Pistil Smut Infection Increases Ovary Production, Seed Yield Components, and Pseudosexual Reproductive Allocation in Buffalograss

Abstract
:
1. Introduction
2. Results and Discussion
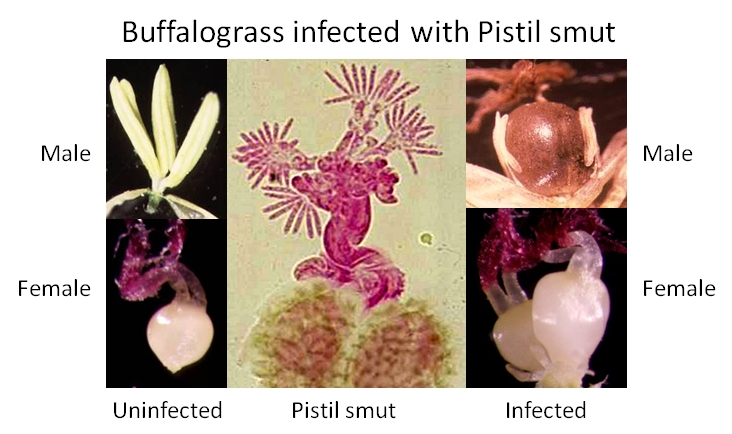
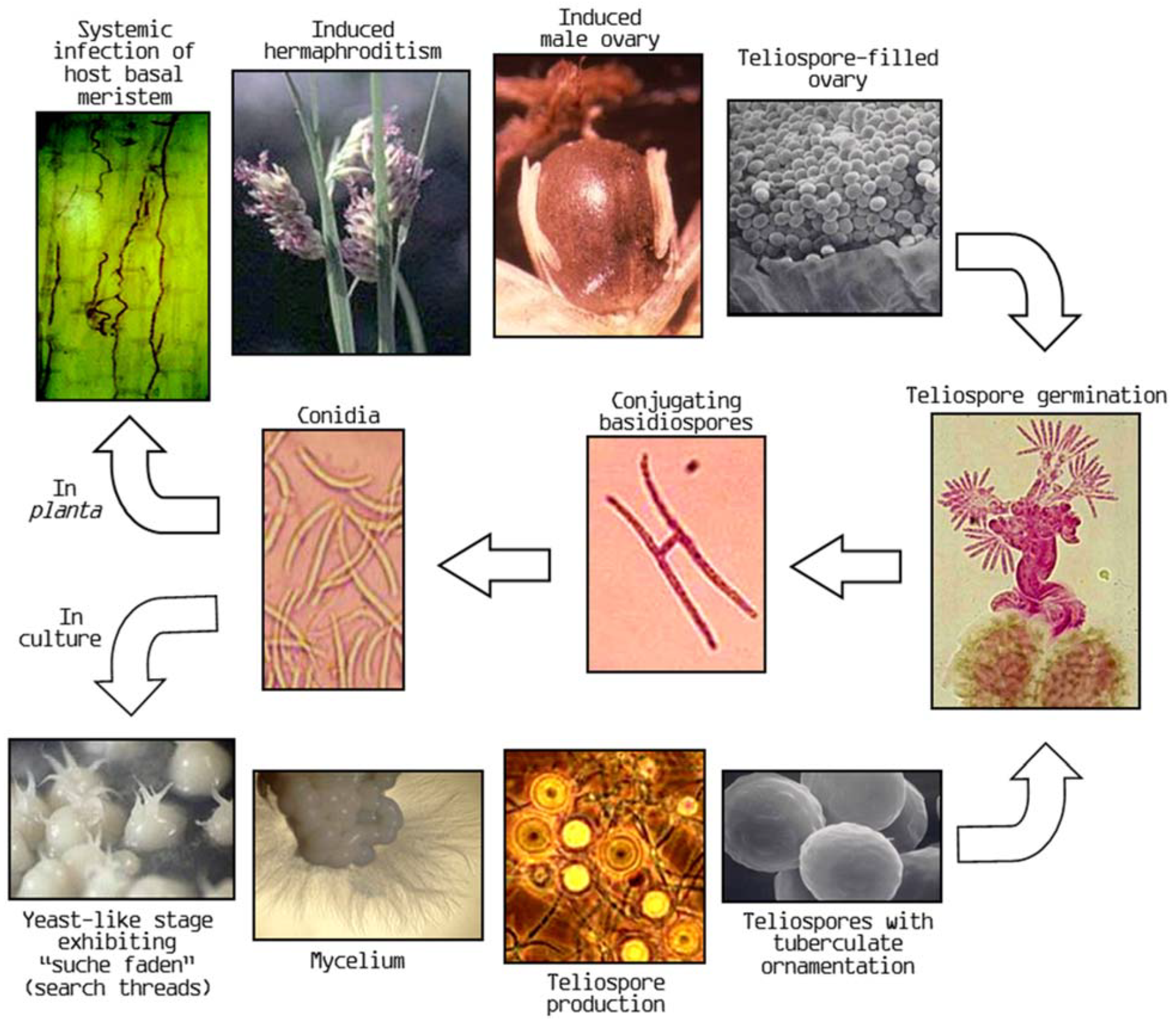
2.1. Pistil Smut Fungus Life Cycle and Secondary Effects of Pistil Smut Infection



2.2. Biomass Accumulation and Resource Partitioning

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomass plant−1 | Resource allocation plant−1 | ||||
|---|---|---|---|---|---|
| Total 1 (g) | Vegetative 2 (g) | Sexual 3 (g) | Vegetative (VRA) 4 | Sexual (SRA) 5 | |
| Female (N = 23) | |||||
| Infected | 85.0 | 68.5 | 16.51 | 0.803 | 0.197 |
| Uninfected | 128.9 | 125.2 | 3.63 | 0.973 | 0.027 |
| Avg. difference | −43.9 *** | −56.7 *** | 12.89 *** | −0.170 *** | 0.170 *** |
| (SEM) | (5.09) | (4.99) | (2.20) | (0.023) | (0.023) |
| Fold difference 6 | −1.52 | −1.83 | 4.55 | −1.21 | 7.21 |
| Male (N = 31) | |||||
| Infected | 77.3 | 64.9 | 12.46 | 0.833 | 0.167 |
| Uninfected | 120.0 | 116.2 | 3.78 | 0.968 | 0.032 |
| Avg. difference | −42.6 *** | −51.3 *** | 8.67 *** | −0.135 *** | 0.135 *** |
| (SEM) | (4.86) | (5.13) | (1.17) | (0.016) | (0.016) |
| Fold difference 6 | −1.55 | −1.79 | 3.29 | −1.16 | 5.22 |
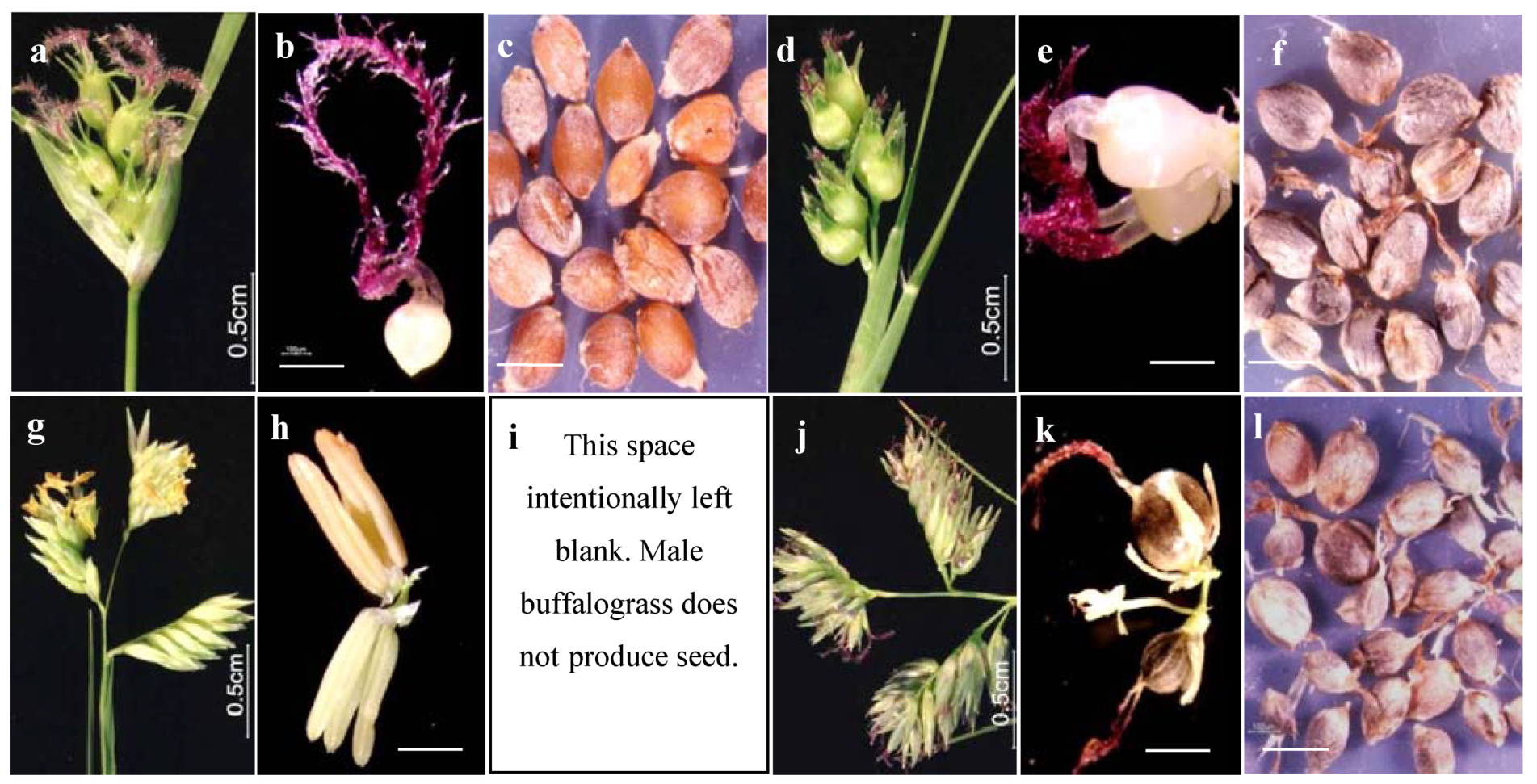
2.3. Seed Yield Components
| Inflorescences plant−1 | Spikes inflor.−1 | Spikelets spike−1 | Florets spikelet−1 | Estimated florets plant−1 1 | |
|---|---|---|---|---|---|
| Female | |||||
| Genotypes (N) | 23 | 20 | 20 | 20 | 20 |
| Infected | 141.9 | 3.58 | 4.84 | 2.00 | 5,683 |
| Uninfected | 21.2 | 3.16 | 2.95 | 1.00 | 219 |
| Avg. difference | 120.7 *** | 0.412 ** | 1.89 *** | 1.00 *** | 5,464 *** |
| (SEM) | (14.00) | (0.150) | (0.473) | (0.079) | (1,002) |
| Fold difference 2 | 6.70 | 1.13 | 1.64 | 2.00 | 25.98 |
| Male | |||||
| Genotypes (N) | 31 | 28 | 28 | 28 | 28 |
| Infected | 127.4 | 3.41 | 12.22 | 2.83 | 15,809 |
| Uninfected | 53.8 | 2.86 | 13.20 | 2.00 | 4,529 |
| Avg. difference | 73.6 *** | 0.55 *** | −0.98 ** | 0.83 *** | 11,280 *** |
| (SEM) | (10.56) | (0.102) | (0.380) | (0.055) | (1,379) |
| Fold difference 2 | 2.37 | 1.19 | −1.08 | 1.39 | 2.37 |
2.4. Potential Seed Yield and Harvest Index
| Estimated ovaries plant−1 1 | Weight of 100 seeds or smut balls (sb) 2 (mg) | Potential seed or smut ball yield plant−1 3 (g) | Potential harvest index plant−1 4 (%) | |
|---|---|---|---|---|
| Female | ||||
| Genotypes (N) | 20 | 6 | 20 | 20 |
| Infected | 5683 | 64.4 (sb) | 3.660 (sb) | 4.697 |
| Uninfected | 219 | 78.3 (seed) | 0.171 (seed) | 0.134 |
| Avg. difference | 5464 *** | −13.9 ** | 3.489 *** | 4.563 *** |
| (SEM) | (1002) | (3.50) | (0.647) | (1.006) |
| Fold difference 5 | 25.98 | −1.22 | 21.37 | 35.15 |
| Male | ||||
| Genotypes (N) | 28 | 6 | 28 | 28 |
| Infected | 15019 | 40.0 (sb) | 6.01 (sb) | 8.19 (sb) |
| Uninfected | 0 | 0 | 0 | 0 |
| Avg. difference | - | - | - | - |
| (SEM) | - | - | - | - |
| Fold difference 5 | N/A | N/A | N/A | N/A |
2.5. Potential Seed Yield and Harvest Index for a Male-Female Pair of Buffalograss
| Male + Female pair | Potential seed or smut ball yield pair−1 1 (g) | Potential harvest index pair−1 2 (%) |
|---|---|---|
| Uninfected (Seed) | 0.171 | 0.069 |
| Infected (Smut balls) | 9.670 | 6.022 |
| Fold difference 3 | 56.550 | 86.994 |
3. Experimental Section
3.1. Isolation and Culture of Pistil Smut
3.2. Plant Material
3.3. Host Infection by Pistil Smut
3.4. Resource Partitioning Analysis
3.5. Analysis of Seed Yield Components
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gorischek, A.M.; Afkhami, M.E.; Seifert, E.K.; Rudgers, J.A. Fungal symbionts as manipulators of plant reproductive biology. Am. Nat. 2013, 181, 562–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gundel, P.E.; Garibaldi, L.A.; Martínez-Ghersa, M.A.; Ghersa, C.M. Trade-off between seed number and weight: Influence of a grass—Endophyte symbiosis. Basic Appl. Ecol. 2012, 13, 32–39. [Google Scholar] [CrossRef]
- Shykoff, J.A.; Kaltz, O. Effects of anther smut Microbotryum violaceum on host life-history patterns in Silene latifolia (Caryophyllaceae). Int. J. Plant Sci. 1997, 158, 164–171. [Google Scholar] [CrossRef]
- Shykoff, J.A.; Kaltz, O. Phenotypic changes in host plants diseased by Microbotryum violaceum: Parasite manipulation, side effects, and trade-offs. Int. J. Plant Sci. 1998, 159, 236–243. [Google Scholar] [CrossRef]
- Purdy, L.H.; Kendrick, E.L.; Hoffman, J.A.; Holton, C.S. Dwarf bunt of wheat. Annu. Rev. Microbiol. 1963, 17, 199–222. [Google Scholar] [CrossRef]
- Trione, E.J.; Stockwell, V.O.; Latham, C.J. Floret development and teliospore production in bunt-infected wheat, in planta and in cultured spikelets. Phytopathology 1989, 79, 999–1002. [Google Scholar] [CrossRef]
- Ghareeb, H.; Becker, A.; Iven, T.; Feussner, I.; Schirawski, J. Sporisorium reilianum infection changes inflorescence and branching architectures of maize. Plant Physiol. 2011, 156, 2037–2052. [Google Scholar] [CrossRef] [PubMed]
- Cano, L.M.; Raffaele, S.; Haugen, R.H.; Saunders, D.G.O.; Leonelli, L.; MacLean, D.; Hogenhout, S.A.; Kamoun, S. Major transcriptome reprogramming underlies floral mimicry induced by the rust fungus Puccinia monoica in Boechera stricta. PLoS One 2013, 8, e75293. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Davis, R.E.; Nuss, D.L.; Zhao, Y. Phytoplasmal infection derails genetically preprogrammed meristem fate and alters plant architecture. Proc. Natl. Acad. Sci. USA 2013, 110, 19149–19154. [Google Scholar] [CrossRef] [PubMed]
- Hitchcock, A.S. Manual of the Grasses of the United States, 2nd ed.; Chase, A., Ed.; Dover Publications Inc.: New York, NY, USA, 1950; p. 991. [Google Scholar]
- Huff, D.R. Sex ratios and inheritance of anther and stigma color in diploid buffalograss. Crop Sci. 1992, 31, 328–332. [Google Scholar]
- Huff, D.R.; Wu, L. Distribution and inheritance of inconstant sex forms in natural populations of dioecious buffalograss (Buchloë dactyloides). Am. J. Bot. 1992, 79, 207–215. [Google Scholar] [CrossRef]
- Quinn, J.A.; Engel, J.L. Life-history strategies and sex ratios for a cultivar and a wild population of Buchloe dactyloides (Gramineae). Amer. J. Bot. 1986, 73, 874–881. [Google Scholar] [CrossRef]
- Kinney, M.S.; Columbus, T.; Friar, E.A. Dicliny in Bouteloua (Poaceae: Chloridoideae): implications for evolution of dioecy. Aliso 2007, 23, 605–614. [Google Scholar] [CrossRef]
- Wenger, L.E. Inflorescence variations in buffalo grass, Buchloe dactyloides. J. Am. Soc. Agron. 1940, 32, 274–277. [Google Scholar] [CrossRef]
- Huff, D.R. University of California: Davis, CA, USA; Personal Observation, 1984.
- Kellerman, W.A.; Swingle, W.T. New species of Kansas fungi. J. Mycol. 1889, 5, 11–14. [Google Scholar] [CrossRef]
- Chandra, A.; Huff, D.R. Salmacisia, a new genus of Tilletiales: Reclassification of Tilletia buchloeana causing induced hermaphroditism in buffalograss. Mycologia 2008, 100, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Chandra, A.; Huff, D.R. A fungal parasite regulates a putative female-suppressor gene homologous to maize Tasselseed2 and causes induced hermaphroditism in male buffalograss. Mol. Plant Microbe Interact. 2010, 23, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Baudoin, M. Host castration as a parasitic strategy. Evolution 1975, 29, 335–352. [Google Scholar] [CrossRef]
- Clay, K. Parasitic castration of plants by fungi. Trends Ecol. Evol. 1991, 6, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Kover, P.X. Effects of parasitic castration on plant resource allocation. Oecologia 2000, 123, 48–56. [Google Scholar] [CrossRef]
- Uchida, W.; Matsunaga, S.; Sugiyama, R.; Kazama, Y.; Kawano, S. Morphological development of anthers induced by the dimorphic smut fungus Microbotryum violaceum in female flowers of the dioecious plant Silene latifolia. Planta 2003, 218, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Alexander, H.; Antonovics, J. Disease spread and population dynamics of anther-smut infection of Silene alba caused by the fungus Ustilago violacea. J. Ecol. 1988, 76, 91–104. [Google Scholar] [CrossRef]
- Buono, L.; López-Villavicencio, M.; Shykoff, J.A.; Snirc, A.; Giraud, T. Influence of multiple infection and relatedness on virulence: Disease dynamics in an experimental plant population and its castrating parasite. PLoS One 2014, 9, e98526. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.F. Plant Reproductive Ecology; John Wiley & Sons: New York, NY, USA, 1983. [Google Scholar]
- Plant Resource Allocation. Available online: http://www.sciencedirect.com/science/book/9780120834907 (accessed on 12 October 2014).
- Mironchenko, A.; Kozłowski, J. Optimal allocation patterns and optimal seed mass of a perennial plant. J. Theor. Biol. 2014, 354, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Law, R. The cost of reproduction in annual meadow grass. Am. Nat. 1979, 113, 3–16. [Google Scholar] [CrossRef]
- Borlaug, N.E.; Narvaez, I.; Aresvik, O.; Anderson, R.G. Green revolution yields a golden harvest. Columbia J. World Bus. 1969, 4, 9–19. [Google Scholar]
- Herben, T.; Krahulec, F.; Hadincova, V.; Kovarova, M.; Skalova, H. Morphological constraints of shoot demography of a clonal plant: Extra- and intravaginal tillers of Festuca rubra. Plant Species Biol. 1994, 9, 183–189. [Google Scholar] [CrossRef]
- Liston, A.; Wilson, B.L.; Robinson, W.A.; Doescher, P.S.; Harris, N.R.; Svejcar, T. The relative importance of sexual reproduction versus clonal spread in an aridland bunchgrass. Oecologia 2003, 137, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Harberd, D.J. Observations on population structure and longevity of Festuca rubra L. New Phytol. 1961, 60, 184–206. [Google Scholar] [CrossRef]
- Harberd, D.J. Some observations on natural clones in Festuca ovina. New Phytol. 1962, 61, 85–100. [Google Scholar] [CrossRef]
- Charnov, E.L. The Theory of Sex Allocation; Princeton University Press: Princeton, NJ, USA, 1982; pp. 1–355. [Google Scholar]
- Huff, D.R. Bluegrasses. In Handbook of Plant Breeding: Fodder Crops and Amenity Grasses; Boller, B., Posselt, U.K., Veronesi, F., Eds.; Springer Publishing: New York, NY, USA, 2010; pp. 345–379. [Google Scholar]
- Reynold, M.P.; Borlaug, N.E. Impacts of breeding on international collaborative wheat improvement. J. Agric. Sci. 2006, 144, 3–17. [Google Scholar] [CrossRef]
- Mathre, D.E. Dwarf bunt: Politics, identification, and biology. Annu. Rev. Phytopathol. 1996, 34, 67–85. [Google Scholar] [CrossRef] [PubMed]
- DeHaan, L.R.; van Tassel, D.L.; Cox, T.S. Perennial grain crops: A synthesis of ecology and plant breeding. Renew. Agric. Food Syst. 2006, 20, 5–14. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Belesky, D.P. Adaptations of endophyte-infected cool-season grasses to environmental stresses: Mechanisms of drought and mineral stress tolerance. Crop Sci. 2000, 40, 923–940. [Google Scholar] [CrossRef]
- Parker, D.; Beckmann, M.; Zubair, H.; Enot, D.P.; Caracuel-Rios, Z.; Overy, D.P.; Snowdon, S.; Nicholas, J.; Talbot, N.J.; Draper, J. Metabolomic analysis reveals a common pattern of metabolic re-programming during invasion of three host plant species by Magnaporthe grisea. Plant J. 2009, 59, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.J.; Clay, K. Infection by the systemic fungus Epichloe glyceriae alters clonal growth of its grass host, Glyceria striata. Proc. R. Soc. Lond. B 2003, 270, 1585–1591. [Google Scholar] [CrossRef]
- Engelmann, G. Two new dioecious grasses of the United States; Transactions of the Academy of Science of St. Louis: St. Louis, MO, USA, 1859; pp. 431–443. [Google Scholar]
- Thompson, B.E.; Hake, S. Translational biology: From Arabidopsis flowers to grass inflorescence architecture. Plant Physiol. 2009, 149, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Pautler, M.; Tanak, W.; Hirano, H.; Jackson, D. Grass meristems I: Shoot apical meristem maintenance, axillary meristem determinacy and the floral transition. Plant Cell Physiol. 2013, 54, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Tanak, W.; Pautler, M.; Jackson, D.; Hirano, H. Grass meristems II: Inflorescence architecture, flower development and meristem fate. Plant Cell Physiol. 2013, 54, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Chuck, G.; Meeley, R.; Irish, E.; Sakai, H.; Hake, S. The maize tasselseed4 microRNA controls sex determination and meristem cell fate by targeting Tasselseed6/indeterminate spikelet1. Nat. Genet. 2007, 39, 151715–151721. [Google Scholar]
- Curaba, J.; Talbot, M.; Li, Z.; Helliwell, C.C. Over-expression of microRNA171 affects phase transitions and floral meristem determinancy in barley. BMC Plant Biol. 2013, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Huff, D.R.; Zagory, D.; Wu, L. Report of buffalograss bunt (Tilletia buchloëana) in Oklahoma. Plant Dis. 1987, 71, 651. [Google Scholar] [CrossRef]
- Huff, D.R.; Peakall, R.; Smouse, P.E. RAPD variation within and among natural populations of outcrossing buffalograss. Theor. Appl. Genet. 1993, 86, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Minitab, version 14.2; Minitab Inc.: State College, PA, USA, 2005.
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chandra, A.; Huff, D.R. Pistil Smut Infection Increases Ovary Production, Seed Yield Components, and Pseudosexual Reproductive Allocation in Buffalograss. Plants 2014, 3, 594-612. https://doi.org/10.3390/plants3040594
Chandra A, Huff DR. Pistil Smut Infection Increases Ovary Production, Seed Yield Components, and Pseudosexual Reproductive Allocation in Buffalograss. Plants. 2014; 3(4):594-612. https://doi.org/10.3390/plants3040594
Chicago/Turabian StyleChandra, Ambika, and David R. Huff. 2014. "Pistil Smut Infection Increases Ovary Production, Seed Yield Components, and Pseudosexual Reproductive Allocation in Buffalograss" Plants 3, no. 4: 594-612. https://doi.org/10.3390/plants3040594