LHX2 Mediates the FGF-to-SHH Regulatory Loop during Limb Development

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. FGF Bead Implants, WNT5a Cell Implants, and Harvesting Embryos
2.2. RNA-Seq Analysis
2.3. DNA Microarray Analysis
2.4. Gene Ontology
2.5. Gene Expression Analysis via Whole-Mount In Situ Hybridization (WMISH)
2.6. RT-qPCR Validation of Transcriptome Data and Quantitation of LHX2 Overexpression/Knockdown Analysis
2.7. Comparative Transcriptome Analysis
2.8. Gene Overexpression and Knockdown via Electroporation
Controlling for Off-Target Effects Using Morpholinos
3. Results
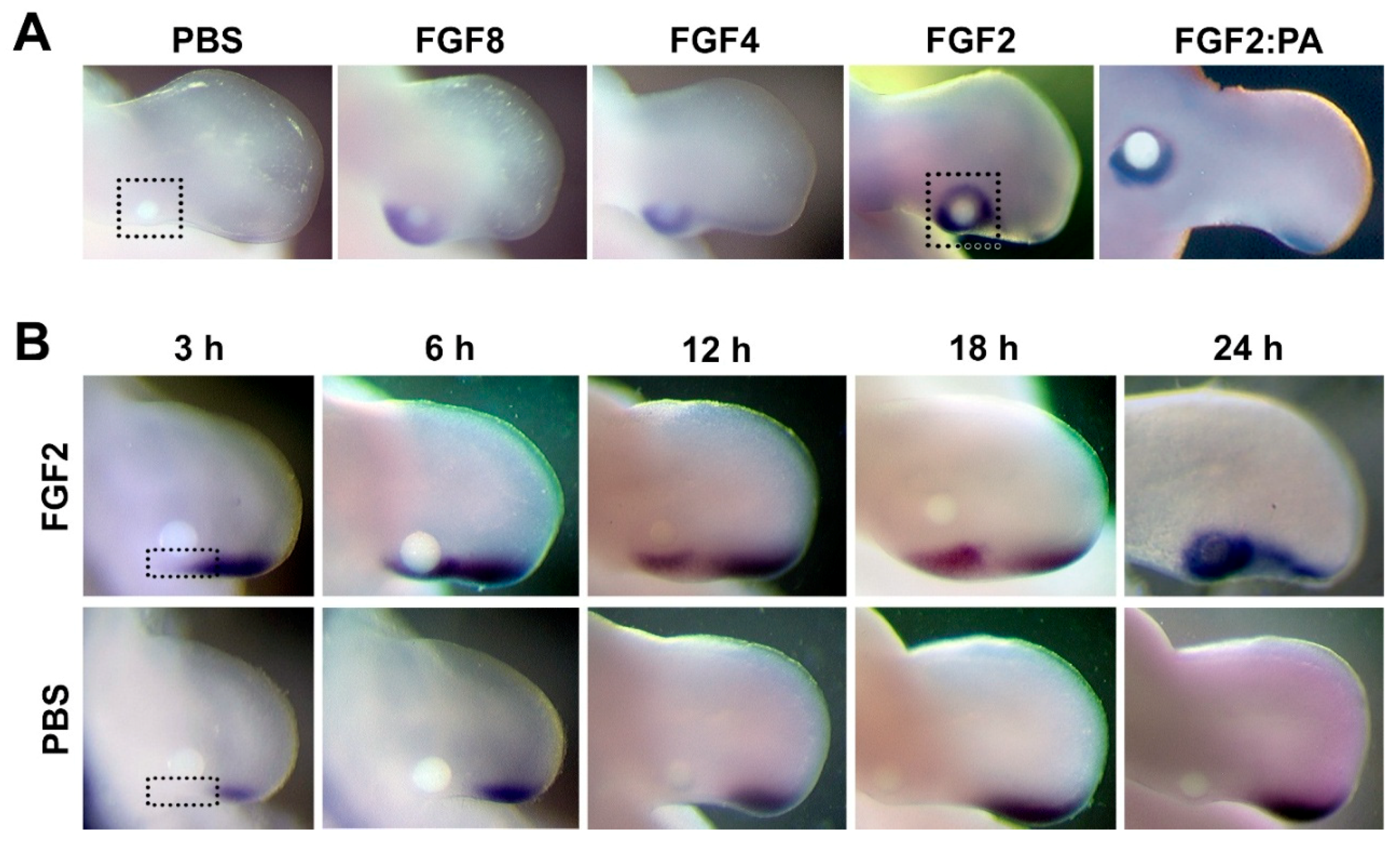
3.1. FGF Can Maintain and Induce SHH Expression in the Posterior Limb Bud
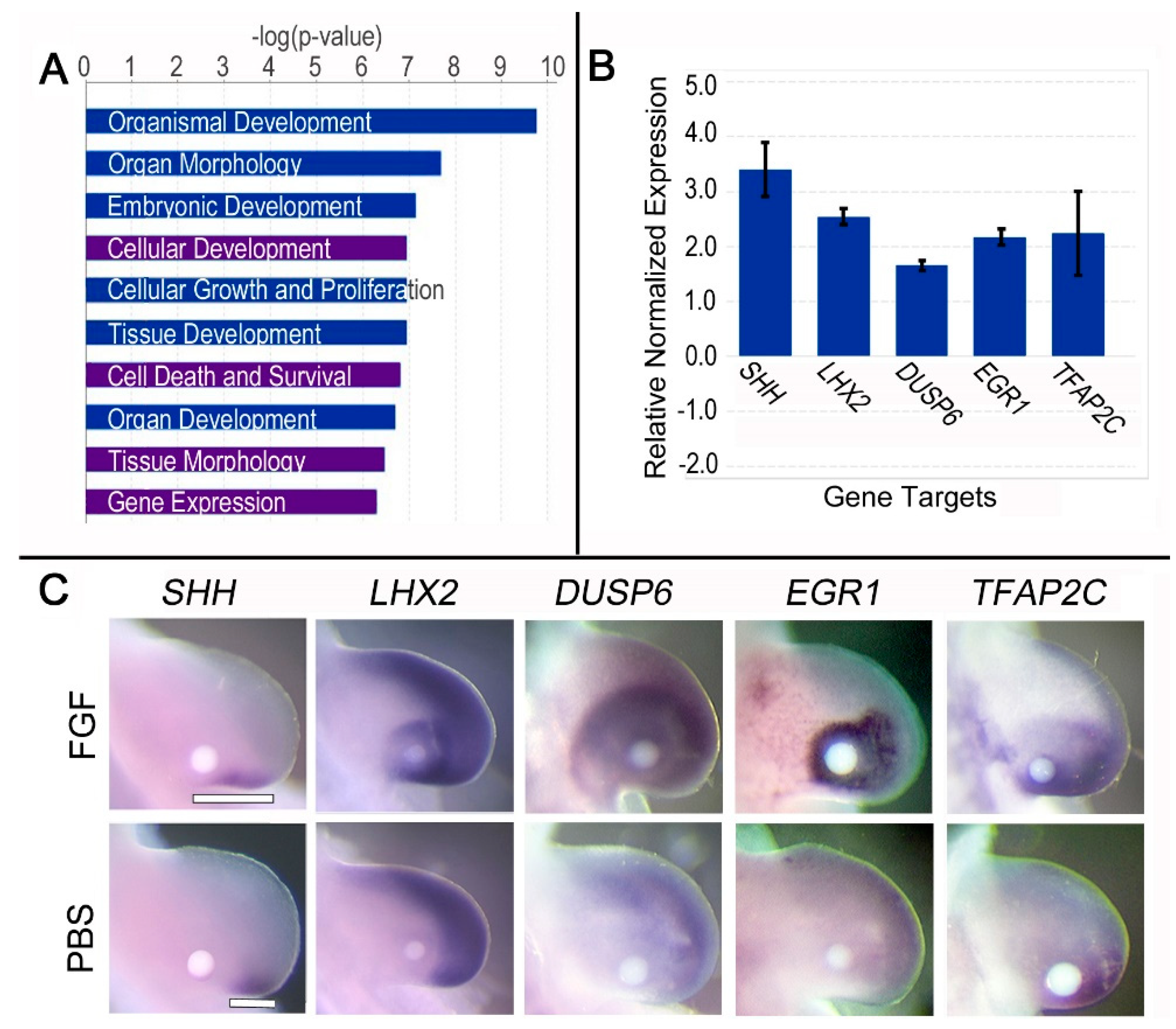
3.2. Brief (3 h) FGF Exposure Affects Biological Processes Associated with Its Role in Gene Expression
3.3. Brief (3 h) FGF Exposure Regulates Genes Involved in FGF Feedback Inhibition, Distalization, and SHH Expression
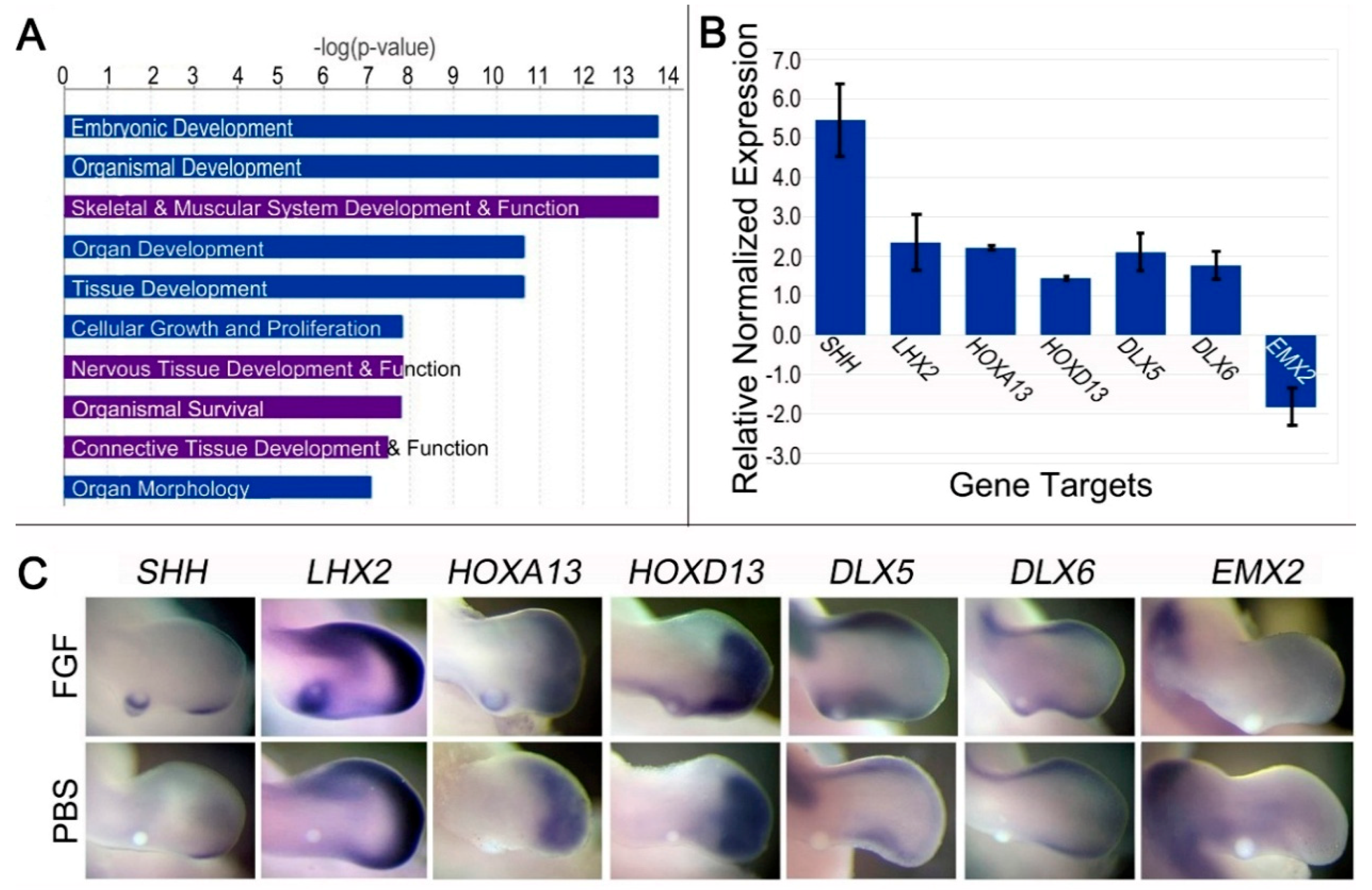
3.4. Prolonged (24 h) FGF Exposure Affects Cell Processes Related to Organ and Organismal Development
3.5. Prolonged FGF (24 h) Regulates Genes Associated with Dedifferentiation, Distalization, and SHH Regulation
3.6. Common Pathways Accentuated by Comparative Transcriptome Analysis
3.6.1. Common Targets Involved in Wnt Signaling
3.6.2. Common Targets Associated with SHH Expression
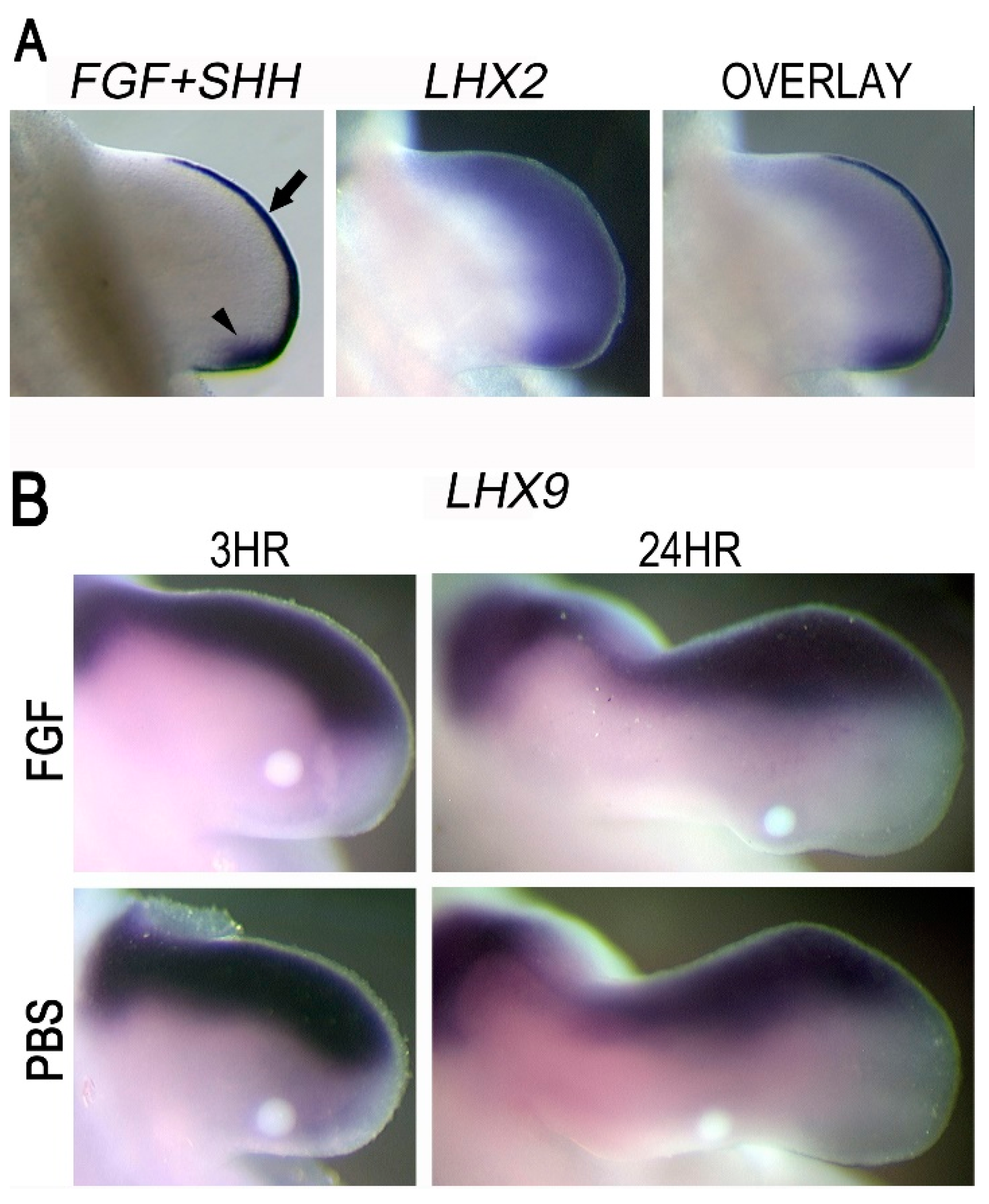
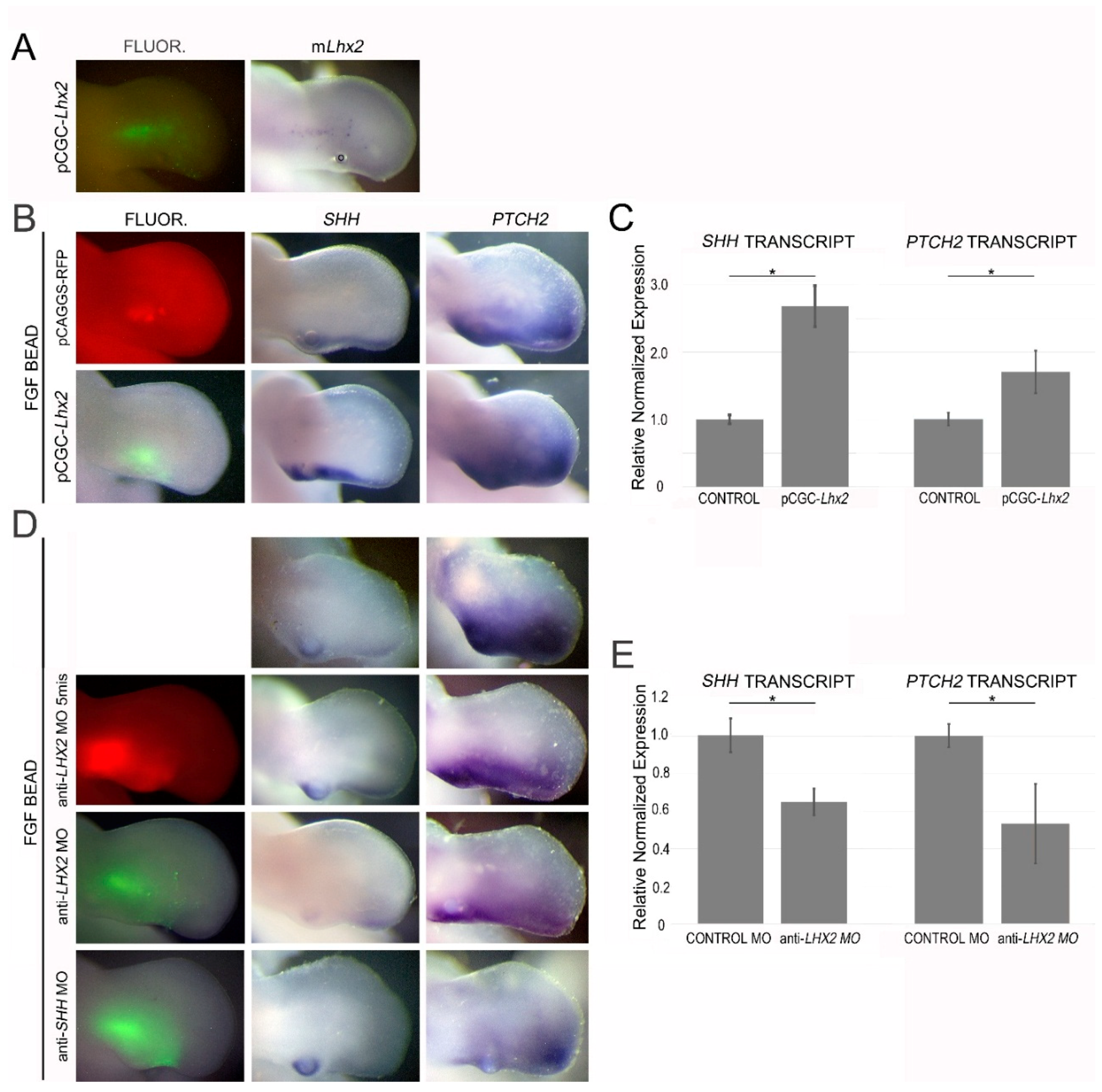
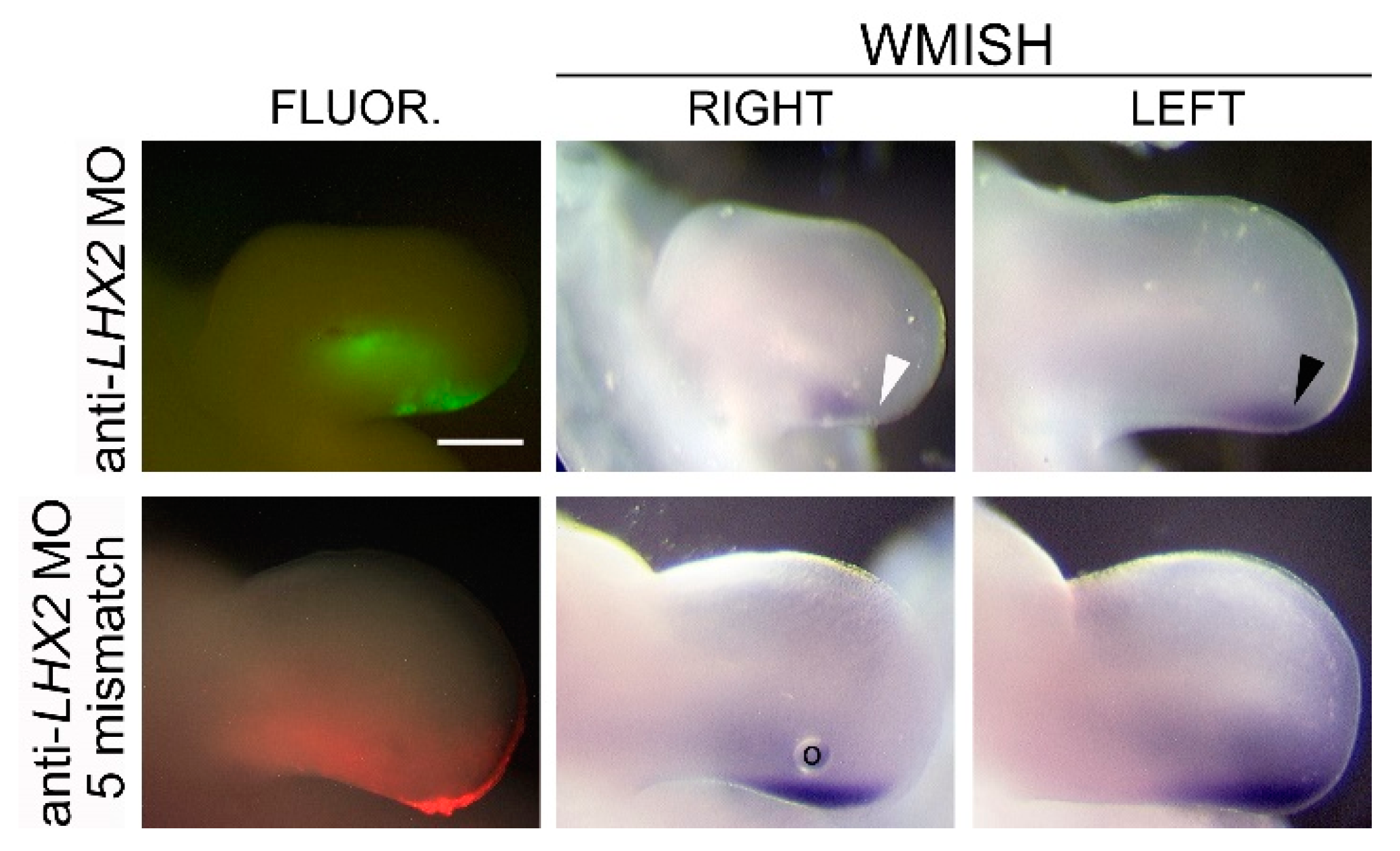
3.7. LHX2 as an Intermediate in FGF-Regulated SHH Expression
4. Discussion
4.1. FGF Regulates Genes that Support SHH Expression and Signaling/Function
4.2. FGF Regulates Factors that Localize SHH Expression
4.3. LHX2, but Not LHX9, Regulates FGF-Mediated SHH Expression during Chicken Limb Development
4.4. LHX2 as a Competency Factor for SHH Expression in the Limb
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Delgado, I.; Torres, M. Coordination of limb development by crosstalk among axial patterning pathways. Dev. Biol. 2017, 429, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Laufer, E.; Nelson, C.E.; Johnson, R.L.; Morgan, B.A.; Tabin, C. Sonic hedgehog and fgf-4 act through a signaling cascade and feedback loop to integrate growth and patterning of the developing limb bud. Cell 1994, 79, 993–1003. [Google Scholar] [CrossRef]
- Niswander, L.; Jeffrey, S.; Martin, G.R.; Tickle, C. A positive feedback loop coordinates growth and patterning in the vertebrate limb. Nature 1994, 371, 609–612. [Google Scholar] [CrossRef] [PubMed]
- Zeller, R.; Haramis, A.G.; Zuniga, A.; McGuigan, C.; Dono, R.; Davidson, G.; Chabanis, S.; Gibson, T. Formin defines a large family of morphoregulatory genes and functions in establishment of the polarising region. Cell Tissue Res. 1999, 296, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Capdevila, J.; Tsukui, T.; Rodriquez Esteban, C.; Zappavigna, V.; Izpisua Belmonte, J.C. Control of vertebrate limb outgrowth by the proximal factor meis2 and distal antagonism of bmps by gremlin. Mol. Cell 1999, 4, 839–849. [Google Scholar] [CrossRef]
- Zuniga, A.; Haramis, A.P.; McMahon, A.P.; Zeller, R. Signal relay by bmp antagonism controls the shh/fgf4 feedback loop in vertebrate limb buds. Nature 1999, 401, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Ros, M.A.; Dahn, R.D.; Fernandez-Teran, M.; Rashka, K.; Caruccio, N.C.; Hasso, S.M.; Bitgood, J.J.; Lancman, J.J.; Fallon, J.F. The chick oligozeugodactyly (ozd) mutant lacks sonic hedgehog function in the limb. Development 2003, 130, 527–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, C.; Litingtung, Y.; Harris, M.P.; Simandl, B.K.; Li, Y.; Beachy, P.A.; Fallon, J.F. Manifestation of the limb prepattern: Limb development in the absence of sonic hedgehog function. Dev. Biol. 2001, 236, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Harfe, B.D.; Scherz, P.J.; Nissim, S.; Tian, H.; McMahon, A.P.; Tabin, C.J. Evidence for an expansion-based temporal shh gradient in specifying vertebrate digit identities. Cell 2004, 118, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Ingham, P.W.; Placzek, M. Orchestrating ontogenesis: Variations on a theme by sonic hedgehog. Nat. Rev. Genet. 2006, 7, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Fallon, J.F.; Lopez, A.; Ros, M.A.; Savage, M.P.; Olwin, B.B.; Simandl, B.K. Fgf-2: Apical ectodermal ridge growth signal for chick limb development. Science 1994, 264, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Anderson, R.; Reginelli, A.D.; Muneoka, K. Fgf-2 influences cell movements and gene expression during limb development. J. Exp. Zool. 1996, 274, 234–247. [Google Scholar] [CrossRef]
- Niswander, L.; Tickle, C.; Vogel, A.; Booth, I.; Martin, G.R. Fgf-4 replaces the apical ectodermal ridge and directs outgrowth and patterning of the limb. Cell 1993, 75, 579–587. [Google Scholar] [CrossRef]
- Vogel, A.; Tickle, C. Fgf-4 maintains polarizing activity of posterior limb bud cells in vivo and in vitro. Development 1993, 119, 199–206. [Google Scholar] [PubMed]
- Crossley, P.H.; Minowada, G.; MacArthur, C.A.; Martin, G.R. Roles for fgf8 in the induction, initiation, and maintenance of chick limb development. Cell 1996, 84, 127–136. [Google Scholar] [CrossRef]
- Vogel, A.; Rodriguez, C.; Izpisua-Belmonte, J.C. Involvement of fgf-8 in initiation, outgrowth and patterning of the vertebrate limb. Development 1996, 122, 1737–1750. [Google Scholar] [PubMed]
- Savage, M.P.; Hart, C.E.; Riley, B.B.; Sasse, J.; Olwin, B.B.; Fallon, J.F. Distribution of fgf-2 suggests it has a role in chick limb bud growth. Dev. Dyn. 1993, 198, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Becic, T.; Kero, D.; Vukojevic, K.; Mardesic, S.; Saraga-Babic, M. Growth factors fgf8 and fgf2 and their receptor fgfr1, transcriptional factors msx-1 and msx-2, and apoptotic factors p19 and rip5 participate in the early human limb development. Acta Histochem. 2018, 120, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Harduf, H.; Halperin, E.; Reshef, R.; Ron, D. Sef is synexpressed with fgfs during chick embryogenesis and its expression is differentially regulated by fgfs in the developing limb. Dev. Dyn. 2005, 233, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Verheyden, J.M.; Lewandoski, M.; Deng, C.; Harfe, B.D.; Sun, X. Conditional inactivation of fgfr1 in mouse defines its role in limb bud establishment, outgrowth and digit patterning. Development 2005, 132, 4235–4245. [Google Scholar] [CrossRef] [PubMed]
- Sheeba, C.J.; Andrade, R.P.; Duprez, D.; Palmeirim, I. Comprehensive analysis of fibroblast growth factor receptor expression patterns during chick forelimb development. Int. J. Dev. Biol. 2010, 54, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Oberg, K.C.; Pira, C.U.; Revelli, J.P.; Ratz, B.; Aguilar-Cordova, E.; Eichele, G. Efficient ectopic gene expression targeting chick mesoderm. Dev. Dyn. 2002, 224, 291–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feenstra, J.M.; Kanaya, K.; Pira, C.U.; Hoffman, S.E.; Eppey, R.J.; Oberg, K.C. Detection of genes regulated by lmx1b during limb dorsalization. Dev. Growth Differ. 2012, 54, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. Ncbi geo: Archive for functional genomics data sets--update. Nucl. Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Szendro, P.I.; Prokscha, A.; Schwartz, R.J.; Eichele, G. Evidence for a role of smad6 in chick cardiac development. Dev. Biol. 1999, 215, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, L.; Sarkar, A.; Shetty, A.S.; Muralidharan, B.; Padmanabhan, H.; Piper, M.; Monuki, E.S.; Bach, I.; Gronostajski, R.M.; Richards, L.J.; et al. Transcription factor lhx2 is necessary and sufficient to suppress astrogliogenesis and promote neurogenesis in the developing hippocampus. Proc. Natl. Acad. Sci. USA 2011, 108, E265–E274. [Google Scholar] [CrossRef] [PubMed]
- Moulton, J.D.; Yan, Y.L. Using Morpholinos to control gene expression. Curr. Protoc. Mol. Biol. 2008, 83, 26.8.1–26.8.29. [Google Scholar]
- Eisen, J.S.; Smith, J.C. Controlling morpholino experiments: Don't stop making antisense. Development 2008, 135, 1735–1743. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Esteban, C.; Schwabe, J.W.; Pena, J.D.; Rincon-Limas, D.E.; Magallon, J.; Botas, J.; Izpisua Belmonte, J.C. Lhx2, a vertebrate homologue of apterous, regulates vertebrate limb outgrowth. Development 1998, 125, 3925–3934. [Google Scholar] [PubMed]
- Tzchori, I.; Day, T.F.; Carolan, P.J.; Zhao, Y.; Wassif, C.A.; Li, L.; Lewandoski, M.; Gorivodsky, M.; Love, P.E.; Porter, F.D.; et al. Lim homeobox transcription factors integrate signaling events that control three-dimensional limb patterning and growth. Development 2009, 136, 1375–1385. [Google Scholar] [CrossRef] [PubMed]
- Bastida, M.F.; Sheth, R.; Ros, M.A. A bmp-shh negative-feedback loop restricts shh expression during limb development. Development 2009, 136, 3779–3789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norrie, J.L.; Lewandowski, J.P.; Bouldin, C.M.; Amarnath, S.; Li, Q.; Vokes, M.S.; Ehrlich, L.I.R.; Harfe, B.D.; Vokes, S.A. Dynamics of bmp signaling in limb bud mesenchyme and polydactyly. Dev. Biol. 2014, 393, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Capellini, T.D.; Di Giacomo, G.; Salsi, V.; Brendolan, A.; Ferretti, E.; Srivastava, D.; Zappavigna, V.; Selleri, L. Pbx1/pbx2 requirement for distal limb patterning is mediated by the hierarchical control of hox gene spatial distribution and shh expression. Development 2006, 133, 2263–2273. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Wellik, D.M. Axial hox9 activity establishes the posterior field in the developing forelimb. Proc. Natl. Acad. Sci. USA 2011, 108, 4888–4891. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, H.; Saito, D.; Abe, G.; Yokoyama, H.; Suzuki, T.; Tamura, K. Upstream regulation for initiation of restricted shh expression in the chick limb bud. Dev. Dyn. 2017, 246, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Osterwalder, M.; Speziale, D.; Shoukry, M.; Mohan, R.; Ivanek, R.; Kohler, M.; Beisel, C.; Wen, X.; Scales, S.J.; Christoffels, V.M.; et al. Hand2 targets define a network of transcriptional regulators that compartmentalize the early limb bud mesenchyme. Dev. Cell 2014, 31, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Lettice, L.; Heaney, S.; Hill, R. 2 Preaxial polydactyly in human and mouse: Regulatory anomalies in digit patterning. J. Anat. 2002, 201, 417. [Google Scholar] [PubMed]
- Church, V.L.; Francis-West, P. Wnt signalling during limb development. Int. J. Dev. Biol. 2002, 46, 927–936. [Google Scholar] [PubMed]
- Geetha-Loganathan, P.; Nimmagadda, S.; Scaal, M. Wnt signaling in limb organogenesis. Organogenesis 2008, 4, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, J.; McGlinn, E.; Huang, P.; Tabin, C.J.; McMahon, A.P. Fgf-dependent etv4/5 activity is required for posterior restriction of sonic hedgehog and promoting outgrowth of the vertebrate limb. Dev. Cell 2009, 16, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Verheyden, J.M.; Hassell, J.A.; Sun, X. Fgf-regulated etv genes are essential for repressing shh expression in mouse limb buds. Dev. Cell 2009, 16, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Lettice, L.A.; Williamson, I.; Wiltshire, J.H.; Peluso, S.; Devenney, P.S.; Hill, A.E.; Essafi, A.; Hagman, J.; Mort, R.; Grimes, G.; et al. Opposing functions of the ets factor family define shh spatial expression in limb buds and underlie polydactyly. Dev. Cell 2012, 22, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Niswender, K.D.; Ji, Q.; van der Meer, R.; Keeney, D.; Magnuson, M.A.; Wisdom, R. Polydactyly and ectopic zpa formation in alx-4 mutant mice. Development 1997, 124, 3999–4008. [Google Scholar] [PubMed]
- Shimomura, Y.; Agalliu, D.; Vonica, A.; Luria, V.; Wajid, M.; Baumer, A.; Belli, S.; Petukhova, L.; Schinzel, A.; Brivanlou, A.H.; et al. Apcdd1 is a novel wnt inhibitor mutated in hereditary hypotrichosis simplex. Nature 2010, 464, 1043–1047. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, J.; Fagotto, F.; van der Velde, H.; van Schie, M.; Noordermeer, J.; Fornerod, M. Ranbp3 enhances nuclear export of active (beta)-catenin independently of crm1. J. Cell Biol. 2005, 171, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, S.; Rosin, J.M.; Ambasta, A.; Hui, K.; Shaneman, V.; Crowder, R.; Vickerman, L.; Cobb, J. A conditional allele of rspo3 reveals redundant function of r-spondins during mouse limb development. Genesis 2012, 50, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, P.; Liu, Y.; Lv, L.; Zhang, X.; Liu, H.; Zhou, Y. Rspo3-lgr4 regulates osteogenic differentiation of human adipose-derived stem cells via erk/fgf signalling. Sci. Rep. 2017, 7, 42841. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Wang, Q.; Guo, K.; Qin, W.; Liao, W.; Wang, S.; Ding, Y.; Lin, J. Tusc3 promotes colorectal cancer progression and epithelial-mesenchymal transition (emt) through wnt/beta-catenin and mapk signalling. J. Pathol. 2016, 239, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Ten Berge, D.; Brugmann, S.A.; Helms, J.A.; Nusse, R. Wnt and fgf signals interact to coordinate growth with cell fate specification during limb development. Development 2008, 135, 3247–3257. [Google Scholar] [CrossRef] [PubMed]
- Probst, S.; Kraemer, C.; Demougin, P.; Sheth, R.; Martin, G.R.; Shiratori, H.; Hamada, H.; Iber, D.; Zeller, R.; Zuniga, A. Shh propagates distal limb bud development by enhancing cyp26b1-mediated retinoic acid clearance via aer-fgf signalling. Development 2011, 138, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Guerrero-Netro, H.; Estienne, A.; Cao, B.; Price, C.A. Regulation and action of early growth response 1 in bovine granulosa cells. Reproduction 2017, 154, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Lejard, V.; Blais, F.; Guerquin, M.J.; Bonnet, A.; Bonnin, M.A.; Havis, E.; Malbouyres, M.; Bidaud, C.B.; Maro, G.; Gilardi-Hebenstreit, P.; et al. Egr1 and egr2 involvement in vertebrate tendon differentiation. J. Biol. Chem. 2011, 286, 5855–5867. [Google Scholar] [CrossRef] [PubMed]
- Sakakini, N.; Turchi, L.; Bergon, A.; Holota, H.; Rekima, S.; Lopez, F.; Paquis, P.; Almairac, F.; Fontaine, D.; Baeza-Kallee, N.; et al. A positive feed-forward loop associating egr1 and pdgfa promotes proliferation and self-renewal in glioblastoma stem cells. J. Biol. Chem. 2016, 291, 10684–10699. [Google Scholar] [CrossRef] [PubMed]
- Law, L.Y.; Lin, J.S.; Becker, D.L.; Green, C.R. Knockdown of connexin43-mediated regulation of the zone of polarizing activity in the developing chick limb leads to digit truncation. Dev. Growth Differ. 2002, 44, 537–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrowolski, R.; Hertig, G.; Lechner, H.; Worsdorfer, P.; Wulf, V.; Dicke, N.; Eckert, D.; Bauer, R.; Schorle, H.; Willecke, K. Loss of connexin43-mediated gap junctional coupling in the mesenchyme of limb buds leads to altered expression of morphogens in mice. Hum. Mol. Genet. 2009, 18, 2899–2911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuxman Bass, J.I.; Sahni, N.; Shrestha, S.; Garcia-Gonzalez, A.; Mori, A.; Bhat, N.; Yi, S.; Hill, D.E.; Vidal, M.; Walhout, A.J.M. Human gene-centered transcription factor networks for enhancers and disease variants. Cell 2015, 161, 661–673. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Seo, S.; Bugge, K.; Stone, E.M.; Sheffield, V.C. Bbs proteins interact genetically with the ift pathway to influence shh-related phenotypes. Hum. Mol. Genet. 2012, 21, 1945–1953. [Google Scholar] [CrossRef] [PubMed]
- Haycraft, C.J.; Banizs, B.; Aydin-Son, Y.; Zhang, Q.; Michaud, E.J.; Yoder, B.K. Gli2 and gli3 localize to cilia and require the intraflagellar transport protein polaris for processing and function. PLoS Genet. 2005, 1, e53. [Google Scholar] [CrossRef] [PubMed]
- Sheeba, C.J.; Andrade, R.P.; Palmeirim, I. Getting a handle on embryo limb development: Molecular interactions driving limb outgrowth and patterning. Semin. Cell Dev. Biol. 2016, 49, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Manouvrier-Hanu, S.; Moerman, A.; Lefevre, J. Bardet-biedl syndrome with preaxial polydactyly. Am. J. Med. Genet. 1999, 84, 75. [Google Scholar] [CrossRef]
- M'Hamdi, O.; Ouertani, I.; Chaabouni-Bouhamed, H. Update on the genetics of bardet-biedl syndrome. Mol. Syndromol. 2014, 5, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Tayeh, M.K.; Yen, H.J.; Beck, J.S.; Searby, C.C.; Westfall, T.A.; Griesbach, H.; Sheffield, V.C.; Slusarski, D.C. Genetic interaction between bardet-biedl syndrome genes and implications for limb patterning. Hum. Mol. Genet. 2008, 17, 1956–1967. [Google Scholar] [CrossRef] [PubMed]
- Kraus, P.; Fraidenraich, D.; Loomis, C.A. Some distal limb structures develop in mice lacking sonic hedgehog signaling. Mech. Dev. 2001, 100, 45–58. [Google Scholar] [CrossRef]
- Smyth, J.R., Jr.; Sreekumar, G.P.; Coyle, C.A.; Bitgood, J.J. A new recessive ametapodia mutation in the chicken (gallus domesticus). J. Hered. 2000, 91, 340–342. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, J.; Lettice, L.; Hecksher-Sorensen, J.; Fox, M.; Hill, R.; Krumlauf, R. Identification of sonic hedgehog as a candidate gene responsible for the polydactylous mouse mutant sasquatch. Curr. Biol. 1999, 9, 97–100. [Google Scholar] [CrossRef]
- Roberson, M.S.; Schoderbek, W.E.; Tremml, G.; Maurer, R.A. Activation of the glycoprotein hormone alpha-subunit promoter by a lim-homeodomain transcription factor. Mol. Cell Biol. 1994, 14, 2985–2993. [Google Scholar] [CrossRef] [PubMed]
- Kvon, E.Z.; Kamneva, O.K.; Melo, U.S.; Barozzi, I.; Osterwalder, M.; Mannion, B.J.; Tissieres, V.; Pickle, C.S.; Plajzer-Frick, I.; Lee, E.A.; et al. Progressive loss of function in a limb enhancer during snake evolution. Cell 2016, 167, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Bertuzzi, S.; Porter, F.D.; Pitts, A.; Kumar, M.; Agulnick, A.; Wassif, C.; Westphal, H. Characterization of lhx9, a novel lim/homeobox gene expressed by the pioneer neurons in the mouse cerebral cortex. Mech. Dev. 1999, 81, 193–198. [Google Scholar] [CrossRef]
- Peukert, D.; Weber, S.; Lumsden, A.; Scholpp, S. Lhx2 and lhx9 determine neuronal differentiation and compartition in the caudal forebrain by regulating wnt signaling. PLoS Biol. 2011, 9, e1001218. [Google Scholar] [CrossRef] [PubMed]
- Nohno, T.; Kawakami, Y.; Wada, N.; Ishikawa, T.; Ohuchi, H.; Noji, S. Differential expression of the two closely related lim-class homeobox genes lh-2a and lh-2b during limb development. Biochem. Biophys. Res. Commun. 1997, 238, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wilson, M.J. Lhx9 gene expression during early limb development in mice requires the fgf signalling pathway. Gene Expr. Patterns 2015, 19, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Tucker, A.S.; Al Khamis, A.; Ferguson, C.A.; Bach, I.; Rosenfeld, M.G.; Sharpe, P.T. Conserved regulation of mesenchymal gene expression by fgf-8 in face and limb development. Development 1999, 126, 221–228. [Google Scholar] [PubMed]
- Jurata, L.W.; Pfaff, S.L.; Gill, G.N. The nuclear lim domain interactor nli mediates homo- and heterodimerization of lim domain transcription factors. J. Biol. Chem. 1998, 273, 3152–3157. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.C.; Revelli, J.P.; Goering, L.; Thaller, C.; Eichele, G. Retinoid signaling is required for the establishment of a zpa and for the expression of hoxb-8, a mediator of zpa formation. Development 1997, 124, 1643–1651. [Google Scholar] [PubMed]
- Charite, J.; McFadden, D.G.; Olson, E.N. The bhlh transcription factor dhand controls sonic hedgehog expression and establishment of the zone of polarizing activity during limb development. Development 2000, 127, 2461–2470. [Google Scholar] [PubMed]
- Fernandez-Teran, M.; Piedra, M.E.; Kathiriya, I.S.; Srivastava, D.; Rodriguez-Rey, J.C.; Ros, M.A. Role of dhand in the anterior-posterior polarization of the limb bud: Implications for the sonic hedgehog pathway. Development 2000, 127, 2133–2142. [Google Scholar] [PubMed]
- Chen, Y.; Knezevic, V.; Ervin, V.; Hutson, R.; Ward, Y.; Mackem, S. Direct interaction with hoxd proteins reverses gli3-repressor function to promote digit formation downstream of shh. Development 2004, 131, 2339–2347. [Google Scholar] [CrossRef] [PubMed]
- Zhulyn, O.; Li, D.; Deimling, S.; Vakili, N.A.; Mo, R.; Puviindran, V.; Chen, M.H.; Chuang, P.T.; Hopyan, S.; Hui, C.C. A switch from low to high shh activity regulates establishment of limb progenitors and signaling centers. Dev. Cell 2014, 29, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, A.A.; Paparidis, Z.; Malik, S.; Bangs, F.; Schmidt, A.; Koch, S.; Lopez-Rios, J.; Grzeschik, K.H. Human intronic enhancers control distinct sub-domains of gli3 expression during mouse cns and limb development. BMC Dev. Biol. 2010, 10, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Sui, P.; Dong, A.; Hassell, J.; Cserjesi, P.; Chen, Y.T.; Behringer, R.R.; Sun, X. Preaxial polydactyly: Interactions among etv, twist1 and hand2 control anterior-posterior patterning of the limb. Development 2010, 137, 3417–3426. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S.; Furukawa, S.; Kitada, S.; Mori, M.; Saito, T.; Kawakami, K.; Belmonte, J.C.I.; Kawakami, Y.; Ito, Y.; Sato, T.; et al. Analysis of transcription factors expressed at the anterior mouse limb bud. PLoS ONE 2017, 12, e0175673. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watson, B.A.; Feenstra, J.M.; Van Arsdale, J.M.; Rai-Bhatti, K.S.; Kim, D.J.H.; Coggins, A.S.; Mattison, G.L.; Yoo, S.; Steinman, E.D.; Pira, C.U.; et al. LHX2 Mediates the FGF-to-SHH Regulatory Loop during Limb Development. J. Dev. Biol. 2018, 6, 13. https://doi.org/10.3390/jdb6020013
Watson BA, Feenstra JM, Van Arsdale JM, Rai-Bhatti KS, Kim DJH, Coggins AS, Mattison GL, Yoo S, Steinman ED, Pira CU, et al. LHX2 Mediates the FGF-to-SHH Regulatory Loop during Limb Development. Journal of Developmental Biology. 2018; 6(2):13. https://doi.org/10.3390/jdb6020013
Chicago/Turabian StyleWatson, Billy A., Jennifer M. Feenstra, Jonathan M. Van Arsdale, Karndeep S. Rai-Bhatti, Diana J. H. Kim, Ashley S. Coggins, Gennaya L. Mattison, Stephen Yoo, Eric D. Steinman, Charmaine U. Pira, and et al. 2018. "LHX2 Mediates the FGF-to-SHH Regulatory Loop during Limb Development" Journal of Developmental Biology 6, no. 2: 13. https://doi.org/10.3390/jdb6020013