amiri, K.I.; Ha, H.C.; Smulson, M.E.; Richmond, A. Differential regulation of CXC ligand 1 transcription in melanoma cell lines by poly(ADP-ribose) polymerase-1. Oncogene 2006, 25, 7714–7722. [Google Scholar] [CrossRef]
- Chu, S.; Xu, H.; Ferro, T.J.; Rivera, P.X. Poly(ADP-ribose) polymerase-1 regulates vimentin expression in lung cancer cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, L1127–1134. [Google Scholar] [CrossRef]
- Maruyama, T.; Nara, K.; Yoshikawa, H.; Suzuki, N. Txk, a member of the non-receptor tyrosine kinase of the tec family, forms a complex with poly(ADP-ribose) polymerase 1 and elongation factor 1alpha and regulates interferon-gamma gene transcription in Th1 cell. Clin. Exp. Immunol. 2007, 147, 164–175. [Google Scholar]
- Wang, J.; Bian, C.; Li, J.; Couch, F.J.; Wu, K.; Zhao, R.C. Poly(ADP-ribose) polymerase-1 down-regulates BRCA2 expression through the brca2 promoter. J. Biol. Chem. 2008, 283, 36249–36256. [Google Scholar]
- Ono, T.; Kaneda, T.; Muto, A.; Yoshida, T. Positive transcriptional regulation of the human micro opioid receptor gene by poly(ADP-ribose) polymerase-1 and increase of its DNA binding affinity based on polymorphism of G-172 ->T. J. Biol. Chem. 2009, 284, 20175–20183. [Google Scholar]
- Reinemund, J.; Seidel, K.; Steckelings, U.M.; Zaade, D.; Klare, S.; Rompe, F.; Katerbaum, M.; Schacherl, J.; Li, Y.; Menk, M., et al. Poly(ADP-ribose) polymerase-1 (parp-1) transcriptionally regulates angiotensin AT2 receptor (AT2R) and AT2R binding protein (ATBP) genes. Biochem. Pharmacol. 2009, 77, 1795–1805. [Google Scholar]
- Kang, X.; Kim, H.J.; Ramirez, M.; Salameh, S.; Ma, X. The septic shock-associated Il-10 -1082 A > G polymorphism mediates allele-specific transcription via poly(ADP-ribose) polymerase 1 in macrophages engulfing apoptotic cells. J. Immunol. 2010, 184, 3718–3724. [Google Scholar] [CrossRef]
- Zaniolo, K.; Desnoyers, S.; Leclerc, S.; Guerin, S.L. Regulation of poly(ADP-ribose) polymerase-1 (PARP-1) gene expression through the post-translational modification of SP1: A nuclear target protein of PARP-1. BMC Mol. Biol. 2007, 8, 96. [Google Scholar]
- Huang, D.; Yang, C.; Wang, Y.; Liao, Y.; Huang, K. PARP-1 suppresses adiponectin expression through poly(ADP-ribosyl)ation of PPAR gamma in cardiac fibroblasts. Cardiovasc. Res. 2009, 81, 98–107. [Google Scholar] [CrossRef]
- Lonn, P.; van der Heide, L.P.; Dahl, M.; Hellman, U.; Heldin, C.H.; Moustakas, A. PARP-1 attenuates Smad-mediated transcription. Mol. Cell 2010, 40, 521–532. [Google Scholar] [CrossRef]
- Gao, F.; Kwon, S.W.; Zhao, Y.; Jin, Y. Parp1 poly(ADP-ribosyl)ates Sox2 to control Sox2 protein levels and FGF4 expression during embryonic stem cell differentiation. J. Biol. Chem. 2009, 284, 22263–22273. [Google Scholar] [CrossRef]
- Hassa, P.O.; Covic, M.; Hasan, S.; Imhof, R.; Hottiger, M.O. The enzymatic and DNA binding activity of PARP-1 are not required for NF-kappa B coactivator function. J. Biol. Chem. 2001, 276, 45588–45597. [Google Scholar]
- Hassa, P.O.; Hottiger, M.O. The functional role of poly(ADP-ribose)polymerase 1 as novel coactivator of NF-kappaB in inflammatory disorders. Cell. Mol. Life. Sci. 2002, 59, 1534–1553. [Google Scholar] [CrossRef]
- Erener, S.; Petrilli, V.; Kassner, I.; Minotti, R.; Castillo, R.; Santoro, R.; Hassa, P.O.; Tschopp, J.; Hottiger, M.O. Inflammasome-activated caspase 7 cleaves PARP1 to enhance the expression of a subset of NF-kappaB target genes. Mol. Cell 2012, 46, 200–211. [Google Scholar] [CrossRef]
- Roy, R.; Chun, J.; Powell, S.N. BRCA1 and BRCA2: Different roles in a common pathway of genome protection. Nat. Rev. Cancer 2011, 12, 68–78. [Google Scholar] [CrossRef]
- Thorslund, T.; West, S.C. BRCA2: A universal recombinase regulator. Oncogene 2007, 26, 7720–7730. [Google Scholar] [CrossRef]
- Jensen, R.B.; Carreira, A.; Kowalczykowski, S.C. Purified human BRCA2 stimulates Rad51-mediated recombination. Nature 2010, 467, 678–683. [Google Scholar]
- Huang, J.; Gong, Z.; Ghosal, G.; Chen, J. SOSS complexes participate in the maintenance of genomic stability. Mol. Cell 2009, 35, 384–393. [Google Scholar] [CrossRef]
- Navadgi-Patil, V.M.; Burgers, P.M. A tale of two tails: Activation of DNA damage checkpoint kinase Mec1/ATR by the 9-1-1 clamp and by Dpb11/TopBP1. DNA Repair (Amst) 2009, 8, 996–1003. [Google Scholar]
- De Piccoli, G.; Torres-Rosell, J.; Aragon, L. The unnamed complex: What do we know about SMC5-SMC6? Chromosome Res. 2009, 17, 251–263. [Google Scholar] [CrossRef]
- Fekairi, S.; Scaglione, S.; Chahwan, C.; Taylor, E.R.; Tissier, A.; Coulon, S.; Dong, M.Q.; Ruse, C.; Yates, J.R., 3rd; Russell, P., et al. Human SLX4 is a holliday junction resolvase subunit that binds multiple DNA repair/recombination endonucleases. Cell 2009, 138, 78–89. [Google Scholar]
- Svendsen, J.M.; Smogorzewska, A.; Sowa, M.E.; O'Connell, B.C.; Gygi, S.P.; Elledge, S.J.; Harper, J.W. Mammalian BTBD12/SLX4 assembles a holliday junction resolvase and is required for DNA repair. Cell 2009, 138, 63–77. [Google Scholar] [CrossRef]
- Li, W.; Ma, H. Double-stranded DNA breaks and gene functions in recombination and meiosis. Cell Res. 2006, 16, 402–412. [Google Scholar] [CrossRef]
- Regnell, C.E.; H., G.A.; Sejersted, Y.; Medin, T.; Moldestad, O.; Rolseth, V.; Krokeide, S. Z.; Suganthan, R.; Luna, L.; Bjoras, M.; Bergersen, L.H. Hippocampal adult neurogenesis is maintained by NEIL3-dependent repair of oxidative DNA lesions in neural progenitor cells. Cell Reports 2012. [Google Scholar] [CrossRef]
- Zhu, J.K. Active DNA demethylation mediated by DNA glycosylases. Annu. Rev. Genet. 2009, 43, 143–166. [Google Scholar] [CrossRef]
- Bellacosa, A. Role of MED1 (MBD4) gene in DNA repair and human cancer. J. Cell Physiol. 2001, 187, 137–144. [Google Scholar] [CrossRef]
- Nishi, R.; Okuda, Y.; Watanabe, E.; Mori, T.; Iwai, S.; Masutani, C.; Sugasawa, K.; Hanaoka, F. Centrin 2 stimulates nucleotide excision repair by interacting with Xeroderma Pigmentosum group C protein. Mol. Cell. Biol. 2005, 25, 5664–5674. [Google Scholar] [CrossRef]
- Aas, P.A.; Otterlei, M.; Falnes, P.O.; Vagbo, C.B.; Skorpen, F.; Akbari, M.; Sundheim, O.; Bjoras, M.; Slupphaug, G.; Seeberg, E., et al. Human and bacterial oxidative demethylases repair alkylation damage in both RNA and DNA. Nature 2003, 421, 859–863. [Google Scholar]
- Duncan, T.; Trewick, S.C.; Koivisto, P.; Bates, P.A.; Lindahl, T.; Sedgwick, B. Reversal of DNA alkylation damage by two human dioxygenases. Proc. Natl. Acad. Sci. USA 2002, 99, 16660–16665. [Google Scholar] [CrossRef]
- Deans, A.J.; West, S.C. DNA interstrand crosslink repair and cancer. Nat. Rev. Cancer 2011, 11, 467–480. [Google Scholar] [CrossRef]
- Chou, D.M.; Adamson, B.; Dephoure, N.E.; Tan, X.; Nottke, A.C.; Hurov, K.E.; Gygi, S.P.; Colaiacovo, M.P.; Elledge, S.J. A chromatin localization screen reveals poly (ADP ribose)-regulated recruitment of the repressive polycomb and NuRD complexes to sites of DNA damage. Proc. Natl. Acad. Sci. USA 2010, 107, 18475–18480. [Google Scholar]
- Asher, G.; Reinke, H.; Altmeyer, M.; Gutierrez-Arcelus, M.; Hottiger, M.O.; Schibler, U. Poly(ADP-ribose) polymerase 1 participates in the phase entrainment of circadian clocks to feeding. Cell 2010, 142, 943–953. [Google Scholar] [CrossRef]
- Vidakovic, M.; Gluch, A.; Qiao, J.; Oumard, A.; Frisch, M.; Poznanovic, G.; Bode, J. PARP-1 expression in the mouse is controlled by an autoregulatory loop: PARP-1 binding to an upstream S/MAR element and to a novel recognition motif in its promoter suppresses transcription. J. Mol. Biol. 2009, 388, 730–750. [Google Scholar] [CrossRef]
- Xu, X.L.; Xing, B.C.; Han, H.B.; Zhao, W.; Hu, M.H.; Xu, Z.L.; Li, J.Y.; Xie, Y.; Gu, J.; Wang, Y., et al. The properties of tumor-initiating cells from a hepatocellular carcinoma patient's primary and recurrent tumor. Carcinogenesis 2009, 31, 167–174. [Google Scholar]
- Shimizu, S.; Nomura, F.; Tomonaga, T.; Sunaga, M.; Noda, M.; Ebara, M.; Saisho, H. Expression of poly(ADP-ribose) polymerase in human hepatocellular carcinoma and analysis of biopsy specimens obtained under sonographic guidance. Oncol. Rep. 2004, 12, 821–825. [Google Scholar]
- Goldberg, M. S.; Xing, D.; Ren, Y.; Orsulic, S.; Bhatia, S. N.; Sharp, P. A. Nanoparticle-mediated delivery of siRNA targeting Parp1 extends survival of mice bearing tumors derived from Brca1-deficient ovarian cancer cells. Proc. Natl. Acad. Sci. USA 2011, 108, 745–750. [Google Scholar]
- Mabley, J.G.; Horvath, E.M.; Murthy, K.G.; Zsengeller, Z.; Vaslin, A.; Benko, R.; Kollai, M.; Szabo, C. Gender differences in the endotoxin-induced inflammatory and vascular responses: Potential role of poly(ADP-ribose) polymerase activation. J. Pharmacol. Exp. Ther. 2005, 315, 812–820. [Google Scholar] [CrossRef]
- Liu, T.T.; Fang, Y.; Xiong, H.; Chen, T.Y.; Ni, Z.P.; Luo, J.F.; Zhao, N.Q.; Shen, X.Z. A case-control study of the relationship between hepatitis B virus DNA level and risk of hepatocellular carcinoma in qidong, china. World J. Gastroenterol. 2008, 14, 3059–3063. [Google Scholar]
- Chan, H.L.; Wong, V.W.; Wong, G.L.; Chim, A.M.; Lai, L.H.; Sung, J.J. Evaluation of impact of serial hepatitis B virus DNA levels on development of hepatocellular carcinoma. J. Clin. Microbiol. 2009, 47, 1830–1836. [Google Scholar] [CrossRef]
- Kwon, J.H.; Choi, J.Y.; Jang, J.W.; Bae, S.H.; Yoon, S.K.; Yang, J.M.; Han, N.I.; Lee, C.D.; Lee, Y.S.; Chung, K.W. Impact of serial hepatitis B virus DNA on hepatocellular carcinoma development in patients with liver cirrhosis. Intervirology 2009, 53, 111–118. [Google Scholar]
- Hagen, T.M.; Huang, S.; Curnutte, J.; Fowler, P.; Martinez, V.; Wehr, C.M.; Ames, B.N.; Chisari, F.V. Extensive oxidative DNA damage in hepatocytes of transgenic mice with chronic active hepatitis destined to develop hepatocellular carcinoma. Proc. Natl. Acad. Sci. USA 1994, 91, 12808–12812. [Google Scholar]
- Ohsaki, E.; Ueda, K.; Sakakibara, S.; Do, E.; Yada, K.; Yamanishi, K. Poly(ADP-ribose) polymerase 1 binds to kaposi's sarcoma-associated herpesvirus (KSHV) terminal repeat sequence and modulates KSHV replication in latency. J. Virol. 2004, 78, 9936–9946. [Google Scholar] [CrossRef]
- Wang, Y.; Li, H.; Tang, Q.; Maul, G.G.; Yuan, Y. Kaposi's sarcoma-associated herpesvirus ori-lyt-dependent DNA replication: Involvement of host cellular factors. J. Virol. 2008, 82, 2867–2882. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Ko, H.L.; Ren, E.C. Functional Aspects of PARP1 in DNA Repair and Transcription. Biomolecules 2012, 2, 524-548. https://doi.org/10.3390/biom2040524
AMA Style
Ko HL, Ren EC. Functional Aspects of PARP1 in DNA Repair and Transcription. Biomolecules. 2012; 2(4):524-548. https://doi.org/10.3390/biom2040524
Chicago/Turabian StyleKo, Hui Ling, and Ee Chee Ren. 2012. "Functional Aspects of PARP1 in DNA Repair and Transcription" Biomolecules 2, no. 4: 524-548. https://doi.org/10.3390/biom2040524