
TORC1-Dependent Phosphorylation Targets in Fission Yeast

1
Laboratory of Cell Responses, National Institute for Basic Biology, Nishigonaka 38, Myodaiji, Okazaki, Aichi 444-8585, Japan
2
Biosignal Research Center, Kobe University, 1-1 Rokkodai-cho, Nada-ku, Kobe 657-8501, Japan
3
Department of Basic Biology, School of Life Science, SOKENDAI (The Graduate University for Advanced Studies), Nishigonaka 38, Myodaiji, Okazaki, Aichi 444-8585, Japan
*
Author to whom correspondence should be addressed.
Biomolecules 2017, 7(3), 50; https://doi.org/10.3390/biom7030050
Submission received: 2 June 2017
/
Revised: 27 June 2017
/
Accepted: 28 June 2017
/
Published: 3 July 2017
(This article belongs to the Special Issue TOR Signaling Pathway)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Target of rapamycin (TOR) kinase controls cell metabolism and growth in response to environmental cues such as nutrients, growth factors, and stress. TOR kinase is widely conserved across eukaryotes. As in other organisms, the fission yeast Schizosaccharomyces pombe has two types of TOR complex, namely TOR complex 1 (TORC1) and TORC2. It is interesting that the two TOR complexes in S. pombe have opposite roles in sexual differentiation, which is induced by nutrient starvation. TORC1, which contains Tor2 as a catalytic subunit, promotes vegetative growth and represses sexual differentiation in nutrient-rich conditions, while TORC2 is required for the initiation of sexual differentiation. Multiple targets of TORC1 have been identified. Some of these, such as S6 kinase and an autophagy regulator Atg13, are known targets in other organisms. In addition, there is a novel group of TORC1 targets involved in the regulation of sexual differentiation. Here, we review recent findings on phosphorylation targets of TORC1 in S. pombe. Furthermore, we briefly report a novel S. pombe target of TORC1.
1. Introduction
It is a fundamental property of cells to grow and decide when to divide by sensing environmental conditions. Target of rapamycin (TOR), a highly conserved serine/threonine kinase, acts as a key factor in this process, and orchestrates various growth-related functions in response to environmental conditions [1]. TOR exists in two distinct complexes termed TOR complex 1 (TORC1) and 2 (TORC2). TORC1 facilitates anabolic processes such as protein synthesis, transcription, and ribosome biogenesis, and inhibits catabolic processes such as autophagy. The macrolide rapamycin, which is known to be an immunosuppressive and antiproliferative agent, acutely and preferentially inhibits TORC1 activity [2]. Mammalian cells have a single TOR kinase, mTOR (mammalian or mechanistic TOR), and both TORC1 and TORC2 share mTOR as the catalytic subunit [3]. In yeast species, TORC1 and TORC2 contain different TOR kinases. In the budding yeast Saccharomyces cerevisiae, in which TOR kinase was originally identified [4], TORC1 contains Tor1 or Tor2, and TORC2 contains Tor2 [5].
While TORC1-mediated signaling is unambiguously important and indispensable for eukaryotic cells, the current knowledge regarding TORC1 targets is still restricted. The most well-known substrates of TORC1 are S6K1 (p70 ribosomal protein S6 kinase 1), which is a protein kinase to phosphorylate ribosomal protein S6, and 4E-BP1 (eIF4E-binding protein). TORC1 promotes protein synthesis through phosphorylation of these effectors. S6K1 is a member of the AGC (protein kinase A/G/C) kinase family, which includes PKA (protein kinase A, also known as cAMP (cyclic adenosine monophosphate)-dependent protein kinase), AKT (also known as protein kinase B or PKB), SGK (serum- and glucocorticoid-induced protein kinase), and PKC (protein kinase C) [6]. AGC kinases are highly conserved and play an important role in growth, proliferation, and survival. A number of AGC kinases are shown to be activated by phosphorylation. S6K1 is phosphorylated by TORC1 at a threonine residue (Thr389 in human) at the C-terminal hydrophobic motif [7,8]. Phosphorylation of 4E-BP1 by TORC1 leads to its dissociation from eIF4E and alleviates the inhibitory effect of 4E-BP1 on initiation of translation [9,10].
Fission yeast Schizosaccharomyces pombe is a unicellular microorganism, which is one of the best model organisms for the study of molecular mechanisms underlying various eukaryotic cellular processes, as well as the budding yeast S. cerevisiae. An intriguing feature of S. pombe is that sensing of nutritional (especially nitrogen) availability is intimately linked to the change of growth mode from vegetative to reproductive. Under nutrient-rich conditions, the fission yeast normally proliferates by mitotic division in the haploid state, displaying one of two mating types, h+ or h−. When starved of nitrogen, the cells begin sexual differentiation, that is, cells exit from the G1 phase of the mitotic cycle and conjugate with cells of the opposite mating type to form diploid zygotes, which then undergo meiosis and spore formation.
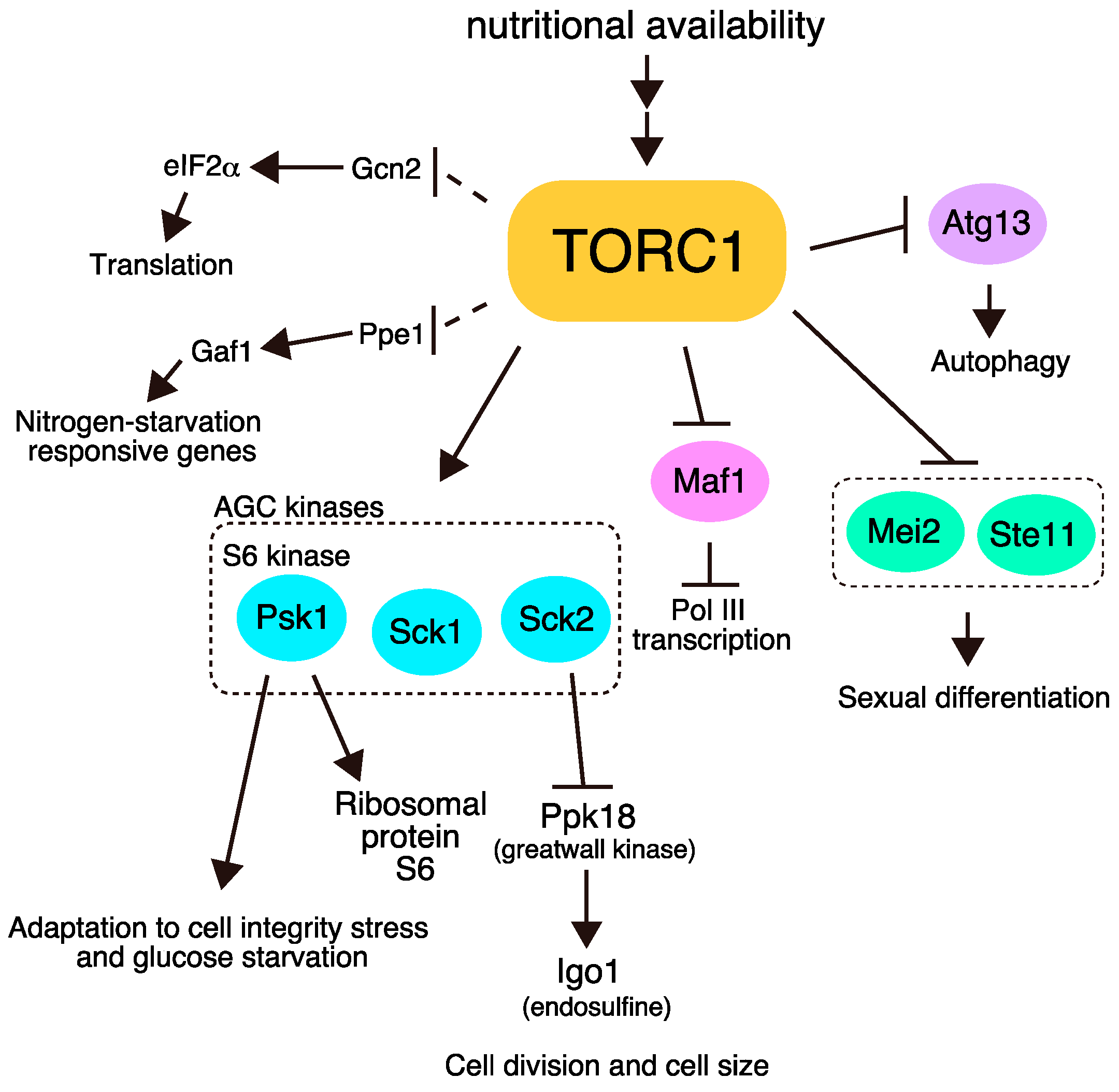
S. pombe has two TOR homologs, Tor1 and Tor2 [11,12]. Similar to other eukaryotes, S. pombe contains two forms of TOR complex, namely TORC1 and TORC2 [13,14,15]. TORC1, which contains Tor2 as the catalytic subunit, and TORC2, which contains Tor1, play opposite roles in the initiation of sexual differentiation. TORC1 is essential for vegetative growth and represses sexual differentiation under nitrogen-rich conditions [13,14,15,16,17]. Rapamycin does not affect vegetative growth of wild-type S. pombe cells [18], although TORC1 is downregulated by treatment of rapamycin [19,20,21,22]. Cells with reduced TORC1 activity, due to a temperature-sensitive tor2 mutation for instance, stop proliferation and initiate sexual differentiation ectopically even in the presence of ample nitrogen. Conversely, TORC2 is required for sexual differentiation. Cells lacking TORC2 cannot enter sexual differentiation [11,12,23]. It has been demonstrated that the major downstream target of TORC2 is Gad8, an ortholog of AKT [23]. In contrast, although multiple phosphorylation targets of TORC1 have been identified, its critical target(s) remains ambiguous. In this review, we overview TORC1-phosphorylation targets in S. pombe (Figure 1).
2. Conserved Phosphorylation Targets of TORC1
2.1. Psk1
AGC family kinase Psk1 is an S6K1 homolog in S. pombe [20]. Similar to mammalian S6K1, several conserved regulatory motifs in Psk1 are phosphorylated by TORC1 and Ksg1 kinase, a homolog of PDK1. Of these target sites, phosphorylation of the hydrophobic motif (Thr415) and the turn motif (Thr392) is regulated by TORC1 [20]. Direct phosphorylation of these two sites by TORC1 has been demonstrated in vitro. Consistent with direct regulation of Psk1 by TORC1, Psk1 is phosphorylated in a nutrient (nitrogen, glucose, and glutamine)-dependent and rapamycin-sensitive manner. Since a commercially available phospho-specific antibody against the phosphorylated hydrophobic motif in mammalian S6K, namely anti-phospho-p70 S6K (Thr389) antibody, can recognize the phosphorylation of Thr415 in Psk1, its phosphorylation status can serve as an excellent readout of TORC1 activity (Figure 2).
Psk1 phosphorylates ribosomal protein S6, i.e., Rps601 and Rps602, in response to nutritional availability, depending on the TORC1 activity [21]. However, lack of Psk1 does not phenocopy loss of TORC1 function. Whereas the reduction of the TORC1 activity leads to ectopic entry into sexual differentiation, the disruption of the psk1 gene shows little effect on the growth mode [20]. It has been recently suggested that Psk1 cooperates with TORC2-target Gad8 to adapt to cell integrity stress and glucose starvation, although the precise mechanism remains unclear [25].
2.2. Sck1 and Sck2
In addition to Psk1, two other AGC kinases, Sck1 and Sck2, are phosphorylated by TORC1 in vitro [20]. In vivo phosphorylation of Sck1 and Sck2 is indeed dependent on TORC1 [20]. These observations suggest that both Sck1 and Sck2 are direct effectors of TORC1. It was originally shown that both Sck1 and Sck2 have overlapping function with Pka1, which is the catalytic subunit of cAMP-dependent protein kinase (PKA) [26,27]. The interconnection between TORC1 and PKA, both of which regulate the onset of sexual differentiation, by Sck1 has been suggested [28], while the precise relationship between them remains elusive. Recently, it has been demonstrated that TORC1 promotes the activity of the protein phosphatase PP2A-B55 through inhibiting the Greatwall-Endosulfine (Ppk18-Igo1 in S. pombe) pathway. PP2A-B55 antagonizes CDK activity in the G2 phase, resulting in increasing cell size in G2 [29,30]. The involvement of Sck2 in the TORC1 and Greatwall-Endosulfine pathway has been suggested, owing to the fact that the overexpression of Sck2 inhibits Greatwall kinase Ppk18 activity [29]. It has also been reported that the overexpression of Sck2 leads to cell elongation [31,32].
2.3. Maf1
RNA polymerase III (pol III) generates small untranslated structural RNAs for protein synthesis, such as 5S rRNA and tRNA. In response to nutrient availability and cellular stress, pol III transcription is controlled by a conserved repressor Maf1, which is regulated by phosphorylation [33,34,35]. Dephosphorylated Maf1 represses pol III-dependent transcription. In the budding yeast S. cerevisiae, the major Maf1 kinase is Sch9, which is an ortholog of S6K1/Psk1 and a target of TORC1 [36,37], whereas S. cerevisiae TORC1 could weakly phosphorylate Maf1 in vitro [38]. In mammalian cells, TORC1 is responsible for Maf1 phosphorylation [39,40,41]. In S. pombe, Maf1 phosphorylation depends on TORC1, but not TORC2, although it remains elusive whether TORC1 is directly involved [22].
2.4. Atg13
Autophagy is a highly conserved intracellular degradation mechanism in eukaryotes [42]. TORC1 negatively regulates autophagy under nutrient-rich conditions. In the budding yeast S. cerevisiae, the conserved serine/threonine kinase Atg1 is pivotal in the regulation of autophagy, which forms a multimeric protein complex [43,44,45]. Atg13 is an essential regulatory member of the Atg1 kinase complex. In S. cerevisiae, Atg13 is highly phosphorylated in nutrient-rich conditions, and becomes rapidly dephosphorylated upon nutrient deprivation or inhibition of TORC1 by rapamycin [43]. It has been shown that TORC1 directly phosphorylates Atg13 at multiple serine residues in vitro [46]. TORC1 inhibition, which leads to dephosphorylation of Atg13, promotes Atg1 complex formation, increases Atg1 activity, and results in the induction of autophagy [43,46]. Since Atg1 also contains a phosphorylation site that is downregulated upon rapamycin addition [47], Atg1 might also be a substrate of TORC1 in S. cerevisiae.
In mammals, TORC1 also negatively regulates autophagy [48]. Mammalian homologs of Atg1 are uncoordinated-51-like kinase 1 and 2 (ULK1 and ULK2). As in S. cerevisiae, ULK1 forms a multimeric protein complex, including mammalian Atg13 [49,50,51]. While TORC1 has been shown to phosphorylate both ULK1 and Atg13 in vitro [50,51], TORC1 has no impact on the formation of the ULK1 complex [50,52]. In vivo phosphorylation of ULK1 by TORC1 prevents the interaction between ULK1 and AMP-activated protein kinase (AMPK) [53]. It has also been shown that TORC1 phosphorylates an autophagy regulator Atg14 in mammalian cells [54].
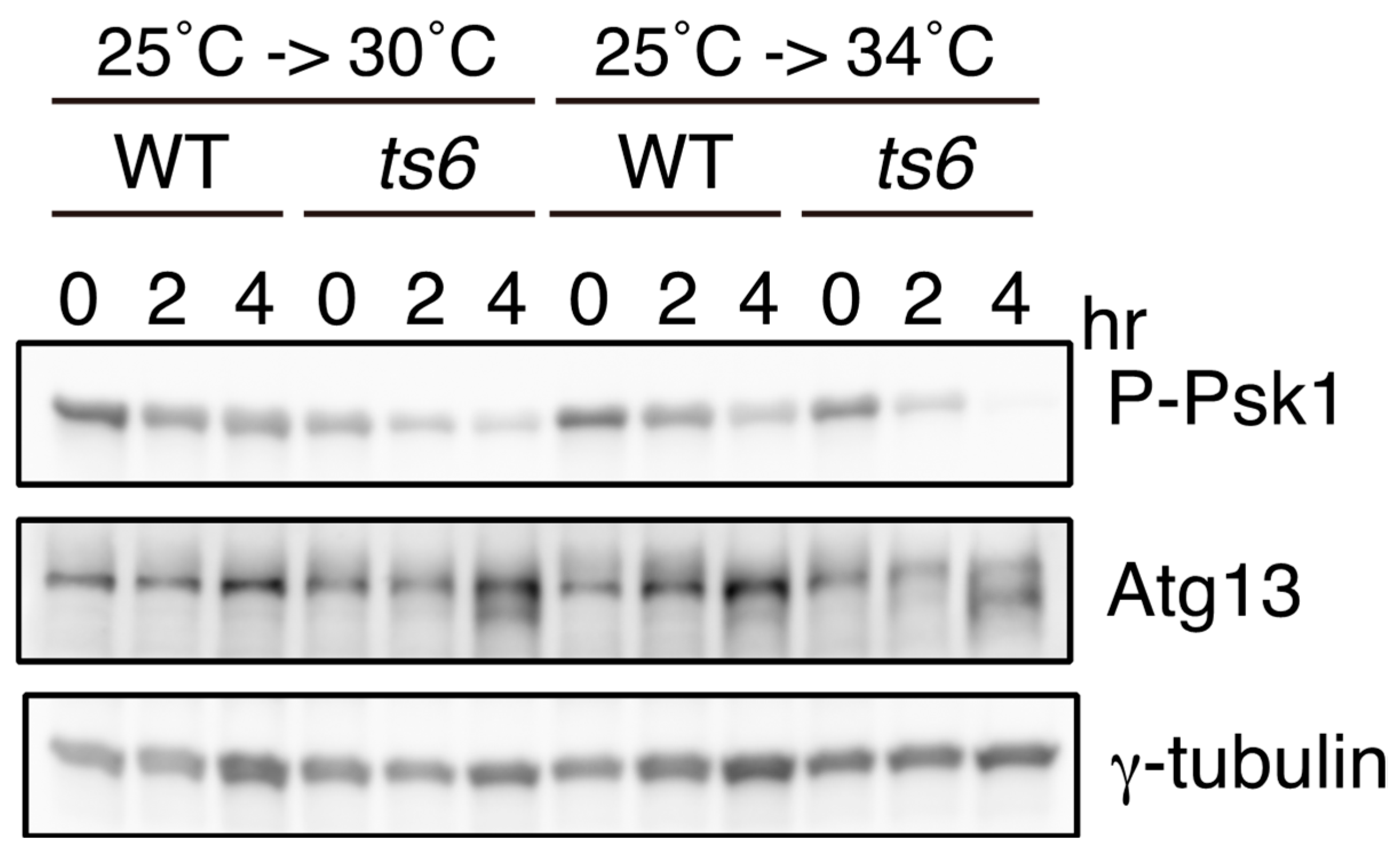
S. pombe carries an autophagy system similar to that of other eukaryotes. Autophagy-deficient mutants lose their viability under conditions of prolonged nitrogen starvation [24]. In addition, these mutants are unable to initiate sexual differentiation under nitrogen starvation conditions [24]. As in S. cerevisiae, it has been demonstrated that S. pombe Atg13 is highly phosphorylated under nutrient-rich conditions, and this phosphorylation decreases upon nitrogen starvation, which induces the inactivation of TORC1 [24]. Here, we present data to confirm that the phosphorylation of Atg13 is dependent on TORC1, as shown using a temperature-sensitive tor2 mutant (Figure 2). In tor2-ts6 cells, Atg13 migrated faster when cells were shifted to a restrictive temperature of 30 °C or 34 °C, indicating that the phosphorylation of Atg13 is dependent on TORC1. This also implies that Atg13 serves as a good readout of TORC1 activity, as is the case with Psk1. While the phosphorylation of S. pombe Atg1 is also alleviated upon nitrogen starvation [24], it remains unknown whether Atg1 is a TORC1 target.
3. Direct Phosphorylation Targets of TORC1 that Regulate Sexual Differentiation
An RNA-binding protein, Mei2, is a master regulator of meiosis in S. pombe [55,56,57]. In addition to the regulation of meiosis, Mei2 functions in the earlier stages of sexual differentiation, and the loss of Mei2 results in a partial deficiency in G1 arrest and conjugation [55,58]. Mip1, which is the S. pombe ortholog of mammalian Raptor, an essential component of mTORC1, was originally isolated as a genetic interactor of Mei2 and was shown to physically interact with Mei2 [59]. Physical interaction between Mei2 and Tor2 has also been demonstrated [13]. These observations suggested the close relationship between Mei2 and TORC1, and it has indeed been demonstrated that TORC1 phosphorylates multiple sites of Mei2 in vivo and in vitro [58]. Mei2 phosphorylation by TORC1 induces polyubiquitination and proteasomal degradation of Mei2, leading to the suppression of sexual differentiation [58]. Mei2-like protein is widely conserved in plants [60,61]. It is intriguing that an Arabidopsis Raptor homolog, named AtRaptor1B, binds to Mei2-like protein AML1 as in S. pombe [60], suggesting a conserved regulatory mechanism between S. pombe and plants.
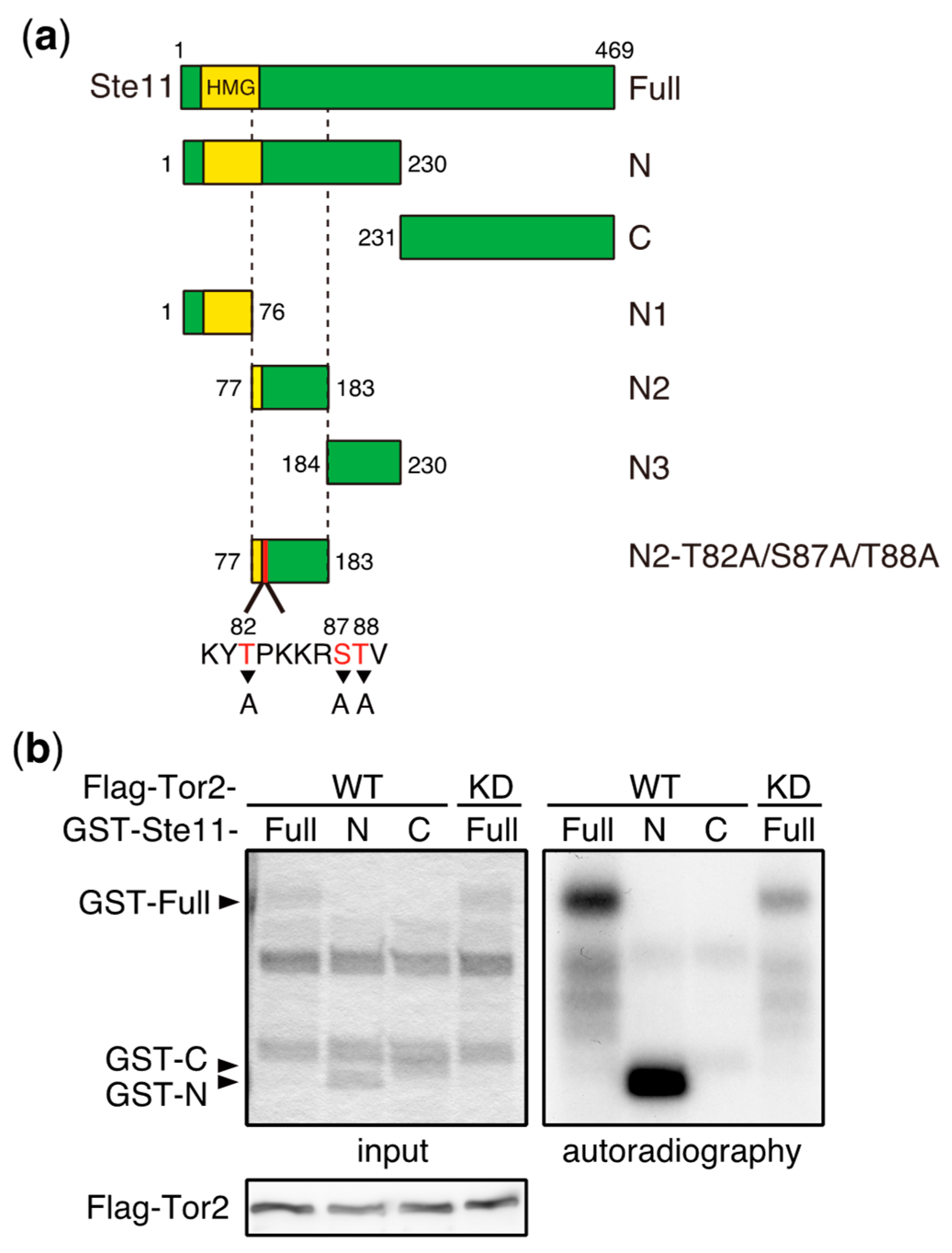
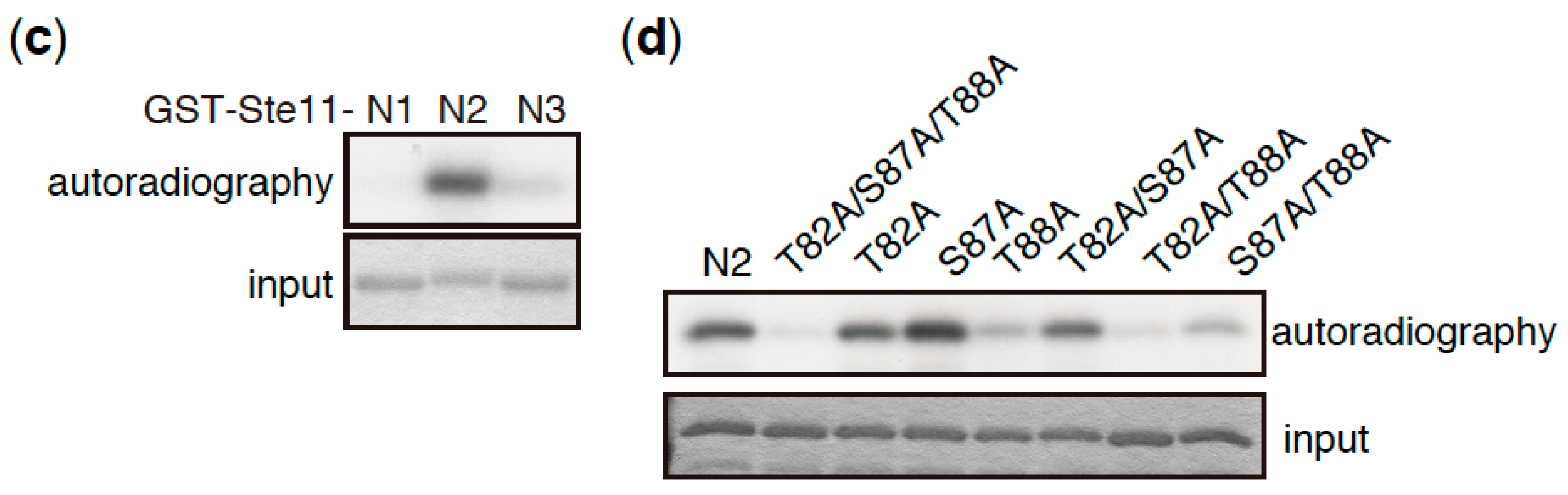
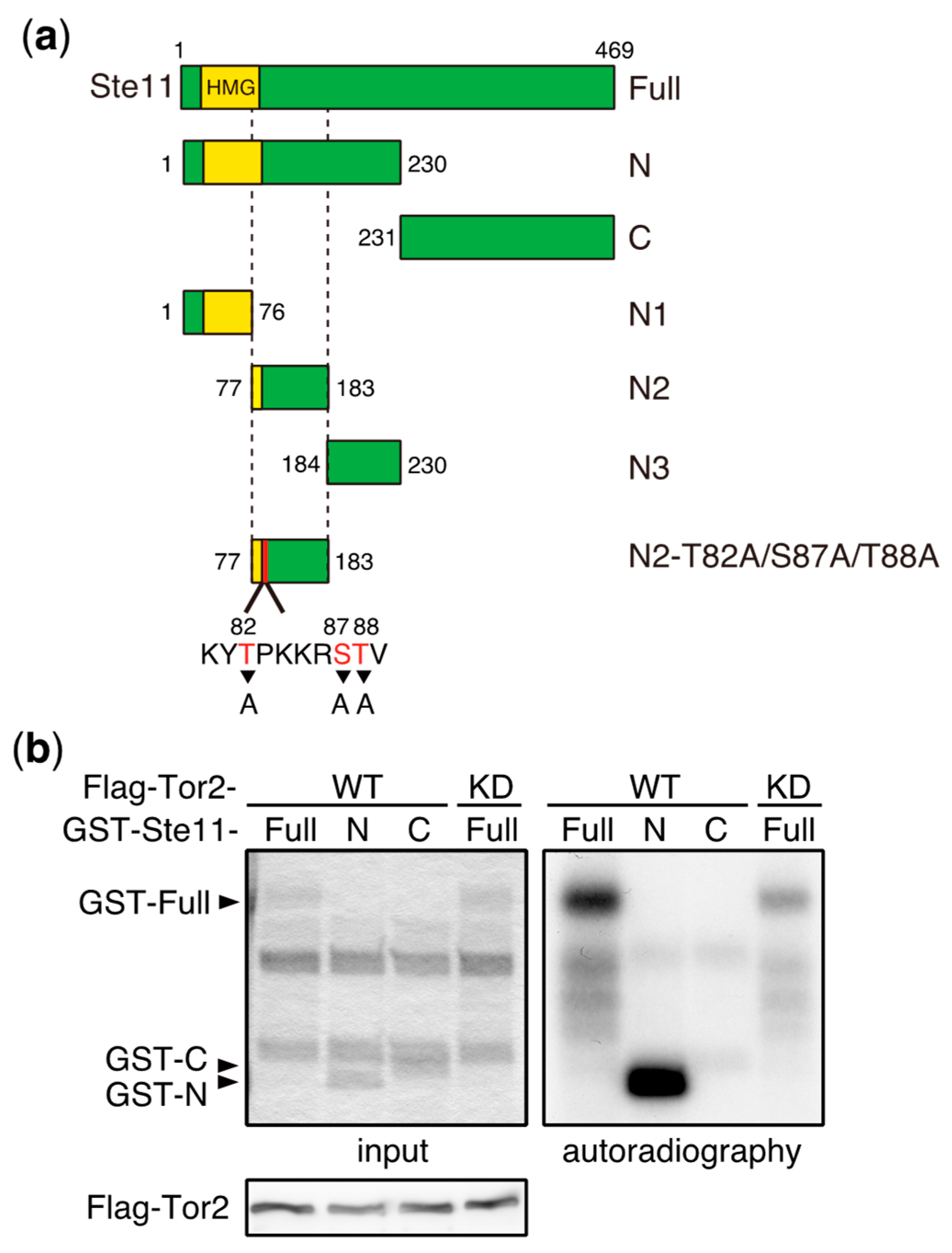
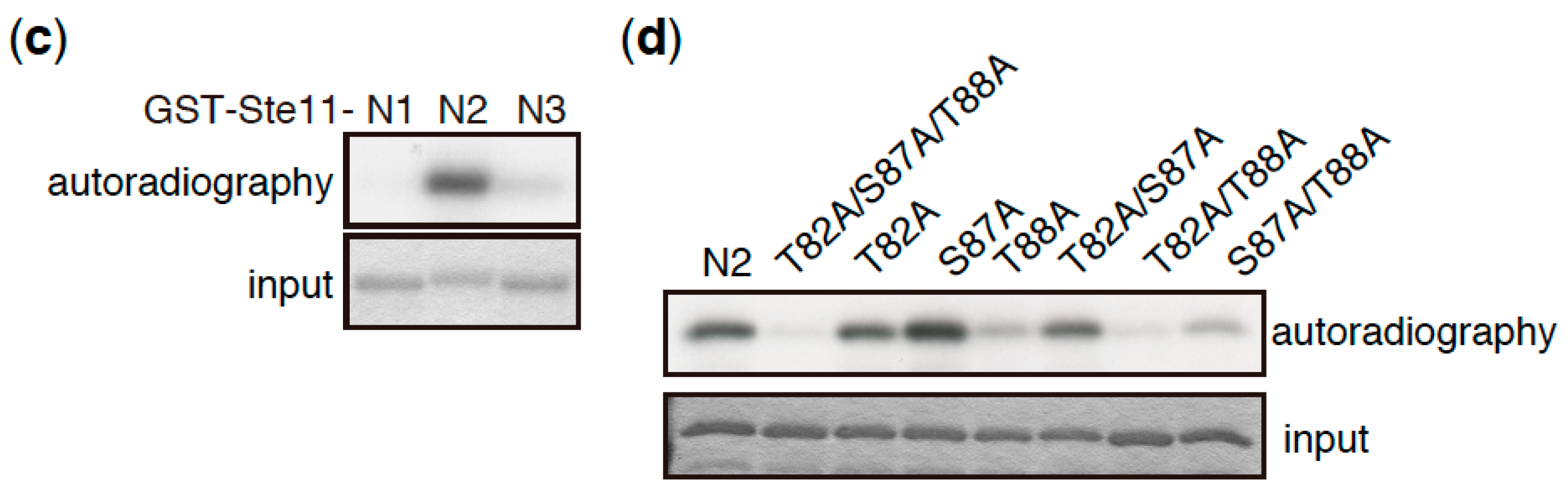
Whereas TORC1 phosphorylates and induces the degradation of Mei2, the deletion of the mei2 gene only partially suppresses the phenotype of tor2 mutant cells, which initiate sexual differentiation under nutrient-rich conditions [58]. This suggests that Mei2 is not the sole TORC1 target that regulates the initiation of sexual differentiation. Consistent with this idea, we found that Ste11, a major transcription factor controlling the switch from cellular proliferation to sexual differentiation in S. pombe, is phosphorylated by TORC1 (Figure 3). Ste11 activates many genes required for the initiation of sexual differentiation, including mei2 [62,63,64]. In vitro kinase assays have demonstrated that the N-terminal fragment of Ste11 (N2, residues 77–183) was phosphorylated by immunoprecipitated TORC1 (Figure 3a–c). An analysis of alanine substitution mutants indicated that Thr88 was a major phosphorylation residue on Ste11, and Thr82 was also phosphorylated, although less efficiently (Figure 3d). It has been shown that the inactivation of TORC1 causes nuclear accumulation of Ste11 [65], suggesting that TORC1 regulates nuclear localization of Ste11 by phosphorylation. Future studies may shed light on the significance of Ste11 phosphorylation by TORC1 with regard to the regulation of sexual differentiation.
4. Indirect Effectors of TORC1
4.1. Ribosomal Protein S6
Ribosomal protein S6 (Rps6) is an evolutionarily conserved protein in eukaryotes and is involved in many cellular processes [68]. In S. pombe, Rps6 is encoded by two genes, rps601 and rps602 [21]. Rps6 is phosphorylated at conserved Ser235 and Ser236 residues by Psk1, a S6k1 homolog, when TORC1 is active [20,21]. The anti-phospho-Akt substrate (PAS) antibody can be used to detect the phosphorylation of Rps6 at these sites [21]. It has been shown that the other TORC1 effector Sck2 is also involved in Rps6 phosphorylation at these residues in a medium-dependent manner, although it is unknown whether Sck2 directly phosphorylates Rps6 [32]. AGC kinase Gad8, which is the major effector of TORC2, has also been shown to contribute to Rps6 phosphorylation [22]. Since the phosphorylation of Rps6 at Ser235 and Ser236 is dispensable for cell viability [21], their phosphorylation might play a role(s) in particular conditions.
4.2. Ppk18 and Igo1
TORC1 negatively regulates the Greatwall-Endosulfine, Ppk18-Igo1, via the inhibition of the Greatwall kinase Ppk18 as mentioned above. The downregulation of Ppk18 has been suggested to be mediated by Sck2, which is a direct effector of TORC1, since it was observed that Sck2 overexpression inhibits Ppk18 activity [29]. It has also been demonstrated that Ppk18-dependent phosphorylation of Igo1 is induced by the reduction of TORC1 activity in tor2 mutant cells, or by the treatment with TOR inhibitors such as rapamycin and Torin1 [29].
4.3. eIF2α
The phosphorylation of eukaryotic initiation factor 2 alpha (eIF2α) causes repression of general protein synthesis in response to various forms of environmental stress. Gcn2 is known to phosphorylate eIF2α in mammals and in the budding yeast S. cerevisiae [69]. In S. cerevisiae, TOR prevents the phosphorylation of eIF2α by Gcn2 [70,71]. Similarly, in S. pombe, Gcn2 phosphorylates eIF2α at serine 52 under TORC1-inactive conditions such as mutation of Tor2 and nitrogen deprivation, while the precise mechanisms underlying Gcn2 activation remain unknown [72]. Serine 52 phosphorylation of S. pombe eIF2α can be detected by the commercially available anti-phospho-eIF2α antibody [72].
4.4. Gaf1
S. pombe TORC1 promotes the phosphorylation of the GATA transcription factor Gaf1 by inhibiting the PP2A-like phosphatase Ppe1 [73]. Phosphorylated Gaf1 is retained in the cytoplasm and does not exert its activity as a transcription factor. When TORC1 is inactivated, Gaf1 is dephosphorylated through the activation of Ppe1 phosphatase and translocates into the nucleus. Thus, the localization of Gaf1 can be a readout for TORC1 activity. Gaf1 positively regulates early response genes under nitrogen stress, such as the 2-oxoglutarate-Fe(II)-dependent oxygenase isp7, and negatively regulates later nitrogen stress genes, such as ste11 [73,74]. The regulation of GATA transcription factors downstream of TORC1 is conserved in S. cerevisiae [75,76,77].
5. Conclusions
In this review, we have overviewed the S. pombe TORC1-dependent phosphorylation targets (Figure 1). Recent studies in S. pombe have revealed a variety of TORC1-related regulatory mechanisms. In particular, S. pombe TORC1 (and TORC2) regulates the change of growth mode in response to environmental conditions, which could be regarded as a simple model of cell differentiation. Hopefully, S. pombe research in the future will bring a new perspective to the study of TOR-mediated cellular regulation.
Since the disruption of gad8, which is phosphorylated and activated by TORC2, results in the same phenotype as the lack of TORC2, such as temperature sensitivity and sterility, Gad8 is thought to be the major TORC2 phosphorylation target [23]. In contrast, the deletion of any TORC1 phosphorylation target fails to phenocopy the TORC1 mutant, suggesting that the key target(s) of TORC1 remains to be identified; nonetheless, we cannot rule out the possibility that TORC1 has multiple important targets and that simultaneous deletion of such factors is required to induce the phenotypes of the TORC1 mutant. In either case, further investigation of TORC1 targets in S. pombe will contribute to the clarification of the essence of the TOR signaling pathway.
Acknowledgments
This work was supported by Japan Society for the Promotion of Science KAKENHI, Grant Numbers 15J40037 and 16K18542 to Y.O., and 15H04333 to A.Y., and by the joint research program of Biosignal Research Center, Kobe University (Grant Number: 281009).
Author Contributions
Y.O. and A.Y. conceived the idea of the review and performed experiments. All the authors contributed to the writing and approved with the final version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Loewith, R.; Jacinto, E.; Wullschleger, S.; Lorberg, A.; Crespo, J.L.; Bonenfant, D.; Oppliger, W.; Jenoe, P.; Hall, M.N. Two TOR complexes, only one of which is rapamycin sensitive, have distinct roles in cell growth control. Mol. Cell 2002, 10, 457–468. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR signaling in growth, metabolism, and disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed]
- Heitman, J.; Movva, N.R.; Hall, M.N. Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 1991, 253, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Loewith, R.; Hall, M.N. Target of rapamycin (TOR) in nutrient signaling and growth control. Genetics 2011, 189, 1177–1201. [Google Scholar] [CrossRef] [PubMed]
- Jacinto, E.; Lorberg, A. TOR regulation of AGC kinases in yeast and mammals. Biochem. J. 2008, 410, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.B.; Dennis, P.B.; Han, J.W.; Williamson, N.A.; Kozma, S.C.; Wettenhall, R.E.; Thomas, G. The principal target of rapamycin-induced p70s6k inactivation is a novel phosphorylation site within a conserved hydrophobic domain. EMBO. J. 1995, 14, 5279–5287. [Google Scholar] [PubMed]
- Burnett, P.E.; Barrow, R.K.; Cohen, N.A.; Snyder, S.H.; Sabatini, D.M. RAFT1 phosphorylation of the translational regulators p70 S6 kinase and 4E-BP1. Proc. Natl. Acad. Sci. USA 1998, 95, 1432–1437. [Google Scholar] [CrossRef] [PubMed]
- Brunn, G.J.; Hudson, C.C.; Sekulic, A.; Williams, J.M.; Hosoi, H.; Houghton, P.J.; Lawrence, J.C., Jr.; Abraham, R.T. Phosphorylation of the translational repressor PHAS-I by the mammalian target of rapamycin. Science 1997, 277, 99–101. [Google Scholar] [CrossRef] [PubMed]
- Gingras, A.C.; Gygi, S.P.; Raught, B.; Polakiewicz, R.D.; Abraham, R.T.; Hoekstra, M.F.; Aebersold, R.; Sonenberg, N. Regulation of 4E-BP1 phosphorylation: A novel two-step mechanism. Genes Dev. 1999, 13, 1422–1437. [Google Scholar] [CrossRef] [PubMed]
- Kawai, M.; Nakashima, A.; Ueno, M.; Ushimaru, T.; Aiba, K.; Doi, H.; Uritani, M. Fission yeast Tor1 functions in response to various stresses including nitrogen starvation, high osmolarity, and high temperature. Curr. Genet. 2001, 39, 166–174. [Google Scholar] [PubMed]
- Weisman, R.; Choder, M. The fission yeast tor homolog, tor1+, is required for the response to starvation and other stresses via a conserved serine. J. Biol. Chem. 2001, 276, 7027–7032. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, B.; Moreno, S. Fission yeast Tor2 promotes cell growth and represses cell differentiation. J. Cell Sci. 2006, 119, 4475–4485. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Hatanaka, M.; Nagao, K.; Nakaseko, Y.; Kanoh, J.; Kokubu, A.; Ebe, M.; Yanagida, M. Rapamycin sensitivity of the Schizosaccharomyces pombe tor2 mutant and organization of two highly phosphorylated TOR complexes by specific and common subunits. Genes Cells 2007, 12, 1357–1370. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Otsubo, Y.; Urano, J.; Tamanoi, F.; Yamamoto, M. Loss of the TOR kinase Tor2 mimics nitrogen starvation and activates the sexual development pathway in fission yeast. Mol. Cell Biol. 2007, 27, 3154–3164. [Google Scholar] [CrossRef] [PubMed]
- Uritani, M.; Hidaka, H.; Hotta, Y.; Ueno, M.; Ushimaru, T.; Toda, T. Fission yeast Tor2 links nitrogen signals to cell proliferation and acts downstream of the Rheb GTPase. Genes Cells 2006, 11, 1367–1379. [Google Scholar] [CrossRef] [PubMed]
- Weisman, R.; Roitburg, I.; Schonbrun, M.; Harari, R.; Kupiec, M. Opposite effects of Tor1 and Tor2 on nitrogen starvation responses in fission yeast. Genetics 2007, 175, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Weisman, R.; Choder, M.; Koltin, Y. Rapamycin specifically interferes with the developmental response of fission yeast to starvation. J. Bacteriol. 1997, 179, 6325–6334. [Google Scholar] [CrossRef] [PubMed]
- Takahara, T.; Maeda, T. TORC1 of fission yeast is rapamycin-sensitive. Genes Cells 2012, 17, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Otsubo, Y.; Yamashita, A.; Sato, T.; Yamamoto, M.; Tamanoi, F. Psk1, an AGC kinase family member in fission yeast, is directly phosphorylated and controlled by TORC1 and functions as S6 kinase. J. Cell Sci. 2012, 125, 5840–5849. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Sato, T.; Tamanoi, F. Fission yeast TORC1 regulates phosphorylation of ribosomal S6 proteins in response to nutrients and its activity is inhibited by rapamycin. J. Cell Sci. 2010, 123, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Halova, L.; Kirkham, S.; Atkin, J.; Petersen, J. TORC2 and the AGC kinase Gad8 regulate phosphorylation of the ribosomal protein S6 in fission yeast. Biol. Open 2012, 1, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Kubo, Y.; Watanabe, Y.; Yamamoto, M. Schizosaccharomyces pombe AGC family kinase Gad8p forms a conserved signaling module with TOR and PDK1-like kinases. EMBO. J. 2003, 22, 3073–3083. [Google Scholar] [CrossRef] [PubMed]
- Kohda, T.A.; Tanaka, K.; Konomi, M.; Sato, M.; Osumi, M.; Yamamoto, M. Fission yeast autophagy induced by nitrogen starvation generates a nitrogen source that drives adaptation processes. Genes Cells 2007, 12, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Madrid, M.; Vazquez-Marin, B.; Franco, A.; Soto, T.; Vicente-Soler, J.; Gacto, M.; Cansado, J. Multiple crosstalk between TOR and the cell integrity MAPK signaling pathway in fission yeast. Sci. Rep. 2016, 6, 37515. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Fujita, M.; Culley, B.M.; Apolinario, E.; Yamamoto, M.; Maundrell, K.; Hoffman, C.S. sck1, a high copy number suppressor of defects in the cAMP-dependent protein kinase pathway in fission yeast, encodes a protein homologous to the Saccharomyces cerevisiae SCH9 kinase. Genetics 1995, 140, 457–467. [Google Scholar] [PubMed]
- Fujita, M.; Yamamoto, M. S. pombe sck2+, a second homologue of S. cerevisiae SCH9 in fission yeast, encodes a putative protein kinase closely related to PKA in function. Curr. Genet. 1998, 33, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Mudge, D.K.; Yang, F.; Currie, B.M.; Kim, J.M.; Yeda, K.; Bashyakarla, V.K.; Ivey, F.D.; Hoffman, C.S. Sck1 negatively regulates Gpa2-mediated glucose signaling in Schizosaccharomyces pombe. Eukaryot. Cell 2014, 13, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Chica, N.; Rozalen, A.E.; Perez-Hidalgo, L.; Rubio, A.; Novak, B.; Moreno, S. Nutritional control of cell size by the greatwall-endosulfine-PP2A.B55 pathway. Curr. Biol. 2016, 26, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Portantier, M.; Chica, N.; Nyquist-Andersen, M.; Mata, J.; Lopez-Aviles, S. A PP2A-B55-mediated crosstalk between TORC1 and TORC2 regulates the differentiation response in fission yeast. Curr. Biol. 2017, 27, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Weston, L.; Greenwood, J.; Nurse, P. Genome-wide screen for cell growth regulators in fission yeast. J. Cell Sci. 2017, 130, 2049–2055. [Google Scholar] [CrossRef] [PubMed]
- Rallis, C.; Lopez-Maury, L.; Georgescu, T.; Pancaldi, V.; Bahler, J. Systematic screen for mutants resistant to TORC1 inhibition in fission yeast reveals genes involved in cellular ageing and growth. Biol. Open 2014, 3, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Michels, A.A. MAF1: A new target of mTORC1. Biochem. Soc. Trans. 2011, 39, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Boguta, M. Maf1, a general negative regulator of RNA polymerase III in yeast. Biochim. Biophys. Acta 2013, 1829, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Lesniewska, E.; Boguta, M. Novel layers of RNA polymerase III control affecting tRNA gene transcription in eukaryotes. Open Biol. 2017, 7, 170001. [Google Scholar] [CrossRef] [PubMed]
- Huber, A.; Bodenmiller, B.; Uotila, A.; Stahl, M.; Wanka, S.; Gerrits, B.; Aebersold, R.; Loewith, R. Characterization of the rapamycin-sensitive phosphoproteome reveals that Sch9 is a central coordinator of protein synthesis. Genes Dev. 2009, 23, 1929–1943. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Moir, R.D.; Willis, I.M. Regulation of RNA polymerase III transcription involves SCH9-dependent and SCH9-independent branches of the target of rapamycin (TOR) pathway. J. Biol. Chem. 2009, 284, 12604–12608. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Tsang, C.K.; Zheng, X.F. Mechanisms of regulation of RNA polymerase III-dependent transcription by TORC1. EMBO. J. 2009, 28, 2220–2230. [Google Scholar] [CrossRef] [PubMed]
- Michels, A.A.; Robitaille, A.M.; Buczynski-Ruchonnet, D.; Hodroj, W.; Reina, J.H.; Hall, M.N.; Hernandez, N. mTORC1 directly phosphorylates and regulates human MAF1. Mol. Cell Biol. 2010, 30, 3749–3757. [Google Scholar] [CrossRef] [PubMed]
- Kantidakis, T.; Ramsbottom, B.A.; Birch, J.L.; Dowding, S.N.; White, R.J. mTOR associates with TFIIIC, is found at tRNA and 5S rRNA genes, and targets their repressor Maf1. Proc. Natl. Acad. Sci. USA 2010, 107, 11823–11828. [Google Scholar] [CrossRef] [PubMed]
- Shor, B.; Wu, J.; Shakey, Q.; Toral-Barza, L.; Shi, C.; Follettie, M.; Yu, K. Requirement of the mTOR kinase for the regulation of Maf1 phosphorylation and control of RNA polymerase III-dependent transcription in cancer cells. J. Biol. Chem. 2010, 285, 15380–15392. [Google Scholar] [CrossRef] [PubMed]
- Papinski, D.; Kraft, C. Regulation of autophagy by signaling through the Atg1/ULK1 complex. J. Mol. Biol. 2016, 428, 1725–1741. [Google Scholar] [CrossRef] [PubMed]
- Kamada, Y.; Funakoshi, T.; Shintani, T.; Nagano, K.; Ohsumi, M.; Ohsumi, Y. Tor-mediated induction of autophagy via an Apg1 protein kinase complex. J. Cell Biol. 2000, 150, 1507–1513. [Google Scholar] [CrossRef] [PubMed]
- Kawamata, T.; Kamada, Y.; Kabeya, Y.; Sekito, T.; Ohsumi, Y. Organization of the pre-autophagosomal structure responsible for autophagosome formation. Mol. Biol. Cell 2008, 19, 2039–2050. [Google Scholar] [CrossRef] [PubMed]
- Kabeya, Y.; Noda, N.N.; Fujioka, Y.; Suzuki, K.; Inagaki, F.; Ohsumi, Y. Characterization of the Atg17-Atg29-Atg31 complex specifically required for starvation-induced autophagy in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2009, 389, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Kamada, Y.; Yoshino, K.; Kondo, C.; Kawamata, T.; Oshiro, N.; Yonezawa, K.; Ohsumi, Y. Tor directly controls the Atg1 kinase complex to regulate autophagy. Mol. Cell Biol. 2010, 30, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Kijanska, M.; Dohnal, I.; Reiter, W.; Kaspar, S.; Stoffel, I.; Ammerer, G.; Kraft, C.; Peter, M. Activation of Atg1 kinase in autophagy by regulated phosphorylation. Autophagy 2010, 6, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, B.; Vacher, C.; Berger, Z.; Davies, J.E.; Luo, S.; Oroz, L.G.; Scaravilli, F.; Easton, D.F.; Duden, R.; O’Kane, C.J.; et al. Inhibition of mTOR induces autophagy and reduces toxicity of polyglutamine expansions in fly and mouse models of Huntington disease. Nat. Genet. 2004, 36, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Ganley, I.G.; Lam du, H.; Wang, J.; Ding, X.; Chen, S.; Jiang, X. ULK1.ATG13.FIP200 complex mediates mTOR signaling and is essential for autophagy. J. Biol. Chem. 2009, 284, 12297–12305. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, N.; Hara, T.; Kaizuka, T.; Kishi, C.; Takamura, A.; Miura, Y.; Iemura, S.; Natsume, T.; Takehana, K.; Yamada, N.; et al. Nutrient-dependent mTORC1 association with the ULK1-Atg13-FIP200 complex required for autophagy. Mol. Biol. Cell 2009, 20, 1981–1991. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Jun, C.B.; Ro, S.H.; Kim, Y.M.; Otto, N.M.; Cao, J.; Kundu, M.; Kim, D.H. ULK-Atg13-FIP200 complexes mediate mTOR signaling to the autophagy machinery. Mol. Biol. Cell 2009, 20, 1992–2003. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, N.; Sasaki, T.; Iemura, S.; Natsume, T.; Hara, T.; Mizushima, N. Atg101, a novel mammalian autophagy protein interacting with Atg13. Autophagy 2009, 5, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.X.; Russell, R.C.; Guan, K.L. Regulation of PIK3C3/VPS34 complexes by MTOR in nutrient stress-induced autophagy. Autophagy 2013, 9, 1983–1995. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Yamamoto, M. S. pombe mei2+ encodes an RNA-binding protein essential for premeiotic DNA synthesis and meiosis I, which cooperates with a novel RNA species meiRNA. Cell 1994, 78, 487–498. [Google Scholar] [CrossRef]
- Watanabe, Y.; Shinozaki-Yabana, S.; Chikashige, Y.; Hiraoka, Y.; Yamamoto, M. Phosphorylation of RNA-binding protein controls cell cycle switch from mitotic to meiotic in fission yeast. Nature 1997, 386, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Watanabe, Y.; Nukina, N.; Yamamoto, M. RNA-assisted nuclear transport of the meiotic regulator Mei2p in fission yeast. Cell 1998, 95, 115–123. [Google Scholar] [CrossRef]
- Otsubo, Y.; Yamashita, A.; Ohno, H.; Yamamoto, M. S. pombe TORC1 activates the ubiquitin-proteasomal degradation of the meiotic regulator Mei2 in cooperation with Pat1 kinase. J. Cell Sci. 2014, 127, 2639–2646. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki-Yabana, S.; Watanabe, Y.; Yamamoto, M. Novel WD-repeat protein Mip1p facilitates function of the meiotic regulator Mei2p in fission yeast. Mol. Cell Biol. 2000, 20, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.H.; Hanson, M.R. The Arabidopsis Mei2 homologue AML1 binds AtRaptor1B, the plant homologue of a major regulator of eukaryotic cell growth. BMC Plant Biol. 2005, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Sebastian, J.; Siddiqi, I. The Arabidopsis-mei2-like genes play a role in meiosis and vegetative growth in Arabidopsis. Plant Cell 2006, 18, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Otsubo, Y.; Yamamoto, M. Signaling pathways for fission yeast sexual differentiation at a glance. J. Cell Sci. 2012, 125, 2789–2793. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, A.; Iino, Y.; Maeda, T.; Watanabe, Y.; Yamamoto, M. Schizosaccharomyces pombe ste11+ encodes a transcription factor with an HMG motif that is a critical regulator of sexual development. Genes Dev. 1991, 5, 1990–1999. [Google Scholar] [CrossRef] [PubMed]
- Mata, J.; Bahler, J. Global roles of Ste11p, cell type, and pheromone in the control of gene expression during early sexual differentiation in fission yeast. Proc. Natl. Acad. Sci. USA 2006, 103, 15517–15522. [Google Scholar] [CrossRef] [PubMed]
- Valbuena, N.; Moreno, S. TOR and PKA pathways synergize at the level of the Ste11 transcription factor to prevent mating and meiosis in fission yeast. PLoS ONE 2010, 5, e11514. [Google Scholar] [CrossRef] [PubMed]
- Aylett, C.H.; Sauer, E.; Imseng, S.; Boehringer, D.; Hall, M.N.; Ban, N.; Maier, T. Architecture of human mTOR complex 1. Science 2016, 351, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Takahara, T.; Hara, K.; Yonezawa, K.; Sorimachi, H.; Maeda, T. Nutrient-dependent multimerization of the mammalian target of rapamycin through the N-terminal HEAT repeat region. J. Biol. Chem. 2006, 281, 28605–28614. [Google Scholar] [CrossRef] [PubMed]
- Meyuhas, O. Physiological roles of ribosomal protein S6: One of its kind. Int. Rev. Cell Mol. Biol. 2008, 268, 1–37. [Google Scholar] [PubMed]
- Baird, T.D.; Wek, R.C. Eukaryotic initiation factor 2 phosphorylation and translational control in metabolism. Adv. Nutr. 2012, 3, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, J.; Staschke, K.A.; Wek, R.C. Dimerization is required for activation of eIF2 kinase Gcn2 in response to diverse environmental stress conditions. J. Biol. Chem. 2004, 279, 22820–22832. [Google Scholar] [CrossRef] [PubMed]
- Cherkasova, V.A.; Hinnebusch, A.G. Translational control by TOR and TAP42 through dephosphorylation of eIF2α kinase GCN2. Genes Dev. 2003, 17, 859–872. [Google Scholar] [CrossRef] [PubMed]
- Valbuena, N.; Rozalen, A.E.; Moreno, S. Fission yeast TORC1 prevents eIF2α phosphorylation in response to nitrogen and amino acids via Gcn2 kinase. J. Cell Sci. 2012, 125, 5955–5959. [Google Scholar] [CrossRef] [PubMed]
- Laor, D.; Cohen, A.; Kupiec, M.; Weisman, R. TORC1 regulates developmental responses to nitrogen stress via regulation of the GATA transcription factor Gaf1. MBio 2015, 6, e00959. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.; Hoe, K.L.; Yu, Y.M.; Yeon, J.H.; Maeng, P.J. The fission yeast GATA factor, Gaf1, modulates sexual development via direct down-regulation of ste11+ expression in response to nitrogen starvation. PLoS ONE 2012, 7, e42409. [Google Scholar] [CrossRef] [PubMed]
- Beck, T.; Hall, M.N. The TOR signalling pathway controls nuclear localization of nutrient-regulated transcription factors. Nature 1999, 402, 689–692. [Google Scholar] [PubMed]
- Cunningham, T.S.; Andhare, R.; Cooper, T.G. Nitrogen catabolite repression of DAL80 expression depends on the relative levels of Gat1p and Ure2p production in Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 14408–14414. [Google Scholar] [CrossRef] [PubMed]
- Georis, I.; Tate, J.J.; Cooper, T.G.; Dubois, E. Nitrogen-responsive regulation of GATA protein family activators Gln3 and Gat1 occurs by two distinct pathways, one inhibited by rapamycin and the other by methionine sulfoximine. J. Biol. Chem. 2011, 286, 44897–44912. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The major downstream pathways of target of rapamycin complex 1 (TORC1) in Schizosaccharomyces pombe. TORC1 phosphorylates multiple targets and regulates cell growth and sexual differentiation. Dashed lines represent pathways in which direct phosphorylation by TORC1 has not been demonstrated.
Figure 1.
The major downstream pathways of target of rapamycin complex 1 (TORC1) in Schizosaccharomyces pombe. TORC1 phosphorylates multiple targets and regulates cell growth and sexual differentiation. Dashed lines represent pathways in which direct phosphorylation by TORC1 has not been demonstrated.
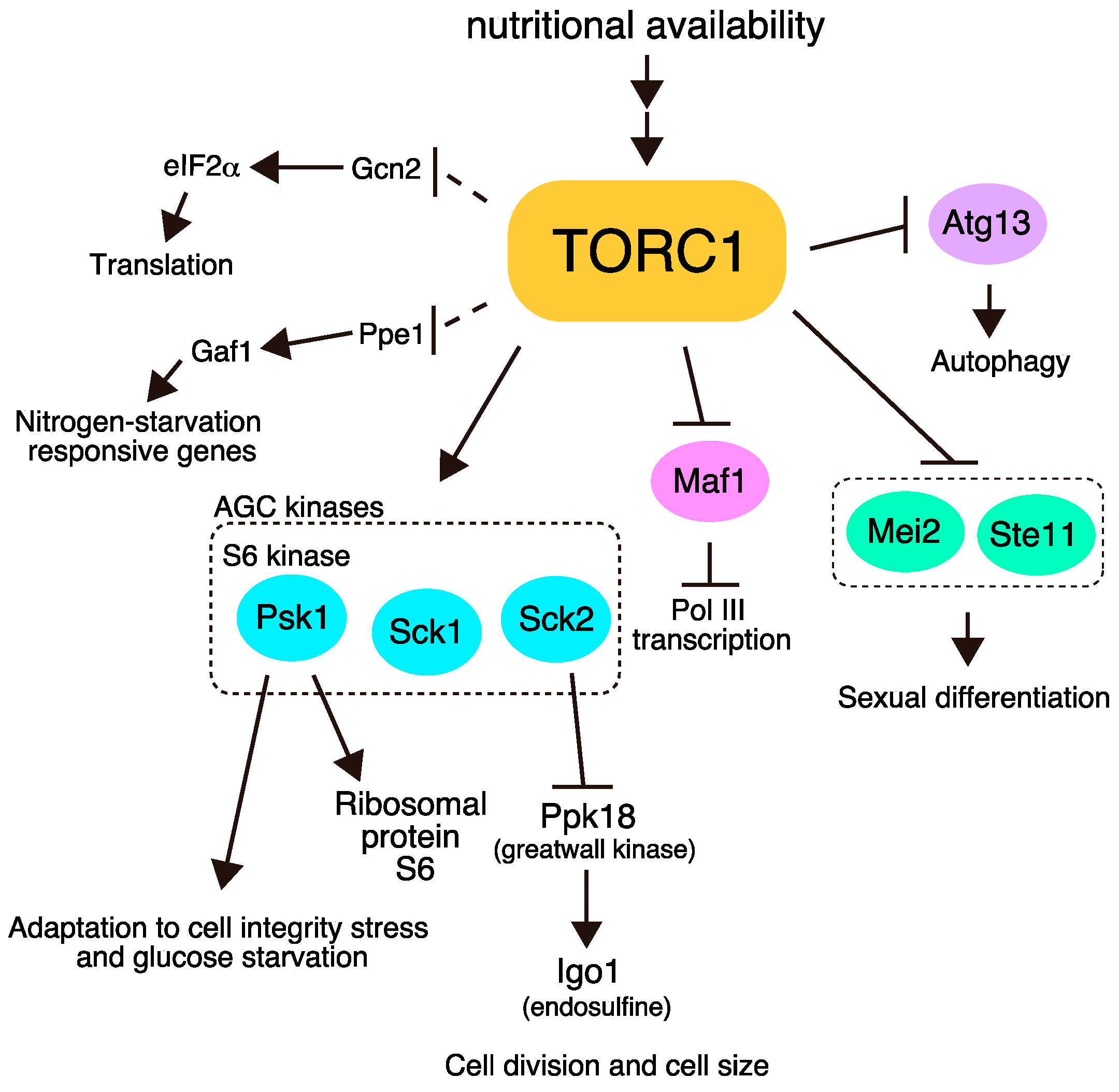
Figure 2.
Phosphorylation of Atg13 and Psk1 is regulated by TORC1 in vivo. Phosphorylation of Atg13 and Psk1 in a temperature-sensitive tor2 mutant. Wild-type (WT; JY3: h90) and tor2-ts6 (JT360: h90 tor2-ts6) cells were grown in YE (Yeast Extract) medium at 25 °C (4 × 106 cells/mL), and then shifted to 30 °C or 34 °C after dilution with the same amount of fresh YE. Cells were disrupted with glass beads in 20% trichloroacetic acid. Cell extracts were subjected to Western blot analysis by using anti-Atg13 antibody (×1000) [24] and anti-phospho-S6 kinase (Thr389) antibody (×1000) (1A5, Cell Signaling Technology, Danvers, MA, USA). Anti-γ-tubulin antibody (GTU-88, Sigma-Aldrich, St. Louis, MO, USA) (×2000) was used as a loading control.
Figure 2.
Phosphorylation of Atg13 and Psk1 is regulated by TORC1 in vivo. Phosphorylation of Atg13 and Psk1 in a temperature-sensitive tor2 mutant. Wild-type (WT; JY3: h90) and tor2-ts6 (JT360: h90 tor2-ts6) cells were grown in YE (Yeast Extract) medium at 25 °C (4 × 106 cells/mL), and then shifted to 30 °C or 34 °C after dilution with the same amount of fresh YE. Cells were disrupted with glass beads in 20% trichloroacetic acid. Cell extracts were subjected to Western blot analysis by using anti-Atg13 antibody (×1000) [24] and anti-phospho-S6 kinase (Thr389) antibody (×1000) (1A5, Cell Signaling Technology, Danvers, MA, USA). Anti-γ-tubulin antibody (GTU-88, Sigma-Aldrich, St. Louis, MO, USA) (×2000) was used as a loading control.

Figure 3.
Ste11 is phosphorylated by TORC1. (a) Schematic illustration of truncation series of Ste11 used in in vitro kinase assays. HMG represents a DNA binding domain, HMG (high mobility group) box. (b–d) In vitro phosphorylation of Ste11 by TORC1. Flag-tagged Tor2 was immunoprecipitated from S. pombe cell extracts using an anti-FLAG antibody. The same immunoprecipitation conditions as described in our previous study were used, in which other components of TORC1 were co-precipitated [15]. The kinase preparation was then incubated with a GST (Glutathione S-transferase)-Ste11 fusion protein, which harbors Ste11 residues 1–469 (Full), 1–230 (N), 231–469 (C), 1–76 (N1), 77–183 (N2), and 184–230 (N3), in the presence of radioactive ATP. Tor2 phosphorylated GST-Ste11-Full and Gst-Ste11-N efficiently, whereas a kinase-dead form of Tor2 (KD) displayed only marginal phosphorylation toward GST-Ste11-Full (b). The exogenously expressed kinase-dead Tor2 in wild-type cells may form a heterodimer with endogenous wild-type Tor2, acquiring a weak activity, as in other organisms [66,67]. Ste11-N2 was also phosphorylated efficiently (c). An alanine substitution series in Ste11-N2 indicated that the major phosphorylation residue was Thr88, while Thr82 was also phosphorylated, but less efficiently (d).
Figure 3.
Ste11 is phosphorylated by TORC1. (a) Schematic illustration of truncation series of Ste11 used in in vitro kinase assays. HMG represents a DNA binding domain, HMG (high mobility group) box. (b–d) In vitro phosphorylation of Ste11 by TORC1. Flag-tagged Tor2 was immunoprecipitated from S. pombe cell extracts using an anti-FLAG antibody. The same immunoprecipitation conditions as described in our previous study were used, in which other components of TORC1 were co-precipitated [15]. The kinase preparation was then incubated with a GST (Glutathione S-transferase)-Ste11 fusion protein, which harbors Ste11 residues 1–469 (Full), 1–230 (N), 231–469 (C), 1–76 (N1), 77–183 (N2), and 184–230 (N3), in the presence of radioactive ATP. Tor2 phosphorylated GST-Ste11-Full and Gst-Ste11-N efficiently, whereas a kinase-dead form of Tor2 (KD) displayed only marginal phosphorylation toward GST-Ste11-Full (b). The exogenously expressed kinase-dead Tor2 in wild-type cells may form a heterodimer with endogenous wild-type Tor2, acquiring a weak activity, as in other organisms [66,67]. Ste11-N2 was also phosphorylated efficiently (c). An alanine substitution series in Ste11-N2 indicated that the major phosphorylation residue was Thr88, while Thr82 was also phosphorylated, but less efficiently (d).
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Otsubo, Y.; Nakashima, A.; Yamamoto, M.; Yamashita, A. TORC1-Dependent Phosphorylation Targets in Fission Yeast. Biomolecules 2017, 7, 50. https://doi.org/10.3390/biom7030050
AMA Style
Otsubo Y, Nakashima A, Yamamoto M, Yamashita A. TORC1-Dependent Phosphorylation Targets in Fission Yeast. Biomolecules. 2017; 7(3):50. https://doi.org/10.3390/biom7030050
Chicago/Turabian StyleOtsubo, Yoko, Akio Nakashima, Masayuki Yamamoto, and Akira Yamashita. 2017. "TORC1-Dependent Phosphorylation Targets in Fission Yeast" Biomolecules 7, no. 3: 50. https://doi.org/10.3390/biom7030050
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.