Live Cell Imaging During Germination Reveals Dynamic Tubular Structures Derived from Protein Storage Vacuoles of Barley Aleurone Cells

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
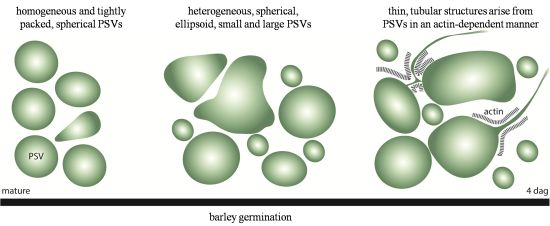
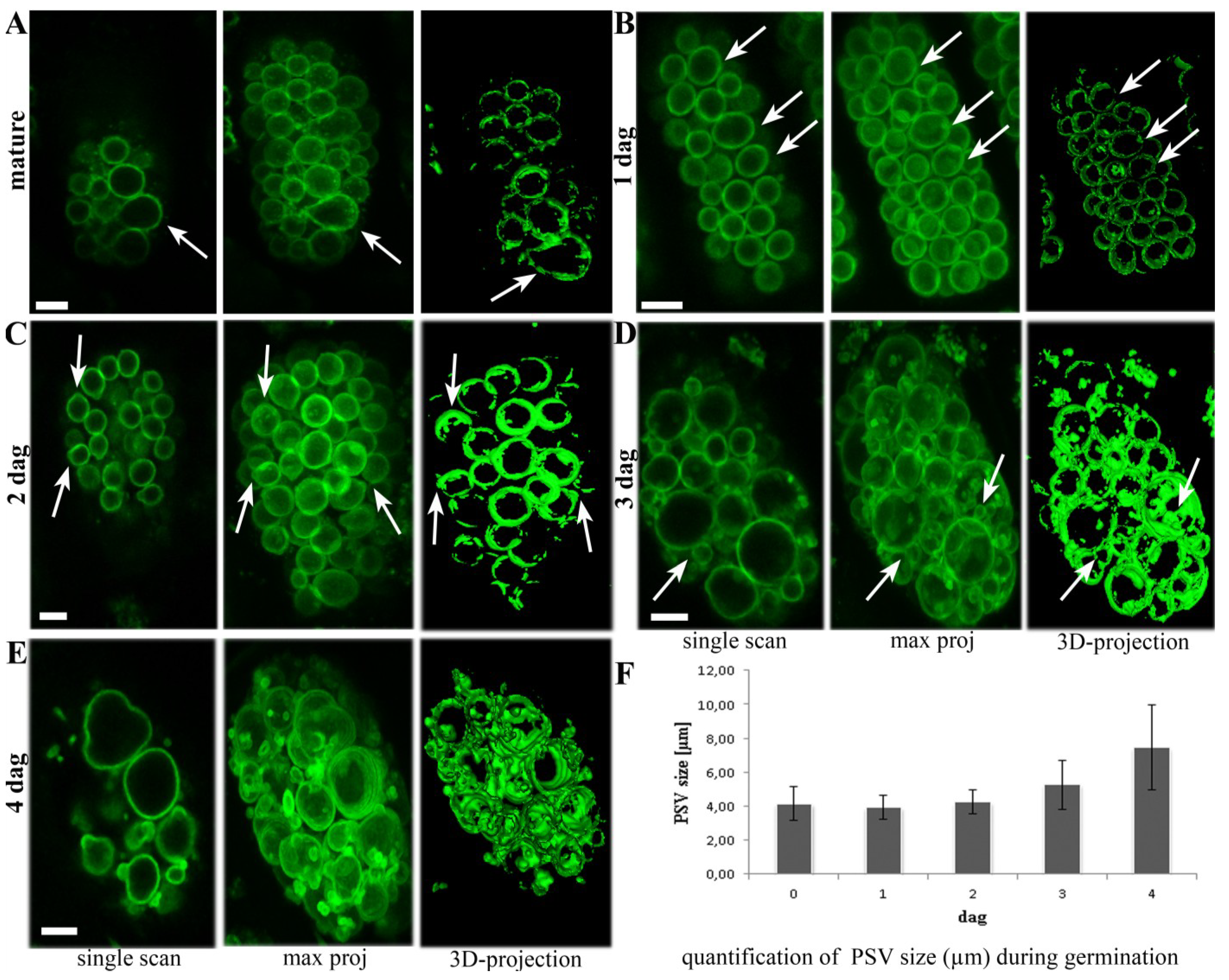
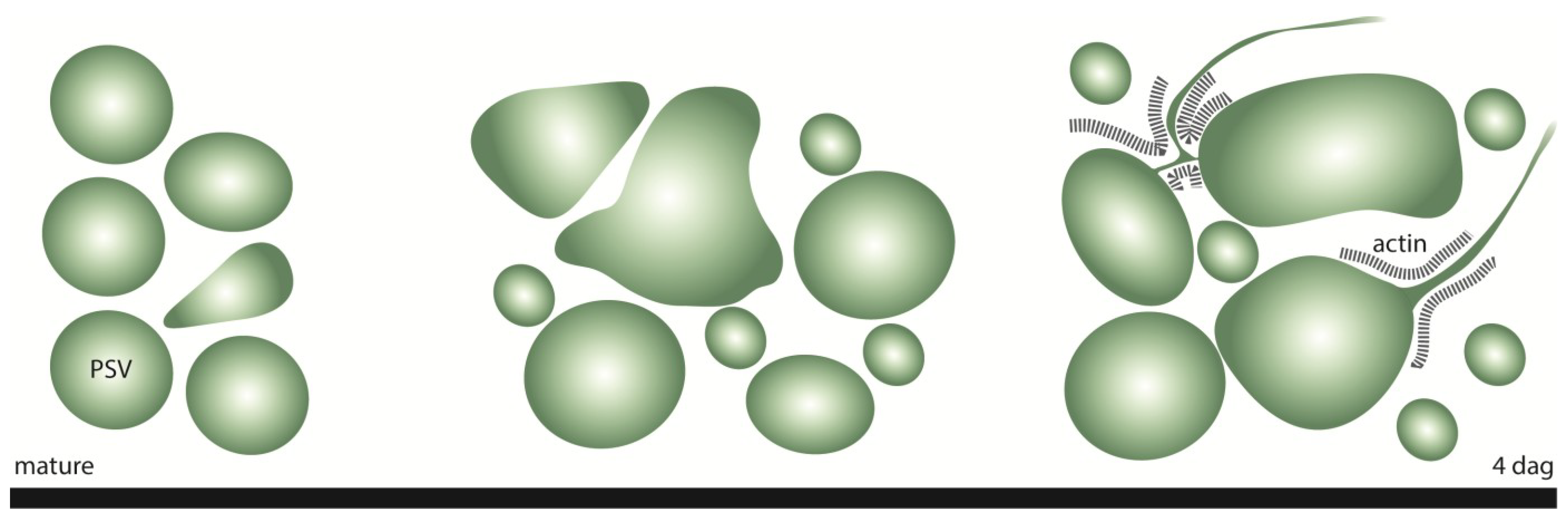
2.1. Aleurone PSVs Become Larger and More Ellipsoid during Germination

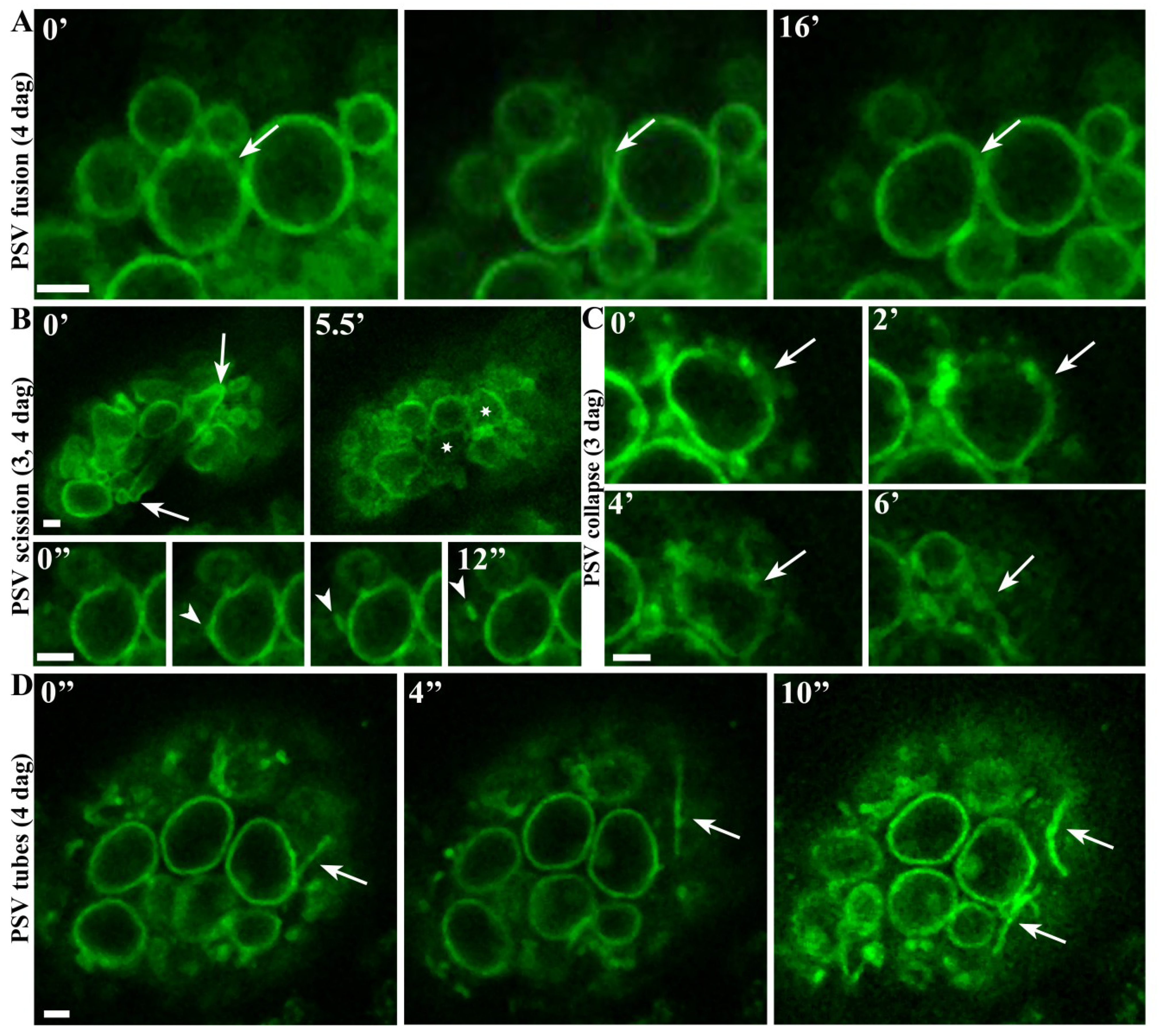
2.2. Real-Time Observations Indicate that Aleurone PSVs may Undergo Fusion, Scission and Collapse
2.3. Highly Mobile Tubular Extension Structures Originate from PSVs


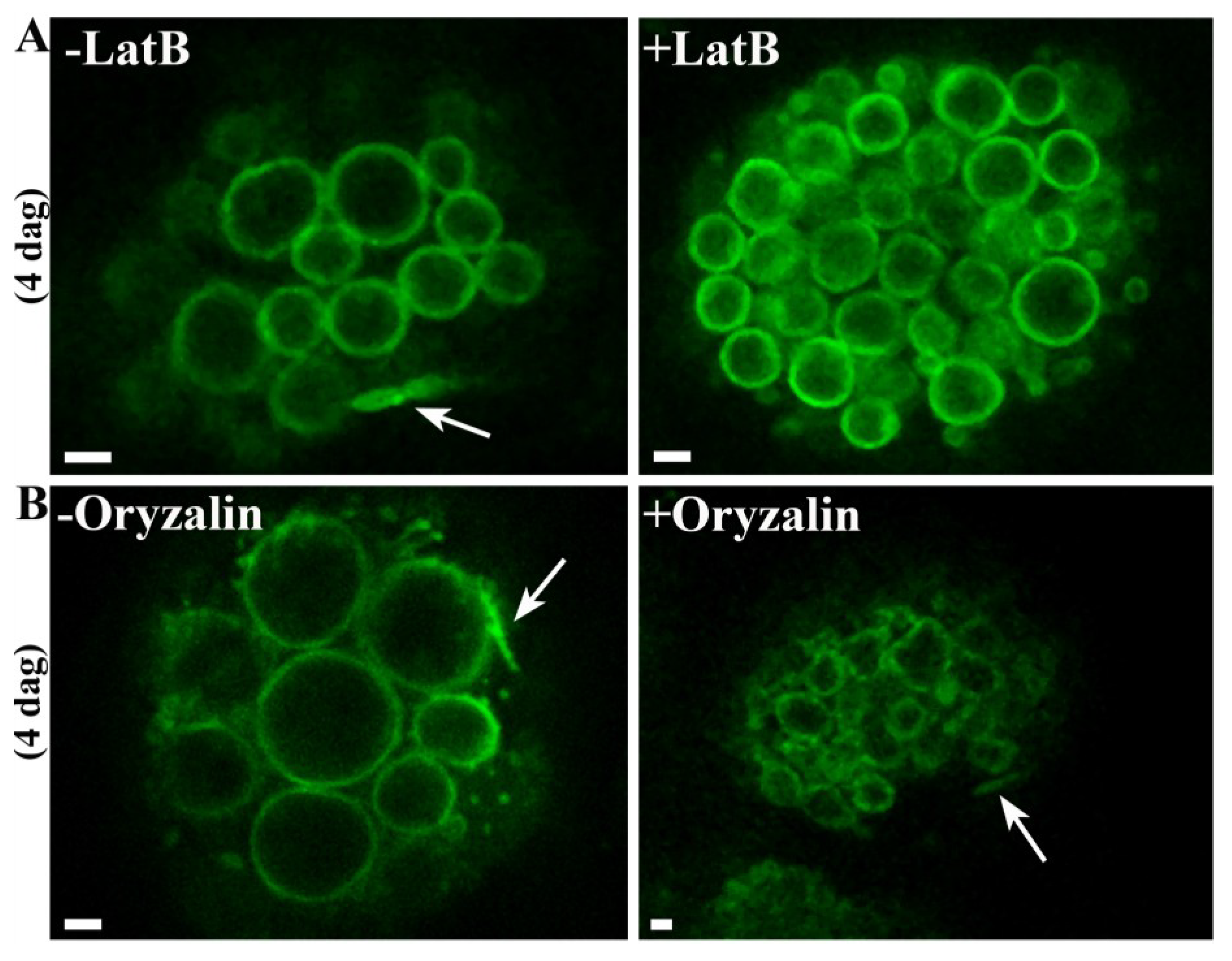
2.4. The Formation of PSV Tubules Depends on Microfilaments but not Microtubules

3. Discussion

4. Experimental
4.1. Plant Material
4.2. Confocal Microcopy
4.3. Calculation of PSV Size
4.4. Inhibitor Studies
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar] [CrossRef]
- Galili, G. Er-derived compartments are formed by highly regulated processes and have special functions in plants. Plant Physiol. 2004, 136, 3411–3413. [Google Scholar] [CrossRef]
- Bassham, D.C.; Raikhel, N.V. Unique features of the plant vacuolar sorting machinery. Curr. Opin. Cell Biol. 2000, 12, 491–495. [Google Scholar] [CrossRef]
- Hunter, P.R.; Craddock, C.P.; Di Benedetto, S.; Roberts, L.M.; Frigerio, L. Fluorescent reporter proteins for the tonoplast and the vacuolar lumen identify a single vacuolar compartment in arabidopsis cells. Plant Physiol. 2007, 145, 1371–1382. [Google Scholar] [CrossRef]
- Herman, E.M.; Larkins, B.A. Protein storage bodies and vacuoles. Plant Cell 1999, 11, 601–614. [Google Scholar] [CrossRef]
- Jiang, L.; Phillips, T.E.; Hamm, C.A.; Drozdowicz, Y.M.; Rea, P.A.; Maeshima, M.; Rogers, S.W.; Rogers, J.C. The protein storage vacuole: A unique compound organelle. J. Cell Biol. 2001, 155, 991–1002. [Google Scholar] [CrossRef]
- Tosi, P.; Parker, M.; Gritsch, C.S.; Carzaniga, R.; Martin, B.; Shewry, P.R. Trafficking of storage proteins in developing grain of wheat. J. Exp. Bot. 2009, 60, 979–991. [Google Scholar] [CrossRef]
- Neuhaus, J.M.; Rogers, J.C. Sorting of proteins to vacuoles in plant cells. Plant Mol. Biol. 1998, 38, 127–144. [Google Scholar] [CrossRef]
- Martinoia, E.; Maeshima, M.; Neuhaus, H.E. Vacuolar transporters and their essential role in plant metabolism. J. Exp. Bot. 2007, 58, 83–102. [Google Scholar] [CrossRef]
- Jauh, G.Y.; Phillips, T.E.; Rogers, J.C. Tonoplast intrinsic protein isoforms as markers for vacuolar functions. Plant Cell 1999, 11, 1867–1882. [Google Scholar]
- Zheng, H.; Staehelin, L.A. Protein storage vacuoles are transformed into lytic vacuoles in root meristematic cells of germinating seedlings by multiple, cell type-specific mechanisms. Plant Physiol. 2011, 155, 2023–2035. [Google Scholar] [CrossRef]
- Bethke, P.C.; Swanson, S.J.; Hillmer, S.; Jones, R.L. From storage compartment to lytic organelle: The metamorphosis of the aleurone protein storage vacuole. Ann. Bot. 1998, 82, 399–412. [Google Scholar] [CrossRef]
- Fath, A.; Bethke, P.; Lonsdale, J.; Meza-Romero, R.; Jones, R. Programmed cell death in cereal aleurone. Plant Mol. Biol. 2000, 44, 255–266. [Google Scholar] [CrossRef]
- Muntz, K. Protein dynamics and proteolysis in plant vacuoles. J. Exp. Bot. 2007, 58, 2391–2407. [Google Scholar] [CrossRef]
- Reyes, F.C.; Chung, T.; Holding, D.; Jung, R.; Vierstra, R.; Otegui, M.S. Delivery of prolamins to the protein storage vacuole in maize aleurone cells. Plant Cell 2011, 23, 769–784. [Google Scholar] [CrossRef]
- Jacobsen, J.V.; Knox, R.B.; Pyliotis, N.A. Structure and composition of aleurone grains in barley aleurone layer. Planta 1971, 101, 189–209. [Google Scholar] [CrossRef]
- Stewart, A.; Nield, H.; Lott, J.N. An investigation of the mineral content of barley grains and seedlings. Plant Physiol. 1988, 86, 93–97. [Google Scholar] [CrossRef]
- Ritchie, S.; Swanson, S.J.; Gilroy, S. Physiology of the aleurone layer and starchy endosperm during grain development and early seedling growth: New insights from cell and molecular biology. Seed Sci. Res. 2000, 10, 193–212. [Google Scholar]
- Sreenivasulu, N.; Usadel, B.; Winter, A.; Radchuk, V.; Scholz, U.; Stein, N.; Weschke, W.; Strickert, M.; Close, T.J.; Stitt, M.; et al. Barley grain maturation and germination: Metabolic pathway and regulatory network commonalities and differences highlighted by new mapman/pageman profiling tools. Plant Physiol. 2008, 146, 1738–1758. [Google Scholar] [CrossRef]
- Jacobsen, J.V.; Zwar, J.A.; Chandler, P.M. Gibberellic-acid-responsive protoplasts from mature aleurone of himalaya barley. Planta 1985, 163, 430–438. [Google Scholar] [CrossRef]
- Bethke, P.C.; Jones, R.L. Cell death of barley aleurone protoplasts is mediated by reactive oxygen species. Plant J. 2001, 25, 19–29. [Google Scholar] [CrossRef]
- Jones, R.L.; Jacobsen, J.V. Regulation of synthesis and transport of secreted proteins in cereal aleurone. Int. Rev. Cytol. 1991, 126, 49–88. [Google Scholar] [CrossRef]
- Fath, A.; Bethke, P.C.; Jones, R.L. Barley aleurone cell death is not apoptotic: Characterization of nuclease activities and DNA degradation. Plant J. 1999, 20, 305–315. [Google Scholar] [CrossRef]
- Bush, D.S.; Cornejo, M.J.; Huang, C.N.; Jones, R.L. Ca-stimulated secretion of alpha-amylase during development in barley aleurone protoplasts. Plant Physiol. 1986, 82, 566–574. [Google Scholar] [CrossRef]
- Swanson, S.J.; Jones, R.L. Gibberellic acid induces vacuolar acidification in barley aleurone. Plant Cell 1996, 8, 2211–2221. [Google Scholar] [CrossRef]
- Holwerda, B.C.; Rogers, J.C. Purification and characterization of aleurain: A plant thiol protease functionally homologous to mammalian cathepsin H. Plant Physiol. 1992, 99, 848–855. [Google Scholar] [CrossRef]
- Swanson, S.J.; Bethke, P.C.; Jones, R.L. Barley aleurone cells contain two types of vacuoles: Characterization of lytic organelles by use of fluorescent probes. Plant Cell 1998, 10, 685–698. [Google Scholar] [CrossRef]
- Hwang, Y.S.; Bethke, P.C.; Cheong, Y.H.; Chang, H.S.; Zhu, T.; Jones, R.L. A gibberellin-regulated calcineurin b in rice localizes to the tonoplast and is implicated in vacuole function. Plant Physiol. 2005, 138, 1347–1358. [Google Scholar] [CrossRef]
- Gattolin, S.; Sorieul, M.; Hunter, P.R.; Khonsari, R.H.; Frigerio, L. In vivo imaging of the tonoplast intrinsic protein family in arabidopsis roots. BMC Plant Biol. 2009, 9, e133. [Google Scholar] [CrossRef]
- Onda, Y.; Kumamaru, T.; Kawagoe, Y. Er membrane-localized oxidoreductase ero1 is required for disulfide bond formation in the rice endosperm. Proc. Natl. Acad. Sci. USA 2009, 106, 14156–14161. [Google Scholar]
- Johnson, K.D.; Herman, E.M.; Chrispeels, M.J. An abundant, highly conserved tonoplast protein in seeds. Plant Physiol. 1989, 91, 1006–1013. [Google Scholar] [CrossRef]
- Gattolin, S.; Sorieul, M.; Frigerio, L. Tonoplast intrinsic proteins and vacuolar identity. Biochem. Soc. Trans. 2010, 38, 769–773. [Google Scholar] [CrossRef]
- Hillmer, S.; Movafeghi, A.; Robinson, D.G.; Hinz, G. Vacuolar storage proteins are sorted in the cis-cisternae of the pea cotyledon golgi apparatus. J. Cell Biol. 2001, 152, 41–50. [Google Scholar] [CrossRef]
- Jiang, L.W.; Rogers, J.C. Sorting of membrane proteins to vacuoles in plant cells. Plant Sci. 1999, 146, 55–67. [Google Scholar] [CrossRef]
- Paris, N.; Stanley, C.M.; Jones, R.L.; Rogers, J.C. Plant cells contain two functionally distinct vacuolar compartments. Cell 1996, 85, 563–572. [Google Scholar]
- Ibl, V.; Kapusi, E.; Arcalis, E.; Kawagoe, Y.; Stoger, E. Fusion, rupture, and degeneration: The fate of in vivo-labelled psvs in developing barley endosperm. J. Exp. Bot. 2014, 65, 3249–3261. [Google Scholar] [CrossRef]
- Ibl, V.; Stoger, E. University of Natural Resources and Life Sciences. Vienna, Austria, Unpublished data. 2014. [Google Scholar]
- Sreenivasulu, N.; Radchuk, V.; Strickert, M.; Miersch, O.; Weschke, W.; Wobus, U. Gene expression patterns reveal tissue-specific signaling networks controlling programmed cell death and ABA-regulated maturation in developing barley seeds. Plant J. 2006, 47, 310–327. [Google Scholar] [CrossRef]
- Greenwood, J.S.; Helm, M.; Gietl, C. Ricinosomes and endosperm transfer cell structure in programmed cell death of the nucellus during ricinus seed development. Proc. Natl. Acad. Sci. USA 2005, 102, 2238–2243. [Google Scholar] [CrossRef]
- Guilliermond, A. The recent development of our idea of the vacuome of plant cells. Am. J. Bot. 1929, 16, 1–22. [Google Scholar]
- Palevitz, B.A.; Okane, D.J. Epifluorescence and video analysis of vacuole motility and development in stomatal cells of allium. Science 1981, 214, 443–445. [Google Scholar]
- Palevitz, B.A.; Okane, D.J.; Kobres, R.E.; Raikhel, N.V. The vacuole system in stomatal cells of allium—Vacuole movements and changes in morphology in differentiating cells as revealed by epifluorescence, video and electron-microscopy. Protoplasma 1981, 109, 23–55. [Google Scholar] [CrossRef]
- Ovecka, M.; Lang, I.; Baluska, F.; Ismail, A.; Illes, P.; Lichtscheidl, I.K. Endocytosis and vesicle trafficking during tip growth of root hairs. Protoplasma 2005, 226, 39–54. [Google Scholar] [CrossRef]
- Hicks, G.R.; Rojo, E.; Hong, S.; Carter, D.G.; Raikhel, N.V. Geminating pollen has tubular vacuoles, displays highly dynamic vacuole biogenesis, and requires vacuoless1 for proper function. Plant Physiol. 2004, 134, 1227–1239. [Google Scholar] [CrossRef]
- Wiltshire, E.J.; Collings, D.A. New dynamics in an old friend: Dynamic tubular vacuoles radiate through the cortical cytoplasm of red onion epidermal cells. Plant Cell Physiol. 2009, 50, 1826–1839. [Google Scholar] [CrossRef]
- Mathur, J.; Radhamony, R.; Sinclair, A.M.; Donoso, A.; Dunn, N.; Roach, E.; Radford, D.; Mohaghegh, P.S.; Logan, D.C.; Kokolic, K.; et al. Meosfp-based green-to-red photoconvertible subcellular probes for plants. Plant Physiol. 2010, 154, 1573–1587. [Google Scholar] [CrossRef]
- Natesan, S.K.; Sullivan, J.A.; Gray, J.C. Stromules: A characteristic cell-specific feature of plastid morphology. J. Exp. Bot. 2005, 56, 787–797. [Google Scholar]
- Kwok, E.Y.; Hanson, M.R. Stromules and the dynamic nature of plastid morphology. J. Microsc. 2004, 214, 124–137. [Google Scholar]
- Hanson, M.R.; Sattarzadeh, A. Stromules: Recent insights into a long neglected feature of plastid morphology and function. Plant Physiol. 2011, 155, 1486–1492. [Google Scholar]
- Hanson, M.R.; Sattarzadeh, A. Trafficking of proteins through plastid stromules. Plant Cell 2013, 25, 2774–2782. [Google Scholar] [CrossRef]
- Itoh, R.; Fujiwara, M.T. Regulation of leucoplast morphology in roots: Interorganellar signaling from mitochondria? Plant Signal. Behav. 2010, 5, 856–859. [Google Scholar] [CrossRef]
- Schattat, M.; Barton, K.; Mathur, J. Correlated behavior implicates stromules in increasing the interactive surface between plastids and er tubules. Plant Signal. Behav. 2011, 6, 715–718. [Google Scholar] [CrossRef]
- Lazzaro, M.D.; Thomson, W.W. The vacuolar-tubular continuum in living trichomes of chickpea (Cicer arietinum) provides a rapid means of solute delivery from base to tip. Protoplasma 1996, 193, 181–190. [Google Scholar] [CrossRef]
- Ashford, A.E.; Allaway, W.G. The role of the motile tubular vacuole system in mycorrhizal fungi. Plant Soil 2002, 244, 177–187. [Google Scholar] [CrossRef]
- Newell, J.M.; Leigh, R.A.; Hall, J.L. Vacuole development in cultured evacuolated oat mesophyll protoplasts. J. Exp. Bot. 1998, 49, 817–827. [Google Scholar] [CrossRef]
- Lovy-Wheeler, A.; Cardenas, L.; Kunkel, J.G.; Hepler, P.K. Differential organelle movement on the actin cytoskeleton in lily pollen tubes. Cell Motil. Cytoskeleton 2007, 64, 217–232. [Google Scholar] [CrossRef]
- Higaki, T.; Kutsuna, N.; Okubo, E.; Sano, T.; Hasezawa, S. Actin microfilaments regulate vacuolar structures and dynamics: Dual observation of actin microfilaments and vacuolar membrane in living tobacco by-2 cells. Plant Cell Physiol. 2006, 47, 839–852. [Google Scholar] [CrossRef]
- Kim, H.; Park, M.; Kim, S.J.; Hwang, I. Actin filaments play a critical role in vacuolar trafficking at the golgi complex in plant cells. Plant Cell 2005, 17, 888–902. [Google Scholar]
- Spector, I.; Shochet, N.R.; Kashman, Y.; Groweiss, A. Latrunculins: Novel marine toxins that disrupt microfilament organization in cultured cells. Science 1983, 219, 493–495. [Google Scholar]
- Sheahan, M.B.; Rose, R.J.; McCurdy, D.W. Actin-filament-dependent remodeling of the vacuole in cultured mesophyll protoplasts. Protoplasma 2007, 230, 141–152. [Google Scholar] [CrossRef]
- Uemura, T.; Yoshimura, S.H.; Takeyasu, K.; Sato, M.H. Vacuolar membrane dynamics revealed by gfp-atvam3 fusion protein. Genes Cells 2002, 7, 743–753. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ibl, V.; Stoger, E. Live Cell Imaging During Germination Reveals Dynamic Tubular Structures Derived from Protein Storage Vacuoles of Barley Aleurone Cells. Plants 2014, 3, 442-457. https://doi.org/10.3390/plants3030442
Ibl V, Stoger E. Live Cell Imaging During Germination Reveals Dynamic Tubular Structures Derived from Protein Storage Vacuoles of Barley Aleurone Cells. Plants. 2014; 3(3):442-457. https://doi.org/10.3390/plants3030442
Chicago/Turabian StyleIbl, Verena, and Eva Stoger. 2014. "Live Cell Imaging During Germination Reveals Dynamic Tubular Structures Derived from Protein Storage Vacuoles of Barley Aleurone Cells" Plants 3, no. 3: 442-457. https://doi.org/10.3390/plants3030442