Why YRNAs? About Versatile RNAs and Their Functions

Martin-Luther-University Halle-Wittenberg, Institute of Molecular Medicine, Section Molecular Cell Biology, ZAMED, Heinrich-Damerow-Str.1, D-6120 Halle, Germany
*
Author to whom correspondence should be addressed.
Biomolecules 2013, 3(1), 143-156; https://doi.org/10.3390/biom3010143
Submission received: 31 December 2012
/
Revised: 27 January 2013
/
Accepted: 31 January 2013
/
Published: 8 February 2013
Abstract
:Y RNAs constitute a family of highly conserved small noncoding RNAs (in humans: 83-112 nt; Y1, Y3, Y4 and Y5). They are transcribed from individual genes by RNA-polymerase III and fold into conserved stem-loop-structures. Although discovered 30 years ago, insights into the cellular and physiological role of Y RNAs remains incomplete. In this review, we will discuss knowledge on the structural properties, associated proteins and discuss proposed functions of Y RNAs. We suggest Y RNAs to be an integral part of ribonucleoprotein networks within cells and could therefore have substantial influence on many different cellular processes. Putative functions of Y RNAs include small RNA quality control, DNA replication, regulation of the cellular stress response and proliferation. This suggests Y RNAs as essential regulators of cell fate and indicates future avenues of research, which will provide novel insights into the role of small noncoding RNAs in gene expression.
1. Introduction
Over recent years, our view of genomic regulation was severely revised when realizing that we have overseen a yet to be explored plethora of long and short non-coding RNAs (ncRNAs). Various reports revealed that ncRNAs are important regulators of diverse cellular processes. MicroRNAs (miRNAs), for instance, were shown to be important regulators of mRNA-fate in the cytoplasm, where they control translation and turnover of specific target transcripts (reviewed in [1]). On the other hand, long non-coding RNAs can serve as scaffolds for the assembly of subnuclear bodies (e.g., paraspeckles and NEAT1-RNA, [2]). These results reflect just two aspects of the various functions facilitated by ncRNAs, which modulate gene expression at various levels.
Y RNAs were originally identified in the 1980’s by immunopurification with auto-antibodies from patients suffering from systemic lupus erythematosus [3]. The proteins Ro60 (TROVE2, SSA) and La (SSB), which are common auto-antigens in autoimmune diseases (like systemic lupus erythematosus and Sjögren’s syndrome (reviewed in [4]), were identified as the major antigens facilitating the association with these small ncRNAs [5,6,7]. Subsequently, these cytoplasmic ncRNAs were named Y RNAs to distinguish them from nuclear U RNAs, another major class of small ncRNAs in the cell [3,8].
2. Structure and Evolution of Y RNAs
Metazoan Y RNAs are transcribed by RNA polymerase III (POLIII, [9]) from distinct promoters. Transcription is terminated at an OligoT stretch (4–6 nt), resulting in an oligo-uridinylated 3’-end of nascent Y RNA transcripts (Figure 1 and Figure 2). This serves as the primary binding site for the La protein [10,11,12]. It was proposed that the OligoU stretch is removed during nuclear maturation in the nucleus to promote nuclear export of trimmed Y RNAs [13]. The trimming of POLIII-transcripts seems to occur frequently and has been observed for various other RNAs [14]. Consistent with a pivotal role of La in the 3’-end processing of Y RNAs, La-binding was shown to prohibit the transport of Y RNAs to the cytoplasm [15]. However, the molecular basis of Y RNA trimming remains poorly understood and the potentially involved nucleases are still not known. Moreover, Y RNAs were shown to possess triphosphates at their 5’-ends and not to contain large amounts of nucleotide modifications [5].
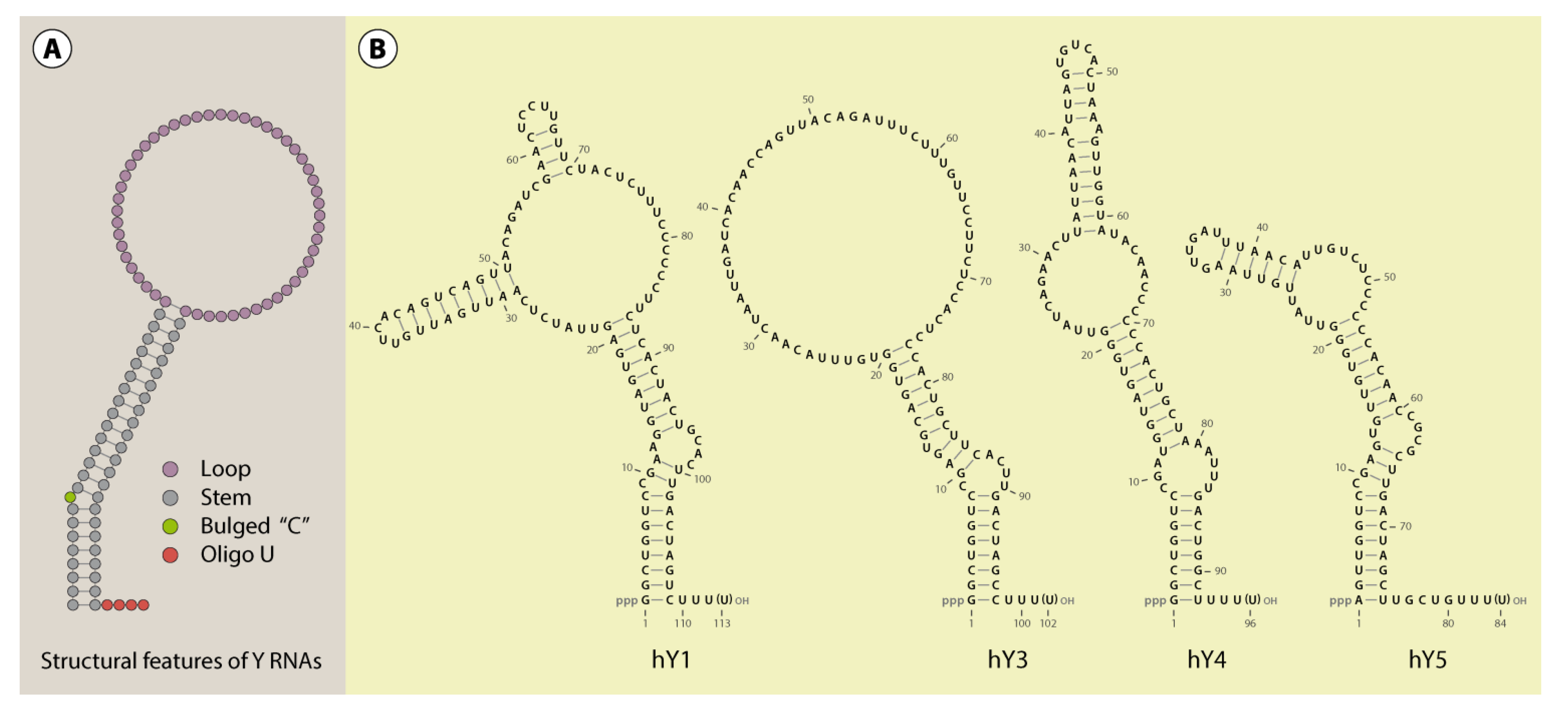
Figure 1.
Y RNA structure. A schematic structure of Y RNAs illustrates the important features common to these RNAs (A). Furthermore, the secondary structure of the human Y RNAs (Y1, Y3, Y4 and Y5) was visualized with VARNA [17], referring to published structure probing experiments (B; [18,19]). According to this data, alternative structures, at least for Y3, are likely [19].
Figure 1.
Y RNA structure. A schematic structure of Y RNAs illustrates the important features common to these RNAs (A). Furthermore, the secondary structure of the human Y RNAs (Y1, Y3, Y4 and Y5) was visualized with VARNA [17], referring to published structure probing experiments (B; [18,19]). According to this data, alternative structures, at least for Y3, are likely [19].

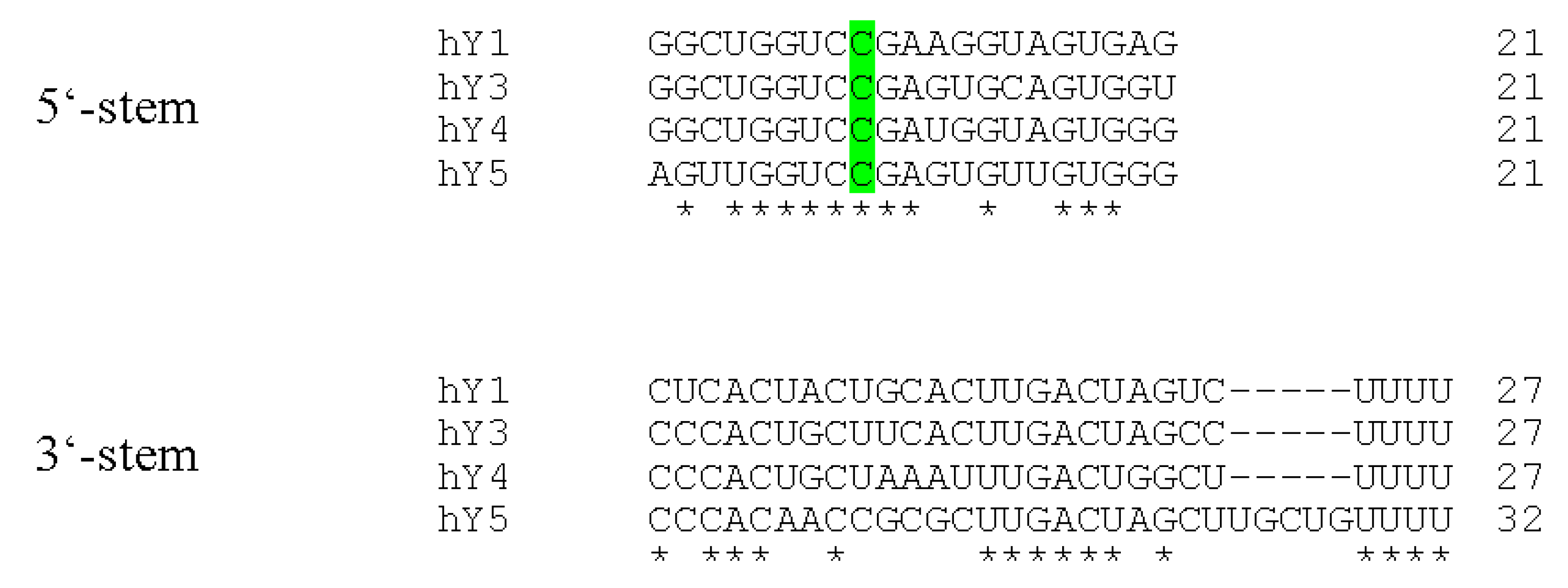
A common characteristic of all reported Y RNAs is their highly conserved stem-loop structure (Figure 1). The terminal 5’- and 3’-sequences (~20–30 nt) of Y RNAs form a double stranded region - the “stem”. This region is the essential structural determinant of Y RNAs allowing there typical stem-loop fold. For some Y RNAs, these structural properties were validated by enzymatic and chemical cleavage [18,19]. The stems of Y RNAs are usually not perfect double strands. Frequently, the “upper” and “lower” parts are separated by bulged regions. One of these, a highly conserved bulged cytosine (C9 in human Y RNAs [20]), constitutes the primary binding site for the Ro60 protein (TROVE2). Deletion or mutation of this site disrupts Ro60-binding and destabilizes the entire Y RNA fold (Figure 2, [15,20,21]). Ro60-homologs and also Y RNAs have been identified from bacteria to humans, suggesting an ancient origin and co-evolution of the Ro60-Y RNA-complex [21,22,23]. This is supported by analyses confirming the Y RNA stem to be the most conserved part of these ncRNAs, and even the bulged cytosine base in the stem is retained in bacterial Y RNAs (Figure 1 and Figure 2, [23,24]). Accordingly, we propose that organisms, which encode for Ro60 orthologs, presumably also express Y RNAs. Ro60 and its orthologs, as well some Y RNAs, were described or assumed in various bacterial species, lower eukaryotes, like Chlamydomonas reinhardtii, nematodes, like Caenorhabditis elegans, and vertebrates (reviewed in [21]). Putative Ro60 and Y RNA orthologs had also been suggested in arthropods, such as Anopheles gambiae [21,22].
Figure 2.
Sequence alignments of human Y RNA stems. The terminal Y RNA stem sequences were aligned using the TCoffee web server [25,26]. Perfectly conserved nucleotides are marked with asterisks. The remarkably conserved cytosine base at position 9 within the 5’-part of the Y RNAs is highlighted in green.
Figure 2.
Sequence alignments of human Y RNA stems. The terminal Y RNA stem sequences were aligned using the TCoffee web server [25,26]. Perfectly conserved nucleotides are marked with asterisks. The remarkably conserved cytosine base at position 9 within the 5’-part of the Y RNAs is highlighted in green.

RNA sequencing approaches frequently reported fragments comprising parts of Y RNAs [27,28]. In view of the conserved Y RNA structure resembling that of pre-miRNAs, it accordingly was suggested that Y RNAs could serve as miRNA precursors [28,29]. However, experimental validation of Y RNA encoded regulatory microRNAs is still lacking, and thus, the proposed Y RNA fragments could also result from degradation [30]. In support of this, it was shown that the biogenesis of some Y RNA fragments is independent of DICER1 and AGO2, providing further evidence that the identified fragments are not generated by the classical miRNA pathway [31].
The loops of Y RNAs are heterogeneous in nature and the least conserved of the ncRNAs [24,32]. The primary sequence and length of the loop distinguishes the four Y RNAs (Y1, Y3, Y4 and Y5). The longest loop is observed for Y1 (hY1: 65 nt) and the shortest for Y5 (hY5: 31 nt). The structure of the loops differs significantly among the four Y RNAs and was suggested to be largely flexible in nature [19]. The loops of Y1, Y3 and Y5 are rich in pyrimidines (human RNAs: 69 %, 65 % and 65 %, respectively); only Y1 and Y3 contain large, mostly single stranded stretches of pyrimidines. To date, in teleostei, only one Y RNA has been identified, which shows high homology to the human Y1/3-type of Y RNAs. Accordingly, Y3 was suggested to be the most ancestral Y RNA, at least in the vertebrate lineage [24,33]. Furthermore, additional Y RNAs possibly evolved through duplication of this ancestral RNA to fulfill novel functions within cells, which likely involves the loop region [24]. In accordance with this hypothesis, Y RNAs are able to recruit various RNA-binding proteins in a loop-dependent manner (see Table 1).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Alternative Names | Y RNA | Binding Region | Proposed Function | Reference |
|---|---|---|---|---|---|
| SSB | La | 1,3,4,5 | OligoU | nuclear retention, protection of Y RNA 3’ends | [15] |
| TROVE2 | Ro60 | 1,3,4,5 | stem | stabilization, | [15,23,34,35,36,37] |
| nuclear export, | |||||
| RNA quality control | |||||
| APOBEC3G | 1,3,4,5 | ? | ? | [38,39,40] | |
| NCL | nucleolin | 1,3 | loop | ? | [41] |
| PTBP1 | hnRNP I | 1,3 | loop | ? | [42] |
| HNRNPK | 1,3 | loop | ? | [42] | |
| IGF2BP1 | ZBP1, Imp1 | (1),3 | loop | nuclear Export of Ro60 and Y3 | [43,44] |
| PUF60 | RoBP1 | (1,3),5 | ? | ? | [45,46] |
3. Y RNA Localization and Expression
POLIII-transcripts can be transported to the cytoplasm (pre-miRNAs, tRNAs, 5S; [47,48]) or remain in the nucleus after transcription (7SK, U6; [49,50]). The localization of Y1, Y3 and Y4 was described to be mostly cytoplasmic, whereas Y5 seems to be more nuclear [51,52]. As mentioned above, the La protein is thought to interfere with nuclear export of Y RNAs by binding to their 3’-ends [15]. This export block could be released by trimming of Y RNAs, which then lack the OligoU stretch. This would trigger Ro60-dependent nuclear export of these ncRNAs [13,15]. Thus, it is tempting to speculate that Y5 differs in its association with La or nuclear trimming, allowing its nuclear retention. Nascent POLIII transcripts can accumulate in the perinucleolar compartment (PNC, [53,54]). In agreement, at least Y1, Y3 and Y5 were shown to localize to this subnuclear site by fluorescence in in situ hybridization (FISH, [55]). In an in vitro system, where labeled Y RNAs are incubated with G1 nuclei, Y RNAs were found to associate with euchromatin, and Y5 was recruited to nucleoli [56]. Notably, Y RNAs can be encapsidated into viruses, as shown for Moloney murine leukemia virus (MLV, [13]) and also for human immunodeficiency virus type 1 (HIV-1, [38]). This process is independent of Ro60-binding and seems to be initiated while Y RNAs are still in the nucleus. Whether Y RNAs modulate the lifecycle of these viruses significantly remains unknown.
The export pathways used by Y RNAs are not known in detail. It was reported that the export of Y RNAs is dependent on the small GTPase Ran, suggesting members of the karyopherin protein family to serve as nuclear export adapters [57]. Although XPO1 and XPOT are presumably not involved, XPO5 seems to be important to direct cytoplasmic translocation of Y RNAs [15,57]. This protein usually exports minihelix containing dsRNAs, which includes VA1, some tRNAs and pre-miRNAs [58,59]. The Y RNA stem is reminiscent of a minihelix, and consistently, XPO5 was shown to associate in a complex with Y1 and RanGTP [58]. This was also supported by the crystal structure of XPO5, indicating the Y RNA stem acts as a substrate for this karyopherin [47]. Notably, there is no evidence for a re-import of Y RNAs into the nucleus. This is supported by the complete nuclear export of radiolabeled Y RNAs after injection into Xenopus oocytes [15,57]. Notably, the subcellular localization of Y RNAs was reported to be cell cycle-dependent and respond to cellular stress signals, like UV-irradiation [23,56,60]. Accordingly, cells accumulate both Ro60 and Y RNAs in the nucleus after UV irradiation or oxidative stress [35,44,60]. This could result from the stress-induced collapse of the Ran gradient and concomitant impairment of nuclear export [61], but may furthermore imply stress-dependent functions of the nuclear Ro60-Y RNA-complex under these conditions.
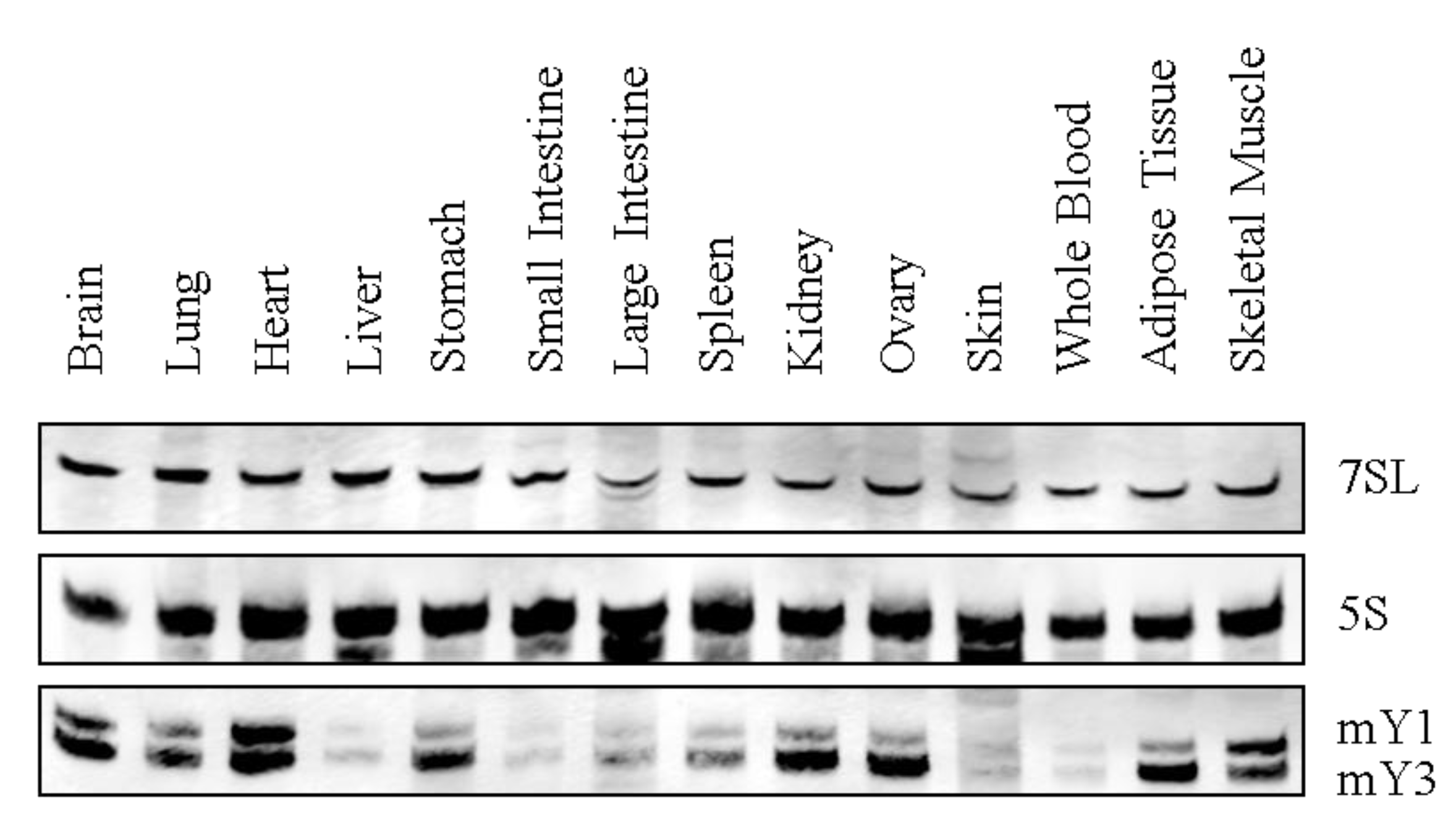
Y RNA expression has been reported in various species, including primary tissue and tumor-derived cell lines [62,63]. However, comprehensive analyses of tissue-specific Y RNA expression profiles are still lacking. Therefore, we analyzed the expression of murine Y RNAs in several adult tissues by Northern blotting (Figure 3). These studies revealed basal expression of murine Y RNAs (mY1 and mY3) in all analyzed tissues. Y RNA abundance varied significantly, with high levels observed in the brain, lung, heart, stomach, kidney, ovary, adipose tissue and skeletal muscle, in contrast to lower mY RNA abundance in the liver, gut, spleen, skin and blood. Intriguingly, the observed mY RNA expression pattern correlated to the expression signature of Ro60 [64]. This supports the view that the Y RNA-Ro60 complex co-evolved and that the protein is essential for Y RNA stabilization [34,36]. In humans, Y RNAs were shown to be significantly upregulated in a variety of tumors, for instance, bladder and kidney carcinomas. Moreover, Y RNAs promote cell proliferation, which is supported by reduced cell cycle progression upon siRNA-directed Y1 and Y3 depletion [63,65]. Little is known about the developmental expression of Y RNAs, which was analyzed exclusively in Xenopus laevis and Danio rerio, where Y RNA levels increased after the midblastula transition (MBT, [66]). Notably, the inhibition of Y RNAs by antisense morpholinos led to lethal developmental defects after MBT before gastrulation.
Figure 3.
Y RNA expression in mouse tissues. A representative Northern blot for murine Y RNA tissue expression is shown. 7SL and 5S rRNA served as a loading control. Note that in mice, just Y1 and Y3 are expressed.
Figure 3.
Y RNA expression in mouse tissues. A representative Northern blot for murine Y RNA tissue expression is shown. 7SL and 5S rRNA served as a loading control. Note that in mice, just Y1 and Y3 are expressed.

4. Is the Functional Role of Y RNA Determined by Associated Proteins?
4.1. Y RNP Core Proteins
All metazoan Y RNAs associate with Ro60 and La, which presumably form the core of nuclear or cytoplasmic Y RNPs (RNP: Ribonucleoprotein). As mentioned above, La binds the OligoU-stretch at the 3’-end of nascent Y RNAs [12]. On the contrary, Ro60 binds to the Y RNA stem [7]. Although La was found at the transcription site of Y RNAs, it remains to be addressed whether the protein has any influence on the transcription process itself [67]. Most likely, the protein protects nascent Y RNA transcripts from being degraded by 3’-exonucleases in the nucleus. Ro60, on the other hand, stabilizes the Y RNA structure and prevents Y RNA degradation [34,36]. In agreement, Y RNA levels are severely reduced in Ro60−/− cells [35]. However, the stabilizing function of Ro60 is presumed to mainly affect the cytoplasmic pool of Y RNAs, as shown for mouse fibroblasts [13]. Hence, it remains elusive whether other factors facilitate a Ro60-like role in modulating Y RNA fate in the nucleus [13,68]. Notably, the stabilization of Y RNAs by Ro60 is conserved in the bacterium Deinococcus radiodurans, where they act as essential facilitators of UV-resistance [23]. Additionally, the bacterial Rsr (Ro sixty related) was shown to assist in 23S rRNA maturation, and Y RNA were reported to sequester Rsr to inhibit this activity [37]. The crystal structure of Ro60-Y RNA complex revealed that Y RNAs associate with the outer surface of the HEAT-repeat-ring of Ro60, which comprises a highly conserved histidine residue (H187 in human and mouse) that is essential for direct contact formation with Y RNAs [69]. This association surface partially overlaps with the binding site for misfolded ncRNAs (like 5S), implying that Y RNAs modulate the proposed role of Ro60 in the quality surveillance of ncRNAs [60,69,70,71].
All four human Y RNAs associate with the antiviral cytidine deaminase APOBEC3G, which accordingly was observed in Ro60- and La-RNPs [38,39,40]. Mutational inactivation of the zinc-binding domain of APOBEC3G (W127A) strongly reduced its interaction with Y1 [38]. Hence, although the function of APOBEC3G-Y RNA complexes remains unknown, one could speculate that the protein facilitates C-to-U-editing to modulate Y RNA-functions and/or suppress retro-transposition of these ncRNAs [39]. Notably, retro-pseudogenes derived from Y RNAs have been described, but it is unclear if they can be re-expressed or have any other functional relevance [72]. The existence of another putative Y RNA core protein, which binds the upper Y RNA stem, is still controversial. Based on sequence requirements, this protein could be involved in nuclear export of Y RNAs and the proposed involvement of Y RNAs in DNA-replication [57,73]. In vitro, all four human Y RNAs were reported to associate with proteins of the pre-replication, as well as the origin recognition complex (e.g., ORC2 and CDT1, [56,66]). However, the functional composition of these Y RNPs remains to be deciphered, since the Y RNP core proteins, Ro60 and La, as well as nucleolin association, are not a prerequisite for Y RNAs to function in DNA replication [74].
4.2. Accessory Y RNA Binding Proteins
Apart from proteins associating at the Y RNA stem, additional proteins have been shown to bind Y RNAs. Most of these associate with distinct Y RNA loops and, thus, may impose Y RNA specific functions or play a role in directing the potentially distinct lifecycle of each Y RNA (for a summary, see Table 1 and Figure 4). Only the Y1- and Y3-RNAs contain pyrimidine-rich tracts within their loops. Accordingly, these Y RNAs associate with OligoU/C-binding proteins, including hnRNP I (PTBP1), hnRNP K (HNRNPK) and Nucleolin (NCL) [41,42]. Additionally, RoBP1 (PUF60) was shown to bind Y5 and also associates with Y1 and Y3, in vivo and in vitro [45,46]). The purification procedure used for PUF60-Y5-complexes led to the identification of RPL5 as a putative binding partner of Y5. In support of this, Y5 was shown to associate with 5S-rRNA-variants, suggesting a role of this Y RNA in the biogenesis of ribosomal RNAs [46], as demonstrated for Ro60 and La [75,76]. The identification of Ro60 in IGF2BP1 containing RNPs suggested an association of this protein with Y RNAs [77]. Recently, we and the Wolin lab could confirm this by demonstrating that mouse IGF2BP1, like its chicken ortholog ZBP1 (Zipcode binding protein), directly associates with Y3 and, to a lesser extent, with Y1 [43,44]. The depletion of IGF2BP1 resulted in the nuclear accumulation of Ro60 and Y3, suggesting an involvement of the protein in nuclear export of this Ro60-Y RNA complex [44].
Despite the various proteins described to associate with Y RNAs, the composition and properties of cellular Y RNPs remains largely elusive. Gel filtration studies indicate that Y RNPs range in size from 150–550 kDa. This suggests that one Y RNA can associate with more than one protein simultaneously. In vitro, we and others observed ternary complexes comprising La, Y3 and ZBP1 [43,78]. These findings suggest that Y RNPs contain at least one Y RNA, one core protein (e.g., Ro60) and one or two directly loop-associated accessory proteins (e.g., ZBP1), which could serve as binding scaffolds for additional proteins or promote oligomerization of Y RNPs (Figure 4).
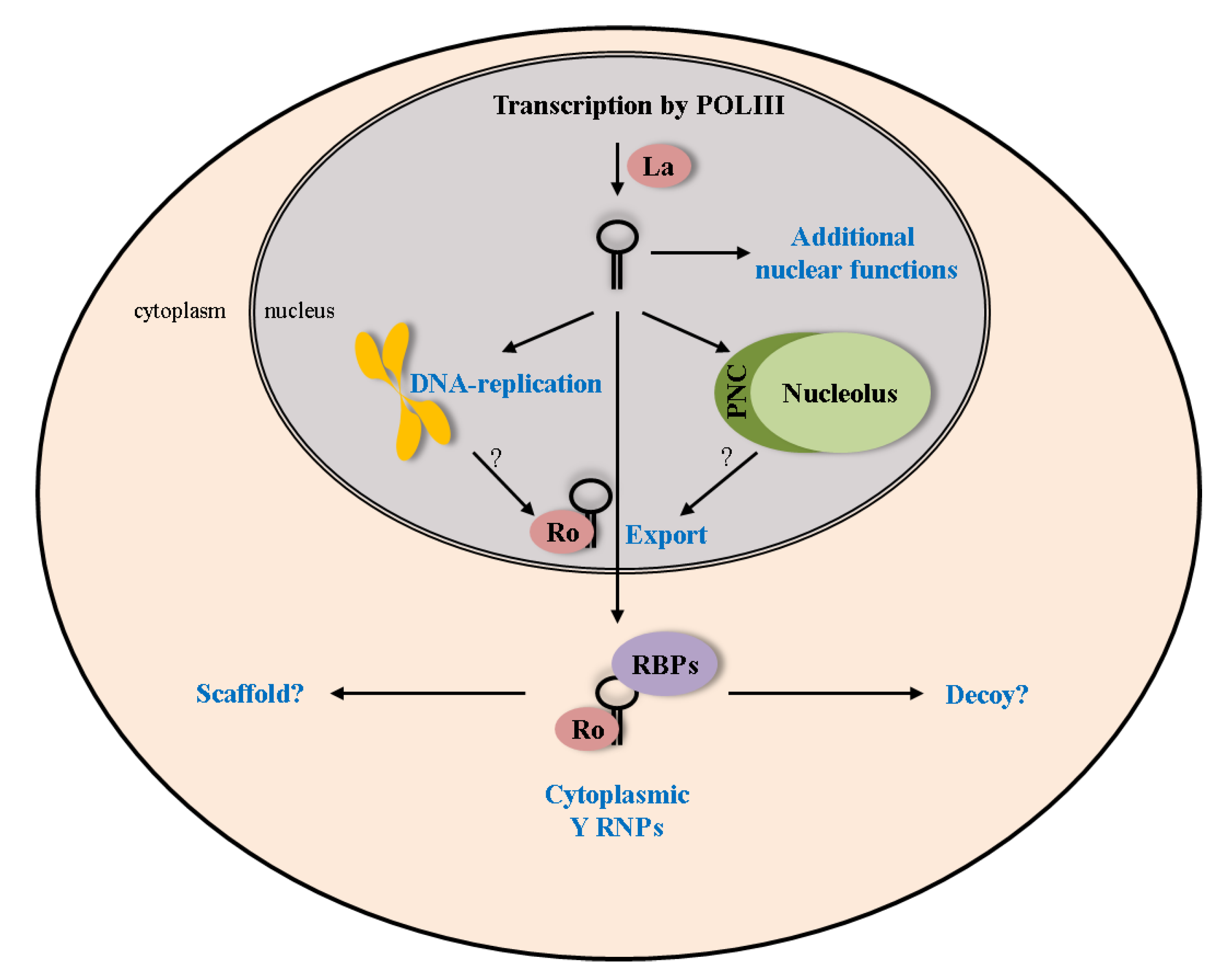
Figure 4.
Lifecycle of Y RNAs. The proposed cellular functions of Y RNAs rely mostly on the association with their core proteins Ro60 and La. These interactions influence various parts of the Y RNA lifecycle (e.g., nuclear export together with Ro60). Additional functions and interactions with RNA binding proteins (RBPs) have to be assumed in the nucleus, as well as the cytoplasm.
Figure 4.
Lifecycle of Y RNAs. The proposed cellular functions of Y RNAs rely mostly on the association with their core proteins Ro60 and La. These interactions influence various parts of the Y RNA lifecycle (e.g., nuclear export together with Ro60). Additional functions and interactions with RNA binding proteins (RBPs) have to be assumed in the nucleus, as well as the cytoplasm.

5. Future Perspectives and Conclusions
Although various Y RNA-associated proteins have been reported, the role of this highly conserved family of small ncRNAs and, in particular, the reason for their diversification remains sparse. Only two Y RNA functions have been proposed so far. During the cell cycle, they were suggested to stimulate DNA replication (Figure 4; [65,73,79]). This assumption is mainly based on in vitro evidence, using isolated G1-phase nuclei incubated with cellular extracts. These studies also indicated that Y RNAs are required for the establishment of new replication forks, but not for DNA elongation [79]. Although the model of Y RNAs being involved in DNA replication is intriguing, more information regarding the molecular mechanisms and in vivo regulation of these processes are required. However, a role of Y RNAs in DNA replication could be supported by their high conservation, as well as the devastating developmental defects and the proliferation decrease upon Y RNA inhibition [63,66]. On the other hand, Y RNAs were reported to inhibit the function of Ro60 in RNA quality control [37]. Accordingly, it was also shown that the binding site for Y RNAs in Ro60 partially overlaps with the one for misfolded RNAs, suggesting a mutually exclusive Ro60-RNP [69]. Therefore, we favor a model that Y RNAs and bound Ro60 act as cellular stress sensors. Accordingly, Ro60 can dissociate from Y RNAs under conditions, like UV-irradiation, to assist in cellular recovery by salvaging misfolded RNAs (reviewed in [21]). We propose that, in addition to Ro60, other RBPs could be sequestered by Y RNAs in a similar fashion. This is in accordance with the proposed functions of long ncRNAs, which can act as decoys and/or scaffolds to regulate gene expression [80]. A putative scaffolding role could provide both a sequestering of these regulators acting like a “molecular sink” or a chaperoning function to expedite function of the associated proteins [80]. We thus expect that Y RNAs modulate additional regulatory processes controlling the fate of mRNAs, which could include RNA processing, as well as cytoplasmic regulation of gene expression. The latter assumption is supported by the observation that Y RNAs are predominantly cytoplasmic at steady state. Surprisingly, most of the proposed roles of Y RNAs involve nuclear functions like DNA-replication or small RNA quality control. Future work will thus have to address the compartment-specific function of Y RNPs, which we expect to reveal that these small ncRNAs serve functions in modulating cytoplasmic mRNA fate. This role could essentially rely on the capability of Y RNAs to associate with various RBPs observed in mRNPs, for instance, IGF2BP1 (reviewed in [81]). By controlling the accessibility of such factors, Y RNAs could modulate the function of these regulatory RBPs in mRNA turnover, translation and, potentially, mRNA localization at regular or stress conditions. In addition to revealing the cellular role of Y RNAs, future studies also have to address the physiological significance of these small ncRNAs by the use of genetic models to address their function in development and diseases.
6. Materials and Methods: Isolation of Total RNA and Northern Blot
Tissues were isolated from athymic Nude-Foxn1nu-mice (Harlan) and rapidly mixed with TRIZOL-reagent (Life Technologies). RNA-extraction was then performed using chloroform and precipitation with isopropanol. For Northern Blotting, 2.5 µg of total RNA was resolved on a 15% denaturing TBE-Urea-gel and subsequently blotted onto nylon membranes (Roche). The membranes were then UV-crosslinked (Stratalinker 2400) and pre-hybridized with PerfectHyb Plus (Sigma-Aldrich). Northern probes (Atto680 or DY-782 label) were diluted to 100 ng/µL in PerfectHyb Plus and hybridized at 30 °C for 2 hours. Detection was conducted using the Odyssey Scanner (LI-COR). Northern Probes: 7SL: GGCATAGCGCACTACAGCCCAGAACTCCTG; Y1: ATAACTCACTACCTTCGGACCAGCC; Y3: CTGTAACTGGTTGTGATCAATTAGT; and 5S: AAGTACTAACCAGGCCCGAC.
Acknowledgments
This work was funded by the SFB 610 (DFG) to Stefan Hüttelmaier.
Conflict of Interest
The authors declare no conflict of interest.
References
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar]
- Bond, C.S.; Fox, A.H. Paraspeckles: nuclear bodies built on long noncoding RNA. J. Cell Biol. 2009, 186, 637–644. [Google Scholar] [CrossRef]
- Lerner, M.R.; Boyle, J.A.; Hardin, J.A.; Steitz, J.A. Two novel classes of small ribonucleoproteins detected by antibodies associated with lupus erythematosus. Science 1981, 211, 400–402. [Google Scholar]
- Franceschini, F.; Cavazzana, I. Anti-Ro/SSA and La/SSB antibodies. Autoimmunity 2005, 38, 55–63. [Google Scholar] [CrossRef]
- Hendrick, J.P.; Wolin, S.L.; Rinke, J.; Lerner, M.R.; Steitz, J.A. Ro small cytoplasmic ribonucleoproteins are a subclass of La ribonucleoproteins: further characterization of the Ro and La small ribonucleoproteins from uninfected mammalian cells. Mol. Cell. Biol. 1981, 1, 1138–1149. [Google Scholar]
- Francoeur, A.M.; Mathews, M.B. Interaction between VA RNA and the lupus antigen La: formation of a ribonucleoprotein particle in vitro. Proc. Natl. Acad. Sci. USA 1982, 79, 6772–6776. [Google Scholar] [CrossRef]
- Wolin, S.L.; Steitz, J.A. The Ro small cytoplasmic ribonucleoproteins: identification of the antigenic protein and its binding site on the Ro RNAs. Proc. Natl. Acad. Sci. USA 1984, 81, 1996–2000. [Google Scholar] [CrossRef]
- Will, C.L.; Lührmann, R. Splicing of a rare class of introns by the U12-dependent spliceosome. Biol. Chem. 2005, 386, 713–724. [Google Scholar]
- Wolin, S.L.; Steitz, J.A. Genes for two small cytoplasmic Ro RNAs are adjacent and appear to be single-copy in the human genome. Cell 1983, 32, 735–744. [Google Scholar] [CrossRef]
- Stefano, J.E. Purified lupus antigen La recognizes an oligouridylate stretch common to the 3' termini of RNA polymerase III transcripts. Cell 1984, 36, 145–154. [Google Scholar]
- Mathews, M.B.; Francoeur, A.M. La antigen recognizes and binds to the 3'-oligouridylate tail of a small RNA. Mol. Cell. Biol. 1984, 4, 1134–1140. [Google Scholar]
- Pruijn, G.J.; Slobbe, R.L.; van Venrooij, W.J. Analysis of protein--RNA interactions within Ro ribonucleoprotein complexes. Nucleic Acids Res. 1991, 19, 5173–5180. [Google Scholar]
- Garcia, E.L.; Onafuwa-Nuga, A.; Sim, S.; King, S.R.; Wolin, S.L.; Telesnitsky, A. Packaging of host mY RNAs by murine leukemia virus may occur early in Y RNA biogenesis. J. Virol. 2009, 83, 12526–12534. [Google Scholar]
- van Hoof, A.; Lennertz, P.; Parker, R. Three conserved members of the RNase D family have unique and overlapping functions in the processing of 5S, 5.8S, U4, U5, RNase MRP and RNase P RNAs in yeast. EMBO J. 2000, 19, 1357–1365. [Google Scholar] [CrossRef]
- Simons, F.H.; Rutjes, S.A.; van Venrooij, W.J.; Pruijn, G.J. The interactions with Ro60 and La differentially affect nuclear export of hY1 RNA. RNA 1996, 2, 264–273. [Google Scholar]
- Gruber, A.R.; Lorenz, R.; Bernhart, S.H.; Neuböck, R.; Hofacker, I.L. The Vienna RNA websuite. Nucleic Acids Res. 2008, 36, W70–W74. [Google Scholar]
- Darty, K.; Denise, A.; Ponty, Y. VARNA: Interactive drawing and editing of the RNA secondary structure. Bioinformatics 2009, 25, 1974–1975. [Google Scholar] [Green Version]
- van Gelder, C.W.; Thijssen, J.P.; Klaassen, E.C.; Sturchler, C.; Krol, A.; van Venrooij, W.J.; Pruijn, G.J. Common structural features of the Ro RNP associated hY1 and hY5 RNAs. Nucleic Acids Res. 1994, 22, 2498–2506. [Google Scholar]
- Teunissen, S.W.; Kruithof, M.J.; Farris, A.D.; Harley, J.B.; Venrooij, W.J.; Pruijn, G.J. Conserved features of Y RNAs: a comparison of experimentally derived secondary structures. Nucleic Acids Res. 2000, 28, 610–619. [Google Scholar]
- Green, C.D.; Long, K.S.; Shi, H.; Wolin, S.L. Binding of the 60-kDa Ro autoantigen to Y RNAs: evidence for recognition in the major groove of a conserved helix. RNA 1998, 4, 750–765. [Google Scholar]
- Sim, S.; Wolin, S.L. Emerging roles for the Ro 60-kDa autoantigen in noncoding RNA metabolism. Wiley Interdiscip. Rev. RNA 2011, 2, 686–699. [Google Scholar]
- Perreault, J.; Perreault, J.-P.; Boire, G. Ro-associated Y RNAs in metazoans: evolution and diversification. Mol. Biol. Evol. 2007, 24, 1678–1689. [Google Scholar]
- Chen, X.; Quinn, A.M.; Wolin, S.L. Ro ribonucleoproteins contribute to the resistance of Deinococcus radiodurans to ultraviolet irradiation. Genes Dev. 2000, 14, 777–782. [Google Scholar]
- Mosig, A.; Guofeng, M.; Stadler, B.M.R.; Stadler, P.F. Evolution of the vertebrate Y RNA cluster. Theory Biosci. 2007, 126, 9–14. [Google Scholar]
- Poirot, O.; O'Toole, E.; Notredame, C. Tcoffee@igs: A web server for computing, evaluating and combining multiple sequence alignments. Nucleic Acids Res. 2003, 31, 3503–3506. [Google Scholar]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-Coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar]
- Liao, J.-Y.; Ma, L.-M.; Guo, Y.-H.; Zhang, Y.-C.; Zhou, H.; Shao, P.; Chen, Y.-Q.; Qu, L.-H. Deep sequencing of human nuclear and cytoplasmic small RNAs reveals an unexpectedly complex subcellular distribution of miRNAs and tRNA 3' trailers. PLoS ONE 2010, 5, e10563. [Google Scholar]
- Meiri, E.; Levy, A.; Benjamin, H.; Ben-David, M.; Cohen, L.; Dov, A.; Dromi, N.; Elyakim, E.; Yerushalmi, N.; Zion, O.; et al. Discovery of microRNAs and other small RNAs in solid tumors. Nucleic Acids Res. 2010, 38, 6234–6246. [Google Scholar] [CrossRef]
- Verhagen, A.P.M.; Pruijn, G.J.M. Are the Ro RNP-associated Y RNAs concealing microRNAs? Y RNA-derived miRNAs may be involved in autoimmunity. Bioessays 2011, 33, 674–682. [Google Scholar]
- Rutjes, S.A.; van der Heijden, A.; Utz, P.J.; van Venrooij, W.J.; Pruijn, G.J. Rapid nucleolytic degradation of the small cytoplasmic Y RNAs during apoptosis. J. Biol. Chem. 1999, 274, 24799–24807. [Google Scholar]
- Nicolas, F.E.; Hall, A.E.; Csorba, T.; Turnbull, C.; Dalmay, T. Biogenesis of Y RNA-derived small RNAs is independent of the microRNA pathway. FEBS Lett. 2012, 586, 1226–1230. [Google Scholar]
- Farris, A.D.; Koelsch, G.; Pruijn, G.J.; van Venrooij, W.J.; Harley, J.B. Conserved features of Y RNAs revealed by automated phylogenetic secondary structure analysis. Nucleic Acids Res. 1999, 27, 1070–1078. [Google Scholar] [CrossRef]
- Farris, A.D.; O'Brien, C.A.; Harley, J.B. Y3 is the most conserved small RNA component of Ro ribonucleoprotein complexes in vertebrate species. Gene 1995, 154, 193–198. [Google Scholar] [CrossRef]
- Xue, D.; Shi, H.; Smith, J.D.; Chen, X.; Noe, D.A.; Cedervall, T.; Yang, D.D.; Eynon, E.; Brash, D.E.; Kashgarian, M.; et al. A lupus-like syndrome develops in mice lacking the Ro 60-kDa protein, a major lupus autoantigen. Proc. Natl. Acad. Sci. USA 2003, 100, 7503–7508. [Google Scholar]
- Sim, S.; Weinberg, D.E.; Fuchs, G.; Choi, K.; Chung, J.; Wolin, S.L. The subcellular distribution of an RNA quality control protein, the Ro autoantigen, is regulated by noncoding Y RNA binding. Mol. Biol. Cell 2009, 20, 1555–1564. [Google Scholar] [CrossRef]
- Labbé, J.C.; Hekimi, S.; Rokeach, L.A. The levels of the RoRNP-associated Y RNA are dependent upon the presence of ROP-1, the Caenorhabditis elegans Ro60 protein. Genetics 1999, 151, 143–150. [Google Scholar]
- Chen, X.; Wurtmann, E.J.; van Batavia, J.; Zybailov, B.; Washburn, M.P.; Wolin, S.L. An ortholog of the Ro autoantigen functions in 23S rRNA maturation in D. radiodurans. Genes Dev. 2007, 21, 1328–1339. [Google Scholar] [CrossRef]
- Wang, T.; Tian, C.; Zhang, W.; Luo, K.; Sarkis, P.T.N.; Yu, L.; Liu, B.; Yu, Y.; Yu, X.-F. 7SL RNA mediates virion packaging of the antiviral cytidine deaminase APOBEC3G. J. Virol. 2007, 81, 13112–13124. [Google Scholar] [CrossRef]
- Chiu, Y.-L.; Witkowska, H.E.; Hall, S.C.; Santiago, M.; Soros, V.B.; Esnault, C.; Heidmann, T.; Greene, W.C. High-molecular-mass APOBEC3G complexes restrict Alu retrotransposition. Proc. Natl. Acad. Sci. USA 2006, 103, 15588–15593. [Google Scholar]
- Gallois-Montbrun, S.; Holmes, R.K.; Swanson, C.M.; Fernández-Ocaña, M.; Byers, H.L.; Ward, M.A.; Malim, M.H. Comparison of cellular ribonucleoprotein complexes associated with the APOBEC3F and APOBEC3G antiviral proteins. J. Virol. 2008, 82, 5636–5642. [Google Scholar]
- Fouraux, M.A.; Bouvet, P.; Verkaart, S.; van Venrooij, W.J.; Pruijn, G.J.M. Nucleolin associates with a subset of the human Ro ribonucleoprotein complexes. J. Mol. Biol. 2002, 320, 475–488. [Google Scholar]
- Fabini, G.; Raijmakers, R.; Hayer, S.; Fouraux, M.A.; Pruijn, G.J.; Steiner, G. The heterogeneous nuclear ribonucleoproteins I and K interact with a subset of the ro ribonucleoprotein-associated Y RNAs in vitro and in vivo. J. Biol. Chem. 2001, 276, 20711–20718. [Google Scholar]
- Köhn, M.; Lederer, M.; Wächter, K.; Hüttelmaier, S. Near-infrared (NIR) dye-labeled RNAs identify binding of ZBP1 to the noncoding Y3-RNA. RNA 2010, 16, 1420–1428. [Google Scholar]
- Sim, S.; Yao, J.; Weinberg, D.E.; Niessen, S.; Yates, J.R.; Wolin, S.L. The zipcode-binding protein ZBP1 influences the subcellular location of the Ro 60-kDa autoantigen and the noncoding Y3 RNA. RNA 2012, 18, 100–110. [Google Scholar] [CrossRef]
- Bouffard, P.; Barbar, E.; Brière, F.; Boire, G. Interaction cloning and characterization of RoBPI, a novel protein binding to human Ro ribonucleoproteins. RNA 2000, 6, 66–78. [Google Scholar]
- Hogg, J.R.; Collins, K. Human Y5 RNA specializes a Ro ribonucleoprotein for 5S ribosomal RNA quality control. Genes Dev. 2007, 21, 3067–3072. [Google Scholar]
- Okada, C.; Yamashita, E.; Lee, S.J.; Shibata, S.; Katahira, J.; Nakagawa, A.; Yoneda, Y.; Tsukihara, T. A high-resolution structure of the pre-microRNA nuclear export machinery. Science 2009, 326, 1275–1279. [Google Scholar]
- Murdoch, K.; Loop, S.; Rudt, F.; Pieler, T. Nuclear export of 5S rRNA-containing ribonucleoprotein complexes requires CRM1 and the RanGTPase cycle. Eur. J. Cell Biol. 2002, 81, 549–556. [Google Scholar] [CrossRef]
- Peterlin, B.M.; Brogie, J.E.; Price, D.H. 7SK snRNA: a noncoding RNA that plays a major role in regulating eukaryotic transcription. Wiley Interdiscip. Rev. RNA 2012, 3, 92–103. [Google Scholar]
- Spiller, M.P.; Boon, K.-L.; Reijns, M.A.M.; Beggs, J.D. The Lsm2-8 complex determines nuclear localization of the spliceosomal U6 snRNA. Nucleic Acids Res. 2007, 35, 923–929. [Google Scholar]
- Gendron, M.; Roberge, D.; Boire, G. Heterogeneity of human Ro ribonucleoproteins (RNPS): nuclear retention of Ro RNPS containing the human hY5 RNA in human and mouse cells. Clin. Exp. Immunol. 2001, 125, 162–168. [Google Scholar] [CrossRef]
- Farris, A.D.; Puvion-Dutilleul, F.; Puvion, E.; Harley, J.B.; Lee, L.A. The ultrastructural localization of 60-kDa Ro protein and human cytoplasmic RNAs: association with novel electron-dense bodies. Proc. Natl. Acad. Sci. USA 1997, 94, 3040–3045. [Google Scholar]
- Wang, C.; Politz, J.C.; Pederson, T.; Huang, S. RNA polymerase III transcripts and the PTB protein are essential for the integrity of the perinucleolar compartment. Mol. Biol. Cell 2003, 14, 2425–2435. [Google Scholar]
- Pollock, C.; Huang, S. The perinucleolar compartment. J. Cell. Biochem. 2009, 107, 189–193. [Google Scholar]
- Matera, A.G.; Frey, M.R.; Margelot, K.; Wolin, S.L. A perinucleolar compartment contains several RNA polymerase III transcripts as well as the polypyrimidine tract-binding protein, hnRNP I. J. Cell Biol. 1995, 129, 1181–1193. [Google Scholar]
- Zhang, A.T.; Langley, A.R.; Christov, C.P.; Kheir, E.; Shafee, T.; Gardiner, T.J.; Krude, T. Dynamic interaction of Y RNAs with chromatin and initiation proteins during human DNA replication. J. Cell. Sci. 2011, 124, 2058–2069. [Google Scholar]
- Rutjes, S.A.; Lund, E.; van der Heijden, A.; Grimm, C.; van Venrooij, W.J.; Pruijn, G.J. Identification of a novel cis-acting RNA element involved in nuclear export of hY RNAs. RNA 2001, 7, 741–752. [Google Scholar]
- Gwizdek, C.; Ossareh-Nazari, B.; Brownawell, A.M.; Doglio, A.; Bertrand, E.; Macara, I.G.; Dargemont, C. Exportin-5 mediates nuclear export of minihelix-containing RNAs. J. Biol. Chem. 2003, 278, 5505–5508. [Google Scholar]
- Calado, A.; Treichel, N.; Müller, E.-C.; Otto, A.; Kutay, U. Exportin-5-mediated nuclear export of eukaryotic elongation factor 1A and tRNA. EMBO J. 2002, 21, 6216–6224. [Google Scholar]
- Chen, X.; Smith, J.D.; Shi, H.; Yang, D.D.; Flavell, R.A.; Wolin, S.L. The Ro autoantigen binds misfolded U2 small nuclear RNAs and assists mammalian cell survival after UV irradiation. Curr. Biol. 2003, 13, 2206–2211. [Google Scholar]
- Miyamoto, Y.; Saiwaki, T.; Yamashita, J.; Yasuda, Y.; Kotera, I.; Shibata, S.; Shigeta, M.; Hiraoka, Y.; Haraguchi, T.; Yoneda, Y. Cellular stresses induce the nuclear accumulation of importin alpha and cause a conventional nuclear import block. J. Cell Biol. 2004, 165, 617–623. [Google Scholar] [CrossRef]
- Pruijn, G.J.; Wingens, P.A.; Peters, S.L.; Thijssen, J.P.; van Venrooij, W.J. Ro RNP associated Y RNAs are highly conserved among mammals. Biochim. Biophys. Acta 1993, 1216, 395–401. [Google Scholar]
- Christov, C.P.; Trivier, E.; Krude, T. Noncoding human Y RNAs are overexpressed in tumours and required for cell proliferation. Br. J. Cancer 2008, 98, 981–988. [Google Scholar]
- Wang, D.; Buyon, J.P.; Chan, E.K. Cloning and expression of mouse 60 kDa ribonucleoprotein SS-A/Ro. Mol. Biol. Rep. 1996, 23, 205–210. [Google Scholar]
- Christov, C.P.; Gardiner, T.J.; Szüts, D.; Krude, T. Functional requirement of noncoding Y RNAs for human chromosomal DNA replication. Mol. Cell. Biol. 2006, 26, 6993–7004. [Google Scholar]
- Collart, C.; Christov, C.P.; Smith, J.C.; Krude, T. The midblastula transition defines the onset of Y RNA-dependent DNA replication in Xenopus laevis. Mol. Cell. Biol. 2011, 31, 3857–3870. [Google Scholar]
- Fairley, J.A.; Kantidakis, T.; Kenneth, N.S.; Intine, R.V.; Maraia, R.J.; White, R.J. Human La is found at RNA polymerase III-transcribed genes in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 18350–18355. [Google Scholar]
- Simons, F.H.; Pruijn, G.J.; van Venrooij, W.J. Analysis of the intracellular localization and assembly of Ro ribonucleoprotein particles by microinjection into Xenopus laevis oocytes. J. Cell Biol. 1994, 125, 981–988. [Google Scholar]
- Stein, A.J.; Fuchs, G.; Fu, C.; Wolin, S.L.; Reinisch, K.M. Structural insights into RNA quality control: the Ro autoantigen binds misfolded RNAs via its central cavity. Cell 2005, 121, 529–539. [Google Scholar]
- Hogg, J.R.; Collins, K. Structured non-coding RNAs and the RNP Renaissance. Curr. Opin. Chem. Biol. 2008, 12, 684–689. [Google Scholar]
- Macrae, I.J.; Doudna, J.A. Ro's role in RNA reconnaissance. Cell 2005, 121, 495–496. [Google Scholar]
- Perreault, J.; Noël, J.-F.; Brière, F.; Cousineau, B.; Lucier, J.-F.; Perreault, J.-P.; Boire, G. Retropseudogenes derived from the human Ro/SS-A autoantigen-associated hY RNAs. Nucleic Acids Res. 2005, 33, 2032–2041. [Google Scholar]
- Gardiner, T.J.; Christov, C.P.; Langley, A.R.; Krude, T. A conserved motif of vertebrate Y RNAs essential for chromosomal DNA replication. RNA 2009, 15, 1375–1385. [Google Scholar] [CrossRef]
- Langley, A.R.; Chambers, H.; Christov, C.P.; Krude, T. Ribonucleoprotein particles containing non-coding Y RNAs, Ro60, La and nucleolin are not required for Y RNA function in DNA replication. PLoS ONE 2010, 5, e13673. [Google Scholar]
- Shi, H.; O'Brien, C.A.; van Horn, D.J.; Wolin, S.L. A misfolded form of 5S rRNA is complexed with the Ro and La autoantigens. RNA 1996, 2, 769–784. [Google Scholar]
- O'Brien, C.A.; Wolin, S.L. A possible role for the 60-kD Ro autoantigen in a discard pathway for defective 5S rRNA precursors. Genes Dev. 1994, 8, 2891–2903. [Google Scholar]
- Jønson, L.; Vikesaa, J.; Krogh, A.; Nielsen, L.K.; Hansen, T.v.; Borup, R.; Johnsen, A.H.; Christiansen, J.; Nielsen, F.C. Molecular composition of IMP1 ribonucleoprotein granules. Mol. Cell Proteomics 2007, 6, 798–811. [Google Scholar] [CrossRef]
- Fabini, G.; Rutjes, S.A.; Zimmermann, C.; Pruijn, G.J.; Steiner, G. Analysis of the molecular composition of Ro ribonucleoprotein complexes. Identification of novel Y RNA-binding proteins. Eur. J. Biochem. 2000, 267, 2778–2789. [Google Scholar] [CrossRef]
- Krude, T.; Christov, C.P.; Hyrien, O.; Marheineke, K. Y RNA functions at the initiation step of mammalian chromosomal DNA replication. J. Cell. Sci. 2009, 122, 2836–2845. [Google Scholar]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar]
- Bell, J.L.; Wächter, K.; Mühleck, B.; Pazaitis, N.; Köhn, M.; Lederer, M.; Hüttelmaier, S. Insulin-like growth factor 2 mRNA-binding proteins (IGF2BPs): post-transcriptional drivers of cancer progression? Cell. Mol. Life Sci. 2012. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Köhn, M.; Pazaitis, N.; Hüttelmaier, S. Why YRNAs? About Versatile RNAs and Their Functions. Biomolecules 2013, 3, 143-156. https://doi.org/10.3390/biom3010143
AMA Style
Köhn M, Pazaitis N, Hüttelmaier S. Why YRNAs? About Versatile RNAs and Their Functions. Biomolecules. 2013; 3(1):143-156. https://doi.org/10.3390/biom3010143
Chicago/Turabian StyleKöhn, Marcel, Nikolaos Pazaitis, and Stefan Hüttelmaier. 2013. "Why YRNAs? About Versatile RNAs and Their Functions" Biomolecules 3, no. 1: 143-156. https://doi.org/10.3390/biom3010143