Calmodulin: A Multitasking Protein in Kv7.2 Potassium Channel Functions



Instituto Biofisika, Consejo Superior de Investigaciones Científicas, CSIC, UPV/EHU, Barrio Sarriena s/n, 48940 Leioa, Spain
*
Authors to whom correspondence should be addressed.
Biomolecules 2018, 8(3), 57; https://doi.org/10.3390/biom8030057
Submission received: 26 June 2018
/
Revised: 13 July 2018
/
Accepted: 14 July 2018
/
Published: 18 July 2018
(This article belongs to the Special Issue Calcium Binding Proteins: Structure, Properties, Functions)
{kind=link}
{kind=link}
Abstract
:The ubiquitous calcium transducer calmodulin (CaM) plays a pivotal role in many cellular processes, regulating a myriad of structurally different target proteins. Indeed, it is unquestionable that CaM is the most relevant transductor of calcium signals in eukaryotic cells. During the last two decades, different studies have demonstrated that CaM mediates the modulation of several ion channels. Among others, it has been indicated that Kv7.2 channels, one of the members of the voltage gated potassium channel family that plays a critical role in brain excitability, requires CaM binding to regulate the different mechanisms that govern its functions. The purpose of this review is to provide an overview of the most recent advances in structure–function studies on the role of CaM regulation of Kv7.2 and the other members of the Kv7 family.
1. Calmodulin: A Ubiquitous Protein Involved in Many Different Functions
Calmodulin (CaM) is the member of EF-hand proteins superfamily which are important Ca2+ sensors in eukaryotic cells that have increasingly received more attention. Initially, CaM was described as an activator of cyclic nucleotide phosphodiesterases in heart and brain extracts [1,2]. However, more recently, it has been recognized for the multitasking nature and the ubiquitous localization of this Ca2+-binding protein that lacks enzymatic activity on its own [3]. CaM is a small (148 aa), soluble, thermostable and acidic protein ubiquitously found in animals, plants, fungi and protozoa, but absent in prokaryotic cells. In mammalians, it is widely distributed in all body tissues [4], being particularly abundant in the brain where its concentration ranges from 1 to 10 μM, or even more (~0.5% of brain proteins) [5,6]. Structurally, CaM presents two similar globular domains, the N-lobe and the C-lobe, connected to each other by a central flexible linker region. Each lobe is composed of two EF-hands, namely α-helix-loop-α-helix motifs, which are responsible for the binding of up to four Ca2+ ions per CaM molecule. Remarkably, the ability of CaM to interact with a large number of proteins is mostly due to its structural flexibility, particularly its’ adaptable binding surfaces [7,8].
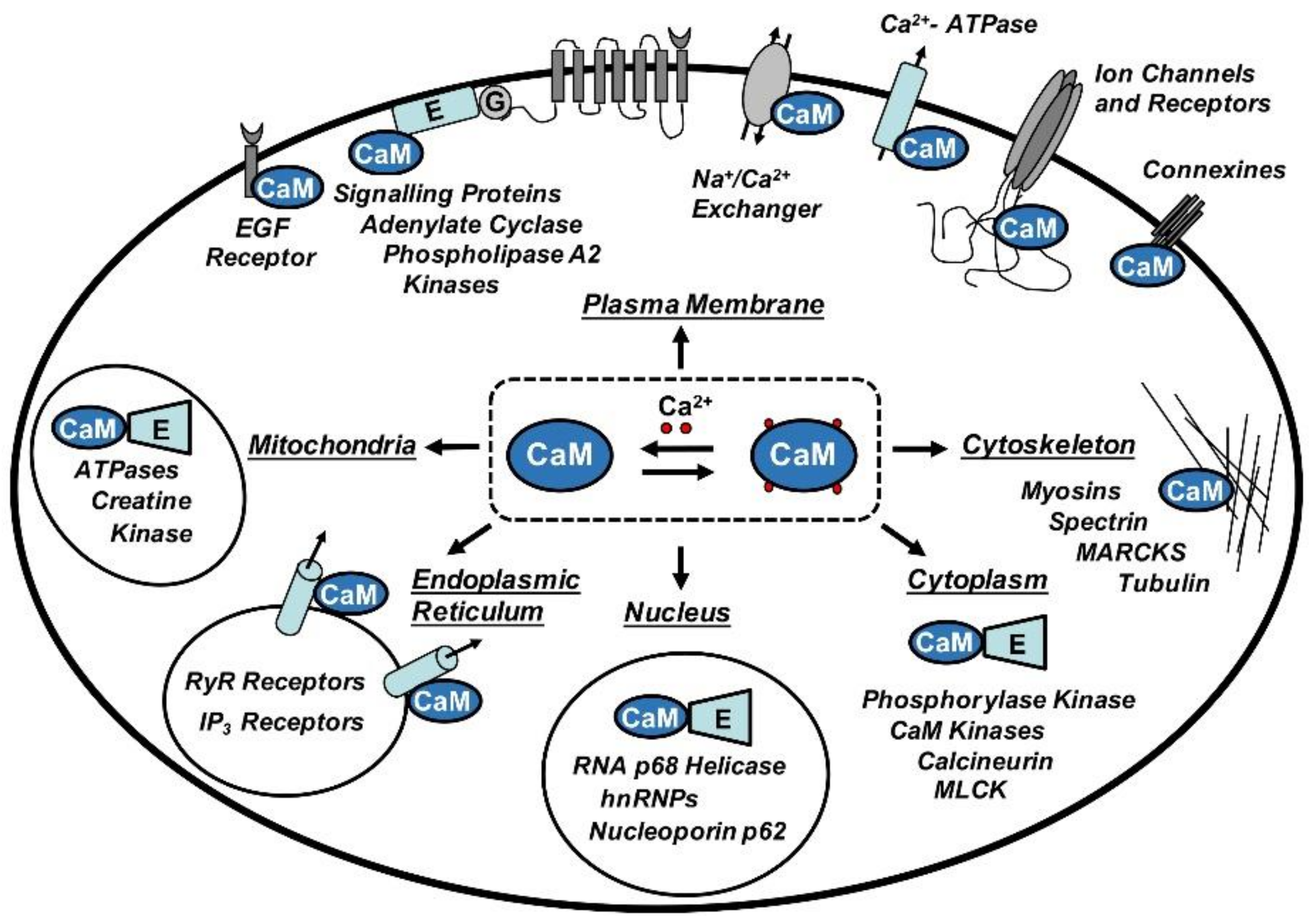
CaM is involved in crucial cellular pathways where it acts as a sensor or transductor of Ca2+ signals by regulating the function of other proteins. Among them, we find enzymes, cytoskeleton proteins, membrane transporters, receptors and ion channels implicated in a plethora of cellular functions, including inflammatory processes and immune response, smooth muscle contraction, cellular division and proliferation, gene expression, hormone and neurotransmitter secretion, apoptosis and others [9] (Figure 1).
In 1978, Brehm and Eckert described for the first time the Ca2+-mediated inhibition of a voltage gated ion channel in Paramecium [10]. Nevertheless, the molecular mechanism underlying this Ca2+-dependent regulation of an ion channel remained obscure until the ‘90s when it was suggested that CaM could mediate the regulation of Ca2+-dependent Na+- and K+-channels in Paramecium [11,12]. Since then, over 40 years later, several papers have demonstrated that CaM plays a pivotal role in the modulation of ion channels and receptors, such as Ca2+-activated K+ channels [13], N-methyl-D-aspartate (NMDA) glutamate receptors [14], cyclic nucleotide-gated ion channels [15], transient receptor potential (TRP) channels [16], voltage-gated Ca2+-, Na+- or K+-channels [17,18,19,20,21] and many others [5].
This review aims to summarize the functional findings together with the different mechanisms of action proposed for the interaction of CaM with Kv7.2 channels. Finally, we will discuss the most recent structure-function studies on the role of CaM regulation of Kv7.2 and the other members of the Kv7 family.
2. Multiple Roles of Calmodulin in the Regulation of Kv7.2 Channels
The human genome contains about 70 genes encoding for K+ channel subunits, being de facto the most diversified family of ion channels. Among them, KCNQ genes encode five Kv7 voltage-gated K+-channels (Kv7.1–Kv7.5). Kv7.1 is principally localized in the heart where it is responsible for the slow potassium current IKs, while the other members of the family (Kv7.2–Kv7.5) are mainly expressed in the nervous system [22,23]. Kv7.2 and Kv7.3 are the main subunits of the low-threshold voltage-gated K+ channel termed “M-channel”, which widely regulates neuronal excitability [24,25,26,27]. Kv7 channels form homo- or hetero-tetramers and each subunit present six transmembrane segments with intracellular N- and C-terminals. Recent cryo-electron microscopy studies by MacKinnon [28] shows that the C-terminal displays four predicted α-helices (ABCD) conserved in all Kv7 family subunits [20]. Atomic structural models have revealed the existence of another helix (TW helix or post-helix A, between helices A and B). Additionally, the extended C-terminal region presents domains that are essential for the interaction with modulatory molecules, such as the membrane phospholipid phosphatidylinositol-4,5-bisphosphate (PIP2), and for the tetrameric assembly [29].
In 2002, two independent laboratories demonstrated that CaM is a binding partner of Kv7 channels [20,21]. The CaM binding domain of Kv7 channels is made up of two discontinuous sites, one located in helix A (hA) and the other in helix B (hB) [20], separated by approximately 135 residues, though another site between hA and hB in Kv7.2 (“TW helix”) could also assist in CaM binding [30]. All the members of Kv7 channels bind CaM [20], however, there is a disagreement about the Ca2+-dependence of this interaction. It has been proposed that Ca2+ does not influence the association of CaM with Kv7.2/7.3 heteromers [21], or that the interaction of Kv7.2 with CaM is weaker [31] or stronger in the absence of Ca2+ [20,32,33,34,35,36]. Furthermore, in vitro assays have demonstrated that apoCaM or Ca2+-CaM can bind peptides containing the sequences of Kv7.2 hA or hB [20,21,33,37,38,39]. In summary, it appears that the strength of the interaction of CaM with Kv7.2 channels is altered by Ca2+ occupancy.
Similar to small conductance Ca2+-activated K+ channels (SK) [13], CaM was first defined as an integral subunit constitutively tethered to the C-terminal region of Kv7.2/3 channels since Kv7.2 mutants that were deficient in CaM binding were unable to generate measurable currents [5,21]. However, this model has been questioned since Kv7.2 channels, carrying a hB mutation [40] or Kv7.4 hA mutated channels [41] that do not bind CaM, can still reach the plasma membrane and are functional. In brief, it appears that the constitutive tethering of CaM is not an absolute requirement for M-channel function.
CaM is essential in the generation of functional M-current (Kv7.2/Kv7.3) in heterologous cells [37] and in neurons [42]. It is also believed to mediate the Ca2+-dependent inhibition [43] of Kv7.2/7.3 heterotetramers by bradykinin or UTP in sympathetic neurons [34,37,44,45]. This “Ca2+ sensor” function influences channel gating in a Kv7 subunit-specific manner, suppressing Kv7.2, Kv7.4 and Kv7.5 currents but stimulating Kv7.1 and IKS channels (Kv7.1 + KCNE1) activity [46,47]. CaM binding to Kv7.1 is also required for appropriate folding of the C-terminus and it is also necessary for correct channel trafficking to the plasma membrane [47,48].
In 2007, our laboratory demonstrated for the first time the crucial role of CaM in Kv7.2 channels trafficking. We found that mutations in hA and hB underlying benign familiar neonatal convulsions (BFNC), an autosomal dominant form of neonatal epilepsy, weakened CaM binding, leading to reduced currents as a consequence of an endoplasmic reticulum (ER) retention of Kv7.2 subunits. Due to this retention, a reduced number of channels reached the plasma membrane [49]. Furthermore, we confirmed the critical role played by CaM in the intracellular transport of Kv7.2 proteins, proposing a model in which CaM needs to adopt an “active” conformation to promote the exit from the ER [32]. Subsequently, this was also observed in hippocampal neurons where CaM regulates the trafficking and the enrichment of Kv7.2/Kv7.3 channels at the axonal surface [50,51].
In the last few years, several studies have demonstrated new and intriguing roles of CaM in the regulation of Kv7.2 channels. These channels, as with all the members of the Kv7 family, necessitate phospholipid PIP2 for their regular function [52,53,54]. Kv7 channels are inhibited after stimulation of Gq- and/or G11-protein-coupled receptors [52,55] as a consequence of the depletion of PIP2 upon activation of phospholipase C [56,57]. Mutations in the Kv7.2 CaM binding domain interfere with the PIP2 activation of the channel [57], suggesting that CaM might compete with PIP2 [58]. Accordingly, recent data indicate the influence of CaM in Kv.7 channels gating by producing changes in the voltage dependence of activation [59,60,61].
Lately, two research laboratories have provided evidence concerning the interconnection between these modulatory molecules in Kv7.2 channels function. The Naoto Hoshi’s group demonstrated that the phosphorylation of CaM, mediated by casein kinase 2, enhanced the binding with Kv7.2, induced resistance to PIP2 depletion, thus, leading to an augmentation on Kv7.2 current amplitude [62]. In accordance with these findings, Gomis-Perez and colleagues found that expression and availability of apoCaM alter the PIP2 regulation of Kv7.2 and Kv7.3 channels [60]. Another paper, performing a live-cell FRET study combined with an electrophysiological-based analysis, provided evidence of a functional connection between CaM binding, PIP2 dependency and the distal coiled-coil tetramerization domain in Kv7.2 channels [35]. Therefore, these data suggest that CaM affects PIP2 sensitivity. Finally, new observations underlined the reciprocal connection between the hAB domain and the coiled-coil module through CaM-mediated regulation of the stability of the distal tetramerization domain of Kv7.2 channels [63].
3. Structure-Function Studies on Calmodulin-Kv7 Channel Complexes
The Protein Data Bank (https://www.rcsb.org/) contains hundreds of structures of CaM, alone or forming complexes with different proteins. Importantly, the number of CaM structures with peptides of ion channels is still growing, advancing our understanding of CaM regulation of these membrane proteins (reviewed by [8,64]).
Concerning Kv7 channels, protein expression for structural studies, demanding huge amounts of soluble and correctly folded proteins, very often has been challenged by the poor solubility of the Kv7 proximal C-termini, resulting in insoluble and aggregated proteins. Currently, a successful approach to obtaining high quality protein samples include the elimination of disposable hydrophobic regions of Kv7 C-terminal likely to be unstructured, along with the co-expression of CaM. This strategy has produced excellent results and is consonant with the assumption that CaM may have a chaperone-like role for Kv7 family C-termini domains [47,48,65,66,67].
Biochemical, functional and structural studies have determined the oligomeric state of the Kv7 hAB domain-CaM complex. The results clearly indicated that CaM binds simultaneously to the hAB sites of Kv7 in a 1:1 stoichiometry. Hence, the complex is modeled as a tetramer with four subunits of Kv7.1 [48,66,68], Kv7.2 [35,67,69] or Kv7.4 [59,70] COOH-terminal and four bound molecules of CaM.
Early biochemical and functional studies have paved the way for understanding how CaM binds these channels. Thanks to a focused research effort, we know that the CaM binding domain is composed by two sites, referred to as hA and hB, which adopt an alpha helical configuration [20,68] and that the binding of CaM can take place on the same or on different subunits of the tetrameric channel [33,66]. Additionally, CaM can bind individually hA or hB, both in the presence or absence of Ca2+ [33]. Mruck and co-workers used an alternative approach to generate an elegant model of the Kv7.2/7.3–CaM complex [71]. In this model, the hAB-CaM complex is located at ~40 Å from the vestibule of the pore, thus, when CaM binds Ca2+, it can easily modulate the channel gating. Our group found that hA of Kv7.2 presents a noticeable preference for the C-lobe, while hB binds more favorably to the N-lobe of CaM [38]. Significantly, this arrangement is also displayed on the crystallographic complexes of CaM with Kv7.1 [66], Kv7.4, Kv7.5 [59] and a chimera between hA of Kv7.3 and hB of Kv7.2 [69]. As described below, this arrangement is also evident in the nuclear magnetic resonance (NMR) Kv7.2hAB-CaM complex [67] and in the cryo-EM structure of Kv7.1 [28].
In the last five years, the complex formed by CaM and Kv7 family subunits has been explored using X-ray crystallography [59,66,69,70], NMR spectroscopy [67] and cryo-electron microscopy [28] (Figure 2).
The X-ray crystallographic structure of Ca2+-CaM/Kv7.4 hB resembles the classical CaM 1-14 binding motif [72], with one molecule of Ca2+-CaM wrapping around an α-helix, namely the Kv7.4 hB segment. Likewise, in this structure, of the two CaM lobes, the N-lobe establishes more contacts with hB [70]. The results obtained by Sachyani and co-workers revealed that CaM hugs hA and hB of Kv7.1 channels, with the apo-C lobe bound to hA and the Ca2+-loaded N-lobe associated with hB [66]. Strikingly, this conformation is similar to that of the C-lobe of CaM-SK2 channel complex [73], which underscore structural analogies between Kv7 and SK channels. Lately, single-particle cryo-electron microscopy was used to determine the whole structure of Kv7.1 in complex with CaM [28]. Using this powerful tool, MacKinnon and colleagues have uncovered a novel interaction of CaM with the intracellular S2–S3 linker, providing evidence of an alternative functional coupling between the voltage sensor and the pore of Kv7.1 channels [28].
Two recent studies have determined the structures of Kv7.2-CaM complexes. Firstly, Hirsch and co-workers solved the crystal structure of the chimera Kv7.3hA-Kv7.2hB complexed with CaM at high Ca2+ concentration [69]. The complex displays hA bound to the CaM C-lobe, whilst Kv7.2 hB interacts principally with the N-lobe. The two helices form an antiparallel coiled-coil that resembles the Kv7.1 hAB domain described above [66]. Finally, the association of hAB domain of Kv7.2 with CaM was characterized in solution using NMR spectroscopy [67]. The authors of this study reported the structures of the CaM-Kv7.2hAB at two different Ca2+ concentrations: normal cytosolic Ca2+ concentrations (<100 nM) are able to calcify only the CaM N-lobe, while when Ca2+ levels increase (>1 μM), the C-lobe also can bind this cation. Precisely, the binding of Ca2+ to the C-lobe and the subsequent conformational rearrangement could be the structural signal responsible for modulation of channel gating. As in the previous structures, CaM wraps the Kv7.2 helices, in particular, hA binds the apoC-lobe whilst hB interacts with the calcified N-lobe. In conclusion, CaM-Kv7.2hAB structure is strikingly similar to the CaM-Kv7.1hAB, CaM-Kv7.3hA/Kv7.2hB and CaM-Kv7.4hAB or Kv7.5hAB complexes [28,59,66,69], suggesting a common, conserved CaM-mediated mechanism for transduction of Ca2+ signals among the members of the Kv7 family.
4. Conclusions
During the past 40 years, the number of ion channels found to be modulated by CaM has increased dramatically. Today, several models and accurate structures of ion channels in complex with CaM are available, improving our knowledge about the regulation of these membrane proteins.
In light of the latest progress in structure-function studies, we have summarized the most recent knowledge of Kv7 channels regulation by CaM, emphasizing numerous fundamental aspects, for which the explanation was previously hindered by the absence of structural data. In conclusion, we are confident that the recent progress on the structure-function of Kv7-CaM complexes will help to identify and evaluate the comprehensive mechanisms for mutation-induced Kv7 channel dysfunctions, such as epilepsy and long QT (LQT) syndrome.
Author Contributions
A.A. and A.V. designed the work, wrote and revised the manuscript.
Funding
This research was funded by grants from the Department of Industry, Tourism and Trade of the Government of the Autonomous Community of the Basque Country (Elkartek BG2015) and from the Spanish Ministry of Economy, Industry and Competitiveness (BFU2015-66910-R and CSD2008-00005).
Acknowledgments
We would like to thank Juncal Fernández-Orth for editing and critical reading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cheung, W.Y. Cyclic 3′,5′-nucleotide phosphodiesterase. Demonstration of an activator. Biochem. Biophys. Res. Commun. 1970, 38, 533–538. [Google Scholar] [CrossRef]
- Kakiuchi, S.; Yamazaki, R. Calcium dependent phosphodiesterase activity and its activating factor (PAF) from brain studies on cyclic 3′,5′-nucleotide phosphodiesterase (3). Biochem. Biophys. Res. Commun. 1970, 41, 1104–1110. [Google Scholar] [CrossRef]
- Jurado, L.A.; Chockalingam, P.S.; Jarrett, H.W. Apocalmodulin. Physiol. Rev. 1999, 79, 661–682. [Google Scholar] [CrossRef] [PubMed]
- Chin, D.; Mens, A.R. Calmodulin: A prototypical calcium sensor. Trends Cell Biol. 2000, 10, 322–328. [Google Scholar] [CrossRef]
- Saimi, Y.; Kung, C. Calmodulin as an ion channel subunit. Annu. Rev. Physiol. 2002, 64, 289–311. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Storm, D.R. The role of calmodulin as a signal integrator for synaptic plasticity. Nat. Rev. Neurosci. 2005, 6, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Yamniuk, A.P.; Vogel, H.J. Calmodulin’s flexibility allows for promiscuity in its interactions with target proteins and peptides. Mol. Biotechnol. 2004, 27, 33–57. [Google Scholar] [CrossRef]
- Villarroel, A.; Taglialatela, M.; Bernardo-Seisdedos, G.; Alaimo, A.; Agirre, J.; Alberdi, A.; Gomis-Perez, C.; Soldovieri, M.V.; Ambrosino, P.; Malo, C.; et al. The ever changing moods of calmodulin: How structural plasticity entails transductional adaptability. J. Mol. Biol. 2014, 426, 2717–2735. [Google Scholar] [CrossRef] [PubMed]
- Vetter, S.W.; Leclerc, E. Novel aspects of calmodulin target recognition and activation. Eur. J. Biochem. 2003, 270, 404–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brehm, P.; Eckert, R. Calcium entry leads to inactivation of calcium channel in Paramecium. Science 1978, 202, 1203–1206. [Google Scholar] [CrossRef] [PubMed]
- Kink, J.A.; Maley, M.E.; Preston, R.R.; Ling, K.Y.; Wallen-Friedman, M.A.; Saimi, Y.; Kung, C. Mutations in paramecium calmodulin indicate functional differences between the C-terminal and N-terminal lobes in vivo. Cell 1990, 62, 165–174. [Google Scholar] [CrossRef]
- Saimi, Y.; Kung, C. Ion channel regulation by calmodulin binding. FEBS Lett. 1994, 350, 155–158. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.M.; Fakler, B.; Rivard, A.; Wayman, G.; Johanson-Pais, T.; Keen, J.E.; Ishii, T.; Hirschber, B.; Bond, C.T.; Lutsenko, S.; et al. Mechanism of calcium gating in small-conductance calcium-activated potassium channels. Nature 1998, 395, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, M.D.; Zhang, S.; Bernhardt, J.P.; Huganir, R.L. Inactivation of NMDA receptors by direct interaction of calmodulin with the NR1 subunit. Cell 1996, 84, 745–755. [Google Scholar] [CrossRef]
- Liu, M.; Chen, T.Y.; Ahamed, B.; Li, J.; Yau, K.W. Calcium-calmodulin modulation of the olfactory cyclic nucleotide-gated cation channel. Science 1994, 266, 1348–1354. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.M.; Bull, A.; Kelly, LE. Identification of a Drosophila gene encoding a calmodulin-binding protein with homology to the trp phototransduction gene. Neuron 1992, 8, 631–642. [Google Scholar] [CrossRef]
- Zühlke, R.D.; Pitt, G.S.; Deisseroth, K.; Tsien, R.W.; Reuter, H. Calmodulin supports both inactivation and facilitation of L-type calcium channels. Nature 1999, 399, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.Z.; DeMaria, C.D.; Adelman, J.P.; Yue, D.T. Calmodulin is the Ca2+ sensor for Ca2+-dependent inactivation of L-type calcium channels. Neuron 1999, 22, 549–558. [Google Scholar] [CrossRef]
- Mori, M.; Konno, T.; Ozawa, T.; Murata, M.; Imoto, K.; Nagayama, K. Novel interaction of the voltage-dependent sodium channel (VDSC) with calmodulin: Does VDSC acquire calmodulin-mediated Ca2+-sensitivity? Biochemistry 2000, 39, 1316–1323. [Google Scholar] [CrossRef] [PubMed]
- Yus-Nájera, E.; Santana-Castro, I.; Villarroel, A. The identification and characterization of a noncontinuous calmodulin-binding site in noninactivating voltage-dependent KCNQ potassium channels. J. Biol. Chem. 2002, 277, 28545–28553. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Levitan, I.B. Calmodulin is an auxiliary subunit of KCNQ2/3 potassium channels. J. Neurosci. 2002, 22, 7991–8001. [Google Scholar] [CrossRef] [PubMed]
- Jentsch, T.J. Neuronal KCNQ potassium channels: Physiology and role in disease. Nat. Rev. Neurosci. 2000, 1, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J. KCNQ potassium channels: Physiology, pathophysiology, and pharmacology. Pharmacol. Ther. 2001, 90, 1–19. [Google Scholar] [CrossRef]
- Brown, D.A.; Adams, P.R. Muscarinic suppression of a novel voltage-sensitive K+ current in a vertebrate neurone. Nature 1980, 283, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A. M currents. Ion. Channels 1988, 1, 55–94. [Google Scholar] [PubMed]
- Marrion, N.V. Control of M-current. Annu. Rev. Physiol. 1997, 59, 483–504. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.S.; Pan, Z.; Shi, W.; Brown, B.S.; Wymore, R.S.; Cohen, I.S.; Dixon, J.E.; McKinnon, D. KCNQ2 and KCNQ3 potassium channel subunits: Molecular correlates of the M-channel. Science 1998, 282, 1890–1893. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; MacKinnon, R. Cryo-EM Structure of a KCNQ1/CaM Complex Reveals Insights into Congenital Long QT Syndrome. Cell 2017, 169, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Haitin, Y.; Attali, B. The C-terminus of Kv7 channels: A multifunctional module. J. Physiol. 2008, 586, 1803–1810. [Google Scholar] [CrossRef] [PubMed]
- Gomis-Perez, C.; Alaimo, A.; Fernandez-Orth, J.; Alberdi, A.; Aivar-Mateo, P.; Bernardo-Seisdedos, G.; Malo, C.; Areso, P.; Felipe, A.; Villarroel, A. Unconventional calmodulin anchoring site within the AB module of Kv7.2 channels. J. Cell Sci. 2015, 128, 3155–3163. [Google Scholar] [CrossRef] [PubMed]
- Bal, M.; Zaika, O.; Martin, P.; Shapiro, M.S. Calmodulin binding to M-type K+ channels assayed by TIRF/FRET in living cells. J. Physiol. 2008, 586, 2307–2320. [Google Scholar] [CrossRef] [PubMed]
- Alaimo, A.; Gomez-Posada, J.C.; Aivar, P.; Etxeberria, A.; Rodriguez-Alfaro, J.A.; Areso, P.; Villarroel, A. Calmodulin activation limits the rate of KCNQ2 K+ channel exit from the endoplasmic reticulum. J. Biol. Chem. 2009, 284, 20668–20675. [Google Scholar] [CrossRef] [PubMed]
- Alaimo, A.; Alberdi, A.; Gomis-Perez, C.; Fernandez-Orth, J.; Gomez-Posada, J.C.; Areso, P.; Villarroel, A. Cooperativity between calmodulin-binding sites in Kv7.2 channels. J. Cell Sci. 2013, 126, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Kosenko, A.; Hoshi, N. A Change in Configuration of the Calmodulin-KCNQ Channel Complex Underlies Ca2+-Dependent Modulation of KCNQ Channel Activity. PLoS ONE 2013, 8, e82290. [Google Scholar] [CrossRef] [PubMed]
- Alberdi, A.; Gomis-Perez, C.; Bernardo-Seisdedos, G.; Alaimo, A.; Malo, C.; Aldaregia, J.; Lopez-Robles, C.; Areso, P.; Butz, E.; Wahl-Schott, C.; et al. Uncoupling PIP2-calmodulin regulation of Kv7.2 channels by an assembly de-stabilizing epileptogenic mutation. J. Cell Sci. 2015, 128, 4014–4023. [Google Scholar] [CrossRef] [PubMed]
- Ambrosino, P.; Alaimo, A.; Bartollino, S.; Manocchio, L.; DeMaria, M.; Mosca, I.; Gomis-Perez, C.; Alberdi, A.; Scambia, G.; Lesca, G.; et al. Epilepsy-causing mutations in Kv7.2 C-terminus affect binding and functional modulation by calmodulin. Biochim. Biophys. Acta 2015, 1852, 1856–1866. [Google Scholar] [CrossRef] [PubMed]
- Gamper, N.; Shapiro, M.S. Calmodulin mediates Ca2+-dependent modulation of M-type K+ channels. J. Gen. Physiol. 2003, 122, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Alaimo, A.; Alberdi, A.; Gomis-Perez, C.; Fernandez-Orth, J.; Bernardo-Seisdedos, G.; Malo, C.; Millet, O.; Areso, P.; Villarroel, A. Pivoting between Calmodulin Lobes Triggered by Calcium in the Kv7.2/Calmodulin Complex. PLoS ONE 2014, 9, e86711. [Google Scholar] [CrossRef] [PubMed]
- Bonache, M.A.; Alaimo, A.; Malo, C.; Millet, O.; Villarroel, A.; González-Muñiz, R. Clicked bis-PEG-Peptide Conjugated for Studying Calmodulin-Kv7.2 Channel Binding. Org. Biomol. Chem. 2014, 12, 8877–8887. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Posada, J.C.; Aivar, P.; Alberdi, A.; Alaimo, A.; Etxeberria, A.; Fernandez-Orth, J.; Zamalloa, T.; Roura-Ferrer, M.; Villace, P.; Areso, P.; et al. Kv7 Channels Can Function without Constitutive Calmodulin Tethering. PLoS ONE 2011, 6, e25508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sihn, C.R.; Kim, H.J.; Woltz, R.L.; Yarov-Yarovoy, V.; Yang, P.C.; Xu, J.; Clancy, C.E.; Zhang, X.D.; Chiamvimonvat, N.; Yamoah, E.N. Mechanisms of Calmodulin Regulation of Different Isoforms of Kv7.4 K+ Channels. J. Biol. Chem. 2016, 291, 2499–2509. [Google Scholar] [CrossRef] [PubMed]
- Shahidullah, M.; Santarelli, L.C.; Wen, H.; Levitan, I.B. Expression of a calmodulin-binding KCNQ2 potassium channel fragment modulates neuronal M-current and membrane excitability. Proc. Natl. Acad. Sci. USA 2005, 102, 16454–16459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selyanko, A.A.; Brown, D.A. Intracellular calcium directly inhibits potassium M channels in excised membrane patches from rat sympathetic neurons. Neuron 1996, 16, 151–162. [Google Scholar] [CrossRef]
- Cruzblanca, H.; Koh, D.S.; Hille, B. Bradykinin inhibits M current via phospholipase C and Ca2+ release from IP3-sensitive Ca2+ stores in rat sympathetic neurons. Proc. Natl. Acad. Sci. USA 1998, 95, 7151–7156. [Google Scholar] [CrossRef] [PubMed]
- Zaika, O.; Tolstykh, G.P.; Jaffe, D.B.; Shapiro, M.S. Inositol triphosphate-mediated Ca2+ signals direct purinergic P2Y receptor regulation of neuronal ion channels. J. Neurosci. 2007, 27, 8914–8926. [Google Scholar] [CrossRef] [PubMed]
- Gamper, N.; Li, Y.; Shapiro, M.S. Structural requirements for differential sensitivity of KCNQ K+ channels to modulation by Ca2+/calmodulin. Mol. Biol. Cell 2005, 16, 3538–3551. [Google Scholar] [CrossRef] [PubMed]
- Shamgar, L.; Ma, L.J.; Schmitt, N.; Haitin, Y.; Peretz, A.; Wiener, R.; Hirsch, J.; Pongs, O.; Attali, B. Calmodulin is essential for cardiac IKS channel gating and assembly—Impaired function in long-QT mutations. Circ. Res. 2006, 98, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Nunziato, D.A.; Pitt, G.S. KCNQ1 assembly and function is blocked by long-QT syndrome mutations that disrupt interaction with calmodulin. Circ. Res. 2006, 98, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Etxeberria, A.; Aivar, P.; Rodriguez-Alfaro, J.A.; Alaimo, A.; Villace, P.; Gomez-Posada, J.C.; Areso, P.; Villarroel, A. Calmodulin regulates the trafficking of KCNQ2 potassium channels. FASEB J. 2008, 22, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Cavaretta, J.P.; Sherer, K.R.; Lee, K.Y.; Kim, E.H.; Issema, R.S.; Chung, H.J. Polarized axonal surface expression of neuronal KCNQ potassium channels is regulated by calmodulin interaction with KCNQ2 subunit. PLoS ONE 2014, 9, e103655. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Devaux, J.J. Calmodulin orchestrates the heteromeric assembly and the trafficking of KCNQ2/3 (Kv7.2/3) channels in neurons. Mol. Cell Neurosci. 2014, 58, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Delmas, P.; Brown, D.A. Pathways modulating neural KCNQ/M (Kv7) potassium channels. Nat. Rev. Neurosci. 2005, 6, 850–862. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gamper, N.; Hilgemann, D.W.; Shapiro, M.S. Regulation of Kv7 (KCNQ) K+ channel open probability by phosphatidylinositol 4,5-bisphosphate. J. Neurosci. 2005, 25, 9825–9835. [Google Scholar] [CrossRef] [PubMed]
- Suh, B.C.; Inoue, T.; Meyer, T.; Hille, B. Rapid chemically induced changes of Ptdlns(4,5)P2 gate KCNQ ion channels. Science 2006, 314, 1454–1457. [Google Scholar] [CrossRef] [PubMed]
- Selyanko, A.A.; Hadley, J.K.; Wood, I.C.; Abogadie, F.C.; Jentsch, T.J.; Brown, D.A. Inhibition of KCNQ1-4 potassium channels expressed in mammalian cells via M1 muscarinic acetylcholine receptors. J. Physiol. 2000, 522, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Suh, B.C.; Hille, B. Recovery from muscarinic modulation of M current channels requires phosphatidylinositol 4,5-bisphosphate synthesis. Neuron 2002, 35, 507–520. [Google Scholar] [CrossRef]
- Zhang, H.L.; Craciun, L.C.; Mirshahi, T.; Rohacs, T.; Lopes, C.M.B.; Jin, T.H.; Logothetis, D.E. PIP2 activates KCNQ channels, and its hydrolysis underlies receptor-mediated inhibition of M currents. Neuron 2003, 37, 963–975. [Google Scholar] [CrossRef]
- Tobelaim, W.S.; Dvir, M.; Lebel, G.; Cui, M.; Buki, T.; Peretz, A.; Marom, M.; Haitin, Y.; Logothetis, D.E.; Hirsch, J.A.; et al. Ca2+-Calmodulin and PIP2 interactions at the proximal C-terminus of Kv7 channels. Channels 2017, 11, 686–695. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.; Abderemane-Ali, F.; Hura, G.L.; Rossen, N.D.; Gate, R.E.; Minor, D.L., Jr. A Calmodulin C-Lobe Ca2+-Dependent Switch Governs Kv7 Channel Function. Neuron 2018, 97, 836–852. [Google Scholar] [CrossRef] [PubMed]
- Gomis-Perez, C.; Soldovieri, M.V.; Malo, C.; Ambrosino, P.; Taglialatela, M.; Areso, P.; Villarroel, A. Differential Regulation of PI(4,5)P2 Sensitivity of Kv7.2 and Kv7.3 Channels by Calmodulin. Front. Mol. Neurosci. 2017, 10, 117. [Google Scholar] [CrossRef] [PubMed]
- Tobelaim, W.S.; Dvir, M.; Lebel, G.; Cui, M.; Buki, T.; Peretz, A.; Marom, M.; Haitin, Y.; Logothetis, D.E.; Hirsch, J.A.; et al. Competition of calcified calmodulin N lobe and PIP2 to an LQT mutation site in Kv7.1 channel. Proc. Natl. Acad. Sci. USA 2017, 114, E869–E878. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Xu, M.; Cooper, E.C.; Hoshi, N. Channel anchored protein kinase CK2 and protein phosphatase 1 reciprocally regulate KCNQ2-containing M-channels via phosphorylation of calmodulin. J. Biol. Chem. 2014, 289, 11536–11544. [Google Scholar] [CrossRef] [PubMed]
- Alaimo, A.; Nuñez, E.; Aivar, P.; Fernández-Orth, J.; Gomis-Perez, C.; Bernardo-Seisdedos, G.; Malo, C.; Villarroel, A. Calmodulin confers calcium sensitivity to the stability of the distal intracellular assembly domain of Kv7.2 channels. Sci. Rep. 2017, 7, 13425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalevskaya, N.V.; van de Waterbeemd, M.; Bokhovchuk, F.M.; Bate, N.; Bindels, R.J.; Hoenderop, J.G.; Vuister, G.W. Structural analysis of calmodulin binding to ion channels demonstrates the role of its plasticity in regulation. Pflugers Arch. 2013, 465, 1507–1519. [Google Scholar] [CrossRef] [PubMed]
- Etzioni, A.; Siloni, S.; Chikvashvilli, D.; Strulovich, R.; Sachyani, D.; Regev, N.; Greitzer-Antes, D.; Hirsch, J.A.; Lotan, I. Regulation of neuronal M-channel gating in an isoform-specific manner: Functional interplay between calmodulin and syntaxin 1A. J. Neurosci. 2011, 31, 14158–14171. [Google Scholar] [CrossRef] [PubMed]
- Sachyani, D.; Dvir, M.; Strulovich, R.; Tria, G.; Tobelaim, W.; Peretz, A.; Pongs, O.; Svergun, D.; Attali, B.; Hirsch, J.A. Structural basis of a Kv7.1 potassium channel gating module: Studies of the intracellular C-terminal domain in complex with calmodulin. Structure 2014, 22, 1582–1594. [Google Scholar] [CrossRef] [PubMed]
- Bernardo-Seisdedos, G.; Nunez, E.; Gomis, C.; Malo, C.; Villarroel, A.; Millet, O. Structural basis and energy landscape for the Ca2+ gating and calmodulation of the Kv7.2 K+ channel. Proc. Natl. Acad. Sci. USA 2018, 115, 2395–2400. [Google Scholar] [CrossRef] [PubMed]
- Wiener, R.; Haitin, Y.; Shamgar, L.; Fernandez-Alonso, M.C.; Martos, A.; Chomsky-Hecht, O.; Rivas, G.; Attali, B.; Hirsch, J.A. The KCNQ1 (Kv7.1) COOH terminus, a multitiered scaffold for subunit assembly and protein interaction. J. Biol. Chem. 2008, 283, 5815–5830. [Google Scholar] [CrossRef] [PubMed]
- Strulovich, R.; Tobelaim, W.S.; Attali, B.; Hirsch, J.A. Structural Insights into the M-Channel Proximal C-Terminus/Calmodulin Complex. Biochemistry 2016, 55, 5353–5365. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Chang, A.; Tolia, A.; Minor, D.L., Jr. Structure of a Ca(2+)/CaM:Kv7.4 (KCNQ4) B-helix complex provides insight into M current modulation. J. Mol. Biol. 2013, 425, 378–394. [Google Scholar] [CrossRef] [PubMed]
- Mruk, K.; Shandilya, S.M.; Blaustein, R.O.; Schiffer, C.A.; Kobertz, W.R. Structural insights into neuronal K+ channel-calmodulin complexes. Proc. Natl. Acad. Sci. USA 2012, 109, 13579–13583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhoads, A.R.; Friedberg, F. Sequence motifs for calmodulin recognition. FASEB J. 1997, 11, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.A.; Rivard, A.F.; Bächinger, H.P.; Adelman, J.A. Structure of the gating domain of a Ca2+-activated K+ channel complexed with Ca2+/calmodulin. Nature 2001, 410, 1120–1124. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The central role played by calmodulin (CaM) in the regulation of a wide spectrum of binding partners. Abbreviations: hnRNPs: Heterogeneous nuclear ribonucleoproteins; MARCKS: Myristoylated alanine-rich C-kinase substrate; MLCK: Myosin light-chain kinase.
Figure 1.
The central role played by calmodulin (CaM) in the regulation of a wide spectrum of binding partners. Abbreviations: hnRNPs: Heterogeneous nuclear ribonucleoproteins; MARCKS: Myristoylated alanine-rich C-kinase substrate; MLCK: Myosin light-chain kinase.

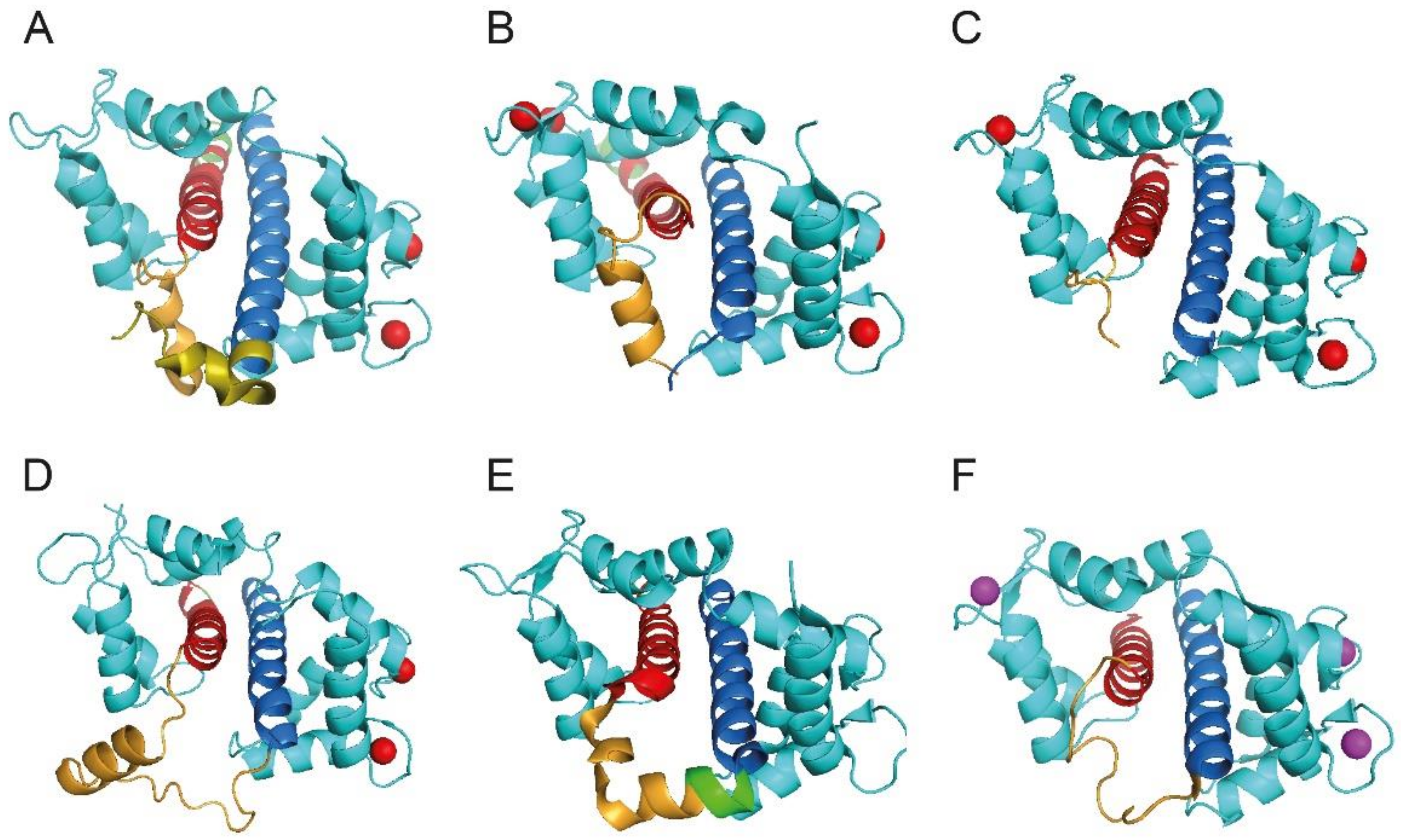
Figure 2.
CaM/Kv7 complexes. Ribbon representation of CaM in complex with AB helices of Kv7 channels. CaM is colored turquoise, the helices A are red, the helices B are blue, the “TW domains” are in dark orange and, finally, Ca2+ and Mg2+ ions are red and magenta, respectively. Structural data were obtained from the Protein Data Bank (PDB), accession codes: (A) CaM/Kv 7.1hAB (4V0C, [66]); (B) CaM/Kv7.3hA-Kv7.2hB (5J03, [69]); (C) CaM/Kv7.1hAB (5VMS, [28]); (D) CaM/Kv7.2hAB (6FEG, [67]); (E) CaM/Kv7.4hAB (6B8L, [59]); (F) CaM/Kv7.5hAB (6B8Q, [59]). Panels A and C were simplified to show only the CaM/hAB complexes. Figures were produced with PyMOL (The PyMOL Molecular Graphics System, Version 1.5.0.4 Schrödinger, LLC.).
Figure 2.
CaM/Kv7 complexes. Ribbon representation of CaM in complex with AB helices of Kv7 channels. CaM is colored turquoise, the helices A are red, the helices B are blue, the “TW domains” are in dark orange and, finally, Ca2+ and Mg2+ ions are red and magenta, respectively. Structural data were obtained from the Protein Data Bank (PDB), accession codes: (A) CaM/Kv 7.1hAB (4V0C, [66]); (B) CaM/Kv7.3hA-Kv7.2hB (5J03, [69]); (C) CaM/Kv7.1hAB (5VMS, [28]); (D) CaM/Kv7.2hAB (6FEG, [67]); (E) CaM/Kv7.4hAB (6B8L, [59]); (F) CaM/Kv7.5hAB (6B8Q, [59]). Panels A and C were simplified to show only the CaM/hAB complexes. Figures were produced with PyMOL (The PyMOL Molecular Graphics System, Version 1.5.0.4 Schrödinger, LLC.).

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alaimo, A.; Villarroel, A. Calmodulin: A Multitasking Protein in Kv7.2 Potassium Channel Functions. Biomolecules 2018, 8, 57. https://doi.org/10.3390/biom8030057
AMA Style
Alaimo A, Villarroel A. Calmodulin: A Multitasking Protein in Kv7.2 Potassium Channel Functions. Biomolecules. 2018; 8(3):57. https://doi.org/10.3390/biom8030057
Chicago/Turabian StyleAlaimo, Alessandro, and Alvaro Villarroel. 2018. "Calmodulin: A Multitasking Protein in Kv7.2 Potassium Channel Functions" Biomolecules 8, no. 3: 57. https://doi.org/10.3390/biom8030057
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.