Calmodulin Gene Expression in Response to Mechanical Wounding and Botrytis cinerea Infection in Tomato Fruit

Abstract
:1. Introduction
2. Results
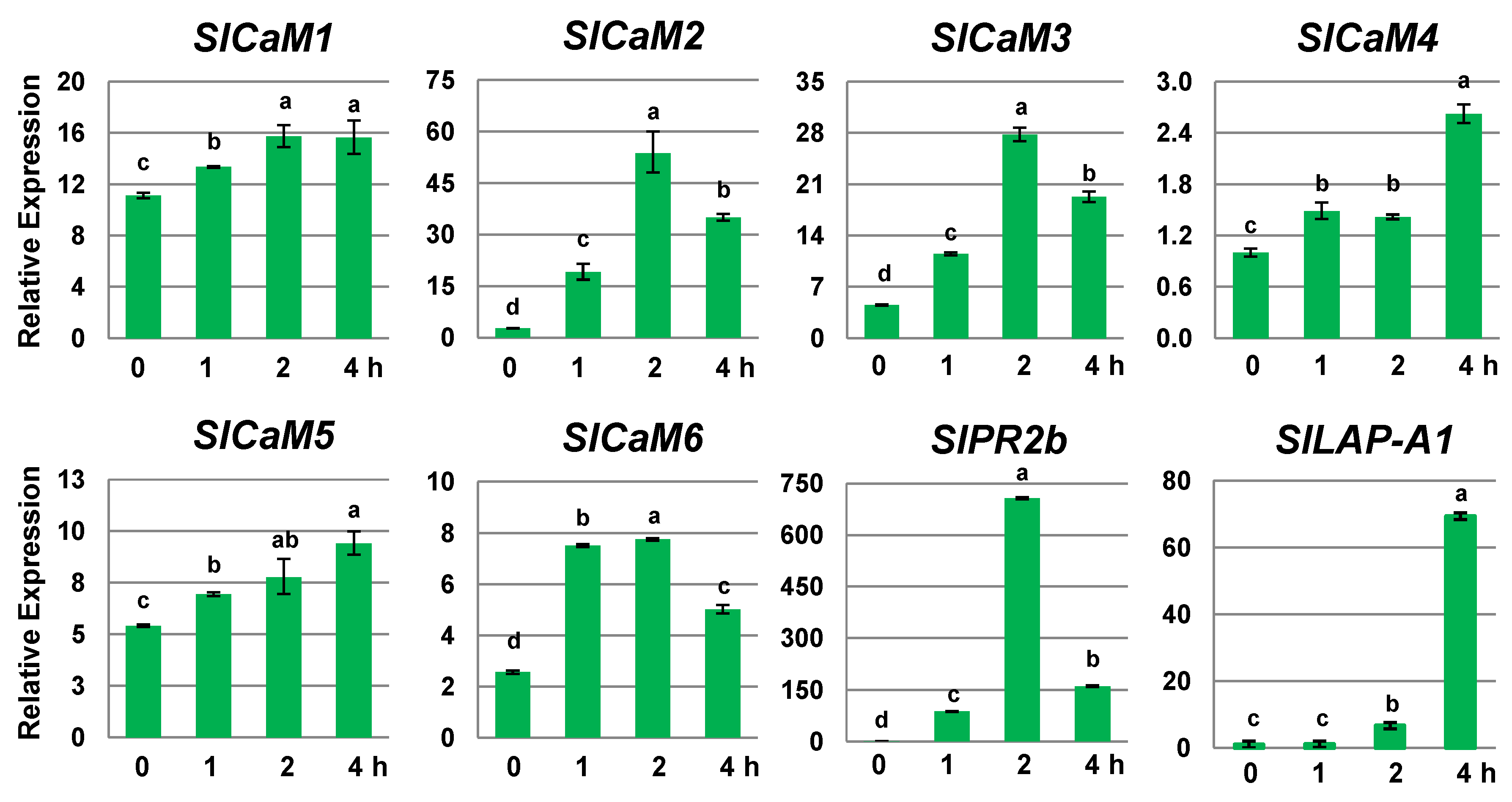
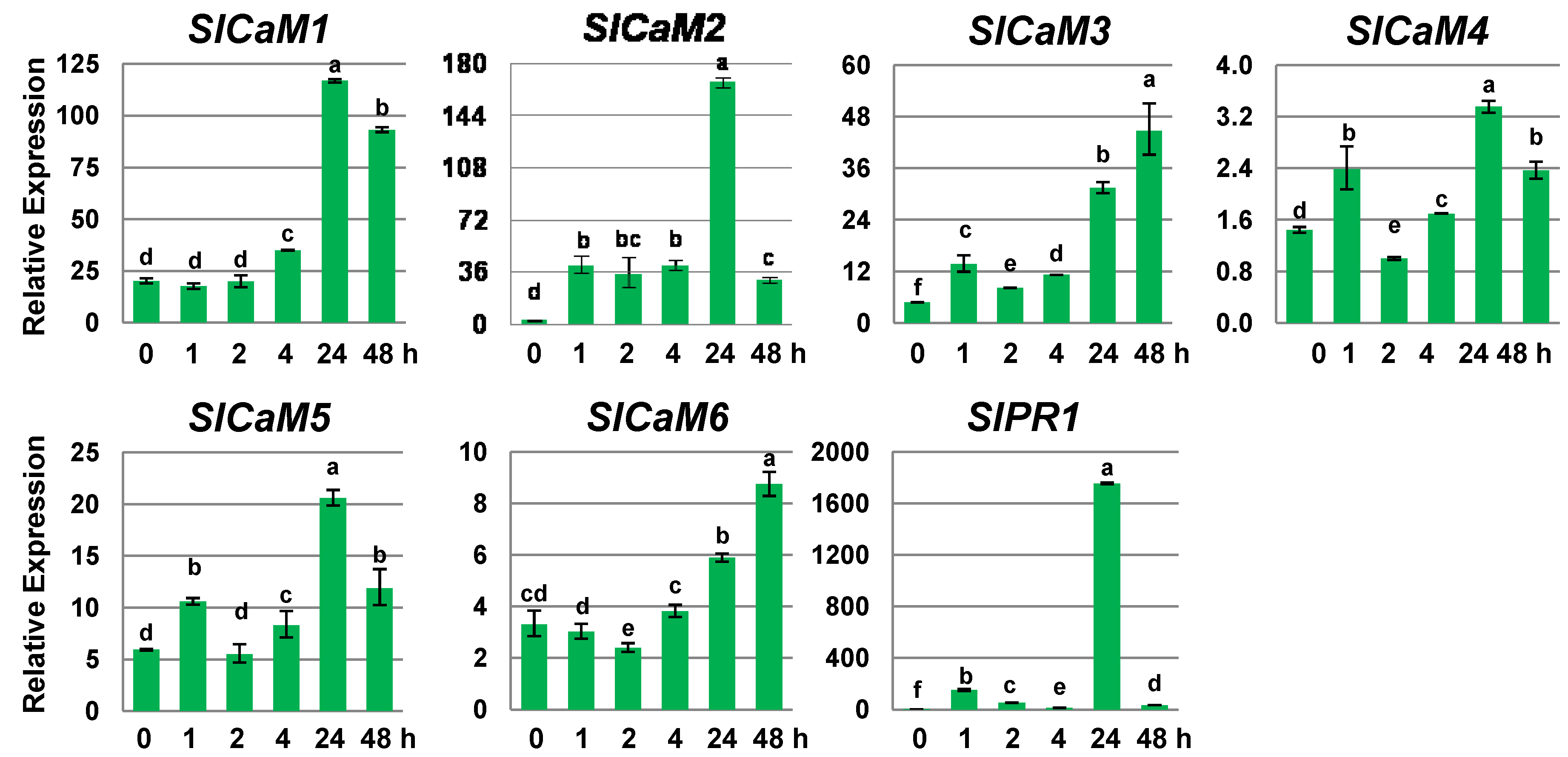
2.1. SlCaMs Are Responsive to Mechanical Wounding

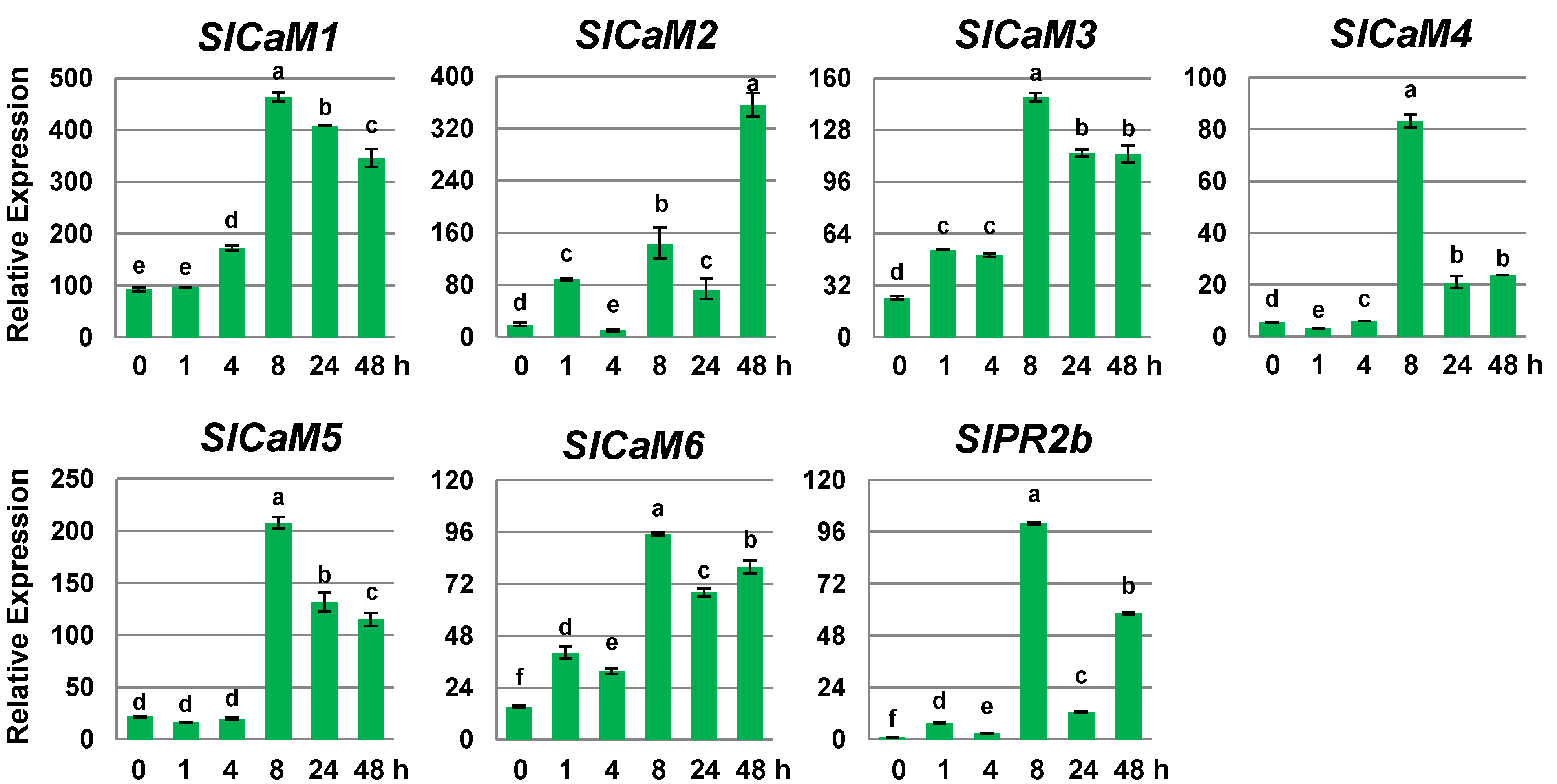
2.2. Pathogen Infection Triggers Calmodulin Gene Expression

2.3. Calmodulin Genes Are Salicylic Acid-Responsive

2.4. Methyl Jasmonate Stimulates Calmodulin Gene Expression

2.5. Overexpression of the SlCaM2 Gene Reduces Symptoms Incited by Botrytis Cinerea

3. Discussion
4. Experimental Section
4.1. Plant Materials
4.2. Mechanical Injury, Methyl Jasmonate and Salicylic Acid Treatments
4.3. Pathogen Infection and Decay Assay
4.4. RNA Extraction and RT-qPCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Oligonucleotides | Gene ID |
|---|---|---|
| SlCaM1-a | CCAGAGTTCCTTAACCTGATGG | Solyc01g008950 |
| SlCaM1-b | CTTTTCGCCTAGGTTTGTCATC | |
| SlCaM2-a | TCTGAGGAGGAGTTGAAAGAGG | Solyc10g081170 |
| SlCaM2-b | TCAACATCAGCTTCCCTAATCA | |
| SlCaM3-a | GATGGTAATGGAACCATCGACT | Solyc10g077010 |
| SlCaM3-b | CATCAGTGAGCTTCTCACCAAG | |
| SlCaM4-a | TCAGATCTCGGAGTTCAAAGAAG | Solyc11g072240 |
| SlCaM4-b | CAGGTTAAGGAACTCAGGGAAGT | |
| SlCaM5-a | TTAACTTGATGGCTCGGAAGAT | Solyc12g099990 |
| SlCaM5-b | ACGAATCATCTCGTCAACCTCT | |
| SlCaM6-a | ATCACTTGGTCAGAATCCCACT | Solyc03g098050 |
| SlCaM6-b | AGCTGCAGAAATAAAGCCATTC | |
| SlPR1-a | CTGTGAAGATGTGGGTTGATGAG | NM-001247429 |
| SlPR1-b | TCTCCAGTTACCTGGTGGATCAT | |
| SlPR2b-a | TCTTGCCCCATTTCAAGTTC | M80608 |
| SlPR2b-b | TGCACGTGTATCCCTCAAAA | |
| SlLAPA1-a | TGTCGCAGCATGTGAAAATATG | Solyc12g010020 |
| SlLAPA1-b | AGCACCAGTTAATGTTGCCAGA | |
| SlActin-a | GAAATAGCATAAGATGGCAGACG | X55749 |
| SlActin-b | ATACCCACCATCACACCAGTAT | |
| SlCaM2-S1 * | ggtggtaccATGGCGGATCAGCTGACGG | |
| SlCaM2-S2 * | ggaggatccCTTGGCCATCATGACCTTAAC | |
| SlCaM2-A1 * | ggtggtaccCTTGGCCATCATGACCTTAAC | |
| SlCaM2-A2 * | ggaggatccATGGCGGATCAGCTGACGG |
4.5. Construction of Ti Plasmids Carrying Sense- and Antisense-SlCaM2 Gene
4.6. Fruit Agroinjection
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol. 2010, 61, 593–620. [Google Scholar] [CrossRef] [PubMed]
- Batistic, O.; Kudla, J. Analysis of calcium signaling pathways in plants. BBA Gen Subj. 2012, 1820, 1283–1293. [Google Scholar] [CrossRef]
- DeFalco, T.A.; Bender, K.W.; Snedden, W.A. Breaking the code: Ca2+ sensors in plant signalling. Biochem. J. 2010, 425, 27–40. [Google Scholar] [CrossRef]
- Reddy, A.S.N.; Ali, G.S.; Celesnik, H.; Day, I.S. Coping with stresses: Roles of calcium- and calcium/calmodulin-regulated gene expression. Plant Cell 2011, 23, 2010–2032. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Poovaiah, B.W. Calcium/calmodulin-mediated signal network in plants. Trends Plant Sci. 2003, 8, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Bouche, N.; Yellin, A.; Snedden, W.A.; Fromm, H. Plant-specific calmodulin-binding proteins. Annu. Rev. Plant Biol. 2005, 56, 435–466. [Google Scholar] [CrossRef] [PubMed]
- Landoni, M.; de Francesco, A.; Galbiati, M.; Tonelli, C. A loss-of-function mutation in calmodulin2 gene affects pollen germination in Arabidopsis thaliana. Plant Mol. Biol. 2010, 74, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhou, R.G.; Gao, Y.J.; Zheng, S.Z.; Xu, P.; Zhang, S.Q.; Sun, D.Y. Molecular and genetic evidence for the key role of AtCaM3 in heat-shock signal transduction in Arabidopsis. Plant Physiol. 2009, 149, 1773–1784. [Google Scholar] [CrossRef] [PubMed]
- Heo, W.D.; Lee, S.H.; Kim, M.C.; Kim, J.C.; Chung, W.S.; Chun, H.J.; Lee, K.J.; Park, C.Y.; Park, H.C.; Choi, J.Y.; et al. Involvement of specific calmodulin isoforms in salicylic acid-independent activation of plant disease resistance responses. Proc. Natl. Acad. Sci. USA 1999, 96, 766–771. [Google Scholar] [CrossRef] [PubMed]
- Hodges, R.J.; Buzby, J.C.; Bennett, B. Postharvest losses and waste in developed and less developed countries: Opportunities to improve resource use. J. Agric. Sci. 2011, 149, 37–45. [Google Scholar] [CrossRef]
- Bergey, D.R.; Ryan, C.A. Wound- and systemin-inducible calmodulin gene expression in tomato leaves. Plant Mol. Biol. 1999, 40, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, W.; Xu, Y.P.; Cao, J.Y.; Braam, J.; Cai, X.Z. Genome-wide identification and functional analyses of calmodulin genes in Solanaceous species. BMC Plant Biol. 2013, 13. [Google Scholar] [CrossRef]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar]
- Chao, W.S.; Gu, Y.Q.; Pautot, V.; Bray, E.A.; Walling, L.L. Leucine aminopeptidase RNAs, proteins, and activities increase in response to water deficit, salinity, and the wound signals systemin, methyl jasmonate, and abscisic acid. Plant Physiol. 1999, 120, 979–992. [Google Scholar] [CrossRef] [PubMed]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [PubMed]
- Asghari, M.; Aghdam, M.S. Impact of salicylic acid on post-harvest physiology of horticultural crops. Trends Food Sci. Technol. 2010, 21, 502–509. [Google Scholar] [CrossRef]
- Tornero, P.; Gadea, J.; Conejero, V.; Vera, P. Two PR-1 genes from tomato are differentially regulated and reveal a novel mode of expression for a pathogenesis-related gene during the hypersensitive response and development. Mol. Plant Microbe Interact. 1997, 10, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Uquillas, C.; Letelier, I.; Blanco, F.; Jordana, X.; Holuigue, L. NPR1-independent activation of immediate early salicylic acid-responsive genes in Arabidopsis. Mol. Plant Microbe Interact. 2004, 17, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar]
- Rohwer, C.L.; Erwin, J.E. Jasmonate-induced changes in polyphenol oxidase, peroxidase, and proteinase inhibitors in horticultural species. Hortscience 2006, 41, 1072–1072. [Google Scholar]
- Yang, T.B.; Peng, H.; Whitaker, B.D.; Jurick, W.M. Differential expression of calcium/calmodulin-regulated SlSRs in response to abiotic and biotic stresses in tomato fruit. Physiol. Plantarum. 2013, 148, 445–455. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. Jasmonate signaling: Toward an integrated view. Plant Physiol. 2008, 146, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Saftner, R.A.; Bai, J.H.; Abbott, J.A.; Lee, Y.S. Sanitary dips with calcium propionate, calcium chloride, or a calcium amino acid chelate maintain quality and shelf stability of fresh-cut honeydew chunks. Postharvest. Biol. Technol. 2003, 29, 257–269. [Google Scholar] [CrossRef]
- Abbott, J.A.; Conway, W.S.; Sams, C.E. Postharvest calcium-chloride infiltration affects textural attributes of apples. J. Am. Soc. Hortic. Sci. 1989, 114, 932–936. [Google Scholar]
- Park, S.; Cheng, N.H.; Pittman, J.K.; Yoo, K.S.; Park, J.; Smith, R.H.; Hirschi, K.D. Increased calcium levels and prolonged shelf life in tomatoes expressing Arabidopsis H+/Ca2+ transporters. Plant Physiol. 2005, 139, 1194–1206. [Google Scholar] [CrossRef] [PubMed]
- Ritenour, M.A.; Stoffella, P.J.; He, Z.L.; Narciso, J.A.; Salvatore, J.J. Postharvest calcium chloride dips of whole tomato fruit reduce postharvest decay under commercial conditions. Hortscience 2006, 41, 1016–1017. [Google Scholar]
- Saftner, R.A.; Conway, W.S.; Sams, C.E. Postharvest calcium infiltration alone and combined with surface coating treatments influence volatile levels, respiration, ethylene production, and internal atmospheres of “Golden Delicious” apples. J. Am. Soc. Hortic. Sci. 1999, 124, 553–558. [Google Scholar]
- Poovaiah, B.W.; Shekhar, V.C. Effects of calcium infiltration of golden delicious apples on fruit firmness and senescence. Hortscience 1978, 13, 357–357. [Google Scholar]
- Sams, C.E.; Conway, W.S.; Abbott, J.A.; Lewis, R.J.; Benshalom, N. Firmness and decay of apples following postharvest pressure infiltration of calcium and heat-treatment. J. Am. Soc. Hortic. Sci. 1993, 118, 623–627. [Google Scholar]
- Conway, W.S.; Sams, C.E. The effects of postharvest infiltration of calcium, magnesium, or strontium on decay, firmness, respiration, and ethylene production in apples. J. Am. Soc. Hortic. Sci. 1987, 112, 300–303. [Google Scholar]
- Lecourieux, D.; Raneva, R.; Pugin, A. Calcium in plant defence-signalling pathways. New Phytol. 2006, 171, 249–269. [Google Scholar] [CrossRef] [PubMed]
- Nimchuk, Z.; Eulgem, T.; Holt, B.F., 3rd; Dangl, J.L. Recognition and response in the plant immune system. Annu. Rev. Genet. 2003, 37, 579–609. [Google Scholar]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: Turning local infection into global defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.J.; Kenton, P.; Atzorn, R.; Miersch, O.; Wasternack, C. The outcomes of concentration-specific interactions between salicylate and jasmonate signaling include synergy, antagonism, and oxidative stress leading to cell death. Plant Physiol. 2006, 140, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, A.; Leon-Reyes, A.; Ritsema, T.; Verhage, A.; den Otter, F.C.; van Loon, L.C.; Pieterse, C.M.J. Kinetics of salicylate-mediated suppression of jasmonate signaling reveal a role for redox modulation. Plant Physiol. 2008, 147, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Abd el Rahman, T.; el Oirdi, M.; Gonzalez-Lamothe, R.; Bouarab, K. Necrotrophic pathogens use the salicylic acid signaling pathway to promote disease development in tomato. Mol. Plant Microbe Interact. 2012, 25, 1584–1593. [Google Scholar]
- El Oirdi, M.; Abd el Rahman, T.; Rigano, L.; el Hadrami, A.; Rodriguez, M.C.; Daayf, F.; Vojnov, A.; Bouarab, K. Botrytis cinerea manipulates the antagonistic effects between immune pathways to promote disease development in tomato. Plant Cell 2011, 23, 2405–2421. [Google Scholar]
- Wang, L.; Tsuda, K.; Truman, W.; Sato, M.; Nguyen, L.V.; Katagiri, F.; Glazebrook, J. CBP60g and SARD1 play partially redundant critical roles in salicylic acid signaling. Plant J. 2011, 67, 1029–1041. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, S.; Ding, P.; Wang, D.; Cheng, Y.T.; He, J.; Gao, M.; Xu, F.; Li, Y.; Zhu, Z.; et al. Control of salicylic acid synthesis and systemic acquired resistance by two members of a plant-specific family of transcription factors. Proc. Natl. Acad. Sci. USA 2010, 107, 18220–18225. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tsuda, K.; Sato, M.; Cohen, J.D.; Katagiri, F.; Glazebrook, J. Arabidopsis CaM binding protein CBP60g contributes to MAMP-induced SA accumulation and is involved in disease resistance against Pseudomonas syringae. PLoS Pathogens 2009, 5, e1000301. [Google Scholar] [CrossRef] [PubMed]
- Du, L.Q.; Ali, G.S.; Simons, K.A.; Hou, J.G.; Yang, T.B.; Reddy, A.S.N.; Poovaiah, B.W. Ca2+/calmodulin regulates salicylic-acid-mediated plant immunity. Nature 2009, 457, 1154–1158. [Google Scholar] [CrossRef] [PubMed]
- Poovaiah, B.W.; Du, L.Q.; Wang, H.Z.; Yang, T.B. Recent advances in calcium/calmodulin-mediated signaling with an emphasis on plant-microbe interactions. Plant Physiol. 2013, 163, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Leon, J.; Rojo, E.; Titarenko, E.; Sanchez-Serrano, J.J. Jasmonic acid-dependent and -independent wound signal transduction pathways are differentially regulated by Ca2+/calmodulin in Arabidopsis thaliana. Mol. Genet. Gen. 1998, 258, 412–419. [Google Scholar] [CrossRef]
- Dombrowski, J.E.; Bergey, D.R. Calcium ions enhance systemin activity and play an integral role in the wound response. Plant Sci. 2007, 172, 335–344. [Google Scholar] [CrossRef]
- Qiu, Y.J.; Xi, J.; Du, L.Q.; Suttle, J.C.; Poovaiah, B.W. Coupling calcium/calmodulin-mediated signaling and herbivore-induced plant response through calmodulin-binding transcription factor AtSR1/CAMTA3. Plant Mol. Biol. 2012, 79, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Laluk, K.; Prasad, K.V.S.K.; Savchenko, T.; Celesnik, H.; Dehesh, K.; Levy, M.; Mitchell-Olds, T.; Reddy, A.S.N. The calmodulin-binding transcription factor signal responsive1 is a novel regulator of glucosinolate metabolism and herbivory tolerance in Arabidopsis. Plant Cell Physiol. 2012, 53, 2008–2015. [Google Scholar] [CrossRef] [PubMed]
- Galon, Y.; Nave, R.; Boyce, J.M.; Nachmias, D.; Knight, M.R.; Fromm, H. Calmodulin-binding transcription activator (CAMTA) 3 mediates biotic defense responses in Arabidopsis. FEBS Lett. 2008, 582, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Sargent, S.A.; Moretti, C.L. Tomato. In Agricultural Handbook Number 66 (HB66); Draft-Updated August 2014; USDA-ARS: USA. Available online: http://www.ba.ars.usda.gov/hb66/tomato.pdf (accessed on 11 August 2014).
- Yan, H.J.; Jurick, W.M., II; Lou, Y.G.; Gaskins, V.L.; Kim, Y.-K. First report of pyrimethanil resistance in Botrytis cinerea from stored apples in pennsylvania. Plant Dis. 2014, 98, 999. [Google Scholar]
- Untergrasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer 3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Orzaez, D.; Mirabel, S.; Wieland, W.H.; Granell, A. Agroinjection of tomato fruits. A tool for rapid functional analysis of transgenes directly in fruit. Plant Physiol. 2006, 140, 3–11. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Peng, H.; Yang, T.; II, W.M.J. Calmodulin Gene Expression in Response to Mechanical Wounding and Botrytis cinerea Infection in Tomato Fruit. Plants 2014, 3, 427-441. https://doi.org/10.3390/plants3030427
Peng H, Yang T, II WMJ. Calmodulin Gene Expression in Response to Mechanical Wounding and Botrytis cinerea Infection in Tomato Fruit. Plants. 2014; 3(3):427-441. https://doi.org/10.3390/plants3030427
Chicago/Turabian StylePeng, Hui, Tianbao Yang, and Wayne M. Jurick II. 2014. "Calmodulin Gene Expression in Response to Mechanical Wounding and Botrytis cinerea Infection in Tomato Fruit" Plants 3, no. 3: 427-441. https://doi.org/10.3390/plants3030427