Tungsten Toxicity in Plants

Department of Botany, School of Biology, Aristotle University of Thessaloniki, 541 24 Thessaloniki, Greece
*
Author to whom correspondence should be addressed.
Plants 2012, 1(2), 82-99; https://doi.org/10.3390/plants1020082
Submission received: 3 September 2012
/
Revised: 9 November 2012
/
Accepted: 13 November 2012
/
Published: 16 November 2012
(This article belongs to the Special Issue New Journal Plants - Feature Papers)
Abstract
:Tungsten (W) is a rare heavy metal, widely used in a range of industrial, military and household applications due to its unique physical properties. These activities inevitably have accounted for local W accumulation at high concentrations, raising concerns about its effects for living organisms. In plants, W has primarily been used as an inhibitor of the molybdoenzymes, since it antagonizes molybdenum (Mo) for the Mo-cofactor (MoCo) of these enzymes. However, recent advances indicate that, beyond Mo-enzyme inhibition, W has toxic attributes similar with those of other heavy metals. These include hindering of seedling growth, reduction of root and shoot biomass, ultrastructural malformations of cell components, aberration of cell cycle, disruption of the cytoskeleton and deregulation of gene expression related with programmed cell death (PCD). In this article, the recent available information on W toxicity in plants and plant cells is reviewed, and the knowledge gaps and the most pertinent research directions are outlined.
Abbreviations
| AO | aldehyde oxidase |
| ER | endoplasmic reticulum |
| mARC | mitochondrial amidoxime reducing component |
| MoCo | molybdenum cofactor |
| Mo-enzymes | molybdoenzymes |
| MT | microtubule |
| NR | nitrate reductase |
| PCD | programmed cell death |
| ROS | reactive oxygen species |
| SO | sulphite oxidase |
| UPR | unfolded protein response |
| W-PCD | tungsten-induced programmed cell death |
| XDH | xanthine dehydrogenase |
1. Introduction
Tungsten (W) is a rare transition element, belonging in the Group VI-B of the Periodic Table of elements, along with chromium, molybdenum (Mo) and seaborgium. Due to its unique physical and chemical properties, W is an extremely useful element having broad industrial, civil and military applications, ranging from daily household goods such as light bulbs and golf clubs, to electronics, hard metal tools, munitions and highly specialized components of advanced modern science and technology [1]. Although W is present in the soil in traces, its concentration can be very high locally due to anthropogenic activities, such as active and abandoned mining sites [2,3,4]. The use in agriculture of phosphate fertilizers that may contain W up to 100 mg kg−1 [5] and the replacement of lead with W in military applications and war practices, e.g., in projectiles, as an environmentally “benign” alternative, have increased surface soil W concentration [6,7] and the potential exposure of humans to W [8]. Post-consumer discharges also contribute to the flow of W in the environment. Although the recycling rates of the metal discharges reach 50%–70%, it has been pointed out that there still exists a possibility for sustainable use and/or better control in the environmental emissions of W [9].
Unlike many other metals, such as copper, cadmium, nickel, aluminium and mercury, which have been well documented as toxic and harmful to living organisms, the existing knowledge on the presumed toxicological profile of W is relatively limited and constitutes a subject of productive discussion within the scientific community [1,10]. Hence, W has been considered either as a non-toxic or non deleterious metal [11] or it has been regarded that it might be tumorous and leukemogenic in animal cells [12].
While in the former USSR investigations concerning the toxicological profile of W have began already by the 1950s and environmental regulations on W pollution were developed since the 1980s, research on W in USA and the European Union has started relatively recently [13]. Commence of research over W toxicology in the USA was encouraged due to cases of the leukemia clusters in Nevada State [14]. Especially the cluster in the city of Fallon, regarded as “one of the most unique …ever reported” [15], provoked extensive research in an attempt to find the preliminary cause. Despite the high levels of W, as well as of arsenic, JP-8 jet fuel and pesticides found in the area, no direct relationship between leukemia and W exposure was found, while no environmental exposure that could explain the cluster of childhood leukemia was ever discovered (among others see [16]). However, due to a lack of experimental data on which regulatory decisions could be established, the National Institute of Health classified W as a priority chemical for toxicological research [16]. In 2008, W was characterized as a substance of concern and an emerging pollutant [13]. A question then arises whether W could be considered as an environmental problem, boosting relevant research over recent years, mostly on microbes, animals and humans ([17,18,19,20,21,22,23,24]; and references therein).
In plants, the relevant information so far refers mainly to the molybdoenzyme (Mo-enzyme) research, where W is used as an inhibitor of Mo-enzymes (Section 2.2). However, recent reports indicate that, apart from its effects on the Mo-enzymes, W inhibits growth, deregulates cell division, disrupts cortical microtubule (MT) integrity and function, and induces programmed cell death (PCD) [25,26,27,28]. Consequently, W toxicity to plants should be analyzed under a broadened view, not solely as a Mo-enzyme inhibitor but also as a heavy metal with further effects [29]. Therefore, this review aims to: (i) recapitulate the existing information on W effects on the Mo-enzymes, (ii) summarize W effects on plant growth, cytology and genome expression under W stress, and (iii) outline the potential mechanisms by which W exerts its effects beyond Mo-enzyme inhibition. As such, this review should be considered as a complement to the few existing reviews in plants [29] and the many more in other organisms (e.g., [10,17,30,31,32,33]).
2. W Effects on Plants
2.1. W Uptake and Accumulation
W is uptaken by the roots and translocated to the upper plant organs via the xylem. W accumulation in the root tissues is much higher than in the shoot [4,34,35]. Anthocyanins may also facilitate W accumulation in some plant species [36]. Plants growing on abandoned mines that are heavily contaminated with several metals and metalloids including W may accumulate metals and evolve tolerance against these pollutants. For instance, plants of Calluna vulgaris naturally growing in an abandoned early twentieth century W mine in UK accumulated W in the root tissues at 56% of the total amount of W calculated in the spoil (1,637 mg kg−1) separated from the rhizosphere [4]. Ryegrass is another plant able to accumulate W; its seedlings germinated in soil amended with high concentrations of metallic W (10,000 mg kg−1) accumulated significant amounts of the metal in the leaves (13,535 ± 6,125 mg kg−1) after two months [19]. Many more plant species have been indentified having the potential of W accumulation, some of which are Digitalis purpurea, Chamaespartium tridentatum, Cistus ladanifer, Pinus pinaster, Erica umbellata and Quercus ilex subsp. ballota [3]. Except natural outcrops, cultivated plants have been shown to uptake W and were used for W and cobalt removal from wastewaters when applied to agricultural fields [37,38]. In particular, cabbage and carrot fields were irrigated with wastewater (or 50% wastewater diluted with unpolluted water) from a W and cobalt processing unit. These experiments have indicated that migration of the metals to the soil is limited when plants are grown in the field as compared to fields without plants. W-tolerant plant species may be useful in mine stabilization and re-vegetation strategies, while W-accumulating ones can potentially be applied in phytoremediation-based technologies for the cleanup of W-contaminated soils [3,4,19,34]. Accordingly, deeper knowledge about the effects of W on plants and the pathway W ions use in entering the cell is essential.
2.2. W-Mo Antagonism: W as an Inhibitor of Mo-Enzyme Activity
As soon as W enters the plant cell it may cause some adverse effects on specific targets. Of the first targets indentified were the Mo-enzymes, mostly due to W-Mo antagonism. Over evolution, due to their similar physical and chemical properties (Figure 1A), W and Mo have been incorporated into the active sites of key enzymes, highly broadening their catalytic diversity in the biological systems. In the hot and anaerobic conditions under which life probably arose, physicochemical properties of elements accounted so that W might have been first acquired by the early life forms than Mo. As the earth’s crust cooled and the atmosphere became aerobic, Mo presumably substituted W in the enzyme active sites [30]. Nowadays, the consensus is that Mo is required by most living organisms and the Mo-activated enzymes are encountered in all aerobic ones, whereas W occurs in the obligate, typically thermophilic anaerobes [17,30,39]. Otherwise, W is not necessary for eukaryotes and no “tungsto-enzymes” have so far been identified in them. The W-Mo antagonism for the same enzyme site has been inherited today when W occurs in high concentrations. This was already stated by 1956 in Asprergillus nidulans, where Higgins et al. [40] noticed that when nitrate was the sole nitrogen source, tungstate in a molar ratio of 20:1 to Mo readily induced growth inhibition.
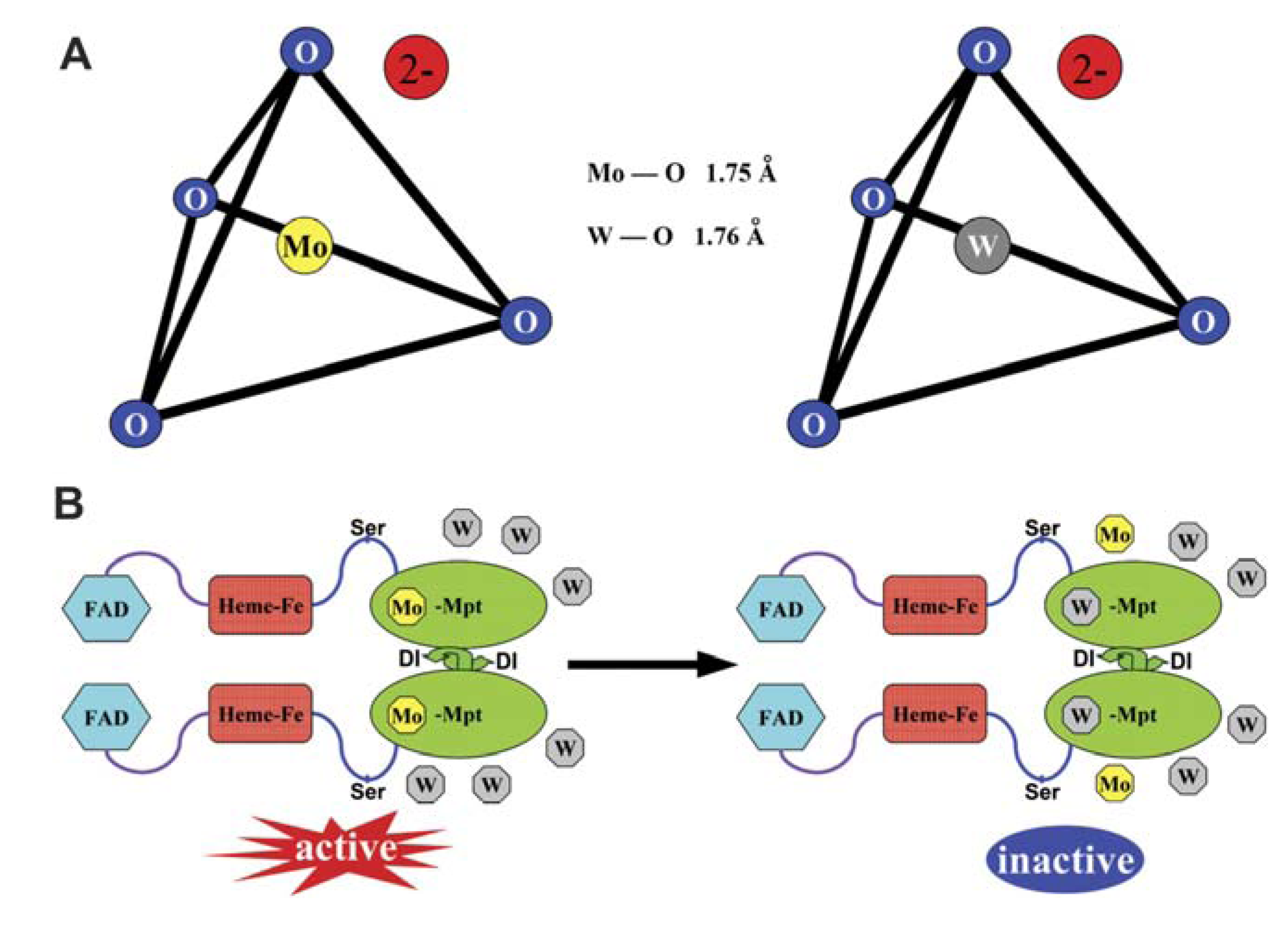
Figure 1.
A comparison of tungstate and molybdate structure that underlies their antagonism for molybdenum cofactor (MoCo). (A) Structural schemes of tungstate and molybdate. The metal atoms are at the centre of a tetrahedron of oxygen atoms. (B) Tungstate competes with molybdate for incorporation into the MoCo, resulting in inactivation of nitrate reductase (NR). FAD: flavin adenine dinucleotide, Mo-Mpt: molybdenum-molybdopterin. Reprinted from [29], with kind permission from both the authors and Oxford University Press.
Figure 1.
A comparison of tungstate and molybdate structure that underlies their antagonism for molybdenum cofactor (MoCo). (A) Structural schemes of tungstate and molybdate. The metal atoms are at the centre of a tetrahedron of oxygen atoms. (B) Tungstate competes with molybdate for incorporation into the MoCo, resulting in inactivation of nitrate reductase (NR). FAD: flavin adenine dinucleotide, Mo-Mpt: molybdenum-molybdopterin. Reprinted from [29], with kind permission from both the authors and Oxford University Press.

Within the cell, W antagonizes effectively and may substitute Mo in the molybdenum cofactor (MoCo) of the Mo-enzymes (Figure 1B) [29]. Mo-enzymes are required for diverse key reactions in carbon, sulphur and nitrogen metabolism. In plants, they can be subdivided into two families: the xanthine oxidase family, represented by xanthine dehydrogenase (XDH) and aldehyde oxidase (AO); and the sulphite oxidase (SO) family, represented by SO and nitrate reductase (NR). Recently, the mitochondrial amidoxime reducing component (mARC) was also identified, but it has not yet been integrated into one of the aforementioned families [41].
Omarov et al. [42] reported that application of 250 μΜ of tungstate resulted in AO inhibition in barley roots and leaves. The authors used the inhibitory effect in order to conclude that Mo is involved in the catalytic centres of the AO. Moreover, in W-treated Zea mays plants or in plants grown in W-enriched soils, AO activity appeared significantly reduced [43]. In Zea mays, inhibition of drought-induced abscisic acid synthesis was noticed and it was suggested that the MoCo of AO may be important for the drought-induced abscisic acid biosynthesis. Similar reduction of AO activity was noticed in barley, when treated with increasing tungstate concentrations [44]. Also, the activity of XDH was sensitive to W. In barley roots, 100 and 500 μΜ tungstate in the nutrient medium drastically reduced XDH activity; again, the drop in enzyme activity was attributed to the reduction of the MoCo [44]. AO, XDH and SO have the ability to normally produce hydrogen peroxide (H2O2) [41]. Recently, it was found that tungstate inhibited the H2O2-generating activity of both SO and AO [45]. Regarding mARC, as far as we are aware no reports about the effect of W on this enzyme exist yet.
Plenty of studies have demonstrated the inhibitory action of W to NR activity, and in the form of sodium tungstate (Na2WO4) it was used as a casual inhibitor of NR [46,47]. So, since W could be easily applied, it was and is still used as a NR inhibitor in plant NR-research, in order to lower the production of nitric oxide (NO) (among others see [29,46,47,48]), since the NR-dependent NO formation is the best-characterized NO source in plants [49].
However, some researchers have questioned the ability of tungstate to affect NO production. Characteristic examples are the studies by Xiong et al. [50] and Wang et al. [51], who have reported that 100 μΜ of tungstate had no effect on endogenous NO concentration in rice and lupin, respectively. Moreover, tungstate inhibited primary and crown root elongation in rice plants [50]. So an assumption was made that tungstate could be toxic to plant roots and that tungstate-induced inhibition on root elongation does not correlate with endogenous NO content. More recently, Kumar et al. [52], while investigating the generation of NO in isolated protoplasts and chloroplasts of Brassica napus, found that NO production was not inhibited by 250 μM Na2WO4. Consequently, it comes as no surprise the question: “Tungstate: is it really a specific NR inhibitor in plant NO research?” [29].
Since there are doubts on the ability of tungstate to affect NO, and tungstate’s toxicity in roots could not be only due to NO inhibition, a question that unavoidably arises is: are Mo-enzymes the only targets of W toxicity? Recent experimentation revealed further subcellular targets, correlated with defects in root growth and morphology.
2.3. W Is More Than a Mo-Enzyme Inhibitor
2.3.1. W Inhibits Plant Growth
The first visible symptom of W toxicity was a general reduction of root and shoot growth. This has been observed in different plant species and some of the results are summarized in Table 1. Except for a reduction in plant growth or biomass production, notable is the case of W application in Brassica rapa, B. juncea and B. oleracea, where it caused the accumulation of a blue complex in the roots [36].

{kind=link}
{kind=link}
| Plant species | Treatments | Effects/Results | Reference |
|---|---|---|---|
| Brassica rapa | Seeds sown in Magenta boxes on ½ MS medium with 50-150 mg L−1 W as Na2WO4. | Increased production of a blue substance correlated with anthocyanin accumulation. | [36] |
| B. juncea | |||
| B. oleracea | |||
| Hordeum vulgare L. cv. Steptoe (barley) | 0.25, 0.5, 1, 10, 100, 500 μM Na2WO4, 9 days. Hydroponics. | Significant reduction in root and shoot biomass at high concentrations (≥10 μM). | [44] |
| Lolium perenne (ryegrass) | Urban and forest soil amended with ammunition grade W powder (average particle size 5 μM): 1, 10, 100, 1,000, 10,000 mg kg−1. Up to 9 months. | Serious reduction of plant growth and death after two months. | [19] |
| Pisum sativum cv. Onmard (pea) | 200, 500 mg L−1 Na2WO4, up to 8 days. Hydroponics. | Inhibition of root elongation and lateral root formation, retardment of seedling growth rate and new leaf emergence. | [25] |
| Gossypium hisutum cv. | |||
| Campo (cotton) | |||
| Helianthus annuus L. (sunflower) | Processed field soil, spiked with 6,500 mg kg−1 metallic W powder, aged for six months. Diluted soil samples contained from 0 to 6,500 mg W kg−1. Plants grown for two or four weeks. | Plant total weight significantly reduced at W concentration ≥2,600 mg kg−1, root and shoot length reduced at ≥3,900 mg kg−1. | [34] |
| Avena sativa cv. Ogle (oat) | Natural soil containing 0 (control), 0.803, 2.41, 7.21, 21.7, 65.0, 195.1, and 586 mg W kg−1 dry soil. | Toxicity was recorded at ~58.6-293 mg W kg−1 dry soil, with lettuce being more sensitive than radish, followed by oat. | [53] |
| Raphanus sativus cv. Crimson Giant | |||
| (radish) | |||
| Lactuca sativa cv. Grand Rapids (lettuce) | |||
| Triticum aestivum L. var Raj4037. | Potted soil watered with 3, 9, 27, 81, 243 mg kg−1 Na2WO4.2H2O solutions plus control. Samples harvested after 60 days. | Lower concentrations (3, 9 mg kg−1) of W had promotive effects in growth, biomass, chlorophyll, carbohydrate and Mo contents, higher ones decreased them. | [35] |
| Vigna unguiculata L. Walp. var. Sephali Shikha-313 | Potted soil watered with 5, 10, 15, 20, 25 μg g−1 Na2WO4.2H2O solutions. Samples harvested after fruiting. | Lower applied doses (5, 10, 15 μg g−1) of W promoted root-shoot length. Higher doses retarded root-shoot length. | [54] |
| Brassica oleracea (cabbage) | Grown in aged W powder-spiked soil containing monomeric and polymeric tungstates provided as Na2WO4·2H2O. | Cabbage growth was impaired at 436 mg W kg−1 W soil. | [23] |
| Lactuca sativa (lettuce) |
However, the mechanism by which W exerted this effect has not been extensively studied, and commonly visible defects were correlated with the decreased activity of the Mo-enzymes in response to increasing amounts of tungstate [29,43]. Several further assumptions have been made. W could affect plants indirectly, such as by lowering soil pH [53], which alters the availability of nutrients in the soil. For instance, W is oxidized to tungstate in soil and in turn it can be polymerized with phosphate, depleting the latter from the soil [55].
Another indirect mechanism of toxicity may be the alteration of soil microflora necessary for plant survival. Reports on W effects on microbes have shown that W can either be beneficial or detrimental for the microbial biomass. For example, Strigul et al. [19] reported a statistically significant decrease in microbial biomass in Bacillus subtilis and Pseudomonas fluorescens. The above study showed that the presence of metallic W resulted in the death of a substantial portion of the bacterial component of the soil community and an increase of fungal biomass. On the other hand, Rigelberg et al. [20] reported a general positive effect of W on soil microbial biomass, as revealed by the increase in Gram-negative and Gram-positive bacterial fatty acids at relatively low W concentrations and negative effects at higher concentrations. Of the most important soil bacteria affected by W are the nitrogen-fixing ones, as for example the Azotobacter vinelandii [20]. W may also affect the N2-reducing capacity of these bacteria by substituting Mo in nitrogenase, as it was found in Rhodobacter capsulatus [56].
W could also affect root growth directly by targeting specific cellular components or biochemical pathways (besides the Mo-enzymes). The effects of W on phosphate-dependent biochemical pathways have been already examined in animal cells [57] and remain yet to be studied in plants. For instance, W may disturb phosphate concentration inside the cell, altering phosphate homeostasis within the tissues, disrupting phosphorylating reactions including cell signalling pathways or even production of adenosine triphosphate (ATP). Moreover, inhibition of root elongation may be related with ultrastructural defects caused by W in root cells [25,26,28]. Root elongation is achieved by both cell division and expansion [58], both of which are reported to be affected by W.
Recent reports maintain that W, like other heavy metals, affected nucleus morphology and cell division [25,28]. In particular, the nuclei of W-treated root cells in Pisum sativum and Gossypium hirsutum seedlings appeared larger and frequently multi-lobed, as compared to the rounded and smooth ones of untreated samples. The chromatin of W-affected nuclei became condensed and peripherally distributed, and in some cases atypical double membrane structures occurred in the nucleoplasm defining chromatin condensation in sub-peripheral compartments. Cytoplasmic components such as ribosomes, mitochondria, membranes, vesicles or small vacuoles and lipid droplets, were frequently entrapped within the nucleoplasm of W-affected nuclei. These intranuclear entrapments imply a defect in cell division. Moreover, nucleoli of W-affected cells were more prominent and contained bigger nucleolar vacuoles than those of untreated ones. Also, in W-treated roots incomplete, misaligned or unilaterally extended cytokinetic walls were readily recognized. These cytokinetic aberrations were most likely attributed to defective MT polymerization and function, as phragmoplast MTs were reduced in number [25,28]. There are many reports stating that toxic metal ions produce several defects in dividing plant cells, including abnormal mitoses, decreased mitotic index, alterations of nucleus and nucleolus morphology, cell cycle aberrations and DNA defects [59]. Similarly, evidence has been reported that W affects plant DNA: W particles caused a breakage of phosphodiester bonds in native DNA at a limited number of sites in wheat embryos after a biolistic transformation [60].
A common target of W toxicity among different plant species are the cortical MTs, as this was revealed by exposing taxonomically diverse land plant taxa representing monocots (Allium cepa, Zea mays), dicots (Arabidopsis thaliana), gymnosperms (Pinus brutia), pterophytes (Adiantum capillus-veneris) and bryophytes (Physcomitrella patens, Tortula muralis), to aqueous Na2WO4 solutions [27]. As cortical MTs are of critical importance for cell shaping and anisotropic elongation, disruption by W could also be responsible for root growth inhibition [26], which is a general trait of heavy metal and metalloid toxicity ([61,62]; and references therein). Although the mechanism by which W induced this effect is unclear, it seems to be a side effect of a more prominent response, probably involving programmed cell death (PCD) processes [28].
2.3.2. W-Induced Programmed Cell Death (PCD)
One of the main responses of plants to developmental and biotic/abiotic environmental stimuli is the execution of cell “suicide” or programmed cell death (PCD) [63,64]. PCD is a genetically determined process found throughout animal and plant kingdoms, involving the actively controlled and precise degradation of cellular components, aiming at selective elimination of harmful, unwanted or damaged cells in eukaryotes [64]. Undesirable PCD may also be instigated by many biotic agents such as pathogens [65] and abiotic factors like extreme temperatures [66]. Different types of PCD in plants have been recognized, all sharing a unique process that depends on vacuole collapse, which releases sequestered hydrolases that degenerate cell components and lead to cell death [67,68,69]. Cleavage of nuclear DNA into oligonucleosomal fragments (DNA laddering) is a potential indicator of PCD in plants [63,70,71]. Plant developmental PCD (e.g., the differentiation of tracheary elements of xylem) has been intensively studied and in most cases it is triggered by plant hormones [64,65,67,69].
The execution of a kind of W-induced PCD in plants was recently documented for the first time in root tip cells of Pisum sativum exposed to 200 mg L−1 Na2WO4 for 12, 24, 48 and 72 h [28], rendering cell death a common feature of W toxicity occurring in both plants and animals [21,22]. However, W-induced PCD (W-PCD for brevity) has been stated to exert certain peculiarities [28], therefore its features are being compared with those of other metal-induced PCD cases, which are summarized in Table 2.
| Metal | Plant species | Conditions and responses | Reference |
|---|---|---|---|
| Cd | Nicotiana tabacum Mill. | Chronic exposure of suspension cells to 50-100 mM CdSO4 induced apoptotic-like PCD, including DNA fragmentation into oligonucleosomal units (50-200 kb fragments). | [72] |
| BY-2 cell line | |||
| Al | Hordeum vulgare | In root-tip cells 0.1-1.0 mM Al treatments for 8 h induced PCD, possibly via a ROS-modified signal transduction pathway, whereas 10-50 mM Al treatments caused necrosis. | [73] |
| Cd | Lycopersicon esculentum Mill. (tomato). Cell line MsK8 | Cadmium (CdSO4) induced apoptotic-like PCD that required increased H2O2 production and activation of phospholipase C and D and ethylene signalling pathways. | [74] |
| Al | Lycopersicon esculentum Mill. (tomato). Cell line MsK8 | Suspension cells treated with 100 μM AlCl3 showed typical features of PCD (nuclear and cytoplasmic condensation), executed by caspase-like proteases. | [75] |
| Cd | Lycopersicon esculentum Mill. (tomato). Cell line MsK8 | 0.1 mM CdSO4 in cell suspension culture induced cell death after 24 h, involving caspase-like proteases, indicating that PCD took place. | [76] |
| Cd | Nicotiana tabacum L. (tobacco) BY-2 cell line | 50 μM CdSO4 induced internucleosomal DNA fragmentation connected with the action of cysteine proteases and the loss of membrane integrity, in particular of tonoplast. | [70] |
| W | Pisum sativum L. cv. Onmard (pea) | Root tip cells of young seedlings exposed to 200 mg L−1 Na2WO4 for 12-72 h executed PCD through ER stress-UPR. The expression of the PCD-related genes DAD-1 and HSR203J was altered. | [28] |
DNA laddering did not seem to be induced by W but only a slight DNA smearing, in contrast to what was observed in the case of cadmium [72] and aluminium [73]. DNA laddering is however a hallmark of the majority but not of all the cases of cells that undergo PCD [70]. Similarly to other metal-induced PCD phenomena, W-PCD appears to involve the participation of caspase-like enzymes (cf. [28], [75,76]). Caspase-like proteases or metacaspases, which have a common substrate with caspases, were found to participate in plant PCD [77,78]. Caspase inhibitors have been used to prevent the action of the above enzymes [78,79]. The pan-caspase inhibitor Z-VAD-FMK has been applied to prevent various kinds of PCD in plant cells [80,81], and it was indicated that caspase-like proteases might participate in W-PCD [28]. Moreover, the involvement of the 26S proteasome in W-PCD was indirectly shown in W-treated roots. Inhibition of the 26S proteasome activity by MG132 (Z-Leu-Leu-Leu-al) reduced considerably the effects of W on the W-affected cells, thus preventing W-PCD. Considering that the 26S proteasome is known to exhibit some caspase-like activity both in animal or yeast cells [82] and plant cells [83], it was concluded that the 26S proteasome is involved in W-PCD in plant cells [28].
How is this W-PCD being triggered? Aluminium-induced PCD is initiated by reactive oxygen species (ROS) production [73]. Low doses of ROS can induce the production of antioxidant enzymes, but when the concentration of ROS reaches a certain threshold a signal transduction pathway that results in PCD is activated, while high doses of ROS result in necrosis [84]. Similar evidence was obtained in cadmium-induced PCD, where the oxidative stress appeared to be “instrumental” in cell death [74]. In fact, redox changes are sensed by the plant cell as a “warning” message and, depending on the situation, genetic programs leading to stress acclimation or PCD are switched on [85]. Information about the participation of ROS signals during W application is currently lacking in plants and could be the objective of a future study. ROS induction has been already observed in mammalian cells treated with W-based nanoparticles [24].
In animal cells, heavy metals could lead to apoptosis through an endoplasmic reticulum (ER) stress response ([86]; and references therein). ER-stress is also found to be associated with a PCD-triggering pathway in plants [87,88]. Adamakis et al. [28] showed that W caused DAD-1 silencing in W-treated roots of Pisum sativum. DAD-1 encodes a subunit of oligosaccharyltransferase, which participates in N-linked protein glycosylation in the ER, a post-transcriptional modification fundamental for proper protein folding [89]. Inhibition of N-linked protein glycosylation, as for example with tunicamycin [90], results in the accumulation of unfolded proteins in the ER lumen, initiating the unfolded protein response (UPR), which in turn triggers PCD [88,91]. DAD-1 silencing along with increased expression of three genes closely related with ER stress and UPR (BiP-D, bZIP28 and bZIP60) and additional indirect supportive evidence derived from co-treatments of W and 4-phenylbutyric acid (a chemical suppressor of ER stress), were all consistent with the general conclusion that W induces a kind of PCD in plant cells triggered by the ER stress-UPR pathway [28]. The details, however, of the mechanisms involved are obscure and remain to be further disclosed.
2.4. A Model for W Entrance in Plant Cells
To sum up, besides Mo-enzyme inhibition W appears to induce a wide range of effects after it enters the plant cells. But how does W enter plant cells? Could it use the same entrance pathway as Mo does? The already described subcellular effects and additional pharmacological evidence [26] permit the proposal of a hypothetical model for W uptake depending on the entrance pathway of Mo, as is discussed below.
The uptake of Mo (as molybdate anion, MoO42−) in plant cells starts with the attachment of Mo to a plasma transmembrane anion transporter. The available data are not clear on the identity of this transporter, although two proteins (MOT1 and MOT2) were identified in Arabidopsis thaliana as Mo transporters. However, none of them was found to be located in the plasma membrane [92]. After its entry, Mo interacts with the Cnx1 protein, which is bound to the actin microfilaments of the cortical cytoplasm [93], in the same way as its mammalian homologue gephyrin binds to tubulin [94]. Cnx1 may interact with a transmembrane anion transporter to facilitate the Mo transport towards molybdopterin to form the MoCo [93].
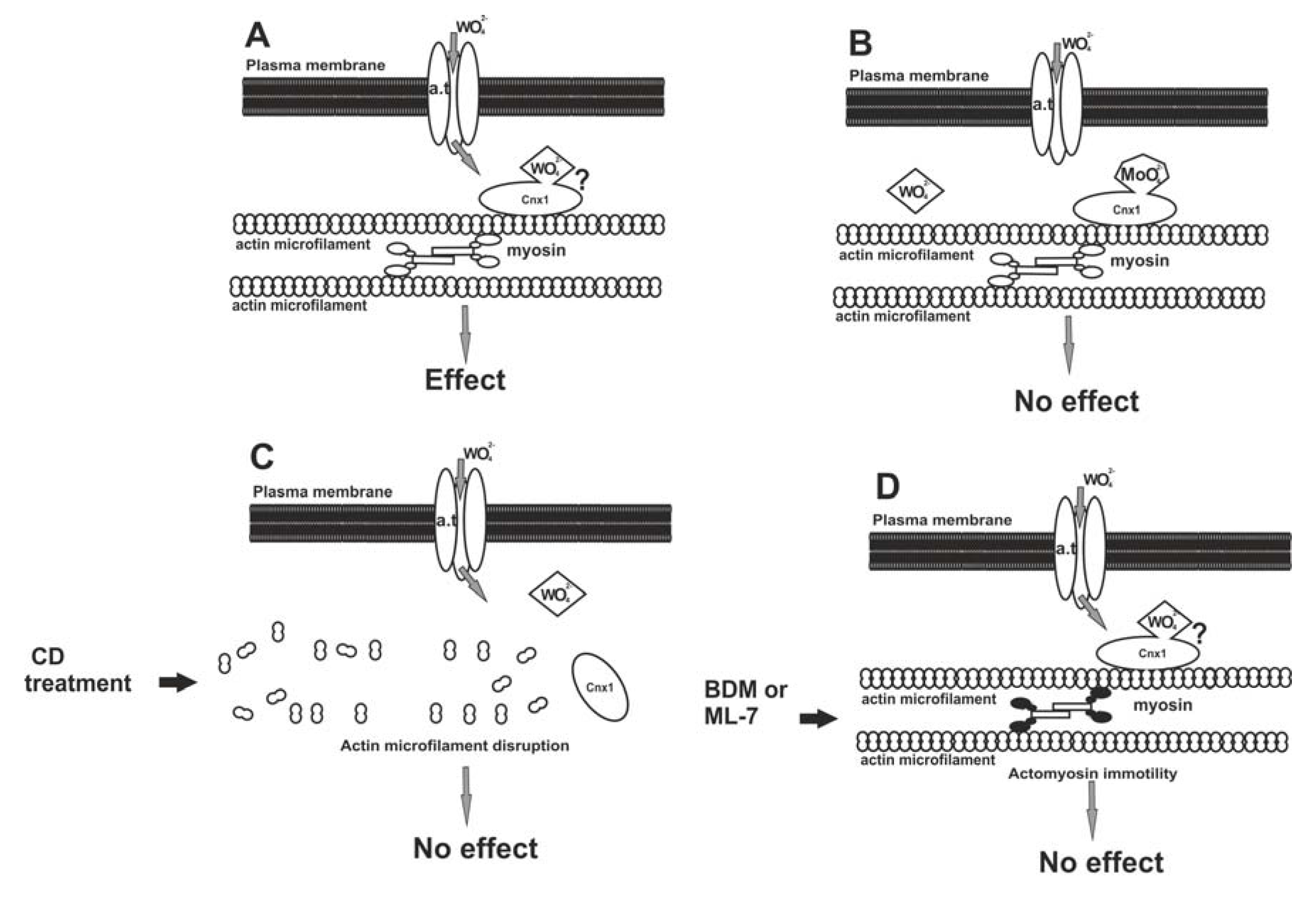
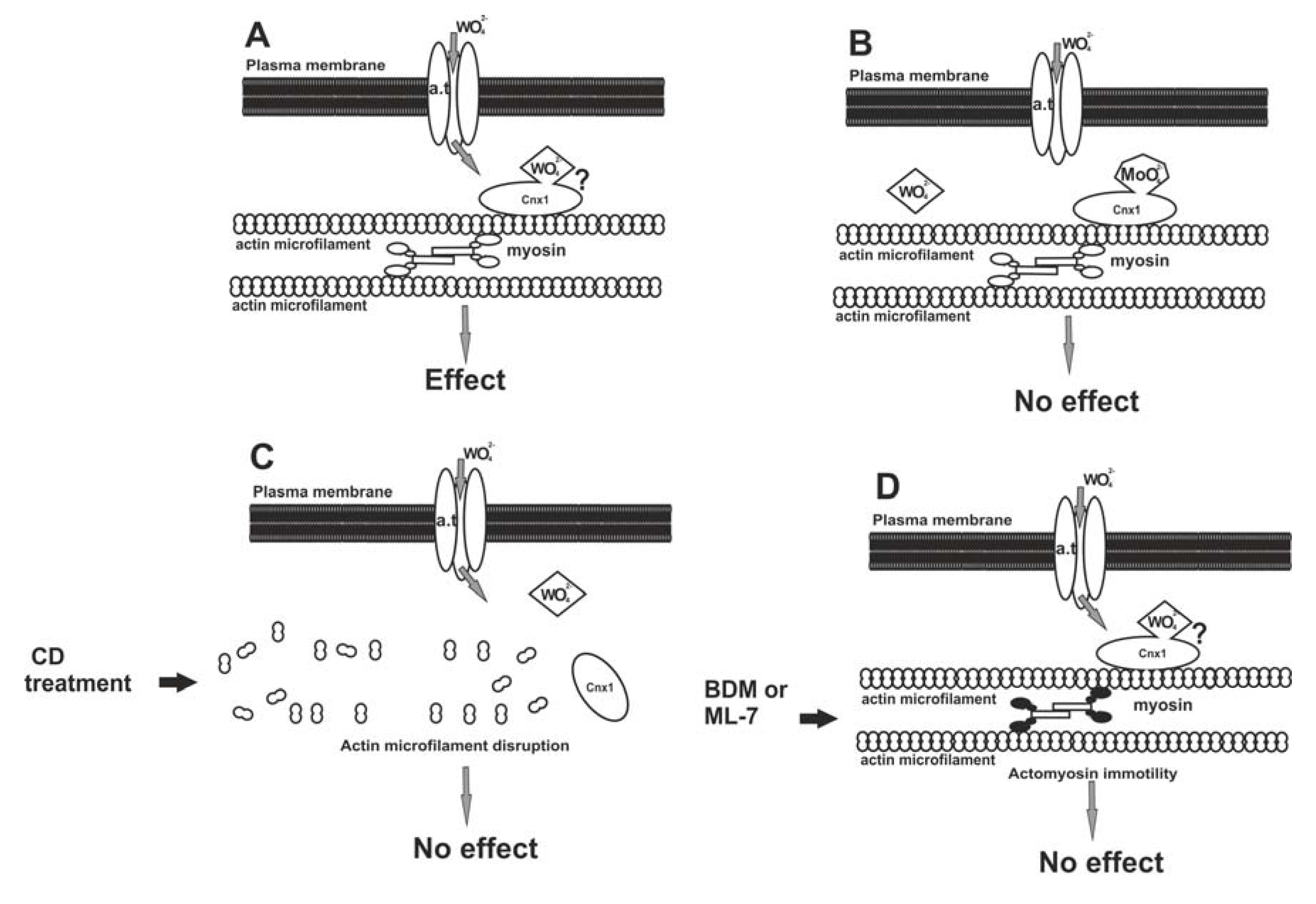
It seems that W may enter the plant cell by exploiting the Mo pathway and binding to the Cnx1 protein at the cell cortex, from where it may be transported in the cytoplasm by actomyosin mobility to exert its deleterious effects (Figure 2A). Should this pathway model be correct, any inhibition of the succession is expected to prevent W to become harmful. Considering this hypothesis, a pharmacological administration involving W, Mo, anti-actin and anti-myosin drugs, singly and in combinations, was staged and the effects were assessed depending on MT morphology [26]. In this research, roots were treated with Na2MoO4, actin microfilaments were disassembled with cytochalasin D and actomyosin mobility was arrested with BDM (2,3-butanedione monoxide) or ML-7 [1-(5-iodo-naphthalene-1-sulphonyl)-1H-hexahydro-1,4-diazepine] prior to W application. When tissues were pre-treated with Mo, subsequent exposure to W did not affect MTs, apparently due to occupation of all binding sites of Cnx1 molecules by Mo, leaving no vacant sites for W (Figure 2B). When actin microfilaments were experimentally disrupted, the Cnx1 protein was most probably misplaced, not providing the anchor for tungstate anions even though they had entered the cell (Figure 2C). If the actomyosin mobility was inactivated, tungstate anions failed to be distributed intracellularly, though bound to Cnx1, and became ineffective on cortical MTs (Figure 2D). This “protection” of MTs by actin microfilament disruption or actomyosin mobility inactivation further supports the above working hypothesis.
However, in order to “solidify” the above model further experimentation is needed. For example besides microtubules actin microfilaments (the other component of plant cytoskeleton) could be affected by W as well. Indeed, actin microfilaments were almost totally disrupted in W-treated roots of Pisum sativum, in parallel with MT disorganization but this effect was correlated with the W-PCD [28], while preliminary experimentation revealed that W treatments induced microfilament bundling [95]. A specific investigation on the effects of W on actin microfilaments is missing and, should that be carried out, it would provide valuable information on the intracellular cascade of events triggered by W toxicity in plant cells.
Figure 2.
A model for the entry of W in the plant cell, through the pathway of Mo. (A) Tungstate anions (WO42−) pass through the plasma membrane by a transmembrane anion transporter (a.t.) protein. The Cnx1 protein is located close to the plasma membrane due to its binding to cortical actin microfilaments. WO42− anions could bind to Cnx1 and be distributed inside the cytoplasm by actomyosin motility. (B) Pre-treatment with molybdate results in occupation of the Mo-binding sites of Cnx1, leaving no vacant sites for WO42− anions. This prevents the latter from exerting their effect. (C) Cnx1 is misplaced due to actin microfilament disruption by cytochalasin D (CD). As a result, WO42− anions cannot bind to Cnx1, fail to be distributed inside the cell and do not exert the W effect. (D) Inhibition of myosin function by BDM or ML-7 results in arrest of actomyosin-based motility. As a consequence, though bound to Cnx1, WO42− anions fail to be distributed. Reprinted from [26], after the model proposed by [93], modified, with kind permission from John Wiley and Sons.
Figure 2.
A model for the entry of W in the plant cell, through the pathway of Mo. (A) Tungstate anions (WO42−) pass through the plasma membrane by a transmembrane anion transporter (a.t.) protein. The Cnx1 protein is located close to the plasma membrane due to its binding to cortical actin microfilaments. WO42− anions could bind to Cnx1 and be distributed inside the cytoplasm by actomyosin motility. (B) Pre-treatment with molybdate results in occupation of the Mo-binding sites of Cnx1, leaving no vacant sites for WO42− anions. This prevents the latter from exerting their effect. (C) Cnx1 is misplaced due to actin microfilament disruption by cytochalasin D (CD). As a result, WO42− anions cannot bind to Cnx1, fail to be distributed inside the cell and do not exert the W effect. (D) Inhibition of myosin function by BDM or ML-7 results in arrest of actomyosin-based motility. As a consequence, though bound to Cnx1, WO42− anions fail to be distributed. Reprinted from [26], after the model proposed by [93], modified, with kind permission from John Wiley and Sons.
3. Conclusions and Perspectives
As already stated “…it appears that environmental obscurity for W and its compounds has ended and environmental scrutiny has emerged” [1]. In line to this statement, recent advances on the toxic attributes of W in plants revealed a range of adverse effects on morphological, cytological and gene expression levels. W effects on plants could therefore be analyzed under a broadened view that W should not be considered solely a Mo-enzyme inhibitor but a heavy metal having a wide range of effects on plants similarly to other heavy metals [29]. The mechanism by which W is trapped in the roots, the motility of the metal inside the plant body, its cumulative ability and the way it enters in the plant cells are interesting issues to be further examined. Research is also needed in the context of W contamination management and development of phytoremediation technologies.
Acknowledgements
This work is part of a PhD thesis (Adamakis S. I-D.) co-financed by the European Union (European Social Fund—ESF) and Greek national funds through the Operational Program “Education and Lifelong Learning” of the National Strategic Reference Framework (NSRF)—Research Funding Program: Heracleitus II. The authors would also like to thank the anonymous reviewers for their constructive suggestions.
References
- Koutsospyros, A.; Braida, W.; Christodoulatos, C.; Dermatas, D.; Strigul, N. A review of tungsten: From environmental obscurity to scrutiny. J. Hazard. Mater. 2006, 136, 1–19. [Google Scholar] [CrossRef]
- Pyatt, F.B.; Pyatt, A.J. The bioaccumulation of tungsten and copper by organisms inhabiting metalliferous areas in North Queensland, Australia: An evaluation of potential health implications. J. Environ. Health Res. 2004, 3, 13–18. [Google Scholar]
- Pratas, J.; Prasad, M.N.V.; Freitas, H.; Conde, L. Plants growing in abandoned mines of Portugal are useful for biogeochemical exploration of arsenic, antimony, tungsten and mine reclamation. J. Geochem. Explor. 2005, 85, 99–107. [Google Scholar] [CrossRef]
- Wilson, B.; Pyatt, F.B. Bio-availability of tungsten in the vicinity of an abandoned mine in the English Lake District and some potential health implications. Sci. Total Environ. 2006, 370, 401–408. [Google Scholar] [CrossRef]
- Senesi, N.; Padovaro, G.; Brunetti, G. Scandium, titanium, tungsten and zirconium content in commercial inorganic fertilizers and their contribution to soil. Environ. Techn. Lett. 1988, 9, 1011–1020. [Google Scholar] [CrossRef]
- Clausen, J.L.; Korte, N. Environmental fate of tungsten from military use. Sci. Total Environ. 2009, 407, 2887–2893. [Google Scholar]
- Karachalios, A.; Wazne, M.; Betancur, J.N.; Christodoulatos, C.; Braida, W.; O’Connor, G. Immobilization of copper, lead, and tungsten in mixed munitions firing rang. Contaminated soils by various amendments. J. Haz. Toxic Radioact. Waste 2011, 15, 151–159. [Google Scholar] [CrossRef]
- Sadiq, M.; Mian, A.A.; Althagafi, K.M. Inter-city comparison of metals in scalp hair collected after the Gulf War 1991. J. Environ. Sci. Health A 1992, 27, 1415–1431. [Google Scholar]
- Harper, E.M.; Graedel, T.E. Illuminating tungsten’s life cycle in the United States: 1975–2000. Environ. Sci. Technol. 2008, 42, 3835–3842. [Google Scholar]
- Clemente, R.; Nicholas, W.L. Tungsten. In Heavy Metals in Soils: Trace Metals and Metalloids in Soils and Their Bioavailability; Alloway, B.J., Ed.; Springer Science and Business Media: Dordrecht, Germany, 2013. [Google Scholar] [CrossRef]
- Thomas, V.G.; Roberts, M.J.; Harrison, P.T. Assessment of the environmental toxicity and carcinogenicity of tungsten-based shot. Ecotoxicol. Environ. Saf. 2009, 72, 1031–1037. [Google Scholar] [CrossRef] [Green Version]
- Kelly, A.D.R.; Lemaire, M.; Young, Y.K.; Eustache, J.H.; Guilbert, C.; Molina, M.F.; Mann, K.K. In vivo tungsten exposure alters B cell development and increases DNA damage in murine bone marrow. Toxicol. Sci. 2012. [Google Scholar] [CrossRef]
- Strigul, N.S.; Koutsospyros, A.; Christodoulatos, C. Tungsten in the former Soviet Union: Review of environmental regulations and related research. Land Contam. Reclam. 2009, 17, 189–215. [Google Scholar] [CrossRef]
- Sheppard, P.R.; Bierman, B.J.; Rhodes, K.; Ridenour, G.; Witten, M.L. Comparison of size and geography of airborne tungsten particles in Fallon, Nevada, and Sweet Home, Oregon, with implications for public health. J. Environ. Public Health 2012. [Google Scholar] [CrossRef]
- Steinmaus, C.; Lu, M.; Todd, R.; Smith, A.H. Probability estimates for the unique childhood leukemia cluster in Fallon, Nevada, and risks near other U.S. military aviation facilities. Environ. Health Perspect. 2004, 112, 766–771. [Google Scholar] [CrossRef]
- CDC (2003). Cross-sectional exposure assessment of environmental contaminants of Churchill County, Nevada. Final Report. Available online: http://www.cdc.gov/nceh/clusters/Fallon/study.htm/ (accessed on 5 November 2012).
- Kletzin, A.; Adams, M.W.W. Tungsten in biological systems. FEMS Microbiol. Rev. 1996, 18, 5–63. [Google Scholar]
- Kalinich, J.F.; Emond, C.A.; Dalton, T.K.; Mog, S.R.; Coleman, G.D.; Kordell, J.E.; Miller, A.C.; McClain, D.E. Embedded weapons-grade tungsten alloy shrapnel rapidly induces metastatic high-grade rhabnomyosarcomas in F344 rats. Environ. Health Perspect. 2005, 113, 729–734. [Google Scholar] [CrossRef]
- Strigul, N.; Koutsospyros, A.; Arienti, P.; Christodoulatos, C.; Dermatas, D.; Braida, W. Effects of tungsten on environmental systems. Chemosphere 2005, 61, 248–258. [Google Scholar]
- Ringelberg, D.B.; Reynolds, C.M.; Winfield, L.E.; Inouye, L.S.; Johnson, D.R.; Bednar, A.J. Tungsten effects on microbial community structure and activity in a soil. J. Environ. Qual. 2009, 38, 103–110. [Google Scholar] [CrossRef]
- Osterburg, A.R.; Robinson, C.T.; Schwemberger, S.; Mokashi, V.; Stockelman, M.; Babcock, G.F. Sodium tungstate (Na2WO4) exposure increases apoptosis in human peripheral blood lymphocytes. J. Immunotoxicol. 2010, 7, 174–182. [Google Scholar] [CrossRef]
- Guilbert, C.; Kelly, A.D.R.; Petruccelli, L.A.; Lemaire, M.; Mann, K.K. Exposure to tungsten induces DNA damage and apoptosis in developing B lymphocytes. Leukemia 2011, 25, 1900–1904. [Google Scholar]
- Kennedy, A.J.; Johnson, D.R.; Seiter, J.M.; Lindsay, J.H.; Boyd, R.E.; Bednar, A.J.; Allison, P.G. Tungsten toxicity, bioaccumulation and compartmentalization into organisms representing two trophic levels. Environ. Sci. Technol. 2012. [Google Scholar] [CrossRef]
- Kühnel, D.; Scheffler, K.; Wellner, P.; Meissner, T.; Potthoff, A.; Busch, W.; Springer, A.; Schirmer, K. Comparative evaluation of particle properties, formation of reactive oxygen species and genotoxic potential of tungsten carbide based nanoparticles in vitro. J. Hazard. Mater. 2012, 227–228, 418–426. [Google Scholar] [CrossRef]
- Adamakis, I.-D.S.; Eleftheriou, E.P.; Rost, T.L. Effects of sodium tungstate on the ultrastructure and growth of pea (Pisum sativum) and cotton (Gossypiumhirsutum) seedlings. Environ. Exp. Bot. 2008, 63, 416–425. [Google Scholar] [CrossRef]
- Adamakis, I.-D.S.; Panteris, E.; Eleftheriou, E.P. Tungsten affects the cortical microtubules of Pisum sativum root cells: Experiments on tungsten-molybdenum antagonism. Plant Biol. 2010, 12, 114–124. [Google Scholar] [CrossRef]
- Adamakis, I.-D.S.; Panteris, E.; Eleftheriou, E.P. The cortical microtubules are a universal target of tungsten toxicity among land plant taxa. J. Biol. Res. (Thessaloniki) 2010, 13, 59–66. [Google Scholar]
- Adamakis, I-D.S.; Panteris, E.; Eleftheriou, E.P. The fatal effect of tungsten on Pisum sativum L. root cells: Indications for endoplasmic reticulum stress-induced programmed cell death. Planta 2011, 234, 21–34. [Google Scholar] [CrossRef]
- Xiong, J.; Fu, G.; Yang, Y.; Zhu, C.; Tao, L. Tungstate: Is it really a specific nitrate reductase inhibitor in plant nitric oxide research? J. Exp. Bot. 2012, 63, 33–41. [Google Scholar] [CrossRef]
- Hille, R. Molybdenum and tungsten in biology. Trends Biochem. Sci. 2002, 27, 360–367. [Google Scholar]
- Keith, L.S.; Moffett, D.B.; Rosemond, Z.A.; Wohlers, D.W. Agency for Toxic Substances and Disease Registry. ATSDR evaluation of health effects of tungsten and relevance to public health. Toxicol. Ind. Health. 2007, 23, 347–387. [Google Scholar] [CrossRef]
- Jonas, W.; van der Voet, G.B.; Todorov, T.I.; Centeno, J.A.; Ives, J.; Mullick, F.G. Metals and health: A clinical toxicological perspective on tungsten and review of the literature. Mil. Med. 2007, 172, 1002–1005. [Google Scholar]
- Jelikić-Stankov, M.; Uskoković-Marković, S.; Holclajtner-Antunović, I.; Todorović, M.; Djurdjević, P. Compounds of Mo, V and W in biochemistry and their biomedical activity. J. Trace Elem. Med. Biol. 2007, 21, 8–16. [Google Scholar]
- Johnson, D.R.; Inouye, L.S.; Bednar, A.J.; Clarke, J.U.; Winfield, L.E.; Boyd, R.E.; Ang, C.Y.; Goss, J. Tungsten bioavailability and toxicity in sunflowers (Helianthus annuus L.). Land Contam. Reclam. 2009, 17, 141–151. [Google Scholar] [CrossRef]
- Kumar, A.; Aery, N.C. Effect of tungsten on growth, biochemical constituents, molybdenum and tungsten contents in wheat. Plant Soil Environ. 2011, 57, 519–525. [Google Scholar]
- Hale, K.L.; Tufan, H.A.; Pickering, I.J.; George, G.N.; Terry, N.; Pilon, M.; Pilon-Smits, E.A.H. Anthocyanins facilitate tungsten accumulation in Brassica. Physiol. Plant. 2002, 116, 351–358. [Google Scholar] [CrossRef]
- Seidov, I.M. Apropos of the soil method of decontaminating sewage waters containing cobalt and wolfram. Gig. Sanit. 1963, 28, 93–96. [Google Scholar]
- Seidov, I.M. Some data on the effect of vegetables enriched with cobalt and wolfram on the animal body. Vopr. Pitan. 1964, 23, 73–77. [Google Scholar]
- Johnson, M.K.; Rees, D.C.; Adams, M.W.W. Tungstoenzymes. Chem. Rev. 1996, 96, 2817–2839. [Google Scholar] [CrossRef]
- Higgins, E.S.; Richert, D.A.; Westerfeld, W.W. Molybdenum deficiency and tungstate inhibition studies. J. Nutr. 1956, 59, 539–559. [Google Scholar]
- Bittner, F.; Mendel, R.R. Cell biology of molybdenum. In Cell Biology of Metals and Nutrients; Hell, R., Mendel, R.R., Eds.; Springer Press: Heidelberg, Germany, 2010; Volume 17, Plant Cell Monographs. [Google Scholar]
- Omarov, R.T.; Akaba, S.; Koshiba, T.; Lips, S.W. Aldehyde oxidase in roots, leaves and seeds of barley (Hordeum vulgare L.). J. Exp. Bot. 1999, 50, 63–69. [Google Scholar]
- Jiang, F.; Heilmeier, H.; Hartung, W. Abscisic acid relations of plants grown on tungsten enriched substrates. Plant Soil 2007, 301, 37–49. [Google Scholar] [CrossRef]
- Jiang, X.Y.; Omarov, T.; Yesbergenova, S.Z.; Sagi, M. The effect of molybdate and tungstate in the growth medium on abscisic acid content and the Mo-hydroxylases activities in barley (Hordeum vulgare L.). Plant Sci. 2004, 167, 297–300. [Google Scholar] [CrossRef]
- Brychkova, G.; Yarmolinsky, D.; Fluhr, R.; Sagi, M. The determination of sulfite levels and its oxidation in plant leaves. Plant Sci. 2012, 190, 123–130. [Google Scholar] [CrossRef]
- Xu, J.; Yin, H.X.; Liu, X.J.; Yuan, T.; Mi, Q.; Yang, L.L.; Xie, Z.X.; Wang, W.Y. Nitric oxide alleviates Fe deficiency-induced stress in Solanum nigrum. Biol. Plantarum 2009, 53, 784–788. [Google Scholar] [CrossRef]
- Chen, J.; Zhong, Y.M.; Zhang, H.Q.; Shi, Z.Q. Nitrate reductase-dependent nitric oxide production is involved in microcystin-LR-induced oxidative stress in Brassica rapa. Water Air Soil Pollut. 2012, 223, 4141–4152. [Google Scholar] [CrossRef]
- Yu, L.Z.; Wu, X.Q.; Ye, J.R; Zhang, S.N.; Wang, S. NOS-like-mediated nitric oxide is involved in Pinus thunbergii response to the invasion of Bursaphelenchus xylophilu. Plant Cell Reports 2012. [Google Scholar] [CrossRef]
- Kaiser, W.M.; Weine, H.; Kandlbinder, A.; Tsai, C.B.; Rockel, P.; Sonoda, M.; Planchet, E. Modulation of nitrate reductase: Some new insights, an unusual case and a potentially important side reaction. J. Exp. Bot. 2002, 53, 875–882. [Google Scholar] [CrossRef]
- Xiong, J.; Lu, H.; Lu, K.; Duan, Y.; An, L.; Zhu, C. Cadmium decreases crown root number by decreasing endogenous nitric oxide which is indispensable for crown root primordial initiation in rice seedlings. Planta 2009, 230, 599–610. [Google Scholar]
- Wang, B.L.; Tang, X.Y.; Cheng, L.Y.; Zhang, A.Z.; Zhang, W.H.; Zhang, F.S.; Liu, J.Q.; Cao, Y.; Allan, D.L.; Vance, C.P.; et al. Nitric oxide is involved in phosphorus deficiency-induced cluster-root development and citrate exudation in white lupin. New Phytol. 2010, 187, 1112–1123. [Google Scholar]
- Kumar, R.T.; Prommer, J.; Watanabe, M. Endogenous nitric oxide generation in protoplast chloroplasts. Plant Cell Reports 2012. [Google Scholar] [CrossRef]
- Bamford, J.E.; Butler, A.D.; Heim, K.E.; Pittinger, C.A.; Lemus, R.; Staveley, J.P.; Lee, K.B.; Venezia, C.; Pardus, M.J. Toxicity of sodium tungstate to earthworm, oat, radish, and lettuce. Environ. Toxicol. Chem. 2011, 30, 2312–2318. [Google Scholar] [CrossRef]
- Kumar, A.; Aery, N.C. Effect of tungsten on the growth, dry-matter production, and biochemical constituents of cowpea. Commun. Soil Sci. Plant Anal. 2012, 43, 1098–1107. [Google Scholar] [CrossRef]
- Bednar, A.J.; Jones, W.T.; Boyd, R.E.; Ringelberg, D.B.; Larson, S.L. Geochemical parameters influencing tungsten motility in soils. J. Environ. Qual. 2008, 37, 399–402. [Google Scholar]
- Siemann, S.; Schneider, K.; Oley, M.; Mueller, A. Characterization of a tungsten-substituted nitrogenase isolated from Rhodobacter capsulatus. Biochemistry 2003, 42, 3846–3857. [Google Scholar] [CrossRef]
- Johnson, D.R.; Ang, C.; Bednar, A.J.; Inouye, L.S. Tungsten effects on phosphate-dependent biochemical pathways are species and liver cell line dependent. Toxicol. Sci. 2010, 116, 523–532. [Google Scholar] [CrossRef]
- Sanz, L.; James, A.H.M.; Dewitte, W. To divide and to rule; regulating cell division in roots during post-embryonic growth. In Progress in Botany; Ulrich, E.L., Beyschlag, W., Burkhard, B., Francis, D., Eds.; Springer Verlag: Berlin, Germany, 2012; Volume 72, pp. 57–80. [Google Scholar]
- Barceló, J.; Poschenrieder, C. Structural and ultrastructural changes in heavy metal exposed plants. In Heavy Metal Stress in Plants. From Molecules to Ecosystems; Prasad, M.N.V., Hagemeyer, J., Eds.; SpringerVerlag: Berlin, Germany, 1999; pp. 183–205. [Google Scholar]
- Krysiak, C.; Mazuś, B.; Buchowicz, J. Generation of DNA double-strand breaks and inhibition of somatic embryogenesis by tungsten microparticles in wheat. Plant Cell Tissue Organ Cult. 1999, 58, 163–170. [Google Scholar]
- Eleftheriou, E.P.; Adamakis, I.-D.S.; Melissa, P. Effects of hexavalent chromium on microtubule organization, ER distribution and callose deposition in root tip cells of Allium cepa L. Protoplasma 2012, 249, 401–416. [Google Scholar] [CrossRef]
- Eleftheriou, E.P.; Adamakis, I.-D.S.; Fatsiou, M.; Panteris, E. Hexavalent chromium disrupts mitosis by stabilizing microtubules in Lens culinaris Moench. root tip cells. Physiol. Plant. 2012. [Google Scholar] [CrossRef]
- Lam, E.; Pontier, D.; del Pozo, O. Die and let live—Programmed cell death in plants. Curr. Opin. Plant Biol. 1999, 2, 502–507. [Google Scholar]
- Gadjev, I.; Stone, J.M.; Gechev, T.S. Programmed cell death in plants: New insights into redox regulation and the role of hydrogen peroxide. In International Review of Cell and Molecular Biology; Elsevier Inc.: Amsterdam, The Netherland, 2008; Volume 270, pp. 87–144. [Google Scholar]
- Jones, A.M. Programmed cell death in development and defence. Plant Physiol. 2001, 125, 94–97. [Google Scholar] [CrossRef]
- Koukalova, B.; Kovarik, A.; Fajkus, J.; Siroky, J. Chromatin fragmentation associated with apoptotic changes in tobacco cells exposed to cold stress. FEBS Lett. 1997, 414, 289–292. [Google Scholar]
- Obara, K.; Kuriyama, H.; Fukuda, H. Direct evidence of active and rapid nuclear degradation triggered by vacuole rupture during programmed cell death in Zinnia. Plant Physiol. 2001, 125, 615–626. [Google Scholar] [CrossRef]
- Lam, E. Vacuolar proteases livening up programmed cell death. Trends Cell Biol. 2005, 15, 124–127. [Google Scholar]
- Kuriyama, H.; Fukuda, H. Developmental programmed cell death in plants. Curr. Opin. Plant Biol. 2002, 5, 568–573. [Google Scholar]
- Kuthanova, A.; Opatrny, Z.; Fischer, L. Is internucleosomal DNA fragmentation an indicator of programmed death in plant cells? J. Exp. Bot. 2008, 59, 2233–2240. [Google Scholar] [CrossRef]
- Reape, T.J.; Molony, E.M.; McCabe, P.F. Programmed cell death in plants: Distinguishing between different modes. J. Exp. Bot. 2008, 59, 435–444. [Google Scholar]
- Fojtová, M.; Kovařík, A. Genotoxic effect of cadmium is associated with apoptotic changes in tobacco cells. Plant Cell Environ. 2000, 23, 531–537. [Google Scholar] [CrossRef]
- Pan, J.W.; Zhu, M.Y.; Chen, H. Aluminum-induced cell death in root-tip cells of barley. Environ. Exp. Bot. 2001, 46, 71–79. [Google Scholar]
- Yakimova, E.T.; Kapchina-Toteva, V.M.; Laarhoven, L.J.; Harren, F.M.; Woltering, E.J. Involvement of ethylene and lipid signalling in cadmium-induced programmed cell death in tomato suspension cells. Plant Physiol. Biochem. 2006, 44, 581–589. [Google Scholar] [CrossRef]
- Yakimova, E.T.; Kapchina-Toteva, V.M.; Woltering, E.J. Signal transduction events in aluminum-induced cell death in tomato suspension cells. J. Plant Physiol. 2007, 164, 702–708. [Google Scholar]
- Iakimova, E.T.; Woltering, E.J.; Kapchina-Toteva, V.M.; Harren, F.J.M.; Cristescu, S.M. Cadmium toxicity in cultured tomato cells—Role of ethylene, proteases and oxidative stress in cell death signaling. Cell Biol. Int. 2008, 32, 1521–1529. [Google Scholar] [CrossRef]
- Suarez, M.F.; Filonova, L.H.; Smertenko, A.; Savenkov, E.I.; Clapham, D.H.; von Arnold, S.; Zhivotovsky, B.; Bozhkov, P.V. Metacaspase-dependent programmed cell death is essential for plant embryogenesis. Curr. Biol. 2004, 14, R339–R340. [Google Scholar]
- Bonneau, L.; Ge, Y.; Drury, G.E.; Gallois, P. What happened to plant caspases? J. Exp. Bot. 2008, 59, 491–499. [Google Scholar] [CrossRef]
- Rotari, V.I.; He, R.; Gallois, P. Death by proteases in plants: Whodunit. Physiol. Plant. 2005, 123, 376–385. [Google Scholar]
- Elbaz, M.; Avni, A.; Weil, M. Constitutive caspase-like machinery executes programmed cell death in plant cells. Cell Death Differ. 2002, 9, 726–733. [Google Scholar] [CrossRef]
- Mlejnek, P.; Procházka, S. Activation of caspase-like proteases and induction of apoptosis by isopentenyladenosine in tobacco BY-2 cells. Planta 2002, 215, 158–166. [Google Scholar]
- Kisselev, A.F.; Garcia-Calvo, M.; Overkleeft, H.S.; Peterson, E.; Pennington, M.W.; Ploegh, H.L.; Thornberry, N.A.; Goldberg, A.L. The caspase-like sites of proteasomes, their substrate specificity, new inhibitors and substrates, and allosteric interactions with the trypsin-like sites. J. Biol. Chem. 2003, 278, 35869–35877. [Google Scholar]
- Hatsugai, N.; Iwasaki, S.; Tamura, K.; Kondo, M.; Fuji, K.; Ogasawara, K.; Nishimura, M.; Nishimura, I.H. A novel membrane fusion-mediated plant immunity against bacterial pathogens. Genes Dev. 2009, 23, 2496–2506. [Google Scholar] [CrossRef]
- Solomon, M.; Belenghi, B.; Delledonne, M.; Menachem, E.; Levine, A. The involvement of cysteine proteases and protease inhibitor genes in the regulation of programmed cell death in plants. Plant Cell 1999, 11, 431–443. [Google Scholar]
- Gechev, T.S.; van Breusege, F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays 2006, 28, 1091–1101. [Google Scholar]
- Yokouchi, M.; Hiramatsu, N.; Hayakawa, K.; Kasai, A.; Takano, Y.; Yao, J.; Kitamura, M. Atypical, bidirectional regulation of cadmium-induced apoptosis via distinct signaling of unfolded protein response. Cell Death Differ. 2007, 14, 1467–1474. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, W.; Li, B.; Wang, Y.; Li, K.; Sodmergen; Han, C.; Zhang, Y.; Li, X. An endoplasmic reticulum response pathway mediates programmed cell death of root tip induced by water stress in Arabidopsis. New Phytol. 2010, 186, 681–695. [Google Scholar] [CrossRef]
- Cacas, J.L. Devil inside: Does plant programmed cell death involve the endomembrane system? Plant Cell Environ. 2010, 33, 1453–1473. [Google Scholar]
- Schröder, M.; Kaufman, R.J. ER stress and the unfolded protein response. Mutat. Res. 2005, 569, 29–63. [Google Scholar]
- Iwata, Y.; Koizumi, N. Unfolded protein response followed by induction of cell death in cultured tobacco cells treated with tunicamycin. Planta 2005, 220, 804–807. [Google Scholar] [CrossRef]
- Urade, R. The endoplasmic reticulum stress signalling pathways in plants. BioFactors 2009, 35, 326–331. [Google Scholar]
- Mendel, R.R.; Kruse, T. Cell biology of molybdenum in plants and humans. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 1568–1579. [Google Scholar] [CrossRef]
- Schwarz, G.; Schulze, J.; Bittner, F.; Eilers, T.; Kuper, J.; Bollmann, G.; Nerlich, A.; Brinkmann, H.; Mendel, R.R. The molybdenum cofactor biosynthetic protein Cnx1 complements molybdate-repairable mutants, transfers molybdenum to the metal binding pterin, and is associated with the cytoskeleton. Plant Cell 2000, 12, 2455–2471. [Google Scholar]
- Kirsch, J.; Betz, H. Widespread expression of gephyrin, a putative glycine receptor-tubulin linker protein, in rat brain. Brain Res. 1993, 621, 301–310. [Google Scholar] [CrossRef]
- Adamakis, I.D.S.; Panteris, E.; Eleftheriou, E.P. Tungsten effects on the actin microfilament network of meristematic cells of Zea mays L. In Proceedings of the 34th Scientific Conference of Hellenic Association for Biological Sciences, Trikala, Greece, 17–19 May, 2012; p. 3.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Adamakis, I.-D.S.; Panteris, E.; Eleftheriou, E.P. Tungsten Toxicity in Plants. Plants 2012, 1, 82-99. https://doi.org/10.3390/plants1020082
AMA Style
Adamakis I-DS, Panteris E, Eleftheriou EP. Tungsten Toxicity in Plants. Plants. 2012; 1(2):82-99. https://doi.org/10.3390/plants1020082
Chicago/Turabian StyleAdamakis, Ioannis-Dimosthenis S., Emmanuel Panteris, and Eleftherios P. Eleftheriou. 2012. "Tungsten Toxicity in Plants" Plants 1, no. 2: 82-99. https://doi.org/10.3390/plants1020082