
Location, Location, Location: Signals in Muscle Specification


1
Department of Pharmaceutical Sciences, College of Pharmacy, Oregon State University, Corvallis, OR 97331, USA
2
Molecular Cell Biology Graduate Program, Oregon State University, Corvallis, OR 97331, USA
*
Author to whom correspondence should be addressed.
J. Dev. Biol. 2018, 6(2), 11; https://doi.org/10.3390/jdb6020011
Submission received: 9 April 2018
/
Revised: 11 May 2018
/
Accepted: 15 May 2018
/
Published: 18 May 2018
(This article belongs to the Special Issue Crosstalk between FGF, TGF-β, BMP, and Wnt Signaling Pathways during Development)
Abstract
:Muscles control body movement and locomotion, posture and body position and soft tissue support. Mesoderm derived cells gives rise to 700 unique muscles in humans as a result of well-orchestrated signaling and transcriptional networks in specific time and space. Although the anatomical structure of skeletal muscles is similar, their functions and locations are specialized. This is the result of specific signaling as the embryo grows and cells migrate to form different structures and organs. As cells progress to their next state, they suppress current sequence specific transcription factors (SSTF) and construct new networks to establish new myogenic features. In this review, we provide an overview of signaling pathways and gene regulatory networks during formation of the craniofacial, cardiac, vascular, trunk, and limb skeletal muscles.
Keywords:
myogenesis; mesoderm; WNT; FGF; BMP; SHH; RA; NOTCH; ephrins; sequence specific transcription factor1. Introduction
In mammals, skeletal muscles begin to develop shortly after gastrulation and potentially can continue developing throughout their life. During gastrulation, epiblast cells ingress through the primitive streak to give rise to the three basic germ layers: ectoderm, mesoderm, and endoderm. During neurulation, as the primitive streak recedes posteriorly, ectodermal cells at the cranial end of the embryo proliferate, migrate, elevate, fold, and form the notochord and neural tube. Ectodermal cells shed and form the mesoderm, which further subdivides into cranial and trunk mesoderm. Early ingressing mesodermal cells migrate and populate underneath the ectoderm layer to form the lateral plate mesoderm (LPM), whereas the others subdivide into the paraxial (PM) (presomitic) and intermediate mesoderm. The skeletal muscle progenitor cells arise from the paraxial mesoderm, which flanks the axial mesoderm, notochord, and neural tube. Craniofacial muscles and cardiac muscle originate from the unsegmented PM, whereas, the skeletal muscles of body and limbs originate from the segmented PM that gives rise to somites (Figure 1a) [1].
The development of myogenic progenitors has been extensively investigated in vertebrate embryos since the 1990s using grafting [2], lineage tracing [3], in situ hybridization [4] and reporter (LacZ, EGFP) gene replacement [5,6,7] in both avian and mammalian embryos. These studies along with recent genomic wide association studies have established that the craniofacial, trunk, and skeletal musculature have unique anatomical origins and molecular networks [8,9,10,11]. Signals like retinoic acid (RA), sonic hedgehog homolog (SHH), bone morphogenetic proteins (BMP), WNT, fibroblast growth factor (FGF) are emitted from the neural tube, notochord, ectoderm, dorsal laminae, and neural crest [11]. RA and FGF set up the anterior-posterior cues respectively in early gastrulation. High concentration of RA triggers the formation of cranial PM and initiates the expression of cardiac (Pitx2, Tbx1, Pax7) and cranial (Nkx2.5, Tbx5, Gata4) SSTF networks [12,13]. FGFs from the tail end of embryo antagonize RA activity and initiate the formation of trunk PM, segmentation, and somite formation. BMP, SHH, WNT, and NOGGIN promote dorsal-ventral and medial-lateral cues that mediate trunk and abdominal SSTF networks (Six1/4, Pax3, Pax7, Pitx2) that in turn regulate the expression of basic helix-loop-helix (bHLH) myogenic regulatory factors (MRF; Myf5, Myf6, Myod1, Myog) [14]. MRFs regulate the commitment, determination, and differentiation of muscle progenitor cells [15]. MRFs act redundantly but control the cell-autonomous activation of myogenesis and regulate the expression of genes involved in contraction [16,17,18,19,20]. MYF5 and MYOD define the committed myoblasts, whereas MYF6 and MYOG define the differentiated myocytes. Differentiated myocytes exit the cell cycle and undergo morphological changes. In this review, we discuss the signaling pathways and gene networks that control skeletal muscle development in head and trunk, in addition to vascular and cardiac muscle (Table 1).
2. Craniofacial Myogenesis
Sixty muscle groups provide support for seeing, feeding, breathing, and moving of the head and neck. These muscles originate from the cranial mesoderm as a part of the paraxial mesoderm. Cranial mesoderm later specifies into the prechordal, pharyngeal, and lateral mesoderm. Prechordal mesoderm gives rise to the extraocular muscles (EOM) which encompass six muscles that control the eye movement [87]. Pharyngeal mesoderm forms the branchial arches (BA) that give rise to the masticatory, facial, pharyngeal and laryngeal muscles, and the secondary heart field [88,89]. Part of the pharyngeal mesoderm contributes to splanchnic mesoderm, that populates the tongue, axial neck, and heart (Figure 1a).
Craniofacial myogenesis is influenced by RA, BMP, FGF, and WNT and BMP inhibitors. RA activates Pitx2 and inhibits Tbx1 expression in the anterior head mesoderm. As RA is weakened in posterior head mesoderm, RA antagonist, FGF8, activates Tbx1 expression [90,91,92]. FGF and BMP reinforce TBX1 activity and activate Alx4 and Msc expression. FGF signaling spreads posteriorly and establishes the mature head mesoderm patterns, drives MyoR and Tbx1 expression in pharynx, BA muscles, and heart [93]. Grafting experiments in chick embryos have indicated that WNT1 and WNT3A from the dorsal neural tube, WNT13 from the surface ectoderm, and BMP4 and BMP7 from the dorsal neural tube and ectoderm inhibit head muscle myogenesis [94,95]. However, the WNT antagonists, FRZB, and the BMP inhibitors, NOGGIN and GREMLIN, from the cranial neural crest and other surrounding tissues, induce craniofacial myogenesis [95]. Lineage tracing approaches in both avian and mouse models demonstrated that EOM and BA cells contribute to distinct muscles [96]. Homeodomain transcription factors Tbx1, Pitx2, Tcf21 and Lhx2 are expressed in the pharyngeal mesoderm and regulate cranial myogenesis. PITX2 specifies EOM by activating Myf5, Myf6, and Myod1 [14]. During BA myogenesis, PITX2 activates Tbx1, Msc, Tcf21, and Six2 [97,98]. TBX1 induces the expression of Fgf receptors and their ligands. Perturbation of Fgf in mice or fish leads to hypoplastic and asymmetric muscles [99,100]. TBX1 and FGF activate en2 in myogenic cells in the dorsal mandibular pharyngeal arch and contribute to jaw development. Although function of EN2 is still unclear, studies in chick embryos implied that signaling pathways prevent prematurely specification. TBX1, MSC, and TCF21 together activate MYF5 and lead to MYOD1 and MYOG expression (Figure 2) [101]. PAX7 is expressed in MRF+ cells [88,94,102] and controls the formation of head satellite cells [103]. Head satellite cells derive from the MESP1+ cells [96] but only the EOM and pharyngeal satellite cells express ISL1, ALX4 PITX1, PITX2, TCF21, cytokines, and chemokines [14,96,104].
Tongue [105] and neck muscles arise from the medial-dorsal and lateral-ventral domains of the occipital and cervical somites [106]. WNT signaling regulates tongue myogenesis [107] and differentially activation of MYF5 and MYOD1 [108]. WNT from the neural tube induces the expression of Colloid-like1, and reinforces BMP activity in the most anterior medial dermomyotome (DM) and regulates neck myogenesis [31].
3. Cardiac Myogenesis
Cranial mesoderm cells ingress laterally and contribute to the formation of the myocardium and endocardium [109], while medial and lateral splanchnic mesoderm give rise to the chambers of the heart [97]. RA establishes the posterior boundary of the secondary heart field [110], influences the development of the posterior and sinoatrial region [111] and transforms the cranial cells into cardiomyocytes [112]. BMP is expressed at the posterior and lateral region of the embryo and recruits head mesoderm cells to the heart [109,112]. WNT1, WNT3a, and WNT8c from the posterior primitive streak and lateral mesoderm inhibit cardiogenesis [113] and define the migratory limits of the cardiomyocytes. BMP antagonist NOGGIN from the notochord provides timely control to initiate early heart formation [12,13]. Collectively, the above signals promote expression of NKX2.5, TBX5, MEF2C and GATA4 [12,13] and the chromatin remodeling subunit SMARCD3 [114] and force the transition of mesodermal cells into cardiocytes [115].
Just after gastrulation, the cardiac crescent or 1st heart field, forms a tubular structure, with an outer myocardium and an inner endocardium. The linear heart tube connects with the artery at the anterior pole which forms the outflow tract and with a vein at the venous pole, which forms the inflow tract [116]. BMP2 activates NKX2.5 expression in the cardiac crescent. NKX2.5 interacts with GATA4 and TBX5 [117] to promote cell differentiation [118] and suppress the expression of FGF10 to prevent prenatal development [119]. BA-derived cells, 2nd heart field, express FGF8, FGF10, ISL1 [120] and TBX1 [121] proliferate, contribute to the extension of the tubular heart, and the formation of outflow tract and right ventricle. PITX2 activates MSC [97,98] and TBX1 [122], which triggers FGF10 expression to stimulate cardiac muscle progenitors towards migration and proliferation [123]. Cranial neural crest (NC) cells contribute to the formation of valve cushions of the outflow tract [124,125]. Histone deacetylases (HDACS) [126] and TGFβ modulate their migration [127] throughout the dorso-ventral axis [128,129]. TGFβ represses cardiomyocyte specification to prevent premature differentiation [130].
4. Smooth Muscle Formation
Smooth muscle cells (SMC) give rise to gastrointestinal, urogenital and respiratory tract, and blood vessels [131]. The specific mechanisms responsible for SMC determination and differentiation are largely unknown. A heterogeneous population of cells derive from somites [132], secondary heart field [133], splanchnic mesoderm [134], and NC cells [135] contribute to the vascular SMC formation. BMP, NOTCH, SHH, and TGFβ1 promote vascular SMC specification and differentiation by activating serum response factors (SRF) and their cofactors. Activation of SRF leads to expression of ACTA), SM myosin heavy chain, and SM22 (review [136,137]). BMP and NOTCH initiate myogenic patterning in vascular SMC development. BMP2 stimulates their migration and the expression of the Va myosin [138], whereas NOTCH is required for differentiation of the vascular SMC progenitors [139]. SHH regulates vascular SMC proliferation and the formation of coronary vessels [140]. TGFβ1 inhibits vascular SMC growth, increases proliferation of the NC-derived vascular SMC via plasminogen activator inhibitor (PAI) [141]. TGFβ1 also stimulates vascular SMC differentiation by activating ACTA2 [142], SM22 [143], SMAD3 [144], and RhoA [145]. TGFβ/BMP co-receptor ENDOGLIN, a descendent of the PAX3+ vascular SMCs is required for angiogenesis [146]. More recent studies also indicated the PAX3+ DM might act as a stem cell niche for vascular SMCs [147].
5. Trunk Myogenesis
Skeletal muscles of the trunk originate from somites, which are derived from the segmented PM and located on the either side of the neural tube. Somitogenesis occurs progressively in an anterior to posterior sequence, simultaneous with regression of the primitive streak soon after neurulation (Figure 1a). After somites are formed and segmented, cells start to differentiate along the dorsal-ventral axis [1]. Ventral cells undergo an epithelial to mesenchymal transition to form the sclerotome, which will generate cartilage and associated connective tissues of the vertebrate and ribs. Dorsal cells remain epithelial and form the DM, which will generate the dermis, skeletal muscle of the truck and limbs, and brown fat [148]. The first myogenic tissue, myotome, arises later from the dorsomedial (DML) and ventrolateral (VLL) lips of the DM [149]. The DM is subdivided into the lateral hypaxial and medial epaxial DM. Cells of the VLL form the hypaxial myotome that gives rise to the lateral trunk and limb musculature, and cells from the epaxial myotome give rise to the deep muscles of the back (Figure 1c) [150,151].
5.1. Somitogenesis
Cells from the anterior tip of presomitic mesoderm undergo different morphogenetic changes and form as an epithelial ball, termed somitomeres [152,153]. Somitic cells are temporally controlled by the cyclin expression of intrinsic oscillating genes (“clock”) that set the pace [154] and signals secreted from nearby tissues (“wave front”) that define the position of the posterior border of each new somite [155,156]. Clock genes are expressed cyclically to define the time interval. Combinatorial signals (RA, FGF8, WNT3, and SHH) regulate the region where the clock genes are segregated and gives rise to a new pair of somites. Gradient signals provide the spatial clue and define the boundaries of segmentation of newly formed somites. FGF8 and WNT3 are produced in the caudal region of the embryo, RA is secreted by the cranial region of newly formed somites [152,153,157], and SHH is released from the notochord [155]. SHH signaling is required to preserve the FGF8 gradient. FGF8 signaling counteracts an opposing RA gradient [158]. The interaction of WNT and FGF8 regulates not only the separation, but also the structure of somites. At the anterior end of PM, WNT induces the expression of ß-catenin and N-cadherin in the center of somite. N-cadherin regulates the adhesion of the epithelial surface of the somite by forming tight junctions and a basal lamina that separates it from nearby tissues [64]. At the same time, WNT induces its antagonist, NOTCH, at the posterior end of somite, while NOTCH induces Epha4 and either ephrinB2 (chick) or Ripply (mouse). Together, they define the posterior and anterior end of the somite and promote their separation (Figure 1b) [64,159,160].
Mature somites differentiate into different compartments and determine the fates of cells (Figure 1c). Compartmentalization is induced by SHH and NOGGIN from the floor plate of neural tube and notochord, and by WNTs and BMPs from the ectoderm and dorsal laminae of the neural tube. Noggin binds and inactivates Bmp4 and creates morphogenic gradients [161,162]. Cells at the ventromedial part of the somite, located closer to the notochord and the ventral part of neural tube, receive a higher concentration of NOGGIN and SHH. As they proliferate, they lose the expression of N-cadherin and later become mesenchymal cells and develop into sclerotome. They express PAX1 and SOX9 that constitute the cartilage, vertebrae, and ribs and I-MF that inhibits muscle differentiation. Cells located at the dorsomedial part of the somite receive a low concentration of NOGGIN and SHH, remain epithelial, and give rise to DM [163,164,165]. The epithelial DM is divided into dorsomedial (epaxial), central (dermatome), and ventromedial (hypaxial) DM. The epithelial DM is influenced by different signals to activate distinct regulatory gene networks that contribute to specific muscle formation in the body. PAX3 marks the myogenic lineage in the DM. The PAX3+ cells mark the DM-derived cells that give rise to skeletal [166,167] and vascular muscles [168,169]. Ablation of Pax3 leads to the absence of forelimb muscles [170,171]. SIX1, SIX4, and their cofactors EYA1 and EYA2 directly bind on the promoter of Pax3 and activate its expression [15,172,173]. Double mutants of Six1/4 and Eya1/2 exhibit loss of PAX3 expression that leads to migratory defects in hypaxial muscles [172,173].
PAX3 plays a key role in activating the MRF expression. MYF5 is expressed earlier in the dorsomedial lip of DM and then in the hypaxial myotome under the influenced of WNT1 and SHH to give rise to the epaxial muscles [174,175]. MYOD1 expression is induced by WNT7a and inhibited by BMP4 in the ventrolateral DM. Expression of MYF6 and MYOG coincides with the innervation and myofiber formation [18,19].
5.2. Epaxial Muscle Formation
Cells in the epaxial DM receive WNT1 and WNT3a from the dorsal neural tube and low levels of SHH from the floor plate of the neural tube. WNT1 and WNT3 activate Myf5 [176] and Myod1 expression through the WNT canonical pathway [177,178,179,180,181]. Low level of SHH contributes to Myf5 expression through the GLI2 and ZIC1 [182,183]. Epaxial DM receives NOTCH/DELTA1 signaling from migrating NC cells and induces MYF5 expression [184]. PAX3 also regulates Myf5 expression through direct binding to Dmrt2 motifs [15,181,185]. PAX3 and MYF5 trigger MYOd1 expression, which drives myoblast differentiation [175]. MYF6 is expressed in the somites simultaneously with MYF5, although what activates its expression is still unknown [20].
5.3. Dermatome Formation
The dermatome is heterogenic and distinct, marks the boundary between the epaxial and hypaxial DM, and gives rise to the dermis of head and neck, satellite cells, and brown adipocytes [9,151,186,187,188]. The combined influence of SHH from the notochord, WNT1 from the dorsal neural tube, WNT11 from surface ectoderm and dermatome, and the negative influenced of BMP4 from the LPM marks the epaxial-hypaxial border. Homeobox gene engrailed1 (en1) blocks the epaxial-hypaxial interface. EN1 is expressed on the border of the hypaxial maker SIM1, which marks the territories of motor neurons and regulates muscle patterning [189]. The EN1-SIM1 expression boundary marks the epaxial-hypaxial DM boundary and the dermatome. The specification of cells in the dermatome is diverse. A subset of cells in the medial part of dermatome receives BMP from the roof plate of the neural tube and migrates dorsally into the sub-ectodermal space to form cartilage and the medial margin of scapula in birds and mammals [190,191,192,193]. Another subset of cells, which is under the influence of Wnt6 from the dorsal ectoderm, expresses PAX3 and PAX7 and forms the satellite cells [9,151,186,187,188]. PAX3 induces the expression of PAX7 in the central DM. The PAX3+/PAX7+ muscle progenitor cells populate the myotome [167,187,188] and receive neurogulin1 (nrg1) from the migrating NC cells through NRG1 receptor ERBB3. The NRG1-ERBB3 signaling maintains PAX7 expression and prevents premature myogenic differentiation [194]. In addition, DERMO-1 also modulates the development of mesenchymal cell lineages including muscle and dermis [195]. DERMO-1 represses transcriptional activity of MEF2 and MYOD in a dose-dependent fashion [196].
5.4. Hypaxial Muscle Formation
The hypaxial DM gives rise to vertebral muscle, the diaphragm, abdominal muscles and muscles of the limb. Development of hypaxial DM is highly influenced by the LPM [165,197]. Upon receiving WNT7a from the dorsal ectoderm and Bmp4 from the LPM, cells give rise to hypaxial myotome [165,197]. BMP signaling induces somite-derived endothelial cell differentiation and migration via VEGFR2 expression [198,199], whereas the WNT7a/ PKC activates Pax3 in hypaxial DM [200]. PAX3 is critical for the onset of embryonic myogenesis by regulating delamination via C-MET, migration via LBX, and determination of muscle progenitors [166,167] in the limb [201,202,203,204,205]. PITX2 lies genetically downstream of PAX3 in the hypaxial DM [206,207]. Activation of Pitx2 regulates expression of MYOD1 and MYOG, myoblast motility, and skeletal muscle maintenance [208,209]. PAX3 regulates the expression of ITM2a in DM, limb buds, adult muscle fibers, and satellite cells [210]. In turn, ITM2a regulates muscle creatine kinase (CK-M) expression and muscle differentiation [211].
5.5. Myotome Formation
Myotome is the first myogenic structure located between the DM dorsally, and the sclerotome ventrally. Cells of the DM, lose expression of N-cadherin, delaminate, and translocate underneath the DM to form the new structure, the myotome. Surrounding signals trigger the expression of MRFs and establish their myogenic identity as they populate the myotome. Upon arrival, the cells start to differentiate and fuse to become myofibers.
During the first wave of myogenesis, myogenic progenitors from the DM develop into mononucleated myocytes. Cells from the dorsal part of myotome originate from the dorsomedial lip (DML), migrate ventrally as they receive WNT1 and WNT3a from the dorsal neural tube, WNT7a from dorsal ectoderm, and SHH from the ventral region of the neural tube and notochord. The WNT and SHH signaling defines the myogenic properties of migrating muscle precursors by activating the expression of MYF5 [72,180,182] and MYOD1 [148]. Although the cells from the central DM contribute to the myotome, they are less significant than cells from the DML and ventrolateral lip (VLL). Cells in the central DM receive neurophrin-3 and WNT1 from the neural tube and become part of the myotome [212]. Cells from the VLL are essential for lateral extension of the myotome [213]. They receive WNT7a from the dorsal ectoderm and BMP4 and FGF5 from the LPM, inducing the expression of MYOD1, but not MYF5 [214].
After entering the myotome, MYF5 and SHH together activate the expression of Fgf4, which regulates the proliferation and differentiation of the MRF expressing myocytes [215]. Once the myocyte identity is established, they express PAX7 before undergoing terminal differentiation. SHH is crucial to maintain the expression of PAX7 in migrating MYF5+/MYOD1+ cells. Interfered by SHH results in accumulation of PAX7+ cells and a small myotome in chick and mouse embryos [216,217]. Myocytes receive WNT11 as they elongate along the somites and express both slow- and fast-type cytoskeletal proteins, including MYH (slow type I), MYH3 (embryonic), ACTC1, ACTA1, DESMIN, JAG2, ß-ENOLASE, and CA2 [8,217,218]. Myocytes fuse and form the first multinucleated MyHC+ myofibers spreading from the ventral to the dorsal myotome [217]. Six1 and Six4 expression activates the fast muscle program [218].
5.6. Limb Muscle Formation
Limb muscle development is a triphasic process. During embryonic myogenesis in mice (E9-E12), the first multinucleated muscle fibers are formed from PAX3+ embryonic myoblasts, followed by PAX3+LBX1+C-MET+ migratory myoblasts that provide the basic pattern and primary myofibers throughout the limbs. During fetal myogenesis (E13-E16), the PAX3+/PAX7+ fetal muscle progenitors use embryonic fibers as a scaffold to expand the muscle mass. During perinatal myogenesis (E17-P5), PAX7+ muscle progenitors (satellite cells) are located between the basal lamina and the fiber plasma membrane and remain mononucleated and quiescent. After muscle damage, satellite cells can activate, fuse with existing myofibers, and repair damaged muscle (Figure 2) [10,219].
Embryonic (primary) myogenesis begins when the first embryonic myoblast progenitors of the hypaxial DM start to delaminate and migrate to forelimb buds [220]. PAX3 activates expression of c-MET and LBX1, and the PAX3+LBX1+c-MET+ cells delaminate, become mobile, and start to migrate distally [203,204,205]. LBX1 is crucial for the migrating embryonic myoblast progenitors (EMP) [205,221]. The migratory directions are controlled by signals from the surface of limb buds, zone of polarizing activity (ZPA). Misrouted EMPs either migrate slowly or accumulate in the mesoderm of ventral body wall and form smaller or no muscles [222,223]. SHH signals from ZPA regulate the migratory patterns of EMPs, maintenance of limb bug outgrowth, dorso-ventral limb patterning, and development of skeleton, cartilage, and tendons of digits [224,225]. SHH and FGFs from the ectodermal ridge control the expression of HGF/SF, the only known ligand of c-MET [226], which directs the migration of EMPs [203,204]. Migratory EMPs also express the chemokine receptor CXCR4. This receptor responds to chemo-attractant SDF1 from the limb buds, which also directs the migratory routes [227]. SF/HGF, BMP, WNT and SFRP-2 inhibit prematurely differentiating EMPs and allow them to multiply and populate the limb bud parallel to establishing the myogenic program [198,228]. The Wnt/ß-catenin/PITX2 pathway controls motility and proliferation of the EMPs by regulating expression of the growth control genes Ccnd1, Ccnd2 and c-myc, [209]. IGF contributes to the proliferation and determination of EMPs [229]. IGF-1 and IGF-2 play a role in PAX3 and MYOG expression via PI3K and MYOD through FGF18 [230]. Upon their entry into the limb bud, EMPs express high levels of MRFs and become embryonic myoblasts and myofibers. At the same time, SHH initiates the expression of MYF5 though GLIA [231]. The spindle-shaped EMPs receive FGF and TGFβ, and increase in cell number while they align to each other. FGFs maintain the expression of MSX1 to prevent their premature differentiation. The newly formed MYOD1+embryonic myoblasts exit the cell cycle and express MYOG and MEF2 [232]. During differentiation, embryonic myoblasts undergo several changes, including cell aggregation, elongation, metabolic changes, cell membrane fragmentation and cytoskeleton assembly. They start to express MyHC and myosin light chain 1 (MYL1), secrete fibronectin, RGD-binding integrins, cadherins, and cell adhesion molecules that promote the alignment of migratory cells [233,234,235], and fuse to each other to become multinucleated primary myofibers [236].
Fetal myogenesis begins when a subset of PAX3+ myogenic progenitors start to express PAX7 while their PAX3 expression is decreased in the central DM [167,188,237,238]. Mouse Pax7 mutants exhibit defects in fetal myogenesis with smaller muscles, fewer myofibers, and impaired satellite cells, leading to defective muscle regeneration [239,240,241,242,243]. PAX7+ cells constitute the fetal myogenic progenitors (FMP). FMP receive TGFβ, BMP, and WNT/β-catenin signaling from the ectoderm and the dorsal laminae to block the premature differentiation. TGFβ2 from the ventral region of the limb buds and later from the muscle mass [173] represses MYOD1 and MYOG activity [244], induces FMP migration and inhibits their differentiation [245,246]. They increase their number and contribute to all muscles [237] as they migrate and fuse to the primary fibers and each other to form the secondary fibers. PAX7 directly binds and activates Nfix, which marks the fetal myoblasts. NFIX represses the embryonic muscle genes Sox6 and MyHCI, and activates the fetal muscle genes α7-integrin, β-enolase, muscle creatine kinase, and muscle sarcomeric proteins [247]. PITX2 and PITX3 are also expressed in fetal myoblasts [248] as they commit to the fetal program and become myocytes by subsequent expression of MYOD1, MYF5, and MYOG [167]. When differentiated, fetal myocytes fuse to embryonic myofibers and give rise to doughnut-shaped, multinucleated fetal myofibers [188,249].
During birth and the first few weeks after birth, the third wave of myogenesis begins with an increase in number and size of myofibers. PAX7+satellite cells express myogenic (c-MET, HGF/SF, MSX1) and endothelial (CD34) markers [250,251]. Expression of c-met and Hgf/Sf promotes delamination and migration from the basal lamina once activated in response to stress. Activated satellite cells turn off Pax7 and begin to express Myod1 and cell-cycle markers and undergo multiple rounds of asymmetric cell division. A small number of activated satellite cells returns to an undifferentiated state (PAX7+MYOD1-) for self-renewal. PITX2 regulates satellite cell division by stimulating the expression of Ccnd1, Ccnd2 and Myf5, and downregulates miR-15b, miR-106b, miR-23b, and miR-503 [252]. Daughter satellite cells express MYF5 and MYOD, and fuse to pre-existing myofibers to form new fibers [238]. MSTN, a member of TGF family, regulates satellite cell renewal and muscle growth [253]. MSTN inhibits Pax7 expression and binds to activin. Activin activates ALK4/5, mediates SMA2/3 phosphorylation [254,255], and recruits SMAD4. MSTN/activin/SMAD signaling pathway inhibits MRF expression [256], and limits proliferation and differentiation into myofibers via AKT/TORc1/P70S6K pathway [257,258,259]. MSTN antagonist, Follistatin (Fst), promotes PAX7 activity [260,261,262], regulates MRFs expression and muscle fiber formation [263]. Bone morphogenetic protein and activin membrane-bound inhibitor (BAMBI), another member of TGF family, responds to Wnt/β-catenin pathway and plays a role in regulating SMAD activity [264].
6. Abdominal Myogenesis
Abdominal muscles encompass three layers of skeletal muscles, which merge toward the midline and form a sheath to assist breathing and protect the inner organs. Abdominal muscles derive from LPM and somites. LPM gives rise to somatopleure (lateral) and splanchnopleure (medial). The somatopleure and the surface epithelial endoderm give rise to the body wall in three waves similar to trunk myogenesis (Figure 1a) [265,266]. Myogenic cells within a thin sheet of LPM-derived cells proliferate without delamination, remain within the DM epithelium, and migrate ventrally into the somatopleure to cover the abdomen and form the primary body wall [266,267]. The primary body wall is composed of myogenic and epithelial cells that provide the first tissue to cover the endoderm. After several days, a second and third wave of myogenic progenitor cells move into the somatopleure and all layers are joined at the midline [268,269]. The secondary body wall forms muscle, skin, ribs, and sternum. Cells of the secondary body wall are heterogeneous, composed of myogenic, mesenchymal and NC cells. WNT/PITX2 signaling plays a role in the abdominal myogenesis. WNT/β-catenin signaling instructs mesodermal specification of somites and LPM-derived mesenchymal cells in ventral body wall as a paracrine signal [270]. WNT7a from the dorsal ectoderm and BMP from the LPM activate expression of PAX3 and its targets. WNT signaling activates Pitx2 expression in LPM. PAX3+PITX2+ cells proliferate, migrate to the midline, and express MRFs [271]. Ablation of Pitx2 results in repression of T-box and activation of Hox9-11 genes in the abdominal wall [266,272]. AP-2a regulates the closure of all four body folds by regulating the epithelial-mesenchymal interactions and cell-cell communication from the ectoderm to the abdominal mesoderm [268,273]. Defects in these pathways lead to hypoplasia in the abdominal wall or organ exposure. HOXb2 and HOXb4 activate ALX3/4 [274,275] and regulate the primary ventral body formation [276,277,278]. Independent from the PITX2 pathway [279], BMP1 might enhance the activity or availability of TFGß5+ myofibroblasts during the first wave [280] and initiate the migration in the second wave [281]. TGFβ2/3 plays a fundamental role in activating cytoskeletal components (Tagln, α-Sma and desmin) that support myofibroblast migration towards the midline [282]. The signaling pathways contribute to ventral wall closing by involving the assembly and function of extracellular matrix and collagen fibrils [283]. NC cells migrate to the midline and contribute to the second body wall formation. AP-2 is expressed in migrating NC cells [284].
7. Conclusions
Skeletal muscle progenitor cells arise from the PM and LPM that are heavily influenced by signals from the neural tube, notochord, ectoderm, and dorsal laminae. Combinatorial networks of signaling molecules regulate the expression of SSTFs that further regulate the commitment and differentiation to the myogenic lineage. The PM of the cranial domain forms early and gives rise to craniofacial, cardiac and vascular smooth muscles. PITX2+MSC+TBX1+ cells from the paraxial head develop the cardiogenic and head muscle cells. Head myogenesis is inhibited by WNT signaling. The segmented trunk PM forms the somites, in which the PAX3+ cells are influenced by WNT and SHH to commit to myogenic MYOD1+ cells and PAX7+ satellite stem cells.
Acknowledgments
We thank colleagues for inspiring discussions, and Jason Lattier for editing the manuscript. We apologize for not including many pivotal studies due to space limitations. This work was supported by the College of Pharmacy, Oregon State University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Burke, A.C.; Nowicki, J.L. A new view of patterning domains in the vertebrate mesoderm. Dev. Cell 2003, 4, 159–165. [Google Scholar] [CrossRef]
- Borycki, A.G.; Emerson, C.P., Jr. Multiple tissue interactions and signal transduction pathways control somite myogenesis. Curr. Top. Dev. Biol. 2000, 48, 165–224. [Google Scholar] [PubMed]
- Ordahl, C.P.; Le Douarin, N.M. Two myogenic lineages within the developing somite. Development 1992, 114, 339–353. [Google Scholar] [PubMed]
- Pownall, M.E.; Emerson, C.P., Jr. Sequential activation of three myogenic regulatory genes during somite morphogenesis in quail embryos. Dev. Biol. 1992, 151, 67–79. [Google Scholar] [CrossRef]
- Goldhamer, D.J.; Faerman, A.; Shani, M.; Emerson, C.P. Regulatory elements that control the lineage-specific expression of myoD. Science 1992, 256, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Pinney, D.F.; de la Brousse, F.C.; Faerman, A.; Shani, M.; Maruyama, K.; Emerson, C.P., Jr. Quail myoD is regulated by a complex array of cis-acting control sequences. Dev. Biol. 1995, 170, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Tajbakhsh, S.; Rocancourt, D.; Buckingham, M. Muscle progenitor cells failing to respond to positional cues adopt non-myogenic fates in myf-5 null mice. Nature 1996, 384, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Chal, J.; Pourquie, O. Making muscle: Skeletal myogenesis in vivo and in vitro. Development 2017, 144, 2104–2122. [Google Scholar] [CrossRef] [PubMed]
- Christ, B.; Huang, R.; Scaal, M. Amniote somite derivatives. Dev. Dyn. 2007, 236, 2382–2396. [Google Scholar] [CrossRef] [PubMed]
- Biressi, S.; Molinaro, M.; Cossu, G. Cellular heterogeneity during vertebrate skeletal muscle development. Dev. Biol. 2007, 308, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Placzek, M.; Tessier-Lavigne, M.; Yamada, T.; Jessell, T.; Dodd, J. Mesodermal control of neural cell identity: Floor plate induction by the notochord. Science 1990, 250, 985–988. [Google Scholar] [CrossRef] [PubMed]
- Schultheiss, T.M.; Lassar, A.B. Induction of chick cardiac myogenesis by bone morphogenetic proteins. Cold Spring Harb. Symp. Quant. Biol. 1997, 62, 413–419. [Google Scholar] [PubMed]
- Schlange, T.; Andree, B.; Arnold, H.H.; Brand, T. BMP2 is required for early heart development during a distinct time period. Mech. Dev. 2000, 91, 259–270. [Google Scholar] [CrossRef]
- Sambasivan, R.; Gayraud-Morel, B.; Dumas, G.; Cimper, C.; Paisant, S.; Kelly, R.G.; Tajbakhsh, S. Distinct regulatory cascades govern extraocular and pharyngeal arch muscle progenitor cell fates. Dev. Cell 2009, 16, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Tajbakhsh, S.; Borello, U.; Vivarelli, E.; Kelly, R.; Papkoff, J.; Duprez, D.; Buckingham, M.; Cossu, G. Differential activation of Myf5 and MyoD by different Wnts in explants of mouse paraxial mesoderm and the later activation of myogenesis in the absence of Myf5. Development 1998, 125, 4155–4162. [Google Scholar] [PubMed]
- Weintraub, H.; Davis, R.; Tapscott, S.; Thayer, M.; Krause, M.; Benezra, R.; Blackwell, T.K.; Turner, D.; Rupp, R.; Hollenberg, S.; et al. The myoD gene family: Nodal point during specification of the muscle cell lineage. Science 1991, 251, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Delfini, M.C.; Duprez, D. Ectopic Myf5 or MyoD prevents the neuronal differentiation program in addition to inducing skeletal muscle differentiation, in the chick neural tube. Development 2004, 131, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Olson, E.N.; Arnold, H.H.; Rigby, P.W.; Wold, B.J. Know your neighbors: Three phenotypes in null mutants of the myogenic bHLH gene MRF4. Cell 1996, 85, 1–4. [Google Scholar] [CrossRef]
- Hasty, P.; Bradley, A.; Morris, J.H.; Edmondson, D.G.; Venuti, J.M.; Olson, E.N.; Klein, W.H. Muscle deficiency and neonatal death in mice with a targeted mutation in the myogenin gene. Nature 1993, 364, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Kassar-Duchossoy, L.; Gayraud-Morel, B.; Gomes, D.; Rocancourt, D.; Buckingham, M.; Shinin, V.; Tajbakhsh, S. Mrf4 determines skeletal muscle identity in Myf5:Myod double-mutant mice. Nature 2004, 431, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.A.; Israel, D.I.; Kelly, S.; Luxenberg, D.P. Bone morphogenetic protein-2 causes commitment and differentiation in C3H10T1/2 and 3T3 cells. Growth Factors 1993, 9, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Denker, A.E.; Haas, A.R.; Nicoll, S.B.; Tuan, R.S. Chondrogenic differentiation of murine C3H10T1/2 multipotential mesenchymal cells: I. Stimulation by bone morphogenetic protein-2 in high-density micromass cultures. Differentiation 1999, 64, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, T.; Yamaguchi, A.; Komaki, M.; Abe, E.; Takahashi, N.; Ikeda, T.; Rosen, V.; Wozney, J.M.; Fujisawa-Sehara, A.; Suda, T. Bone morphogenetic protein-2 converts the differentiation pathway of C2C12 myoblasts into the osteoblast lineage. J. Cell Biol. 1994, 127, 1755–1766. [Google Scholar] [CrossRef] [PubMed]
- Okubo, Y.; Bessho, K.; Fujimura, K.; Iizuka, T.; Miyatake, S. Expression of bone morphogenetic protein-2 via adenoviral vector in C2C12 myoblasts induces differentiation into the osteoblast lineage. Biochem. Biophys. Res. Commun. 1999, 262, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Patterson, S.E.; Bird, N.C.; Devoto, S.H. BMP regulation of myogenesis in zebrafish. Dev. Dyn. 2010, 239, 806–817. [Google Scholar] [CrossRef] [PubMed]
- Frank, N.Y.; Kho, A.T.; Schatton, T.; Murphy, G.F.; Molloy, M.J.; Zhan, Q.; Ramoni, M.F.; Frank, M.H.; Kohane, I.S.; Gussoni, E. Regulation of myogenic progenitor proliferation in human fetal skeletal muscle by BMP4 and its antagonist Gremlin. J. Cell Biol. 2006, 175, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Noulet, F.; Edom-Vovard, F.; Tozer, S.; Le Grand, F.; Duprez, D. Bmp signaling at the tips of skeletal muscles regulates the number of fetal muscle progenitors and satellite cells during development. Dev. Cell 2010, 18, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Re’em-Kalma, Y.; Lamb, T.; Frank, D. Competition between noggin and bone morphogenetic protein 4 activities may regulate dorsalization during Xenopus development. Proc. Natl. Acad. Sci. USA 1995, 92, 12141–12145. [Google Scholar] [CrossRef] [PubMed]
- Reshef, R.; Maroto, M.; Lassar, A.B. Regulation of dorsal somitic cell fates: BMPs and Noggin control the timing and pattern of myogenic regulator expression. Genes Dev. 1998, 12, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Epperlein, H.H.; Vichev, K.; Heidrich, F.M.; Kurth, T. BMP-4 and Noggin signaling modulate dorsal fin and somite development in the axolotl trunk. Dev. Dyn. 2007, 236, 2464–2474. [Google Scholar] [CrossRef] [PubMed]
- Pais de Azevedo, T.; Zuzarte-Luis, V.; Goncalves, L.; Marques, C.; Palmeirim, I. Tissue regulation of somitic colloid-like1 gene expression. Biochem. Biophys. Res. Commun. 2012, 424, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Schuster-Gossler, K.; Cordes, R.; Gossler, A. Premature myogenic differentiation and depletion of progenitor cells cause severe muscle hypotrophy in Delta1 mutants. Proc. Natl. Acad. Sci. USA 2007, 104, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Hrabe de Angelis, M.; McIntyre, J., II; Gossler, A. Maintenance of somite borders in mice requires the Delta homologue DII1. Nature 1997, 386, 717–721. [Google Scholar] [CrossRef] [PubMed]
- Amthor, H.; Christ, B.; Rashid-Doubell, F.; Kemp, C.F.; Lang, E.; Patel, K. Follistatin regulates bone morphogenetic protein-7 (BMP-7) activity to stimulate embryonic muscle growth. Dev. Biol. 2002, 243, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Costamagna, D.; Mommaerts, H.; Sampaolesi, M.; Tylzanowski, P. Noggin inactivation affects the number and differentiation potential of muscle progenitor cells in vivo. Sci. Rep. 2016, 6, 31949. [Google Scholar] [CrossRef] [PubMed]
- Tylzanowski, P.; Mebis, L.; Luyten, F.P. The Noggin null mouse phenotype is strain dependent and haploinsufficiency leads to skeletal defects. Dev. Dyn. 2006, 235, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Niswander, L.; Martin, G.R. Fgf-4 expression during gastrulation, myogenesis, limb and tooth development in the mouse. Development 1992, 114, 755–768. [Google Scholar] [PubMed]
- Edom-Vovard, F.; Bonnin, M.A.; Duprez, D. Misexpression of Fgf-4 in the chick limb inhibits myogenesis by down-regulating Frek expression. Dev. Biol. 2001, 233, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Edom-Vovard, F.; Schuler, B.; Bonnin, M.A.; Teillet, M.A.; Duprez, D. Fgf4 positively regulates scleraxis and tenascin expression in chick limb tendons. Dev. Biol. 2002, 247, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Dormann, D.; Munsterberg, A.E.; Weijer, C.J. Cell movement patterns during gastrulation in the chick are controlled by positive and negative chemotaxis mediated by FGF4 and FGF8. Dev. Cell 2002, 3, 425–437. [Google Scholar] [CrossRef]
- Burks, P.J.; Isaacs, H.V.; Pownall, M.E. FGF signalling modulates transcriptional repression by Xenopus groucho-related-4. Biol. Cell 2009, 101, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.E.; Isaacs, H.V.; Pownall, M.E. eFGF is required for activation of XmyoD expression in the myogenic cell lineage of Xenopus laevis. Development 2002, 129, 1307–1315. [Google Scholar] [PubMed]
- Hebert, J.M.; Boyle, M.; Martin, G.R. mRNA localization studies suggest that murine FGF-5 plays a role in gastrulation. Development 1991, 112, 407–415. [Google Scholar] [PubMed]
- Clase, K.L.; Mitchell, P.J.; Ward, P.J.; Dorman, C.M.; Johnson, S.E.; Hannon, K. FGF5 stimulates expansion of connective tissue fibroblasts and inhibits skeletal muscle development in the limb. Dev. Dyn. 2000, 219, 368–380. [Google Scholar] [CrossRef]
- Groves, J.A.; Hammond, C.L.; Hughes, S.M. Fgf8 drives myogenic progression of a novel lateral fast muscle fibre population in zebrafish. Development 2005, 132, 4211–4222. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.F.; Tsao, K.C.; Lee, S.J.; Yang, R.B. SCUBE3 (signal peptide-CUB-EGF domain-containing protein 3) modulates fibroblast growth factor signaling during fast muscle development. J. Biol. Chem. 2014, 289, 18928–18942. [Google Scholar] [CrossRef] [PubMed]
- Hammond, C.L.; Hinits, Y.; Osborn, D.P.; Minchin, J.E.; Tettamanti, G.; Hughes, S.M. Signals and myogenic regulatory factors restrict pax3 and pax7 expression to dermomyotome-like tissue in zebrafish. Dev. Biol. 2007, 302, 504–521. [Google Scholar] [CrossRef] [PubMed]
- Omoteyama, K.; Takagi, M. FGF8 regulates myogenesis and induces Runx2 expression and osteoblast differentiation in cultured cells. J. Cell Biochem. 2009, 106, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Gryzik, T.; Muller, H.A. FGF8-like1 and FGF8-like2 encode putative ligands of the FGF receptor Htl and are required for mesoderm migration in the Drosophila gastrula. Curr. Biol. 2004, 14, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Rinon, A.; Lazar, S.; Marshall, H.; Buchmann-Moller, S.; Neufeld, A.; Elhanany-Tamir, H.; Taketo, M.M.; Sommer, L.; Krumlauf, R.; Tzahor, E. Cranial neural crest cells regulate head muscle patterning and differentiation during vertebrate embryogenesis. Development 2007, 134, 3065–3075. [Google Scholar] [CrossRef] [PubMed]
- Kopan, R.; Nye, J.S.; Weintraub, H. The intracellular domain of mouse Notch: A constitutively activated repressor of myogenesis directed at the basic helix-loop-helix region of MyoD. Development 1994, 120, 2385–2396. [Google Scholar] [PubMed]
- Vasyutina, E.; Lenhard, D.C.; Wende, H.; Erdmann, B.; Epstein, J.A.; Birchmeier, C. RBP-J (Rbpsuh) is essential to maintain muscle progenitor cells and to generate satellite cells. Proc. Natl. Acad. Sci. USA 2007, 104, 4443–4448. [Google Scholar] [CrossRef] [PubMed]
- Nofziger, D.; Miyamoto, A.; Lyons, K.M.; Weinmaster, G. Notch signaling imposes two distinct blocks in the differentiation of C2C12 myoblasts. Development 1999, 126, 1689–1702. [Google Scholar] [PubMed]
- Kuroda, K.; Tani, S.; Tamura, K.; Minoguchi, S.; Kurooka, H.; Honjo, T. Delta-induced Notch signaling mediated by RBP-J inhibits MyoD expression and myogenesis. J. Biol. Chem. 1999, 274, 7238–7244. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Taniguchi, Y.; Kurooka, H.; Minoguchi, S.; Sakai, T.; Nomura-Okazaki, S.; Tamura, K.; Honjo, T. Involvement of RBP-J in biological functions of mouse Notch1 and its derivatives. Development 1997, 124, 4133–4141. [Google Scholar] [PubMed]
- Lindsell, C.E.; Shawber, C.J.; Boulter, J.; Weinmaster, G. Jagged: A mammalian ligand that activates Notch1. Cell 1995, 80, 909–917. [Google Scholar] [CrossRef]
- Shawber, C.; Nofziger, D.; Hsieh, J.J.; Lindsell, C.; Bogler, O.; Hayward, D.; Weinmaster, G. Notch signaling inhibits muscle cell differentiation through a CBF1-independent pathway. Development 1996, 122, 3765–3773. [Google Scholar] [PubMed]
- Delfini, M.C.; Hirsinger, E.; Pourquie, O.; Duprez, D. Delta 1-activated notch inhibits muscle differentiation without affecting Myf5 and Pax3 expression in chick limb myogenesis. Development 2000, 127, 5213–5224. [Google Scholar] [PubMed]
- Hirsinger, E.; Malapert, P.; Dubrulle, J.; Delfini, M.C.; Duprez, D.; Henrique, D.; Ish-Horowicz, D.; Pourquie, O. Notch signalling acts in postmitotic avian myogenic cells to control MyoD activation. Development 2001, 128, 107–116. [Google Scholar] [PubMed]
- Morimoto, M.; Takahashi, Y.; Endo, M.; Saga, Y. The Mesp2 transcription factor establishes segmental borders by suppressing Notch activity. Nature 2005, 435, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Morimoto, M.; Takahashi, Y.; Koseki, H.; Saga, Y. Identification of Epha4 enhancer required for segmental expression and the regulation by Mesp2. Development 2006, 133, 2517–2525. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, M.; Sasaki, N.; Oginuma, M.; Kiso, M.; Igarashi, K.; Aizaki, K.; Kanno, J.; Saga, Y. The negative regulation of Mesp2 by mouse Ripply2 is required to establish the rostro-caudal patterning within a somite. Development 2007, 134, 1561–1569. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, N.; Kiso, M.; Kitagawa, M.; Saga, Y. The repression of Notch signaling occurs via the destabilization of mastermind-like 1 by Mesp2 and is essential for somitogenesis. Development 2011, 138, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Glazier, J.A.; Zhang, Y.; Swat, M.; Zaitlen, B.; Schnell, S. Coordinated action of N-CAM, N-cadherin, EphA4, and ephrinB2 translates genetic prepatterns into structure during somitogenesis in chick. Curr. Top. Dev. Biol. 2008, 81, 205–247. [Google Scholar] [PubMed]
- Takahashi, J.; Ohbayashi, A.; Oginuma, M.; Saito, D.; Mochizuki, A.; Saga, Y.; Takada, S. Analysis of Ripply1/2-deficient mouse embryos reveals a mechanism underlying the rostro-caudal patterning within a somite. Dev. Biol. 2010, 342, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Munsterberg, A.E.; Kitajewski, J.; Bumcrot, D.A.; McMahon, A.P.; Lassar, A.B. Combinatorial signaling by Sonic hedgehog and Wnt family members induces myogenic bHLH gene expression in the somite. Genes Dev. 1995, 9, 2911–2922. [Google Scholar] [CrossRef] [PubMed]
- Brill, G.; Kahane, N.; Carmeli, C.; von Schack, D.; Barde, Y.A.; Kalcheim, C. Epithelial-mesenchymal conversion of dermatome progenitors requires neural tube-derived signals: Characterization of the role of Neurotrophin-3. Development 1995, 121, 2583–2594. [Google Scholar] [PubMed]
- Chiang, C.; Litingtung, Y.; Lee, E.; Young, K.E.; Corden, J.L.; Westphal, H.; Beachy, P.A. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature 1996, 383, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Hamade, A.; Deries, M.; Begemann, G.; Bally-Cuif, L.; Genet, C.; Sabatier, F.; Bonnieu, A.; Cousin, X. Retinoic acid activates myogenesis in vivo through Fgf8 signalling. Dev. Biol. 2006, 289, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, T.J.; Brade, T.; Sandell, L.L.; Lewandoski, M.; Trainor, P.A.; Colas, A.; Mercola, M.; Duester, G. Retinoic Acid Activity in Undifferentiated Neural Progenitors Is Sufficient to Fulfill Its Role in Restricting Fgf8 Expression for Somitogenesis. PLoS ONE 2015, 10, e0137894. [Google Scholar] [CrossRef] [PubMed]
- Du, S.J.; Devoto, S.H.; Westerfield, M.; Moon, R.T. Positive and negative regulation of muscle cell identity by members of the hedgehog and TGF-beta gene families. J. Cell Biol. 1997, 139, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Stern, H.M.; Brown, A.M.; Hauschka, S.D. Myogenesis in paraxial mesoderm: Preferential induction by dorsal neural tube and by cells expressing Wnt-1. Development 1995, 121, 3675–3686. [Google Scholar] [PubMed]
- Duprez, D.; Fournier-Thibault, C.; Le Douarin, N. Sonic Hedgehog induces proliferation of committed skeletal muscle cells in the chick limb. Development 1998, 125, 495–505. [Google Scholar] [PubMed]
- Bren-Mattison, Y.; Olwin, B.B. Sonic hedgehog inhibits the terminal differentiation of limb myoblasts committed to the slow muscle lineage. Dev. Biol. 2002, 242, 130–148. [Google Scholar] [CrossRef] [PubMed]
- Kruger, M.; Mennerich, D.; Fees, S.; Schafer, R.; Mundlos, S.; Braun, T. Sonic hedgehog is a survival factor for hypaxial muscles during mouse development. Development 2001, 128, 743–752. [Google Scholar] [PubMed]
- Kuroda, K.; Kuang, S.; Taketo, M.M.; Rudnicki, M.A. Canonical Wnt signaling induces BMP-4 to specify slow myofibrogenesis of fetal myoblasts. Skelet Muscle 2013, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Ikeya, M.; Takada, S. Wnt signaling from the dorsal neural tube is required for the formation of the medial dermomyotome. Development 1998, 125, 4969–4976. [Google Scholar] [PubMed]
- Tanaka, S.; Terada, K.; Nohno, T. Canonical Wnt signaling is involved in switching from cell proliferation to myogenic differentiation of mouse myoblast cells. J. Mol. Signal. 2011, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.; Suk, S.; Shih, Y.R.; Seo, T.; Du, B.; Xie, Y.; Li, Z.; Varghese, S. WNT3A promotes myogenesis of human embryonic stem cells and enhances in vivo engraftment. Sci. Rep. 2014, 4, 5916. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.T. A role for the Wnt3a/beta-catenin signaling pathway in the myogenic program of C2C12 cells. In Vitro Cell. Dev. Biol. Anim. 2016, 52, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Buikema, J.W.; Mady, A.S.; Mittal, N.V.; Atmanli, A.; Caron, L.; Doevendans, P.A.; Sluijter, J.P.; Domian, I.J. Wnt/beta-catenin signaling directs the regional expansion of first and second heart field-derived ventricular cardiomyocytes. Development 2013, 140, 4165–4176. [Google Scholar] [CrossRef] [PubMed]
- Takata, H.; Terada, K.; Oka, H.; Sunada, Y.; Moriguchi, T.; Nohno, T. Involvement of Wnt4 signaling during myogenic proliferation and differentiation of skeletal muscle. Dev. Dyn. 2007, 236, 2800–2807. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, H.; Gay, S.; Fedon, Y.; Vernus, B.; Bonnieu, A.; Bacou, F. Wnt4 activates the canonical beta-catenin pathway and regulates negatively myostatin: Functional implication in myogenesis. Am. J. Physiol. Cell Physiol. 2011, 300, C1122–C1138. [Google Scholar] [CrossRef] [PubMed]
- Geetha-Loganathan, P.; Nimmagadda, S.; Prols, F.; Patel, K.; Scaal, M.; Huang, R.; Christ, B. Ectodermal Wnt-6 promotes Myf5-dependent avian limb myogenesis. Dev. Biol. 2005, 288, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Anakwe, K.; Robson, L.; Hadley, J.; Buxton, P.; Church, V.; Allen, S.; Hartmann, C.; Harfe, B.; Nohno, T.; Brown, A.M.; et al. Wnt signalling regulates myogenic differentiation in the developing avian wing. Development 2003, 130, 3503–3514. [Google Scholar] [CrossRef] [PubMed]
- Gros, J.; Serralbo, O.; Marcelle, C. WNT11 acts as a directional cue to organize the elongation of early muscle fibres. Nature 2009, 457, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Noden, D.M.; Francis-West, P. The differentiation and morphogenesis of craniofacial muscles. Dev. Dyn. 2006, 235, 1194–1218. [Google Scholar] [CrossRef] [PubMed]
- Hacker, A.; Guthrie, S. A distinct developmental programme for the cranial paraxial mesoderm in the chick embryo. Development 1998, 125, 3461–3472. [Google Scholar] [PubMed]
- Rochais, F.; Mesbah, K.; Kelly, R.G. Signaling pathways controlling second heart field development. Circ. Res. 2009, 104, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Abu-Issa, R.; Smyth, G.; Smoak, I.; Yamamura, K.; Meyers, E.N. Fgf8 is required for pharyngeal arch and cardiovascular development in the mouse. Development 2002, 129, 4613–4625. [Google Scholar] [PubMed]
- Mitsiadis, T.A.; Tucker, A.S.; De Bari, C.; Cobourne, M.T.; Rice, D.P. A regulatory relationship between Tbx1 and FGF signaling during tooth morphogenesis and ameloblast lineage determination. Dev. Biol. 2008, 320, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Scheven, G.; Bothe, I.; Ahmed, M.U.; Alvares, L.E.; Dietrich, S. Protein and genomic organisation of vertebrate MyoR and Capsulin genes and their expression during avian development. Gene Expr. Patterns 2006, 6, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Bothe, I.; Tenin, G.; Oseni, A.; Dietrich, S. Dynamic control of head mesoderm patterning. Development 2011, 138, 2807–2821. [Google Scholar] [CrossRef] [PubMed]
- Mootoosamy, R.C.; Dietrich, S. Distinct regulatory cascades for head and trunk myogenesis. Development 2002, 129, 573–583. [Google Scholar] [PubMed]
- Tzahor, E.; Kempf, H.; Mootoosamy, R.C.; Poon, A.C.; Abzhanov, A.; Tabin, C.J.; Dietrich, S.; Lassar, A.B. Antagonists of Wnt and BMP signaling promote the formation of vertebrate head muscle. Genes Dev. 2003, 17, 3087–3099. [Google Scholar] [CrossRef] [PubMed]
- Harel, I.; Nathan, E.; Tirosh-Finkel, L.; Zigdon, H.; Guimaraes-Camboa, N.; Evans, S.M.; Tzahor, E. Distinct origins and genetic programs of head muscle satellite cells. Dev. Cell 2009, 16, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Sun, X.; Liu, W.; Ai, D.; Klysik, E.; Lu, M.F.; Hadley, J.; Antoni, L.; Chen, L.; Baldini, A.; et al. Pitx2 promotes development of splanchnic mesoderm-derived branchiomeric muscle. Development 2006, 133, 4891–4899. [Google Scholar] [CrossRef] [PubMed]
- Shih, H.P.; Gross, M.K.; Kioussi, C. Cranial muscle defects of Pitx2 mutants result from specification defects in the first branchial arch. Proc. Natl. Acad. Sci. USA 2007, 104, 5907–5912. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.G.; Jerome-Majewska, L.A.; Papaioannou, V.E. The del22q11.2 candidate gene Tbx1 regulates branchiomeric myogenesis. Hum. Mol. Genet. 2004, 13, 2829–2840. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Yamagishi, H.; Maeda, J.; McAnally, J.; Yamagishi, C.; Srivastava, D. Tbx1 regulates fibroblast growth factors in the anterior heart field through a reinforcing autoregulatory loop involving forkhead transcription factors. Development 2004, 131, 5491–5502. [Google Scholar] [CrossRef] [PubMed]
- Schubert, F.R.; Singh, A.J.; Afoyalan, O.; Kioussi, C.; Dietrich, S. To roll the eyes and snap a bite—Function, development and evolution of craniofacial muscles. Semin. Cell Dev. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Bothe, I.; Dietrich, S. The molecular setup of the avian head mesoderm and its implication for craniofacial myogenesis. Dev. Dyn. 2006, 235, 2845–2860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, J.M.; Hawrot, K.; Sharpe, C.; Noble, A.; Wood, W.M.; Jorge, E.C.; Goldhamer, D.J.; Kardon, G.; Dietrich, S. The emergence of Pax7-expressing muscle stem cells during vertebrate head muscle development. Front. Aging Neurosci. 2015, 7, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randolph, M.E.; Phillips, B.L.; Choo, H.J.; Vest, K.E.; Vera, Y.; Pavlath, G.K. Pharyngeal Satellite Cells Undergo Myogenesis Under Basal Conditions and Are Required for Pharyngeal Muscle Maintenance. Stem. Cells 2015, 33, 3581–3595. [Google Scholar] [CrossRef] [PubMed]
- Sambasivan, R.; Kuratani, S.; Tajbakhsh, S. An eye on the head: The development and evolution of craniofacial muscles. Development 2011, 138, 2401–2415. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, T.; Ahlberg, P.E.; Kessaris, N.; Iannarelli, P.; Dennehy, U.; Richardson, W.D.; McMahon, A.P.; Koentges, G. Neural crest origins of the neck and shoulder. Nature 2005, 436, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Pelikan, R.C.; Iwata, J. WNT/beta-Catenin Signaling Regulates Multiple Steps of Myogenesis by Regulating Step-Specific Targets. Mol. Cell Biol. 2015, 35, 1763–1776. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Zhao, H.; Mayo, J.; Chai, Y. Different requirements for Wnt signaling in tongue myogenic subpopulations. J. Dent. Res. 2015, 94, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Tirosh-Finkel, L.; Elhanany, H.; Rinon, A.; Tzahor, E. Mesoderm progenitor cells of common origin contribute to the head musculature and the cardiac outflow tract. Development 2006, 133, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Ryckebusch, L.; Wang, Z.; Bertrand, N.; Lin, S.C.; Chi, X.; Schwartz, R.; Zaffran, S.; Niederreither, K. Retinoic acid deficiency alters second heart field formation. Proc. Natl. Acad. Sci. USA 2008, 105, 2913–2918. [Google Scholar] [CrossRef] [PubMed]
- Hochgreb, T.; Linhares, V.L.; Menezes, D.C.; Sampaio, A.C.; Yan, C.Y.; Cardoso, W.V.; Rosenthal, N.; Xavier-Neto, J. A caudorostral wave of RALDH2 conveys anteroposterior information to the cardiac field. Development 2003, 130, 5363–5374. [Google Scholar] [CrossRef] [PubMed]
- Daughters, R.S.; Keirstead, S.A.; Slack, J.M. Transformation of jaw muscle satellite cells to cardiomyocytes. Differentiation 2017, 93, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Marvin, M.J.; Di Rocco, G.; Gardiner, A.; Bush, S.M.; Lassar, A.B. Inhibition of Wnt activity induces heart formation from posterior mesoderm. Genes Dev. 2001, 15, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Sanchez, C.; Garcia-Martinez, V. Molecular determinants of cardiac specification. Cardiovasc. Res. 2011, 91, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, J.K.; Bruneau, B.G. Directed transdifferentiation of mouse mesoderm to heart tissue by defined factors. Nature 2009, 459, 708–711. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.G.; Sater, A.K. Features of embryonic induction. Development 1988, 104, 341–359. [Google Scholar] [PubMed]
- Garg, V.; Kathiriya, I.S.; Barnes, R.; Schluterman, M.K.; King, I.N.; Butler, C.A.; Rothrock, C.R.; Eapen, R.S.; Hirayama-Yamada, K.; Joo, K.; et al. GATA4 mutations cause human congenital heart defects and reveal an interaction with TBX5. Nature 2003, 424, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Bruneau, B.G.; Nemer, G.; Schmitt, J.P.; Charron, F.; Robitaille, L.; Caron, S.; Conner, D.A.; Gessler, M.; Nemer, M.; Seidman, C.E.; et al. A murine model of Holt-Oram syndrome defines roles of the T-box transcription factor Tbx5 in cardiogenesis and disease. Cell 2001, 106, 709–721. [Google Scholar] [CrossRef]
- Prall, O.W.; Menon, M.K.; Solloway, M.J.; Watanabe, Y.; Zaffran, S.; Bajolle, F.; Biben, C.; McBride, J.J.; Robertson, B.R.; Chaulet, H.; et al. An Nkx2-5/Bmp2/Smad1 negative feedback loop controls heart progenitor specification and proliferation. Cell 2007, 128, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.L.; Liang, X.; Shi, Y.; Chu, P.H.; Pfaff, S.L.; Chen, J.; Evans, S. Isl1 identifies a cardiac progenitor population that proliferates prior to differentiation and contributes a majority of cells to the heart. Dev. Cell 2003, 5, 877–889. [Google Scholar] [CrossRef]
- Theveniau-Ruissy, M.; Dandonneau, M.; Mesbah, K.; Ghez, O.; Mattei, M.G.; Miquerol, L.; Kelly, R.G. The del22q11.2 candidate gene Tbx1 controls regional outflow tract identity and coronary artery patterning. Circ. Res. 2008, 103, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Harel, I.; Maezawa, Y.; Avraham, R.; Rinon, A.; Ma, H.Y.; Cross, J.W.; Leviatan, N.; Hegesh, J.; Roy, A.; Jacob-Hirsch, J. Pharyngeal mesoderm regulatory network controls cardiac and head muscle morphogenesis. Proc. Natl. Acad. Sci. USA 2012, 109, 18839–18844. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Zaffran, S.; Kuroiwa, A.; Higuchi, H.; Ogura, T.; Harvey, R.P.; Kelly, R.G.; Buckingham, M. Fibroblast growth factor 10 gene regulation in the second heart field by Tbx1, Nkx2-5, and Islet1 reveals a genetic switch for down-regulation in the myocardium. Proc. Natl. Acad. Sci. USA 2012, 109, 18273–18280. [Google Scholar] [CrossRef] [PubMed]
- Doetschman, T.; Barnett, J.V.; Runyan, R.B.; Camenisch, T.D.; Heimark, R.L.; Granzier, H.L.; Conway, S.J.; Azhar, M. Transforming growth factor beta signaling in adult cardiovascular diseases and repair. Cell Tissue Res. 2012, 347, 203–223. [Google Scholar] [CrossRef] [PubMed]
- Bartram, U.; Molin, D.G.; Wisse, L.J.; Mohamad, A.; Sanford, L.P.; Doetschman, T.; Speer, C.P.; Poelmann, R.E.; Gittenberger-de Groot, A.C. Double-outlet right ventricle and overriding tricuspid valve reflect disturbances of looping, myocardialization, endocardial cushion differentiation, and apoptosis in TGF-beta(2)-knockout mice. Circulation 2001, 103, 2745–2752. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, S.L.; Janardhan, H.P.; Trivedi, C.M. Histone Deacetylase 3 Coordinates Deacetylase-independent Epigenetic Silencing of Transforming Growth Factor-beta1 (TGF-beta1) to Orchestrate Second Heart Field Development. J. Biol. Chem. 2015, 290, 27067–27089. [Google Scholar] [CrossRef] [PubMed]
- Dickson, M.C.; Martin, J.S.; Cousins, F.M.; Kulkarni, A.B.; Karlsson, S.; Akhurst, R.J. Defective haematopoiesis and vasculogenesis in transforming growth factor-beta 1 knock out mice. Development 1995, 121, 1845–1854. [Google Scholar] [PubMed]
- Trainor, P.A.; Tam, P.P. Cranial paraxial mesoderm and neural crest cells of the mouse embryo: co-distribution in the craniofacial mesenchyme but distinct segregation in branchial arches. Development 1995, 121, 2569–2582. [Google Scholar] [PubMed]
- Kirby, M.L.; Gale, T.F.; Stewart, D.E. Neural crest cells contribute to normal aorticopulmonary septation. Science 1983, 220, 1059–1061. [Google Scholar] [CrossRef] [PubMed]
- Willems, E.; Cabral-Teixeira, J.; Schade, D.; Cai, W.; Reeves, P.; Bushway, P.J.; Lanier, M.; Walsh, C.; Kirchhausen, T.; Izpisua Belmonte, J.C.; et al. Small molecule-mediated TGF-beta type II receptor degradation promotes cardiomyogenesis in embryonic stem cells. Cell Stem Cell 2012, 11, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Gays, D.; Hess, C.; Camporeale, A.; Ala, U.; Provero, P.; Mosimann, C.; Santoro, M.M. An exclusive cellular and molecular network governs intestinal smooth muscle cell differentiation in vertebrates. Development 2017, 144, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Pouget, C.; Gautier, R.; Teillet, M.A.; Jaffredo, T. Somite-derived cells replace ventral aortic hemangioblasts and provide aortic smooth muscle cells of the trunk. Development 2006, 133, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Waldo, K.L.; Kumiski, D.H.; Wallis, K.T.; Stadt, H.A.; Hutson, M.R.; Platt, D.H.; Kirby, M.L. Conotruncal myocardium arises from a secondary heart field. Development 2001, 128, 3179–3188. [Google Scholar] [PubMed]
- Rosenquist, T.H.; Kirby, M.L.; van Mierop, L.H. Solitary aortic arch artery. A result of surgical ablation of cardiac neural crest and nodose placode in the avian embryo. Circulation 1989, 80, 1469–1475. [Google Scholar] [CrossRef] [PubMed]
- Le Lievre, C.S.; Le Douarin, N.M. Mesenchymal derivatives of the neural crest: Analysis of chimaeric quail and chick embryos. J. Embryol. Exp. Morphol. 1975, 34, 125–154. [Google Scholar] [PubMed]
- Owens, G.K.; Wise, G. Regulation of differentiation/maturation in vascular smooth muscle cells by hormones and growth factors. Agents Actions Suppl. 1997, 48, 3–24. [Google Scholar] [PubMed]
- Miano, J.M.; Long, X.; Fujiwara, K. Serum response factor: Master regulator of the actin cytoskeleton and contractile apparatus. Am. J. Physiol. Cell Physiol. 2007, 292, C70–C81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yang, M.; Liu, L.P.; Lau, W.B.; Gao, H.; Xin, M.K.; Su, L.X.; Wang, J.; Cheng, S.J.; Fan, Q.; et al. BMP-2 overexpression augments vascular smooth muscle cell motility by upregulating myosin Va via Erk signaling. Oxid. Med. Cell. Longev. 2014, 2014, 294150. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Noseda, M.; Higginson, M.; Ly, M.; Patenaude, A.; Fuller, M.; Kyle, A.H.; Minchinton, A.I.; Puri, M.C.; Dumont, D.J.; et al. Differentiation of vascular smooth muscle cells from local precursors during embryonic and adult arteriogenesis requires Notch signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 6993–6998. [Google Scholar] [CrossRef] [PubMed]
- Lavine, K.J.; Long, F.; Choi, K.; Smith, C.; Ornitz, D.M. Hedgehog signaling to distinct cell types differentially regulates coronary artery and vein development. Development 2008, 135, 3161–3171. [Google Scholar] [CrossRef] [PubMed]
- Topouzis, S.; Majesky, M.W. Smooth muscle lineage diversity in the chick embryo. Two types of aortic smooth muscle cell differ in growth and receptor-mediated transcriptional responses to transforming growth factor-beta. Dev. Biol. 1996, 178, 430–445. [Google Scholar] [CrossRef] [PubMed]
- Hautmann, M.B.; Madsen, C.S.; Owens, G.K. A transforming growth factor beta (TGFbeta) control element drives TGFbeta-induced stimulation of smooth muscle alpha-actin gene expression in concert with two CArG elements. J. Biol. Chem. 1997, 272, 10948–10956. [Google Scholar] [CrossRef] [PubMed]
- Hirschi, K.K.; Rohovsky, S.A.; D’Amore, P.A. PDGF, TGF-beta, and heterotypic cell-cell interactions mediate endothelial cell-induced recruitment of 10T1/2 cells and their differentiation to a smooth muscle fate. J. Cell Biol. 1998, 141, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.Y.; Lam, S.S.; Correia, J.J.; Lin, K. Smad3 allostery links TGF-beta receptor kinase activation to transcriptional control. Genes Dev. 2002, 16, 1950–1963. [Google Scholar] [CrossRef] [PubMed]
- Tsapara, A.; Luthert, P.; Greenwood, J.; Hill, C.S.; Matter, K.; Balda, M.S. The RhoA activator GEF-H1/Lfc is a transforming growth factor-beta target gene and effector that regulates alpha-smooth muscle actin expression and cell migration. Mol. Biol. Cell 2010, 21, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Young, K.; Krebs, L.T.; Tweedie, E.; Conley, B.; Mancini, M.; Arthur, H.M.; Liaw, L.; Gridley, T.; Vary, C. Endoglin is required in Pax3-derived cells for embryonic blood vessel formation. Dev. Biol. 2016, 409, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.J.; Chang, C.N.; Ma, H.Y.; Ramsey, S.A.; Flitz, T.M.; Kioussi, C. FACS-Seq analysis of Pax3-derived cells identifies non-myogenic lineages in the embryonic forelimb. Sci. Rep. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Cossu, G.; Tajbakhsh, S.; Buckingham, M. How is myogenesis initiated in the embryo? Trends Genet. 1996, 12, 218–223. [Google Scholar] [CrossRef]
- Brent, A.E.; Schweitzer, R.; Tabin, C.J. A somitic compartment of tendon progenitors. Cell 2003, 113, 235–248. [Google Scholar] [CrossRef]
- Tajbakhsh, S.; Bober, E.; Babinet, C.; Pournin, S.; Arnold, H.; Buckingham, M. Gene targeting the myf-5 locus with nlacZ reveals expression of this myogenic factor in mature skeletal muscle fibres as well as early embryonic muscle. Dev. Dyn. 1996, 206, 291–300. [Google Scholar] [CrossRef]
- Gros, J.; Manceau, M.; Thome, V.; Marcelle, C. A common somitic origin for embryonic muscle progenitors and satellite cells. Nature 2005, 435, 954–958. [Google Scholar] [CrossRef] [PubMed]
- Oates, A.C.; Morelli, L.G.; Ares, S. Patterning embryos with oscillations: Structure, function and dynamics of the vertebrate segmentation clock. Development 2012, 139, 625–639. [Google Scholar] [CrossRef] [PubMed]
- Mallo, M. Revisiting the involvement of signaling gradients in somitogenesis. FEBS J. 2016, 283, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Palmeirim, I.; Henrique, D.; Ish-Horowicz, D.; Pourquie, O. Avian hairy gene expression identifies a molecular clock linked to vertebrate segmentation and somitogenesis. Cell 1997, 91, 639–648. [Google Scholar] [CrossRef]
- Resende, T.P.; Ferreira, M.; Teillet, M.A.; Tavares, A.T.; Andrade, R.P.; Palmeirim, I. Sonic hedgehog in temporal control of somite formation. Proc. Natl. Acad. Sci. USA 2010, 107, 12907–12912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aulehla, A.; Herrmann, B.G. Segmentation in vertebrates: Clock and gradient finally joined. Genes Dev. 2004, 18, 2060–2067. [Google Scholar] [CrossRef] [PubMed]
- Ferjentsik, Z.; Hayashi, S.; Dale, J.K.; Bessho, Y.; Herreman, A.; De Strooper, B.; del Monte, G.; de la Pompa, J.L.; Maroto, M. Notch is a critical component of the mouse somitogenesis oscillator and is essential for the formation of the somites. PLoS Genet. 2009, 5, e1000662. [Google Scholar] [CrossRef] [PubMed]
- Diez del Corral, R.; Olivera-Martinez, I.; Goriely, A.; Gale, E.; Maden, M.; Storey, K. Opposing FGF and retinoid pathways control ventral neural pattern, neuronal differentiation, and segmentation during body axis extension. Neuron 2003, 40, 65–79. [Google Scholar] [CrossRef]
- Watanabe, T.; Takahashi, Y. Tissue morphogenesis coupled with cell shape changes. Curr. Opin. Genet. Dev. 2010, 20, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Yabe, T.; Hoshijima, K.; Yamamoto, T.; Takada, S. Quadruple zebrafish mutant reveals different roles of Mesp genes in somite segmentation between mouse and zebrafish. Development 2016, 143, 2842–2852. [Google Scholar] [CrossRef] [PubMed]
- Marcelino, J.; Sciortino, C.M.; Romero, M.F.; Ulatowski, L.M.; Ballock, R.T.; Economides, A.N.; Eimon, P.M.; Harland, R.M.; Warman, M.L. Human disease-causing NOG missense mutations: Effects on noggin secretion, dimer formation, and bone morphogenetic protein binding. Proc. Natl. Acad. Sci. USA 2001, 98, 11353–11358. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.; Brunet, L.J.; Howard, O.; Burton, Q.; Wu, D.K. Role of hindbrain in inner ear morphogenesis: Analysis of Noggin knockout mice. Dev. Biol. 2007, 311, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.M.; Tessier-Lavigne, M. Patterning of mammalian somites by surface ectoderm and notochord: Evidence for sclerotome induction by a hedgehog homolog. Cell 1994, 79, 1175–1186. [Google Scholar] [CrossRef]
- Johnson, R.L.; Laufer, E.; Riddle, R.D.; Tabin, C. Ectopic expression of Sonic hedgehog alters dorsal-ventral patterning of somites. Cell 1994, 79, 1165–1173. [Google Scholar] [CrossRef]
- Chen, C.M.; Kraut, N.; Groudine, M.; Weintraub, H. I-mf, a novel myogenic repressor, interacts with members of the MyoD family. Cell 1996, 86, 731–741. [Google Scholar] [CrossRef]
- Williams, B.A.; Ordahl, C.P. Pax-3 expression in segmental mesoderm marks early stages in myogenic cell specification. Development 1994, 120, 785–796. [Google Scholar] [PubMed]
- Kassar-Duchossoy, L.; Giacone, E.; Gayraud-Morel, B.; Jory, A.; Gomes, D.; Tajbakhsh, S. Pax3/Pax7 mark a novel population of primitive myogenic cells during development. Genes Dev. 2005, 19, 1426–1431. [Google Scholar] [CrossRef] [PubMed]
- Etchevers, H.C.; Vincent, C.; Le Douarin, N.M.; Couly, G.F. The cephalic neural crest provides pericytes and smooth muscle cells to all blood vessels of the face and forebrain. Development 2001, 128, 1059–1068. [Google Scholar] [PubMed]
- Goupille, O.; Pallafacchina, G.; Relaix, F.; Conway, S.J.; Cumano, A.; Robert, B.; Montarras, D.; Buckingham, M. Characterization of Pax3-expressing cells from adult blood vessels. J. Cell Sci. 2011, 124, 3980–3988. [Google Scholar] [CrossRef] [PubMed]
- Goulding, M.; Lumsden, A.; Paquette, A.J. Regulation of Pax-3 expression in the dermomyotome and its role in muscle development. Development 1994, 120, 957–971. [Google Scholar] [PubMed]
- Bober, E.; Franz, T.; Arnold, H.H.; Gruss, P.; Tremblay, P. Pax-3 is required for the development of limb muscles: A possible role for the migration of dermomyotomal muscle progenitor cells. Development 1994, 120, 603–612. [Google Scholar] [PubMed]
- Grifone, R.; Demignon, J.; Houbron, C.; Souil, E.; Niro, C.; Seller, M.J.; Hamard, G.; Maire, P. Six1 and Six4 homeoproteins are required for Pax3 and Mrf expression during myogenesis in the mouse embryo. Development 2005, 132, 2235–2249. [Google Scholar] [CrossRef] [PubMed]
- Grifone, R.; Kelly, R.G. Heartening news for head muscle development. Trends Genet. 2007, 23, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Ott, M.O.; Bober, E.; Lyons, G.; Arnold, H.; Buckingham, M. Early expression of the myogenic regulatory gene, myf-5, in precursor cells of skeletal muscle in the mouse embryo. Development 1991, 111, 1097–1107. [Google Scholar] [PubMed]
- Tajbakhsh, S.; Rocancourt, D.; Cossu, G.; Buckingham, M. Redefining the genetic hierarchies controlling skeletal myogenesis: Pax-3 and Myf-5 act upstream of MyoD. Cell 1997, 89, 127–138. [Google Scholar] [CrossRef]
- Borello, U.; Berarducci, B.; Murphy, P.; Bajard, L.; Buffa, V.; Piccolo, S.; Buckingham, M.; Cossu, G. The Wnt/beta-catenin pathway regulates Gli-mediated Myf5 expression during somitogenesis. Development 2006, 133, 3723–3732. [Google Scholar] [CrossRef] [PubMed]
- Rong, P.M.; Teillet, M.A.; Ziller, C.; Le Douarin, N.M. The neural tube/notochord complex is necessary for vertebral but not limb and body wall striated muscle differentiation. Development 1992, 115, 657–672. [Google Scholar] [PubMed]
- Fontaine-Perus, J.; Halgand, P.; Cheraud, Y.; Rouaud, T.; Velasco, M.E.; Cifuentes Diaz, C.; Rieger, F. Mouse-chick chimera: A developmental model of murine neurogenic cells. Development 1997, 124, 3025–3036. [Google Scholar] [PubMed]
- Pownall, M.E.; Strunk, K.E.; Emerson, C.P., Jr. Notochord signals control the transcriptional cascade of myogenic bHLH genes in somites of quail embryos. Development 1996, 122, 1475–1488. [Google Scholar] [PubMed]
- Borycki, A.G.; Mendham, L.; Emerson, C.P., Jr. Control of somite patterning by Sonic hedgehog and its downstream signal response genes. Development 1998, 125, 777–790. [Google Scholar] [PubMed]
- Dietrich, S.; Schubert, F.R.; Lumsden, A. Control of dorsoventral pattern in the chick paraxial mesoderm. Development 1997, 124, 3895–3908. [Google Scholar] [PubMed]
- Gustafsson, M.K.; Pan, H.; Pinney, D.F.; Liu, Y.; Lewandowski, A.; Epstein, D.J.; Emerson, C.P., Jr. Myf5 is a direct target of long-range Shh signaling and Gli regulation for muscle specification. Genes Dev. 2002, 16, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Teboul, L.; Summerbell, D.; Rigby, P.W. The initial somitic phase of Myf5 expression requires neither Shh signaling nor Gli regulation. Genes Dev. 2003, 17, 2870–2874. [Google Scholar] [CrossRef] [PubMed]
- Rios, A.C.; Serralbo, O.; Salgado, D.; Marcelle, C. Neural crest regulates myogenesis through the transient activation of NOTCH. Nature 2011, 473, 532–535. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Rocancourt, D.; Marques, L.; Thorsteinsdottir, S.; Buckingham, M. A Pax3/Dmrt2/Myf5 regulatory cascade functions at the onset of myogenesis. PLoS Genet. 2010, 6, e1000897. [Google Scholar] [CrossRef] [PubMed]
- Relaix, F.; Montarras, D.; Zaffran, S.; Gayraud-Morel, B.; Rocancourt, D.; Tajbakhsh, S.; Mansouri, A.; Cumano, A.; Buckingham, M. Pax3 and Pax7 have distinct and overlapping functions in adult muscle progenitor cells. J. Cell Biol. 2006, 172, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Ben-Yair, R.; Kalcheim, C. Lineage analysis of the avian dermomyotome sheet reveals the existence of single cells with both dermal and muscle progenitor fates. Development 2005, 132, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Relaix, F.; Rocancourt, D.; Mansouri, A.; Buckingham, M. A Pax3/Pax7-dependent population of skeletal muscle progenitor cells. Nature 2005, 435, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.U.; Maurya, A.K.; Cheng, L.; Jorge, E.C.; Schubert, F.R.; Maire, P.; Basson, M.A.; Ingham, P.W.; Dietrich, S. Engrailed controls epaxial-hypaxial muscle innervation and the establishment of vertebrate three-dimensional mobility. Dev. Biol. 2017, 430, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Zhi, Q.; Patel, K.; Wilting, J.; Christ, B. Dual origin and segmental organisation of the avian scapula. Development 2000, 127, 3789–3794. [Google Scholar] [PubMed]
- Huang, R.; Zhi, Q.; Schmidt, C.; Wilting, J.; Brand-Saberi, B.; Christ, B. Sclerotomal origin of the ribs. Development 2000, 127, 527–532. [Google Scholar] [PubMed]
- Ehehalt, F.; Wang, B.; Christ, B.; Patel, K.; Huang, R. Intrinsic cartilage-forming potential of dermomyotomal cells requires ectodermal signals for the development of the scapula blade. Anat. Embryol. 2004, 208, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Valasek, P.; Theis, S.; DeLaurier, A.; Hinits, Y.; Luke, G.N.; Otto, A.M.; Minchin, J.; He, L.; Christ, B.; Brooks, G.; et al. Cellular and molecular investigations into the development of the pectoral girdle. Dev. Biol. 2011, 357, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Van Ho, A.T.; Hayashi, S.; Brohl, D.; Aurade, F.; Rattenbach, R.; Relaix, F. Neural crest cell lineage restricts skeletal muscle progenitor cell differentiation through Neuregulin1-ErbB3 signaling. Dev. Cell 2011, 21, 273–287. [Google Scholar] [PubMed]
- Li, L.; Cserjesi, P.; Olson, E.N. Dermo-1: A novel twist-related bHLH protein expressed in the developing dermis. Dev. Biol. 1995, 172, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.Q.; Li, L. Dermo-1, a multifunctional basic helix-loop-helix protein, represses MyoD transactivation via the HLH domain, MEF2 interaction, and chromatin deacetylation. J. Biol. Chem. 2002, 277, 12310–12317. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, S.; Schubert, F.R.; Healy, C.; Sharpe, P.T.; Lumsden, A. Specification of the hypaxial musculature. Development 1998, 125, 2235–2249. [Google Scholar] [PubMed]
- Pourquie, O.; Fan, C.M.; Coltey, M.; Hirsinger, E.; Watanabe, Y.; Breant, C.; Francis-West, P.; Brickell, P.; Tessier-Lavigne, M.; Le Douarin, N.M. Lateral and axial signals involved in avian somite patterning: A role for BMP4. Cell 1996, 84, 461–471. [Google Scholar] [CrossRef]
- Ben-Yair, R.; Kalcheim, C. Notch and bone morphogenetic protein differentially act on dermomyotome cells to generate endothelium, smooth, and striated muscle. J. Cell Biol. 2008, 180, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Brunelli, S.; Relaix, F.; Baesso, S.; Buckingham, M.; Cossu, G. Beta catenin-independent activation of MyoD in presomitic mesoderm requires PKC and depends on Pax3 transcriptional activity. Dev. Biol. 2007, 304, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Giordani, J.; Bajard, L.; Demignon, J.; Daubas, P.; Buckingham, M.; Maire, P. Six proteins regulate the activation of Myf5 expression in embryonic mouse limbs. Proc. Natl. Acad. Sci. USA 2007, 104, 11310–11315. [Google Scholar] [CrossRef] [PubMed]
- Bajard, L.; Relaix, F.; Lagha, M.; Rocancourt, D.; Daubas, P.; Buckingham, M.E. A novel genetic hierarchy functions during hypaxial myogenesis: Pax3 directly activates Myf5 in muscle progenitor cells in the limb. Genes Dev. 2006, 20, 2450–2464. [Google Scholar] [CrossRef] [PubMed]
- Bladt, F.; Riethmacher, D.; Isenmann, S.; Aguzzi, A.; Birchmeier, C. Essential role for the c-met receptor in the migration of myogenic precursor cells into the limb bud. Nature 1995, 376, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.M.; Vogan, K.; Gros, P.; Park, M. Expression of the met receptor tyrosine kinase in muscle progenitor cells in somites and limbs is absent in Splotch mice. Development 1996, 122, 2163–2171. [Google Scholar] [PubMed]
- Gross, M.K.; Moran-Rivard, L.; Velasquez, T.; Nakatsu, M.N.; Jagla, K.; Goulding, M. Lbx1 is required for muscle precursor migration along a lateral pathway into the limb. Development 2000, 127, 413–424. [Google Scholar] [PubMed]
- Kioussi, C.; Briata, P.; Baek, S.H.; Rose, D.W.; Hamblet, N.S.; Herman, T.; Ohgi, K.A.; Lin, C.; Gleiberman, A.; Wang, J.; et al. Identification of a Wnt/Dvl/beta-Catenin --> Pitx2 pathway mediating cell-type-specific proliferation during development. Cell 2002, 111, 673–685. [Google Scholar] [CrossRef]
- Shih, H.P.; Gross, M.K.; Kioussi, C. Expression pattern of the homeodomain transcription factor Pitx2 during muscle development. Gene Expr. Patterns 2007, 7, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Lagha, M.; Sato, T.; Regnault, B.; Cumano, A.; Zuniga, A.; Licht, J.; Relaix, F.; Buckingham, M. Transcriptome analyses based on genetic screens for Pax3 myogenic targets in the mouse embryo. BMC Genom. 2010, 11, 696. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.L.; Shih, H.P.; Xu, J.; Gross, M.K.; Kioussi, C. Regulation of motility of myogenic cells in filling limb muscle anlagen by Pitx2. PLoS ONE 2012, 7, e35822. [Google Scholar] [CrossRef] [PubMed]
- Lagha, M.; Mayeuf-Louchart, A.; Chang, T.; Montarras, D.; Rocancourt, D.; Zalc, A.; Kormish, J.; Zaret, K.S.; Buckingham, M.E.; Relaix, F. Itm2a is a Pax3 target gene, expressed at sites of skeletal muscle formation in vivo. PLoS ONE 2013, 8, e63143. [Google Scholar] [CrossRef] [PubMed]
- Van den Plas, D.; Merregaert, J. Constitutive overexpression of the integral membrane protein Itm2A enhances myogenic differentiation of C2C12 cells. Cell Biol. Int. 2004, 28, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Delfini, M.C.; De La Celle, M.; Gros, J.; Serralbo, O.; Marics, I.; Seux, M.; Scaal, M.; Marcelle, C. The timing of emergence of muscle progenitors is controlled by an FGF/ERK/SNAIL1 pathway. Dev. Biol. 2009, 333, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Pu, Q.; Abduelmula, A.; Masyuk, M.; Theiss, C.; Schwandulla, D.; Hans, M.; Patel, K.; Brand-Saberi, B.; Huang, R. The dermomyotome ventrolateral lip is essential for the hypaxial myotome formation. BMC Dev. Biol. 2013, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- Deries, M.; Thorsteinsdottir, S. Axial and limb muscle development: Dialogue with the neighbourhood. Cell. Mol. Life Sci. 2016, 73, 4415–4431. [Google Scholar] [CrossRef] [PubMed]
- Fraidenraich, D.; Iwahori, A.; Rudnicki, M.; Basilico, C. Activation of fgf4 gene expression in the myotomes is regulated by myogenic bHLH factors and by sonic hedgehog. Dev. Biol. 2000, 225, 392–406. [Google Scholar] [CrossRef] [PubMed]
- Kahane, N.; Ribes, V.; Kicheva, A.; Briscoe, J.; Kalcheim, C. The transition from differentiation to growth during dermomyotome-derived myogenesis depends on temporally restricted hedgehog signaling. Development 2013, 140, 1740–1750. [Google Scholar] [CrossRef] [PubMed]
- Sacks, L.D.; Cann, G.M.; Nikovits, W., Jr.; Conlon, S.; Espinoza, N.R.; Stockdale, F.E. Regulation of myosin expression during myotome formation. Development 2003, 130, 3391–3402. [Google Scholar] [CrossRef] [PubMed]
- Niro, C.; Demignon, J.; Vincent, S.; Liu, Y.; Giordani, J.; Sgarioto, N.; Favier, M.; Guillet-Deniau, I.; Blais, A.; Maire, P. Six1 and Six4 gene expression is necessary to activate the fast-type muscle gene program in the mouse primary myotome. Dev. Biol. 2010, 338, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Bryson-Richardson, R.J.; Currie, P.D. The genetics of vertebrate myogenesis. Nat. Rev. Genet. 2008, 9, 632–646. [Google Scholar] [CrossRef] [PubMed]
- Relaix, F.; Rocancourt, D.; Mansouri, A.; Buckingham, M. Divergent functions of murine Pax3 and Pax7 in limb muscle development. Genes Dev. 2004, 18, 1088–1105. [Google Scholar] [CrossRef] [PubMed]
- Brohmann, H.; Jagla, K.; Birchmeier, C. The role of Lbx1 in migration of muscle precursor cells. Development 2000, 127, 437–445. [Google Scholar] [PubMed]
- Amthor, H.; Christ, B.; Patel, K. A molecular mechanism enabling continuous embryonic muscle growth—A balance between proliferation and differentiation. Development 1999, 126, 1041–1053. [Google Scholar] [PubMed]
- Bendall, A.J.; Ding, J.; Hu, G.; Shen, M.M.; Abate-Shen, C. Msx1 antagonizes the myogenic activity of Pax3 in migrating limb muscle precursors. Development 1999, 126, 4965–4976. [Google Scholar] [PubMed]
- Riddle, R.D.; Johnson, R.L.; Laufer, E.; Tabin, C. Sonic hedgehog mediates the polarizing activity of the ZPA. Cell 1993, 75, 1401–1416. [Google Scholar] [CrossRef]
- Niswander, L.; Jeffrey, S.; Martin, G.R.; Tickle, C. A positive feedback loop coordinates growth and patterning in the vertebrate limb. Nature 1994, 371, 609–612. [Google Scholar] [CrossRef] [PubMed]
- Scaal, M.; Bonafede, A.; Dathe, V.; Sachs, M.; Cann, G.; Christ, B.; Brand-Saberi, B. SF/HGF is a mediator between limb patterning and muscle development. Development 1999, 126, 4885–4893. [Google Scholar] [PubMed]
- Vasyutina, E.; Stebler, J.; Brand-Saberi, B.; Schulz, S.; Raz, E.; Birchmeier, C. CXCR4 and Gab1 cooperate to control the development of migrating muscle progenitor cells. Genes Dev. 2005, 19, 2187–2198. [Google Scholar] [CrossRef] [PubMed]
- Linker, C.; Lesbros, C.; Stark, M.R.; Marcelle, C. Intrinsic signals regulate the initial steps of myogenesis in vertebrates. Development 2003, 130, 4797–4807. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.J.; Johnson, S.E.; Hannon, K. Insulin-like growth factor I stimulates myoblast expansion and myofiber development in the limb. Dev. Dyn. 2002, 223, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, R.H.; Anderton, H.; Brameld, J.M.; Sweetman, D. Correction: Effects of insulin like growth factors on early embryonic chick limb myogenesis. PLoS ONE 2017, 12, e0189395. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.; Williams, V.C.; Moyon, B.; Daubas, P.; Tajbakhsh, S.; Buckingham, M.E.; Shiroishi, T.; Hughes, S.M.; Borycki, A.G. Sonic hedgehog acts cell-autonomously on muscle precursor cells to generate limb muscle diversity. Genes Dev. 2012, 26, 2103–2117. [Google Scholar] [CrossRef] [PubMed]
- Sassoon, D.; Lyons, G.; Wright, W.E.; Lin, V.; Lassar, A.; Weintraub, H.; Buckingham, M. Expression of two myogenic regulatory factors myogenin and MyoD1 during mouse embryogenesis. Nature 1989, 341, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Menko, A.S.; Boettiger, D. Occupation of the extracellular matrix receptor, integrin, is a control point for myogenic differentiation. Cell 1987, 51, 51–57. [Google Scholar] [CrossRef]
- Boettiger, D.; Enomoto-Iwamoto, M.; Yoon, H.Y.; Hofer, U.; Menko, A.S.; Chiquet-Ehrismann, R. Regulation of integrin alpha 5 beta 1 affinity during myogenic differentiation. Dev. Biol. 1995, 169, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, K.A. The calcium-dependent myoblast adhesion that precedes cell fusion is mediated by glycoproteins. J. Cell Biol. 1985, 101, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Spitz, F.; Demignon, J.; Porteu, A.; Kahn, A.; Concordet, J.P.; Daegelen, D.; Maire, P. Expression of myogenin during embryogenesis is controlled by Six/sine oculis homeoproteins through a conserved MEF3 binding site. Proc. Natl. Acad. Sci. USA 1998, 95, 14220–14225. [Google Scholar] [CrossRef] [PubMed]
- Hutcheson, D.A.; Zhao, J.; Merrell, A.; Haldar, M.; Kardon, G. Embryonic and fetal limb myogenic cells are derived from developmentally distinct progenitors and have different requirements for beta-catenin. Genes Dev. 2009, 23, 997–1013. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Sabourin, L.A.; Girgis-Gabardo, A.; Mansouri, A.; Gruss, P.; Rudnicki, M.A. Pax7 is required for the specification of myogenic satellite cells. Cell 2000, 102, 777–786. [Google Scholar] [CrossRef]
- Gunther, S.; Kim, J.; Kostin, S.; Lepper, C.; Fan, C.M.; Braun, T. Myf5-positive satellite cells contribute to Pax7-dependent long-term maintenance of adult muscle stem cells. Cell Stem Cell 2013, 13, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, V.D.; Punch, V.G.; Kawabe, Y.; Jones, A.E.; Palidwor, G.A.; Porter, C.J.; Cross, J.W.; Carvajal, J.J.; Kockx, C.E.; van IJcken, W.F.; et al. Transcriptional dominance of Pax7 in adult myogenesis is due to high-affinity recognition of homeodomain motifs. Dev. Cell 2012, 22, 1208–1220. [Google Scholar] [CrossRef] [PubMed]
- Von Maltzahn, J.; Jones, A.E.; Parks, R.J.; Rudnicki, M.A. Pax7 is critical for the normal function of satellite cells in adult skeletal muscle. Proc. Natl. Acad. Sci. USA 2013, 110, 16474–16479. [Google Scholar] [CrossRef] [PubMed]
- Oustanina, S.; Hause, G.; Braun, T. Pax7 directs postnatal renewal and propagation of myogenic satellite cells but not their specification. EMBO J. 2004, 23, 3430–3439. [Google Scholar] [CrossRef] [PubMed]
- Kuang, S.; Charge, S.B.; Seale, P.; Huh, M.; Rudnicki, M.A. Distinct roles for Pax7 and Pax3 in adult regenerative myogenesis. J. Cell Biol. 2006, 172, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.F.; Li, L.; Olson, E.N. Repression of myogenin function by TGF-beta 1 is targeted at the basic helix-loop-helix motif and is independent of E2A products. J. Biol. Chem. 1992, 267, 10956–10960. [Google Scholar] [PubMed]
- Bischoff, R. Chemotaxis of skeletal muscle satellite cells. Dev. Dyn. 1997, 208, 505–515. [Google Scholar] [CrossRef]
- Yamagishi, T.; Ando, K.; Nakamura, H.; Nakajima, Y. Expression of the Tgfbeta2 gene during chick embryogenesis. Anat. Rec. 2012, 295, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Messina, G.; Biressi, S.; Monteverde, S.; Magli, A.; Cassano, M.; Perani, L.; Roncaglia, E.; Tagliafico, E.; Starnes, L.; Campbell, C.E.; et al. Nfix regulates fetal-specific transcription in developing skeletal muscle. Cell 2010, 140, 554–566. [Google Scholar] [CrossRef] [PubMed]
- L’Honore, A.; Coulon, V.; Marcil, A.; Lebel, M.; Lafrance-Vanasse, J.; Gage, P.; Camper, S.; Drouin, J. Sequential expression and redundancy of Pitx2 and Pitx3 genes during muscle development. Dev. Biol. 2007, 307, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Duxson, M.J.; Usson, Y.; Harris, A.J. The origin of secondary myotubes in mammalian skeletal muscles: Ultrastructural studies. Development 1989, 107, 743–750. [Google Scholar] [PubMed]
- De Angelis, L.; Berghella, L.; Coletta, M.; Lattanzi, L.; Zanchi, M.; Cusella-De Angelis, M.G.; Ponzetto, C.; Cossu, G. Skeletal myogenic progenitors originating from embryonic dorsal aorta coexpress endothelial and myogenic markers and contribute to postnatal muscle growth and regeneration. J. Cell Biol. 1999, 147, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, J.R.; Heslop, L.; Yu, D.S.; Tajbakhsh, S.; Kelly, R.G.; Wernig, A.; Buckingham, M.E.; Partridge, T.A.; Zammit, P.S. Expression of CD34 and Myf5 defines the majority of quiescent adult skeletal muscle satellite cells. J. Cell Biol. 2000, 151, 1221–1234. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Velasco, E.; Vallejo, D.; Esteban, F.J.; Doherty, C.; Hernandez-Torres, F.; Franco, D.; Aranega, A.E. A Pitx2-MicroRNA Pathway Modulates Cell Proliferation in Myoblasts and Skeletal-Muscle Satellite Cells and Promotes Their Commitment to a Myogenic Cell Fate. Mol. Cell Biol. 2015, 35, 2892–2909. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, C.; Hennebry, A.; Thomas, M.; Plummer, E.; Ling, N.; Sharma, M.; Kambadur, R. Myostatin signals through Pax7 to regulate satellite cell self-renewal. Exp. Cell Res. 2008, 314, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Rebbapragada, A.; Benchabane, H.; Wrana, J.L.; Celeste, A.J.; Attisano, L. Myostatin signals through a transforming growth factor beta-like signaling pathway to block adipogenesis. Mol. Cell. Biol. 2003, 23, 7230–7242. [Google Scholar] [CrossRef] [PubMed]
- Massague, J.; Seoane, J.; Wotton, D. Smad transcription factors. Genes Dev. 2005, 19, 2783–2810. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Black, B.L.; Derynck, R. TGF-beta inhibits muscle differentiation through functional repression of myogenic transcription factors by Smad3. Genes Dev. 2001, 15, 2950–2966. [Google Scholar] [CrossRef] [PubMed]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef] [PubMed]
- McPherron, A.C.; Lee, S.J. Double muscling in cattle due to mutations in the myostatin gene. Proc. Natl. Acad. Sci. USA 1997, 94, 12457–12461. [Google Scholar] [CrossRef] [PubMed]
- Sartori, R.; Milan, G.; Patron, M.; Mammucari, C.; Blaauw, B.; Abraham, R.; Sandri, M. Smad2 and 3 transcription factors control muscle mass in adulthood. Am. J. Physiol. Cell Physiol. 2009, 296, C1248–C1257. [Google Scholar] [CrossRef] [PubMed]
- Iezzi, S.; Di Padova, M.; Serra, C.; Caretti, G.; Simone, C.; Maklan, E.; Minetti, G.; Zhao, P.; Hoffman, E.P.; Puri, P.L.; et al. Deacetylase inhibitors increase muscle cell size by promoting myoblast recruitment and fusion through induction of follistatin. Dev. Cell 2004, 6, 673–684. [Google Scholar] [CrossRef]
- Pisconti, A.; Brunelli, S.; Di Padova, M.; De Palma, C.; Deponti, D.; Baesso, S.; Sartorelli, V.; Cossu, G.; Clementi, E. Follistatin induction by nitric oxide through cyclic GMP: A tightly regulated signaling pathway that controls myoblast fusion. J. Cell Biol. 2006, 172, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Fainsod, A.; Deissler, K.; Yelin, R.; Marom, K.; Epstein, M.; Pillemer, G.; Steinbeisser, H.; Blum, M. The dorsalizing and neural inducing gene follistatin is an antagonist of BMP-4. Mech. Dev. 1997, 63, 39–50. [Google Scholar] [CrossRef]
- Amthor, H.; Nicholas, G.; McKinnell, I.; Kemp, C.F.; Sharma, M.; Kambadur, R.; Patel, K. Follistatin complexes Myostatin and antagonises Myostatin-mediated inhibition of myogenesis. Dev. Biol. 2004, 270, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Shi, X.E.; Song, C.; Sun, S.; Yang, G.; Li, X. BAMBI Promotes C2C12 Myogenic Differentiation by Enhancing Wnt/beta-Catenin Signaling. Int. J. Mol. Sci. 2015, 16, 17734–17745. [Google Scholar] [CrossRef] [PubMed]
- Christ, B.; Jacob, M.; Jacob, H.J. On the origin and development of the ventrolateral abdominal muscles in the avian embryo. An experimental and ultrastructural study. Anat. Embryol. 1983, 166, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Eng, D.; Ma, H.Y.; Xu, J.; Shih, H.P.; Gross, M.K.; Kioussi, C. Loss of abdominal muscle in Pitx2 mutants associated with altered axial specification of lateral plate mesoderm. PLoS ONE 2012, 7, e42228. [Google Scholar] [CrossRef]
- Sadler, T.W. The embryologic origin of ventral body wall defects. Semin. Pediatr. Surg. 2010, 19, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Brewer, S.; Williams, T. Finally, a sense of closure? Animal models of human ventral body wall defects. Bioessays 2004, 26, 1307–1321. [Google Scholar] [CrossRef] [PubMed]
- Sadler, T.W.; Feldkamp, M.L. The embryology of body wall closure: Relevance to gastroschisis and other ventral body wall defects. Am. J. Med. Genet. C Semin. Med. Genet. 2008, 148, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, H.; Yu, J.; Cao, J.; Chen, H.; Zhao, H.; Zhao, J.; Yao, Y.; Cheng, H.; Wang, L.; et al. Ectodermal Wnt signaling regulates abdominal myogenesis during ventral body wall development. Dev. Biol. 2014, 387, 64–72. [Google Scholar] [CrossRef] [PubMed]
- L’Honore, A.; Ouimette, J.F.; Lavertu-Jolin, M.; Drouin, J. Pitx2 defines alternate pathways acting through MyoD during limb and somitic myogenesis. Development 2010, 137, 3847–3856. [Google Scholar] [CrossRef] [PubMed]
- Hilton, T.; Gross, M.K.; Kioussi, C. Pitx2-dependent occupancy by histone deacetylases is associated with T-box gene regulation in mammalian abdominal tissue. J. Biol. Chem. 2010, 285, 11129–11142. [Google Scholar] [CrossRef] [PubMed]
- Brewer, S.; Williams, T. Loss of AP-2alpha impacts multiple aspects of ventral body wall development and closure. Dev. Biol. 2004, 267, 399–417. [Google Scholar] [CrossRef] [PubMed]
- Ten Berge, D.; Brouwer, A.; el Bahi, S.; Guenet, J.L.; Robert, B.; Meijlink, F. Mouse Alx3: An aristaless-like homeobox gene expressed during embryogenesis in ectomesenchyme and lateral plate mesoderm. Dev. Biol. 1998, 199, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Niswender, K.D.; Ji, Q.; van der Meer, R.; Keeney, D.; Magnuson, M.A.; Wisdom, R. Polydactyly and ectopic ZPA formation in Alx-4 mutant mice. Development 1997, 124, 3999–4008. [Google Scholar] [PubMed]
- Doi, T.; Puri, P.; Bannigan, J.; Thompson, J. HoxB2, HoxB4 and Alx4 genes are downregulated in the cadmium-induced omphalocele in the chick model. Pediatr. Surg. Int. 2010, 26, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Manley, N.R.; Barrow, J.R.; Zhang, T.; Capecchi, M.R. Hoxb2 and hoxb4 act together to specify ventral body wall formation. Dev. Biol. 2001, 237, 130–144. [Google Scholar] [CrossRef] [PubMed]
- Barrow, J.R.; Capecchi, M.R. Targeted disruption of the Hoxb-2 locus in mice interferes with expression of Hoxb-1 and Hoxb-4. Development 1996, 122, 3817–3828. [Google Scholar] [PubMed]
- Kitamura, K.; Miura, H.; Miyagawa-Tomita, S.; Yanazawa, M.; Katoh-Fukui, Y.; Suzuki, R.; Ohuchi, H.; Suehiro, A.; Motegi, Y.; Nakahara, Y.; et al. Mouse Pitx2 deficiency leads to anomalies of the ventral body wall, heart, extra- and periocular mesoderm and right pulmonary isomerism. Development 1999, 126, 5749–5758. [Google Scholar] [PubMed]
- Hogan, B.L. Bone morphogenetic proteins: Multifunctional regulators of vertebrate development. Genes Dev. 1996, 10, 1580–1594. [Google Scholar] [CrossRef] [PubMed]
- Dickson, M.C.; Slager, H.G.; Duffie, E.; Mummery, G.; Mummery, C.L.; Akhurst, R.J. RNA and protein localisations of TGF beta 2 in the early mouse embryo suggest an involvement in cardiac development. Development 1993, 117, 625–639. [Google Scholar] [PubMed]
- Aldeiri, B.; Roostalu, U.; Albertini, A.; Wong, J.; Morabito, A.; Cossu, G. Transgelin-expressing myofibroblasts orchestrate ventral midline closure through TGFbeta signalling. Development 2017, 144, 3336–3348. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Labosky, P.A.; Furuta, Y.; Hargett, L.; Dunn, R.; Fogo, A.B.; Takahara, K.; Peters, D.M.; Greenspan, D.S.; Hogan, B.L. Failure of ventral body wall closure in mouse embryos lacking a procollagen C-proteinase encoded by Bmp1, a mammalian gene related to Drosophila tolloid. Development 1996, 122, 3587–3595. [Google Scholar] [PubMed]
- Zhang, J.; Hagopian-Donaldson, S.; Serbedzija, G.; Elsemore, J.; Plehn-Dujowich, D.; McMahon, A.P.; Flavell, R.A.; Williams, T. Neural tube, skeletal and body wall defects in mice lacking transcription factor AP-2. Nature 1996, 381, 238–241. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Signaling Molecules in Myogenesis (a) Paraxial mesoderm (PM) located next to the neural tube (NT). Dorsal PM gives rise to head and pharyngeal muscles while dorsal PM gives rise to somites. Intermediate mesoderm (IM) is located between the PM and the lateral plate mesoderm (LPM). LPM gives rise to the vascular system and skeletal muscles of the abdomen pelvis and some limb muscle. NC, notochord; (b) Anterior-posterior pattern of skeletal muscle is initiated by the RA/FGF8 inhibitory network, with AR promoting the anterior muscles and FGF8 the posterior. Activation of Wnt promotes the central somites and NOTCH signaling that will support formation of the anterior and posterior somites via EPH4 and RIPPLY, respectively; (c) Segmentation of somites into dermomyotome (DM), myotome (MT) and sclerotome (SC) is the result of signaling molecules secreted from the dorsal NT (BMP, WNT1/3a), notochord (SHH), ectoderm (WNT6/11, WNT7a, BMP4), and LPM (BMP4, FGF5). DML, dorsomedial lip; VLL, Ventrolateral lip.
Figure 1.
Signaling Molecules in Myogenesis (a) Paraxial mesoderm (PM) located next to the neural tube (NT). Dorsal PM gives rise to head and pharyngeal muscles while dorsal PM gives rise to somites. Intermediate mesoderm (IM) is located between the PM and the lateral plate mesoderm (LPM). LPM gives rise to the vascular system and skeletal muscles of the abdomen pelvis and some limb muscle. NC, notochord; (b) Anterior-posterior pattern of skeletal muscle is initiated by the RA/FGF8 inhibitory network, with AR promoting the anterior muscles and FGF8 the posterior. Activation of Wnt promotes the central somites and NOTCH signaling that will support formation of the anterior and posterior somites via EPH4 and RIPPLY, respectively; (c) Segmentation of somites into dermomyotome (DM), myotome (MT) and sclerotome (SC) is the result of signaling molecules secreted from the dorsal NT (BMP, WNT1/3a), notochord (SHH), ectoderm (WNT6/11, WNT7a, BMP4), and LPM (BMP4, FGF5). DML, dorsomedial lip; VLL, Ventrolateral lip.

Figure 2.
Gene Networks Involved in Muscle Specification. LacZ staining of E12.5 Pitx2LacZ/+ mouse. Extraocular muscles (EOM) and abdominal wall muscles are specified by PITX2. Branchial arch muscles are specified by PITX2 and TBX1. Forelimb, trunk and hindlimb muscles are specified by PAX3.
Figure 2.
Gene Networks Involved in Muscle Specification. LacZ staining of E12.5 Pitx2LacZ/+ mouse. Extraocular muscles (EOM) and abdominal wall muscles are specified by PITX2. Branchial arch muscles are specified by PITX2 and TBX1. Forelimb, trunk and hindlimb muscles are specified by PAX3.


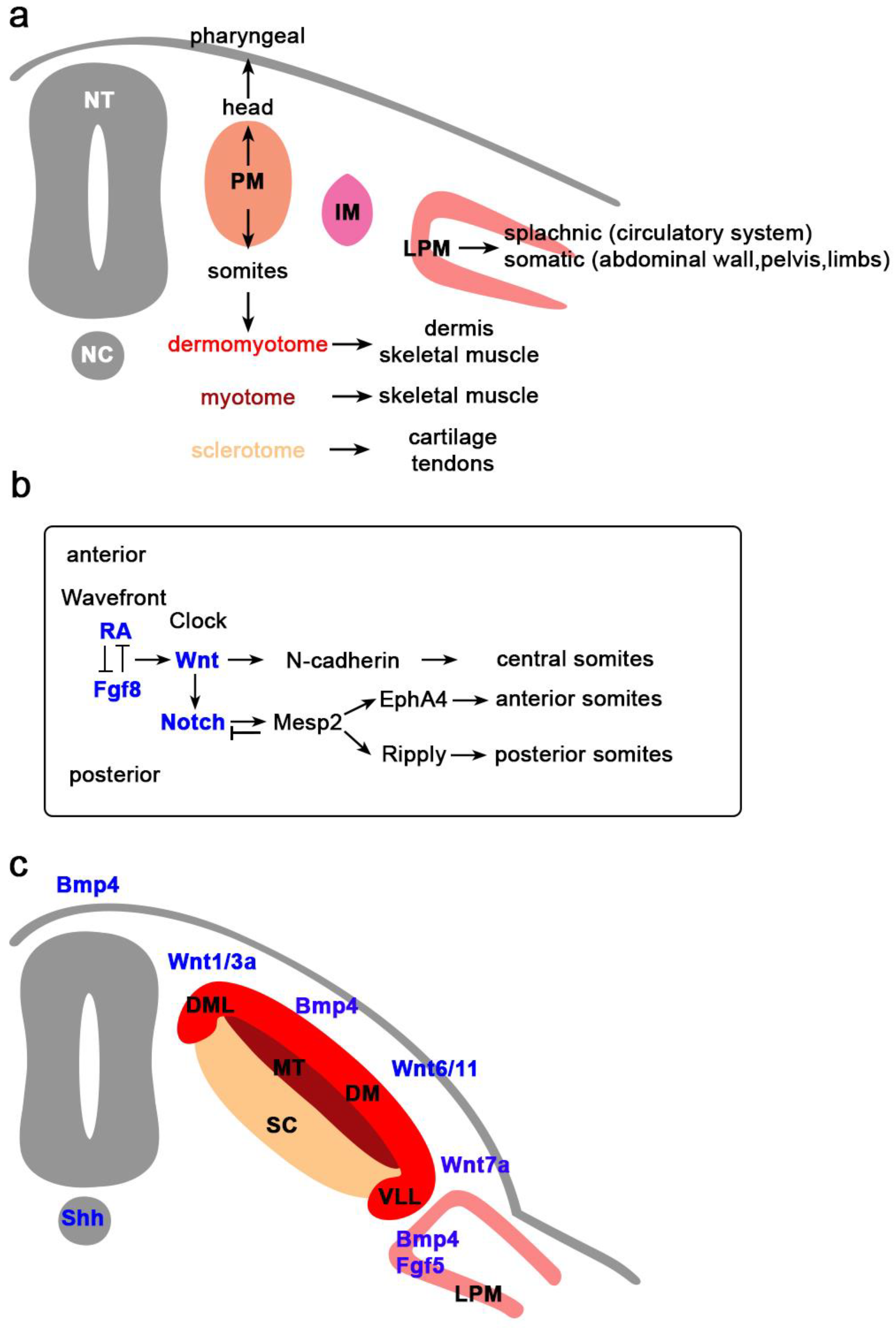{kind=link}
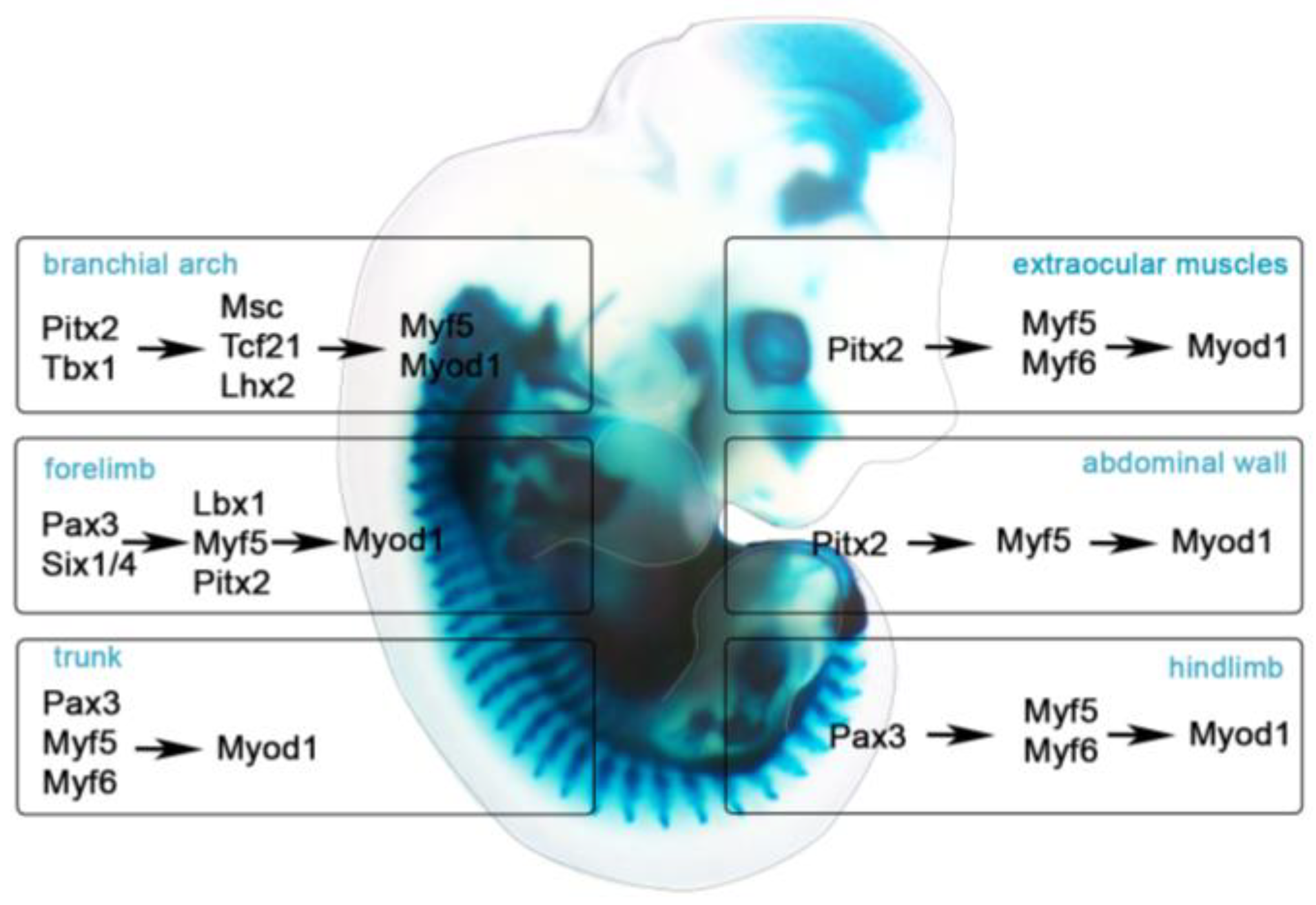{kind=link}
Table 1.
Signaling molecules during myogenesis.
| Gene | Anatomical Location | Model | Function | Reference |
|---|---|---|---|---|
| Bone Morphogenetic Factor (BMP) | ||||
| Bmp2 | murine fibroblasts | prevention of myogenesis | [21,22] | |
| murine myoblasts | inhibition of MyoG expression | [23,24] | ||
| fish | delayed muscle differentiation | [25] | ||
| Bmp4 | human fetal myoblasts | regulation of proliferation | [26] | |
| chick | induction of embryonic and fetal myogenesis | [27] | ||
| frog | induction of ventral mesoderm | [28] | ||
| chick | inhibition of Myod and Myf5 | [29] | ||
| axolotl | somite development | [30] | ||
| colloid-like 1 BMP4 regulator | anterior somites | chick | neck muscle formation | [31] |
| Noggin BMP4 antagonist | Neural Tube; Notochord | mouse | differentiation | [32,33] |
| chick | growth inhibition | [34] | ||
| mouse | fetal myogenesis; migration of muscle progenitor cells | [35,36] | ||
| Bmp7 | dorsal Neural Tube | chick | muscle growth, apoptosis | [34] |
| Fibroblast Growth Factor (FGF) | ||||
| Fgf2 | Central Nervous System | fish | inhibition of muscle pioneer cells | [19,24,25] |
| Fgf4 | myotubes | mouse | mesoderm formation | [37] |
| chick | inhibition of terminal differentiation of limb | [38] | ||
| chick | induction of tendon-specific markers | [39] | ||
| chick | patterning during gastrulation | [40] | ||
| frog | activation of Myod | [41,42] | ||
| Fgf5 | Lateral Plate Mesoderm | mouse | formation of germ layers | [43] |
| chick | inhibition of myogenesis | [44] | ||
| Fgf8 | caudal end of embryo | fish | terminal muscle differentiation | [45,46] |
| chick | tendon formation | [39] | ||
| fish | somitic muscle formation | [47] | ||
| murine myoblasts | myotubes formation | [48] | ||
| fly | mesodermal cell migration | [49] | ||
| chick | mesodermal cell migration | [40] | ||
| chick; mouse | myoblast proliferation | [50] | ||
| Notch Pathway | ||||
| Notch | Ectoderm | mouse | terminal muscle differentiation | [32,33] |
| frog | inhibition of cell fusion; myotube formation | [51] | ||
| mouse | cell fate decisions | [52] | ||
| murine myoblasts | inhibition of muscle differentiation | [53] | ||
| rbp-j NOTCH ligand | trunk somites | frog | determination of dorsolateral and ventral mesoderm | [28] |
| axolotl | formation of the dermomyotome | [30] | ||
| myeloma cells | inhibition of differentiation | [54] | ||
| mouse | differentiation | [32,33] | ||
| murine myoblasts | inhibition of differentiation | [55] | ||
| Jagged-1 NOTCH ligand | trunk mesenchyme, splachnopleure | mouse | differentiation | [32,33] |
| murine myoblasts | inhibition of differentiation | [56,57] | ||
| Delta1 NOTCH ligand | Neural Crest cells | mouse | terminal differentiation | [32,33] |
| chick | inhibition of terminal differentiation | [58] | ||
| chick | terminal differentiation, inhibition of exiting the cell cycle | [59] | ||
| Mesp2 NOTCH ligand | Presomitic Mesoderm | mouse | somitic boundaries | [60] |
| mouse | cellular epithelialization | [61] | ||
| mouse | somitogenesis; rostro-caudal polarity | [62] | ||
| mouse | inhibition of NOTCH targets | [63] | ||
| Ephrin Ligands | ||||
| EphA4 | Presomitic Mesoderm | mouse | somitogenesis | [61] |
| chick | somitogenesis | [64] | ||
| ephinb2 | somites | chick | somitogenesis, expression of N-cadherin | [64] |
| Ripply | Presomitic Mesoderm | mouse | rostro-caudal polarity | [62] |
| mouse | Mesp2 expression | [65] | ||
| neurotrophin 3 | Neural Tube | chick | Pax3 expression induction, somitic myogenesis regulation | [66] |
| chick | dermatome dissociation; epithelial-mesenchymal transition | [67] | ||
| Retinoic Acid (RA) | ||||
| Ra | cranial end of the embryo | mouse | Myf5 expression | [68] |
| fish | somitogenesis; fast muscle differentiation | [69] | ||
| murine myoblasts | inhibition of myoblast proliferation; differentiation | [69] | ||
| mouse | somitogenesis | [70] | ||
| Sonic Hedgehog (SHH) | ||||
| Shh | Neural Tube, Notochord | chick | somitogenesis | [66] |
| fish | formation of slow muscle precursor cells | [71] | ||
| chick | induction of myogenesis | [72] | ||
| fish | inhibition of pax3/7; activation of myf5 and myod | [47] | ||
| chick | induction of pax3 and myod1 | [73] | ||
| chick | activation of slow MyHC | [74] | ||
| mouse | epaxial and hypaxial myogenesis | [75] | ||
| Wnt | ||||
| Wnt1 | Neural Tube | chick | pax3 activation; somitogenesis | [66] |
| murine myoblasts | formation of slow fiber types promotion | [76] | ||
| chick | formation of paraxial mesoderm | [72] | ||
| mouse | Formation of medial and dorsal portion of somites | [77] | ||
| mouse | Myf5 activation | [15] | ||
| Wnt3a | Neural Tube, Apical Ectodermal Ridge (AER) | murine myoblasts | Bmp4 activation; formation of slow fibers | [76] |
| mouse | Formation of dorsomedial part of somites | [77] | ||
| murine myoblasts | myotube formation | [78] | ||
| human embryonic stem cells | myogenic commitment | [79] | ||
| murine myoblasts | myotube formation | [80] | ||
| pluripotent stem cells | cardiomyocytes proliferation | [81] | ||
| Wnt4 | Neural Tube | chickmurine myoblasts | activation of pax7 and myod1; formation of fast myofibers | [82] |
| mouse embryos | activation of Myf5 and Myod1 | [15] | ||
| murine myoblasts | myotube formation | [78] | ||
| murine satellite cells and myoblasts | activation of myogenesis | [83] | ||
| Wnt6 | Paraxial Ectoderm | mouse | activation of Myf5 and Myod1 in paraxial mesoderm | [15] |
| chick | activation of Pax3, Paraxis, Myf5, Myog, Desmin and MyHC | [84] | ||
| Wnt7a | Dorsal Ectoderm | mouse | activation of Myod1 | [15] |
| Wnt11 | somites | chick | formation of fast myofibers | [85] |
| chick | elongation of myocytes | [86] | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chang, C.-N.; Kioussi, C. Location, Location, Location: Signals in Muscle Specification. J. Dev. Biol. 2018, 6, 11. https://doi.org/10.3390/jdb6020011
AMA Style
Chang C-N, Kioussi C. Location, Location, Location: Signals in Muscle Specification. Journal of Developmental Biology. 2018; 6(2):11. https://doi.org/10.3390/jdb6020011
Chicago/Turabian StyleChang, Chih-Ning, and Chrissa Kioussi. 2018. "Location, Location, Location: Signals in Muscle Specification" Journal of Developmental Biology 6, no. 2: 11. https://doi.org/10.3390/jdb6020011
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.