
Physiological Signaling and Structure of the HGF Receptor MET


1
Department Translational Medicine, University Piemonte Orientale, via Solaroli 17, 28100 Novara, Italy
2
Università Vita-Salute San Raffaele, via Olgettina 58, 20132 Milano, Italy
*
Author to whom correspondence should be addressed.
Biomedicines 2015, 3(1), 1-31; https://doi.org/10.3390/biomedicines3010001
Submission received: 30 September 2014
/
Accepted: 9 December 2014
/
Published: 31 December 2014
(This article belongs to the Special Issue New aspects of the Hepatocyte Growth Factor/c-Met System)
Abstract
:The “hepatocyte growth factor” also known as “scatter factor”, is a multifunctional cytokine with the peculiar ability of simultaneously triggering epithelial cell proliferation, movement and survival. The combination of those proprieties results in the induction of an epithelial to mesenchymal transition in target cells, fundamental for embryogenesis but also exploited by tumor cells during metastatization. The hepatocyte growth factor receptor, MET, is a proto-oncogene and a prototypical transmembrane tyrosine kinase receptor. Inhere we discuss the MET molecular structure and the hepatocyte growth factor driven physiological signaling which coordinates epithelial proliferation, motility and morphogenesis.
1. Background Introduction
The Hepatocyte Growth Factor (HGF) was originally identified as a soluble factor promoting hepatocyte growth and liver regeneration [1]. In a parallel way a Scatter Factor (SF) was identified as cytokine secreted by fibroblast promoting dissociation and motility of epithelial cells in culture [2].
Molecular cloning demonstrated that HGF and SF are the same growth factor produced by cells of mesenchymal origin and promoting migration and proliferation depending on the epithelial cell targeted [3]. The HGF receptor (MET) is a prototypal tyrosine kinase receptor which plays a key role in the interaction between mesenchyme and epithelia during embryogenesis and tissue homeostasis [4]. The observation that MET is a proto-oncogene and that its signaling is often subverted in cancer has promoted deep investigations on its molecular structure and signaling proprieties which are reviewed herein.
2. HGF
The HGF gene map to 7q21.11, in the same chromosomal region of its receptor MET. By alternative splicing the gene give rise to several transcripts beside the full length isoform (728 aa corresponding to 83 kDa, increased to 92 kDa after glycosylation) [5]. The full length pre-pro-HGF feature a strong sequence and structural homology to plasminogen [6,7,8], presenting a signal peptide for secretion (residues 1–31), an amino-terminal heparin binding domain (residues 37–123), 4 kringle domains (residues 128–206, 211–288, 305–383 and 391–469) and a serine protease-like domain (residues 495–721) [5,9,10]. A close homolog of HGF is macrophage stimulating protein (MSP) with a 45% identity and a similar organization with 4 kringle domains [11].
Two other HGF splicing isoforms have been extensively characterized consisting of the amino-terminal domain linked in tandem with, respectively, the first one (NK1, 24 kDa) or the first two (NK2, 36 kDa) kringle domains [12]. All three isoforms bind to the receptor tyrosine kinase MET and evoke signaling but with different affinity and potency [13]. Indeed, NK1 stimulates cell proliferation, migration and tubular morphogenesis, though at reduced potency and with greater heparan sulfate (HS) dependence compared to full-length HGF, suggesting that the primary MET binding site is contained within this fragment [14]. Conversely, NK2 acts as a competitive inhibitor of full length HGF promoted mitogenicity, but retains motogenic activity in vitro and in vivo [15,16].
All HGF isoforms are synthesized as pre-peptides that undergo proteolytic cleavage at residue 31 to remove the leader sequence. Full-length pro-HGF also undergoes extensive glycosylation (N-linked at residue 294, 402, 566 and 653, O-linked at residue 476), which is dispensable for biological activity but promotes secretion [17,18]. Full length HGF requires proteolytic cleavage at the beginning of the serine protease like domain (between R494 and V495) to become a biologically active heterodimer consisting of disulfide-linked α (residues 32–494, 69 kDa after glycosylation) and β (residues 495–728, 32–34 kDa depending on the extension of glycosylation) chains [19,20,21]. Several serine proteases in serum are capable of HGF activation in vitro, including urokinase-type plasminogen activator and tissue-type plasminogen activator [22], HGF activator [23,24], matriptase [25], hepsin [26] and blood factors XIa and XIIa [24]. Extracellular processing may play a regulatory role on HGF biological activity as localized activation of pro-HGF has been evidenced in injured tissues [27].
The amino-terminal HS binding domain and secondary binding sites in the first kringle domain [28] play a major role in HGF biology representing a high capacity, medium affinity binding site in many cell types, comprising those unresponsive to HGF stimulation [29]. Those binding sites are constituted by heparan and dermatan sulfates found on extracellular matrix proteins such as decorin, syndecans and biglycan [30,31]. HGF also specifically binds to sulfoglycolipids which may also modulate its activity [32]. The abundance of HGF binding sites in extracellular matrix and basal membranes allow the local accumulation of HGF and its release in a spatially and temporally restricted manner through matrix turnover [31]. Furthermore HGF binding to cell-surface HS increase local HGF concentrations and putatively may allow for HGF dimerization [33] or change in conformation [30], effectively promoting receptor clustering and kinase activation [34,35]. Furthermore HS-modified CD44v3 interacts with MET and increase HGF-induced signal transduction [36]. Conversely, HS interactions with MET are substantially weaker than those with HGF, and their contribution to the stability a ternary HGF-HS-MET complex may not be critical for all HGF responses [37,38].
Lai and Goldschneider [39] reported a naturally occurring hybrid cytokine consisting of the HGF β chain and IL-17 acting as a pre-pro-B cell growth-stimulating factor. Although the molecular mechanisms by which such cytokine is produced are still uncharacterized it represents an efficient way to coordinate signals through both IL-7R and MET promoting B cell development.
3. MET
3.1. Gene and Transcript
The MET gene is located on 7q31 and encodes a 1390 aa protein, with an apparent molecular weight of 190 kDa, which is the most abundant form in a variety of tissues and cell lines. The use of an alternative in-frame splice junction results in a longer transcript variant with 18 additional amino acids in the extracellular region (1408 aa, apparent molecular weight of 170 kDa) [40]. Alternative splicing originates also a variant transcript of MET lacking 47 amino acids in the juxtamembrane region of the cytoplasmic domain present in adult mouse tissues including kidney, liver, and brain at lower levels than the full-length transcript. The deleted region in the cytoplasmic domain contains the S985 phosphorylated by protein kinase C down-regulating of MET kinase activity [41].
Truncated forms, resulting from receptor shedding, were reported in blood and in tumor derived cell lines, those forms are putatively involved in carcinogenesis [42,43,44,45]. Conversely, in stress conditions, the MET receptor is cleaved by caspases at D1002 within its juxtamembrane region, generating a pro-apoptotic intracellular fragment of 40 kDa [46].
MET is predominantly expressed in epithelial derived cells such as the epithelial layer lining the gastrointestinal tract, liver kidney, thyroid and in keratinocytes [47,48]. Lower levels were detected in other cell types such as endothelial cells [49], hematopoietic progenitors [50], B cells [51] and the brain [48]. Conversely, HGF is produced by several mesenchymal cells but is also contained in platelet granules and released during aggregation promoting wound closure and epithelial cell proliferation [5]. Thus the HGF-MET pair are at the base of epithelial mesenchymal interaction during embryogenesis, wound closure and angiogenesis [52].
3.2. MET Structure
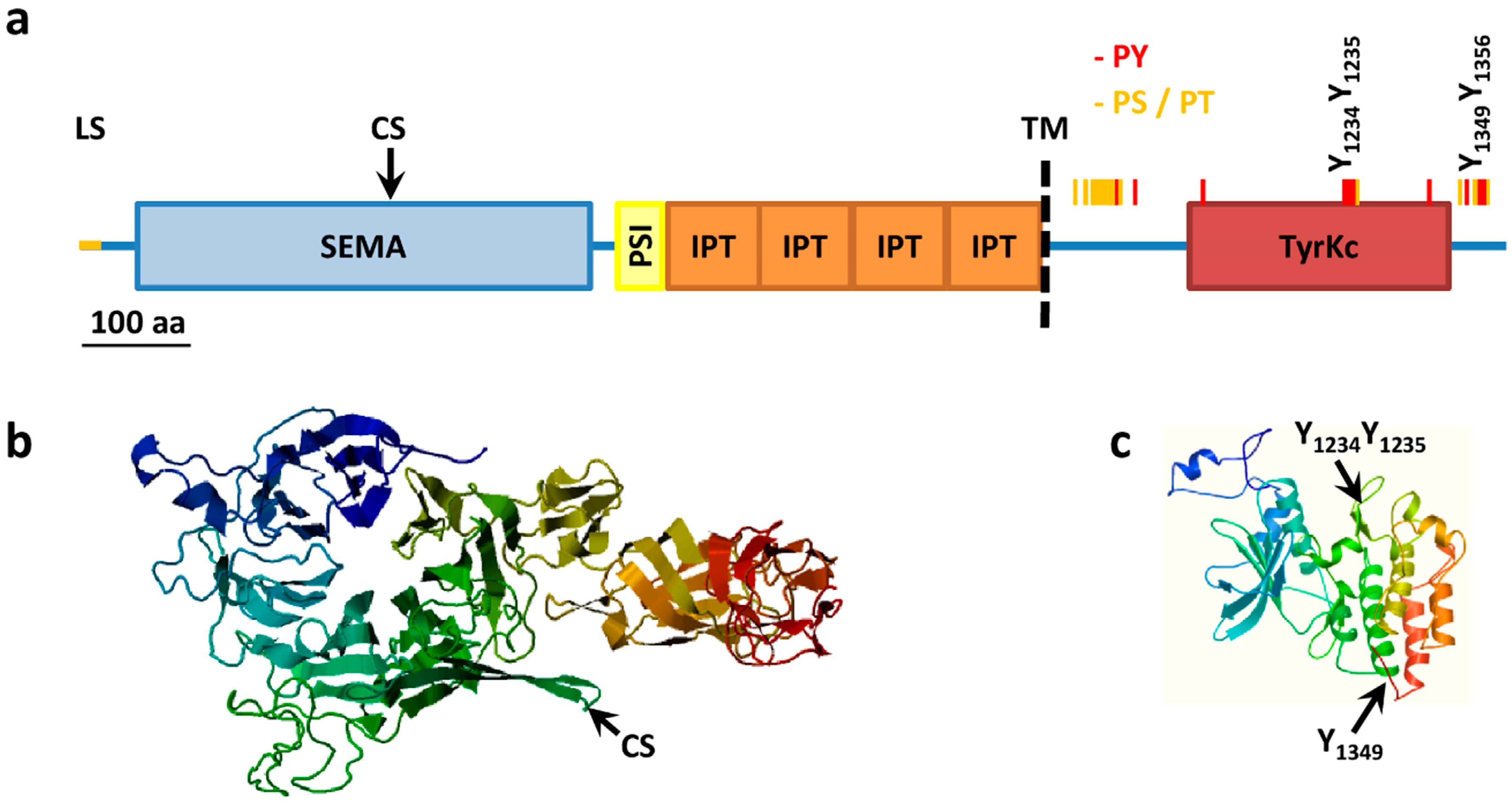
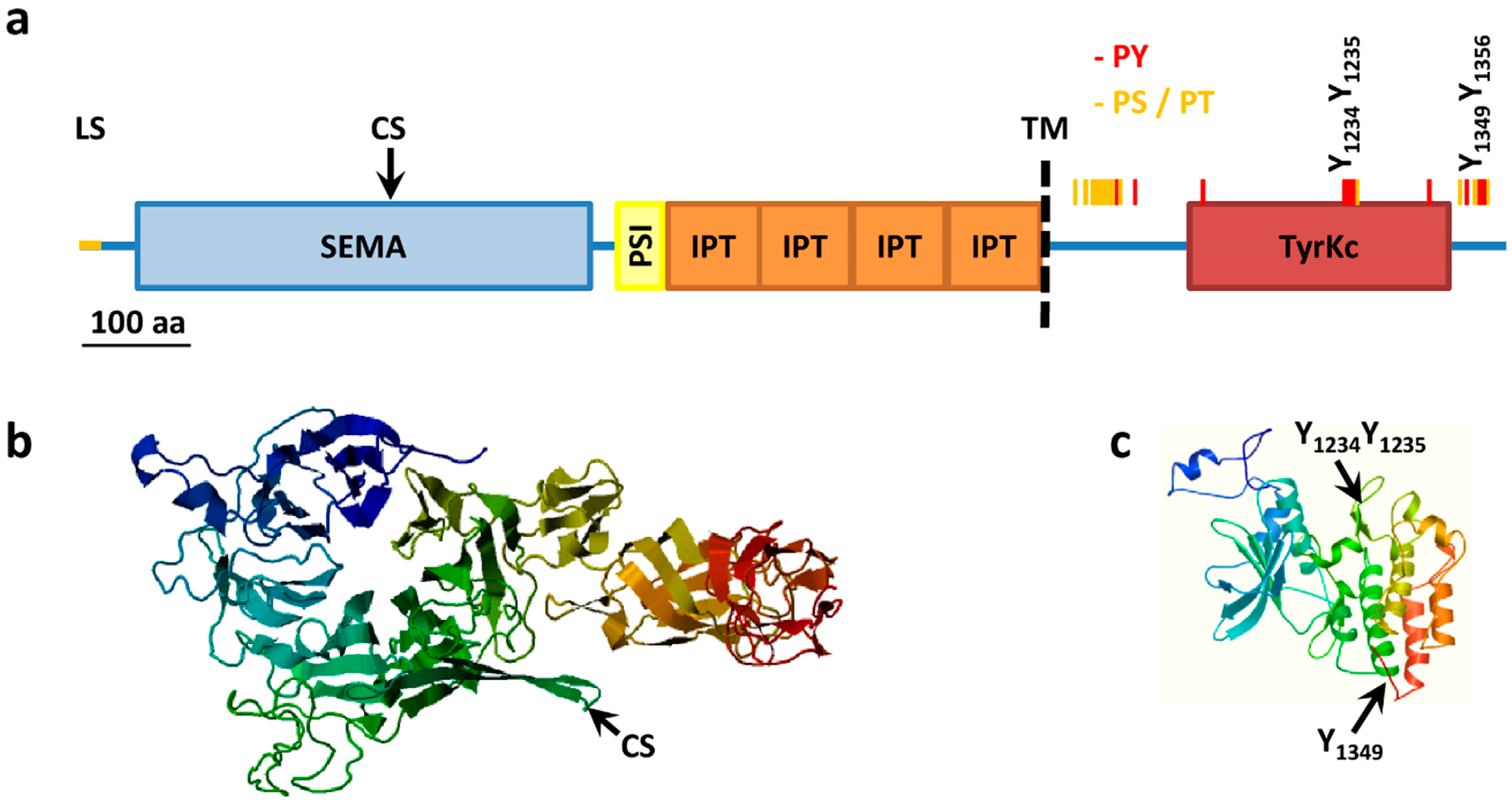
During synthesis in the endoplasmic reticulum the leader sequence (aa 1–24) is removed and MET is extensively co-translationally glycosylated at residue 106 [53] (further putative N-linked glycosylation sites: 45, 149, 202, 399, 405, 607, 635, 785, 879 and 930) to give a 190 kDa single chain precursor with several disulfide bonds [8,54,55]. Glycosylation and folding are a requisite for the following cleavage between residue 307 and 308 to give a mature heterodimer composed of an extracellular 50 kDa α-chain and a transmembrane 140 kDa β-chain, linked together by disulfide bridges. Cleavage is carried out by furin proteases but is not required for receptor activation by HGF [56], indeed the longer transcript variant is exposed on cell surface and phosphorylated but is not processed in an α–β heterodimer [40]. The extracellular α-chain contains the ligand binding pocket while the intracellular portion of the β-chain contains the tyrosine kinase domain and a conserved two-tyrosine multifunctional docking site that interacts with multiple SRC homology 2 (SH2) containing intracellular signal transducers (Figure 1).
Paralleling the similarity between HGF and MSP, the closest homolog of MET is the MSP tyrosine kinase receptor RON, with an overall identity of 33% of amino acids, mainly at the level of the tyrosine kinase domain with a 64% identity [57]. Conversely the extracellular portion presents a series of domains with homology to the semaphorin receptor family (semaphorin domain, SEMA), to plexins and integrins (Plexin SEMAphorins Integrines domain, PSI) and four immunoglobulin-like folds shared by plexins and transcription factors (IPT domain). MET structure and key post-translational modification are summarized in Figure 1.
Figure 1.
MET structure. (a) MET structural domains. MET domains as predicted using SMART. SEMA, Semaphorin domain (aa 52–496); PSI, Plexin semaphorin domain (aa 519–562); IPT, IG like plexins transcription factor (aa 562–655); IPT, IG like plexins transcription factor (aa 656–739); IPT, IG like plexins transcription factor (aa 741–836); IPT, IG like plexins transcription factor (aa 838–934); TM, transmembrane region (aa 933–955); TyrKc, tyrosine kinase (aa 1078–1337); Further features specified: LS, Leader sequence (aa 1–24); CS, Cleavage site (aa 307–308); PY, tyrosine phosphorylation sites (red bars); PS/PT, serine/threonine phosphorylation sites (yellow bars); (b) Model of extracellular portions of MET from the SWISS-MODEL repository aa 42 to 741, Model_id ee0753c4f68e188bddb0c66890beee23_UP000052_4; (c) Model of intracellular portions of MET from the SWISS-MODEL repository, aa 1024 to 1352, Model_id ee0753c4f68e188bddb0c66890beee23_UP000052_3.
Figure 1.
MET structure. (a) MET structural domains. MET domains as predicted using SMART. SEMA, Semaphorin domain (aa 52–496); PSI, Plexin semaphorin domain (aa 519–562); IPT, IG like plexins transcription factor (aa 562–655); IPT, IG like plexins transcription factor (aa 656–739); IPT, IG like plexins transcription factor (aa 741–836); IPT, IG like plexins transcription factor (aa 838–934); TM, transmembrane region (aa 933–955); TyrKc, tyrosine kinase (aa 1078–1337); Further features specified: LS, Leader sequence (aa 1–24); CS, Cleavage site (aa 307–308); PY, tyrosine phosphorylation sites (red bars); PS/PT, serine/threonine phosphorylation sites (yellow bars); (b) Model of extracellular portions of MET from the SWISS-MODEL repository aa 42 to 741, Model_id ee0753c4f68e188bddb0c66890beee23_UP000052_4; (c) Model of intracellular portions of MET from the SWISS-MODEL repository, aa 1024 to 1352, Model_id ee0753c4f68e188bddb0c66890beee23_UP000052_3.
3.3. MET Activation and Signaling
The tyrosine kinase activity of MET transduces the mitogenic and motogenic signals elicited by HGF [58,59]. Notably, beside HGF, the Listeria monocytogenes surface protein InlB is a MET agonist that mimics HGF signaling, inducing bacterial entry through exploitation of a host RTK signaling pathway [60]. While a crystal structure of mature α/β HGF bond to MET is still missing, several studies employed HGF fragments and mutational analysis to identify the contact interface. A direct interaction between HGF β-chain and MET SEMA domain emerges from crystallographic studies but is insufficient to promote receptor activation [7,8]. Conversely, the N terminal HGF region corresponding to NK1 induces signaling by high affinity binding to MET SEMA domain [35,38,61], although other authors reported an interaction of this region with MET IPT domains 3 and 4 [62]. Ligand binding give rise to a change of MET conformation allowing:
- (1)
- (2)
- (3)
The key event inducing signaling is the auto-phosphorylation of the multifunctional docking site made of the tandem arranged Y1349 and Y1356 in the degenerate sequence YVH/NV (Table 1). Those two residues mediate intermediate- to high-affinity interactions with multiple SH2 containing signal transducers. Indeed the mutation of the two tyrosines results in loss of biological function [67,68]. In particular a pivotal role is played by Y1356, as MET containing an Y1356F substitution is unable to transduce motogenic signals. Y1356VNV represents a consensus binding site for multiple effectors such as the adaptor GRB2, the p85 subunit of phosphatidyl inositol 3-kinase (PI3K), phospholipase Cγ (PLCγ), and the tyrosine-protein phosphatase SHP-2 [68,70,71,72]. Together with Y1349 this tyrosine is also required for association and phosphorylation of the adaptor SHC [73]. In vivo, mutation of both residues in the mouse genome caused embryonic lethality, with placenta, liver, and limb muscle defects, mimicking the phenotype of MET null mutants [74].
A fundamental amplification role in the MET signalosome assembly is played by the Y1356 mediated recruitment of the GRB2-associated binding protein (GAB1). Indeed selective disruption of the GRB2 consensus in MET impairs GRB2 and GAB1 association resulting in decreased mitogen-activated protein kinase (MAPK) activation. This reduced level of signaling is sufficient for motility but not for branching morphogenesis and cell transformation [70,75]. In vivo, disrupting the consensus for GRB2 binding allows mice development to proceed to term without affecting placenta and liver, but caused a striking reduction in limb muscle coupled to a generalized deficit of secondary fibers [74]. Conversely, a point mutation which duplicates the GRB2 binding site, super-activates the Ras GTPase pathway and prevents the binding of the other intracellular transducers. This increased the transforming ability of the oncogene but abolished its metastatic potential [76].

{kind=link}
{kind=link}
| SS | MS | aa | Homo sapiens | aa | Mus musculus | aa | Rattus norvegicus |
|---|---|---|---|---|---|---|---|
| 0 | 6 | S966-p | KQIkDLGsELVRyDA | S964 | RKHKDLGSELVRYDA | S967 | RKHKDLGSELVRYDA |
| 0 | 5 | Y971-p | LGsELVRyDARVHtP | Y969 | LGSELVRYDARVHtP | Y972 | LGSELVRYDARVHTP |
| 0 | 6 | T977-p | RyDARVHtPHLDRLV | T975-p | RYDARVHtPHLDRLV | T978 | RYDARVHTPHLDRLV |
| 6 | 0 | S985-p | PHLDRLVsARsVsPt | S983-p | PHLDRLVsARSVsPT | S986-p | PHLDRLVsARSVSPT |
| 0 | 19 | S988-p | DRLVsARsVsPttEM | S986 | DRLVsARSVsPTTEM | S989 | DRLVsARSVSPTTEM |
| 0 | 32 | S990-p | LVsARsVsPttEMVs | S988-p | LVsARSVsPTTEMVs | S991 | LVsARSVSPTTEMVS |
| 0 | 11 | T992-p | sARsVsPttEMVsNE | T990 | sARSVsPTTEMVsNE | T993 | sARSVSPTTEMVSNE |
| 0 | 7 | T993-p | ARsVsPttEMVsNEs | T991 | ARSVsPTTEMVsNEs | T994 | ARSVSPTTEMVSNES |
| 0 | 20 | S997-p | sPttEMVsNEsVDyR | S995-p | sPTTEMVsNEsVDyR | S998 | SPTTEMVSNESVDYR |
| 1 | 43 | S1000-p | tEMVsNEsVDyRAtF | S998-p | TEMVsNEsVDyRATF | S1001 | TEMVSNESVDYRATF |
| 11 | 361 | Y1003-p | VsNEsVDyRAtFPED | Y1001-p | VsNEsVDyRATFPED | Y1004 | VSNESVDYRATFPED |
| 0 | 9 | T1006-p | EsVDyRAtFPEDQFP | T1004 | EsVDyRATFPEDQFP | T1007 | ESVDYRATFPEDQFP |
| 0 | 15 | Y1026-p | GsCRQVQyPLTDMSP | Y1024 | GACRQVQYPLTDLSP | Y1027 | GACRQVQYLLTDLSP |
| 0 | 31 | Y1093-p | RGHFGCVyHGtLLDN | Y1091 | RGHFGCVYHGTLLDN | Y1094 | RGHFGCVYHGTLLDS |
| 4 | 112 | Y1230-p | FGLARDMyDkEyysV | Y1228-p | FGLArDMyDKEyysV | Y1231 | FGLARDMYDKEyySV |
| 39 * | 735 | Y1234-p | RDMyDkEyysVHNkt | Y1232-p | rDMyDKEyysVHNKt | Y1235-p | RDMYDKEyySVHNKT |
| 38 * | 443 | Y1235-p | DMyDkEyysVHNktG | Y1233-p | DMyDKEyysVHNKtG | Y1236-p | DMYDKEyySVHNKTG |
| 1 | 177 | S1236-p | MyDkEyysVHNktGA | S1234-p | MyDKEyysVHNKtGA | S1237 | MYDKEyySVHNKTGA |
| 0 | 5 | T1241-p | yysVHNktGAKLPVK | T1239-p | yysVHNKtGAKLPVK | T1242 | yySVHNKTGAKLPVK |
| 6 | 5 | Y1313-p | EyCPDPLyEVMLkCW | Y1311-p | EYCPDALyEVMLKCW | Y1314 | EYCPDALYEVMLKCW |
| 0 | 6 | T1343-p | RISAIFstFIGEHyV | T1341 | RISSIFSTFIGEHyV | T1344 | RISSIFSTFIGEHYV |
| 24 * | 122 | Y1349-p | stFIGEHyVHVNAty | Y1347-p | STFIGEHyVHVNATy | Y1350 | STFIGEHYVHVNATY |
| 0 | 40 | T1355-p | HyVHVNAtyVNVKCV | T1353 | HyVHVNATyVNVKCV | T1356 | HYVHVNATYVNVKCV |
| 22 * | 120 | Y1356-p | yVHVNAtyVNVKCVA | Y1354-p | yVHVNATyVNVKCVA | Y1357 | YVHVNATYVNVKCVA |
| 7 | 116 | Y1365-p | NVKCVAPyPsLLssE | Y1363-p | NVKCVAPyPSLLPSQ | Y1366 | NVKCVAPYPSLLPSQ |
| 0 | 8 | S1367-p | KCVAPyPsLLssEDN | S1365 | KCVAPyPSLLPSQDN | S1368 | KCVAPYPSLLPSQDN |
Data are from PhosphoSitePlus (www.phosphosite.org) only sites with 5 or more references are reported. SS, site-specific studies; MS, mass spectrometry in discovery mode; * autocatalysis.
The best-characterized MET signal transducers and their role in signaling are summarized below:
GRB2, an adaptor which binds Y1356 inserted in a YVNV motif [72]. Through its SRC homology region 3 (SH3), GRB2 recruits to the receptor the Ras activator SOS, promoting the MAPK signaling cascade. The GRB2 SH3 also binds a GAB1 sequence rich in prolines, promoting the association to the receptor of GAB1, which is in turn heavily phosphorylated [77].
GAB1, a member of IRS family of adaptors, sharing an amino-terminal pleckstrin homology (PH) domain that controls subcellular localization to areas of cell–cell contacts. It is recruited to MET through GRB2, but also contains a MET binding domain [78]. GAB1 presents multiple tyrosine residues that act to further recruit SH2 or phospho-tyrosine binding (PTB) domain-containing substrates [79]. Indeed GAB1 substantially contributes to the recruitment to MET of:
NCK, an adaptor protein, which contains three SH3 domains followed by one SH2 domain. HGF promotes Nck activation and its co-precipitation with PLCγ [87,88]. NCK also regulates actin polymerization and remodeling by recruiting N-WASP and the WAVE complex [89], promoting dorsal ruffles formation [90].
SHC, an SH2 containing adaptor that, after stimulation of the HGF receptor, is phosphorylated on Y317, generating a further high affinity binding site for GRB2 and triggering the Ras pathway [73].
PI3K, the class I phosphatidyl inositol 3-kinase associates via the p85 subunit to GAB1 but also bind directly to the receptor. Upon receptor recruitment to the receptor PI3K is tyrosine phosphorylated and activated [91,92,93]. PI3K activity is critical for MET-mediated chemotaxis and tubulogenesis and less for mitogenesis [94,95]. PI3K also coordinates survival and metabolism via phosphatidylinositol 3,4,5-trisphosphate dependent recruitment and activation of PH containing proteins such as the kinases PDK and AKT.
PLCγ, a phosphoinositide-specific phospholipase, which associates to the receptor and is tyrosine phosphorylated and activated. PLCγ promotes an early peak of inositol 1,4,5-trisphosphate (IP3) and 1,2-diacylglycerol (DG) release [92], while at later times the DG is sustained by phosphatidylcholine-specific PLC activity [96]. PLCγ activity leads to IP3 mediated release of Ca2+ from intracellular stores within seconds and induces Ca2+ oscillation peaking at 2 to 5 hours [97]. The parallel accumulation of Ca2+ and DG sustains classical protein kinase C (PKC) [98], which are not required for MAPK activation but collaborates with p38 and p42/p44 MAPKs in HGF-induced proliferation [99].
HGF also activated phospholipase D (PLD) in a PKC dependent way. PLD hydrolyzes phosphatidylcholine to choline and phosphatidic acid (PA), which is further metabolized to DG by PA phosphohydrolase (PAP). The PLD-PAP pathways quantitatively contributes to DG accumulation and to the expressions of JUN and FOS transcription factors [100].
SRC, the proto-oncogene tyrosine-protein kinase SRC is the prototype of a family of closely related tyrosine kinases among which SRC [101] and FYN [92] bind to MET and are activated upon HGF stimulation. SRC contributes to GAB1 phosphorylation, and is essential for HGF induced motility and transformation but dispensable for proliferation [102]. SRC activation by MET promotes tyrosine phosphorylation of several downstream proteins not directly associated to the receptor such as:
- FAK, the focal adhesion kinase, which lies at the crossroad between integrin and growth factor signaling. The FAK Y194 is also directly phosphorylated by MET contributing to activation [103]. Once activated FAK induces downstream GRB2 binding and MAPK signaling, critically controlling the cytoskeleton [104];
- αDGK (Diacylglycerol kinase alpha), which phosphorylates DG to PA. αDGK is phosphorylated by SRC on Y335 and its activity is crucial for HGF-induced cell motility by promoting PA production at ruffling sites. This drives local recruitment of PA binding proteins involved in migration such as the Rho GDP-dissociation inhibitor (RhoGDI) and atypical PKC [105,106,107,108] as well as integrin recycling such as the Rab11 interactor RCP [109].
STAT3, a transcription factor, which upon HGF treatment is recruited to MET, tyrosine phosphorylated and translocates in the nucleus within in hours. STAT3 directly couples MET signaling to the transcription program required for tubule formation in vitro, without affecting either HGF-induced scattering or growth [110,111].
SHIP-1/2, the SH2 domain-containing inositol 5-phosphatases 1 and 2. SHIP-1/2 binds at Y1356 and positively affects lamellipodia extension and tubulogenesis [112,113].
Phospholipase A2 (PLA2), is not complexed to the receptor but tyrosine phosphorylated by MET and serine phosphorylated by MAPK. PLA2 triggers arachidonic acid release by membrane phospholipids and is functionally coupled with the HGF triggered activation of the COX-2 transcription through the MAPK signaling pathway [114]. The production of arachidonic acid derived mediators by COX-2 amplifies MET signaling and putatively coordinates epithelial sheet responses in a paracrine fashion [115].
Consistently with the pathways described, a global analysis of MET signaling in small cell lung cancer using phospho-arrays identified as positively regulated HGF targets:
- MET (Y1003/Y1230/Y1234/Y1235);
- phosphoproteins that regulate transcriptional control: STAT3 (S727) and CREB (S133);
- cell cycle G1/S checkpoint: RB (S612), RB1 (S780);
- cell survival and apoptosis: AKT1 (S473/T308), JNK (T183/Y185);
- cell proliferation and differentiation: MEK1/2 (S221/S225), ERK1/2 (T185/Y187), ERK1/2 (T202/Y204);
- stress and inflammatory response to cytokines and growth factors: MEK3/6 (S189/S207), p38α (T180/Y182); JNK (T183/Y185);
- Cytoskeletal functions: FAK (Y576/S722/S910), adducin-α (S724) and adducin-γ (S662).
In the same study a modest inhibition of HGF-induced phosphorylation in the following phosphoproteins was detected: PKCα (S657), PKCα/β (T368/641), and PKCδ (T505); the anti-proliferative and pro-apoptotic PKR (T451) and the cell cycle checkpoint regulator CDK1 (T14/Y15) [116].
3.4. Biological Effects of MET Triggering
The interest on HGF–MET signaling has been boosted by its intriguing ability to conjugate in epithelial cells the typical growth factor capability of promoting proliferation and survival with a very strong motogenic activity. The coupling of these two activities in a single “invasive growth program” points to a key role in both embryonic development and tumor metastatization [21]. Indeed a global analysis of HGF/MET dependent transcriptome indicates that targets of the MET pathway included genes involved in the regulation of cell motility, cytoskeletal organization, angiogenesis and oxidative stress responses [117].
Those activities are very reminiscent of epithelial to mesenchymal transition (EMT), a complex program that during embryogenesis enables epithelial cells to acquire mesenchymal migratory phenotype, populate and establish tissues in distant body regions. EMT is also exploited by tumor cell for metastatic dissemination and chemoresistance [118]. Indeed HGF induces EMT master genes such as Snail and Slug via the early growth response protein 1 (Egr1) [119,120]. Furthermore, migration and repopulation of distant sites are reminiscent of a stem phenotype, indeed HGF triggering of MET induces a stem-like phenotype in human prostate cancer cells with a stem-like signature of gene expression and markers of stemness [121]. In untransformed stem cells, HGF and MET are involved in the exit from quiescence and pre-activation [122].
The HGF peculiar capabilities have been characterized in typical in vitro assays, widely used to study HGF and MET activity and signaling, which are described below together with the main signaling pathways involved (Figure 2).
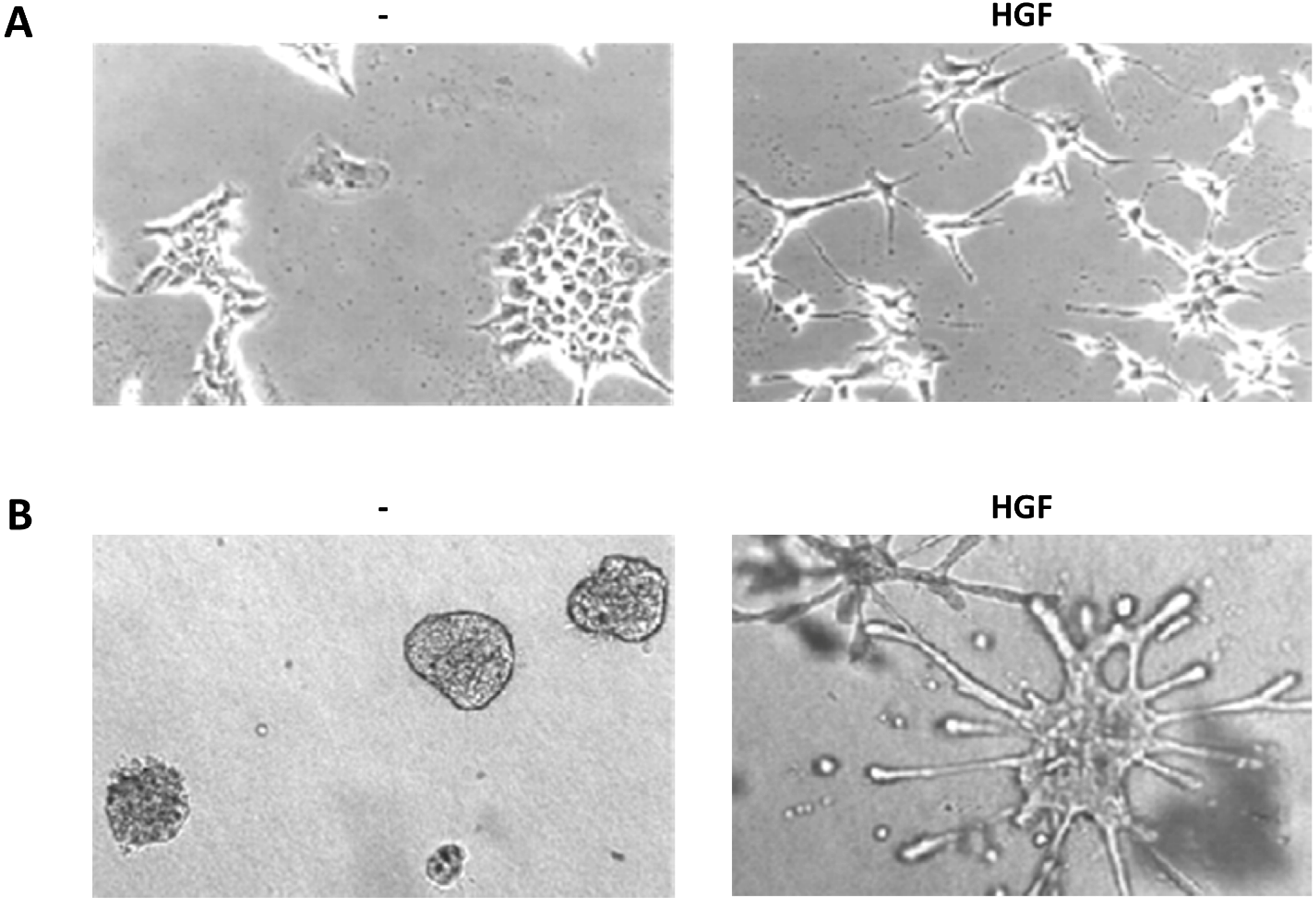
Figure 2.
HGF biological assays. (a) Scatter assay. Mouse embryonic liver derived cells (MLP-29, [123]) cultured on plastic in presence or absence of HGF (5 U/mL, 24 h); (b) Branching morphogenesis assay. MLP-29 cells cultured in collagen gels in presence or absence of HGF (20 U/mL, 7 days).
Figure 2.
HGF biological assays. (a) Scatter assay. Mouse embryonic liver derived cells (MLP-29, [123]) cultured on plastic in presence or absence of HGF (5 U/mL, 24 h); (b) Branching morphogenesis assay. MLP-29 cells cultured in collagen gels in presence or absence of HGF (20 U/mL, 7 days).
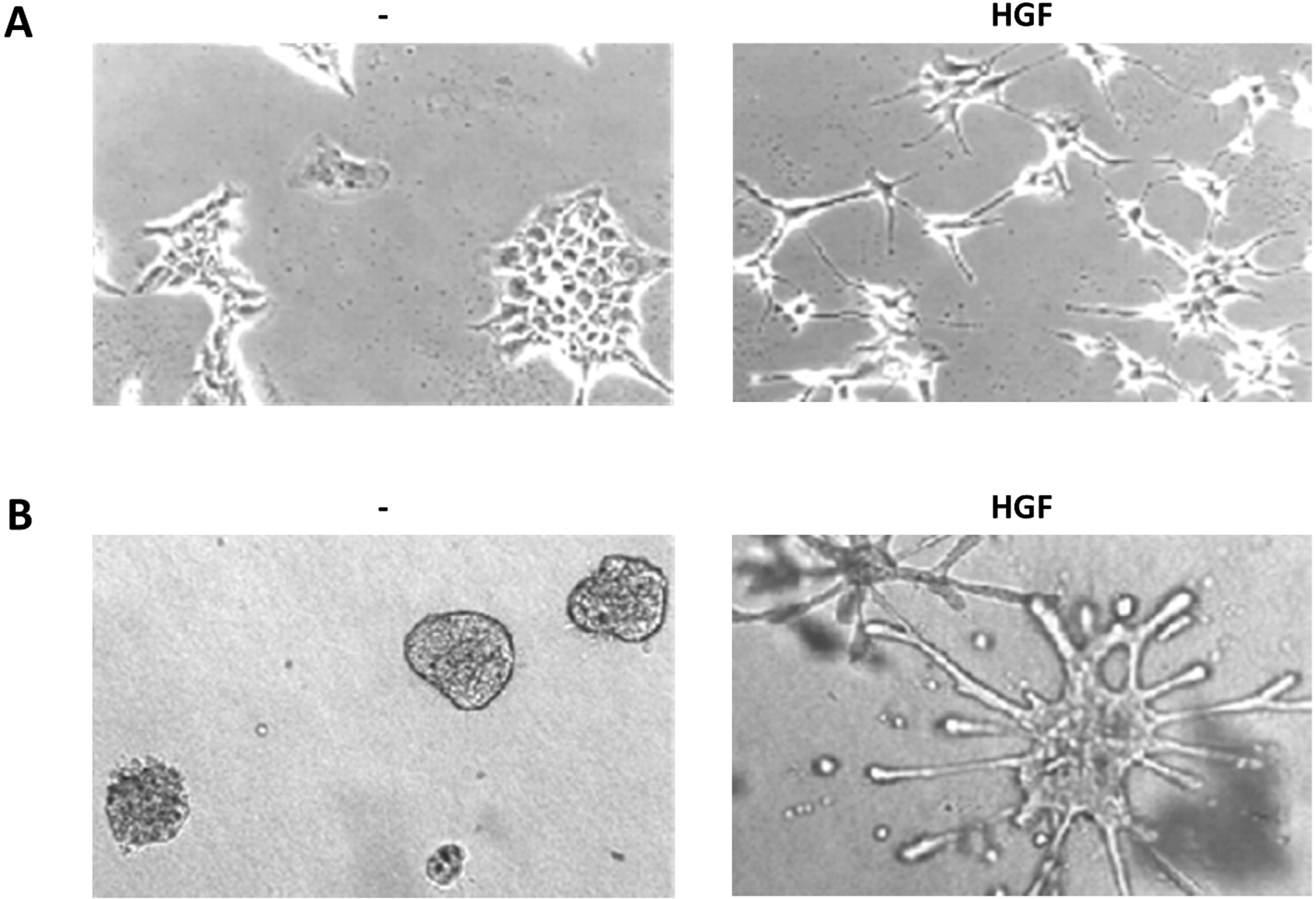
3.4.1. Scattering
Serum-starved untransformed epithelial cells (typically MDCK, Madin-Darby canine kidney epithelial cells [2]) in 2D cultures grow as monolayers with junctional complexes reminiscent of epithelial sheets. HGF stimulation induces a rapid loss of actin stress fibers, which are replaced by smaller peripheral actin filaments, and extensive membrane ruffling. After 4 h of HGF treatment become apparent a loss of junctional complexes (comprising E-cadherin, desmoplakins and the tight junction protein ZO-1) and an increased tyrosine phosphorylation of β-catenin [124]. At the same time the cells loose the cuboidal epithelial shape and the apico-basal polarity to spread and elongate in a fibroblast-like morphology with leading edge-trailing edge polarity, morphologically reproducing EMT [71,125].
HGF-induced cell–cell dissociation and the following dispersion are dependent on Ras and on the downstream activation of both MAPK and PI3K [71,126,127,128,129]. Prolonged activation of Ras-MAPK signaling downstream of HGF is known to promote cell migration, paxillin phosphorylation [127,130,131,132] and expression of matrix metalloproteinase (MMP) 9 via the transcription factors Elk-1 and FOS [133]. Concomitantly the PI3K-AKT-S6K pathway stimulates the expression and proteolytic activity of MMP-9 and matrix invasion [134]. HGF also causes tyrosine phosphorylation and redistribution of β-catenin in the hepatocytes and this effect is attributable to subcellular association of MET and β-catenin. HGF treatment of mouse mammary cells also leads to a transient decrease in GSK3 kinase activity and a parallel increase in the nuclear accumulation of β-catenin and activation of a LEF responsive reporter gene. Part of the EMT program evoked by HGF might be attributable to nuclear β-catenin and the resulting target gene expression [135,136].
In HGF treated epithelial MDCK colonies, MET is transcytosed from the basolateral membrane on Rab4 endosomes, to the apical surface where MET, as well as the MET substrate and scaffold protein, GAB1, localizes and signals at the dorsal ruffles [137]. As dorsal ruffles collapse, MET is internalized into EEA1- and Rab5-positive endosomes and is targeted for degradation through delivery to an Hrs-positive sorting compartment (see below).
The considerable HGF-induced reorganization of the actin cytoskeleton involved in scattering is mediated by the Rho family GTPases downstream to both Ras and PI3K [128]. Indeed HGF promotes activation of Rac1, RhoA and Cdc42 concomitant with the formation of filopodia and lamellipodia [138,139]. Cdc42 and Rac1 activities are required for HGF-induced cell–cell dissociation in MDCK cells but also for inducing ruffling and spreading by promoting activation of the Cdc42/Rac-regulated p21-activated kinase PAK [126,139]. The activation of Rac and Cdc42 is long lasting (till 24 h), consistent with a persistent polarized and migratory phenotype [140]. According with a starter role of Rho in ruffling [141,142] and with the observed prolonged decrease in stress fibers and increased spreading, a transient RhoA activation is observed in the first minutes of stimulation [143,144]. Indeed, inhibition of RhoA blocks HGF-induced cell scattering [128,142] and inhibition of the Rho-dependent kinase (ROCK) results in a reduction in HGF-induced membrane protrusion, reduction of dissociation and impaired motility with cell assuming a typical shape with elongated tails due to an impairment of tail retraction [138,139,145].
In connection with the actin cytoskeleton, there is a remodeling of adhesion sites, with a reduction in large paxillin-associated substratum adhesions particularly in areas of active membrane protrusion, where they are replaced by small peripheral focal contacts, which continuously disassemble or mature to focal adhesions during cell locomotion. Within minutes after exposure of HGF, FAK and paxillin become transiently phosphorylated in coincidence with the conversion to a motile phenotype [146,147]. HGF can also induce serine/threonine phosphorylation of paxillin most probably mediated directly by ERK, resulting in the recruitment and activation of FAK and subsequent enhancement of cell spreading and adhesion [130]. Rho and ROCK activity are required for the formation of mature focal adhesions by promoting cytoskeletal tension and stress fibers in response to HGF [139]. At the same time HGF promotes cell adhesion and invasiveness by increasing the avidity of integrins for their specific ligands in a PI3K dependent way [148,149].
3.4.2. Branching Morphogenesis
When plated in 3D collagen rich reconstituted extracellular matrix, untransformed epithelial cells (also in this case mainly MDCK) forms hollow cysts with typical apico-basal polarity. In polarized epithelial cells MET asymmetrically distributes at the basolateral membranes reflecting association with cell to cell and cell to matrix contacts [150,151]. At the membrane MET partially colocalize with caveolin in detergent resistant membranes and exploit caveolin for signal amplification [152].
Upon HGF treatment, cysts undergo growth and extend in the matrix forming tubular structures with interconnected lumens, in a process reminiscent of the epithelial organogenic program. Early in tubule development, MDCK cells exhibit many features characteristic of EMT forming long, invasive cytoplasmic extensions. Extension formation requires PI3K activity, whereas ROCK controls their number and length [153].
Extensions next proliferate and arrange in rows from one to three cells long, showing elongated cells which invade the matrix. While cells in the monolayer divide with their spindle axis parallel to the monolayer, HGF dislodge the spindle axis so that one of the daughter cells can apparently leave the monolayer to initiate a chain [153]. Those cells loose apico-basal polarity and gain leading edge-trailing edge polarity but maintain some intercellular adhesion representing a partial and transient EMT. Indeed cells in chains redifferentiate [154], loose their mesenchymal qualities and form multilayered cords by expanding regions of cell-cell contact and reestablishing a cuboidal shape. Nascent lumens and incomplete apical and basolateral domains appear. Eventually, cords mature into tubules through formation of a single continuous lumen and coordinated apico-basal polarization of individual cells.
Activation of ERK is necessary and sufficient for the initial step, during which cells depolarize and migrate, while becomes dispensable for the latter stage, during which cells repolarize and differentiate [155]. Conversely MMPs are essential for the late re-differentiation stage of tubulogenesis [155]. MMPs represent a class of HGF effectors positively regulated by HGF and involved in tubulogenesis and scattering. Indeed in glioblastoma cells and endothelial cells prolonged induction of ERK signaling by HGF promotes expression and secretion of MMP-2 and upregulation of MT1-MMP, a cell-surface activator of proMMP-2 [156,157]. Similarly in keratinocytes HGF induced scattering but not proliferation requires MMP-9 induction due to sustained activation of ERK kinases [158].
An emerging player in tubulogenesis is ARF6 which is necessary and sufficient to initiate tubule extension by both regulating the subcellular distribution of Rac1 to tubule extensions but also by inducing ERK mediated expression of the receptor for urokinase type plasminogen activator [159].
3.4.3. Balance between Proliferation and Apoptosis
The invasive growth program induced by HGF critically requires two other key features to succeed: increased resistance to apoptosis and enhanced proliferation.
The increased resistance to apoptosis is a typical feature of EMT and allows the invading epithelial cells to survive without the physiological survival signals provided by cell–cell and cell–matrix contacts, avoiding death for anoikia. HGF triggering of MET protects cells from apoptosis by using mainly the PI3K/Akt and, to a lesser extent, the MAPK pathways [160,161]. The PI3K-AKT pathway triggers Bad phosphorylation, thereby inactivating this pro-apoptotic protein, while simultaneously inducing expression of anti-apoptotic proteins such as Bcl-xL and Mcl-1 [162,163]. Furthermore PI3K/Akt, via the mTOR kinase, promote the translation and nuclear import of Mdm2, which inhibits TP53 activity both in vitro and in vivo [164]. MET induced resistance to apoptosis also allows tumor cells to resist to conditions that they face during tumor progression, i.e., nutrient deprivation or substrate detachment as well as chemotherapeutic treatment. HGF-driven survival of carcinoma requires the engagement of the PI3K/Akt/mTOR/S6K and ERK/MAPK transduction pathways, cooperatively preventing stress induced p38 activity [165].
Proliferation is also a typical response of many cell types to HGF triggering with an early peak of JUN and FOS transcription and activation (0.5–3 h) followed by Myc expression (6–8 h) and increased expression of cyclins A, B, D, and E (12 h) [166,167]. Thus, HGF promotes both increased transcription of AP-1 (FOS/JUN complex) and Myc early response genes [87] but also FOS mRNA translation via the PI3K/mTOR/4E-BP1 [168]. Nuclear factor kappa-B (NF-κB) is a multivalent transcription factor, which potentially controls the apoptosis/proliferation balance depending on the cellular context. HGF promotes NF-κB activation via both the PI3K/AKT and the Ras/MAPK pathway, mediating a survival signal [169] but also proliferation and morphogenesis [170].
Surprisingly HGF is a potent mitogen for a variety of cell types, but it is also known as an anti-mitogenic factor for several types of tumor cell lines. In HepG2 the high intensity ERK signal causes cell cycle arrest at G1 increasing the Cdk inhibitor p16-INK4a [171] and p21 [172], which mediates growth inhibition in the presence of HGF.
3.5. Negative Regulation of MET Signaling
As tyrosine phosphorylation is the key event in MET signaling, several researchers have worked to identify the tyrosine phosphatases (PTP) terminating receptor activation. PTP is a large family of more than 100 genes in humans comprising both soluble enzymes reversibly associating to protein targets and trans-membrane enzymes regulated by both extracellular and intracellular cues. MET phosphorylated in the activation loop is a substrate for cytosolic phosphatases such as PTP-1B and T-cell phosphatase [173]. Also PTP-S binds specifically to the juxtamembrane region of the activated receptor [174]. MET is similarly targeted by receptor-type protein tyrosine phosphatases such as: (1) RPTP-β, which dephosphorylates MET Y1356 and impairs MET tumorigenic activity [175,176]; (2) LAR, which counteracts MET auto-phosphorylation, as well as downstream MAPK and PI3K activation mediating contact inhibition [177,178]; (3) CD148, which dephosphorylates the tyrosines recruiting downstream effectors as well as the associated signal transducers GAB1 and p120 catenin [179].
An independent way of regulating MET activity resides in the juxtamembrane region which contains a cluster of serine/threonine phosphorylation sites (Figure 1 and Table 1). The best characterized of these sites is S985 which is phosphorylated by PKC-δ and -ε and dephosphorylated by protein phosphatase 2A [180]. S985 phosphorylation event inhibits the ligand-induced tyrosine auto-phosphorylation of the receptor and the receptor tyrosine kinase activity on exogenous substrates [181]. The relevance of such region is underscored by the existence of a splicing isoform of MET without such region and endowed with enhanced transforming ability [41,182].
MET and its associated proteins undergo ligand induced internalization that allows efficient signaling from endosomes but also couple with degradation of both receptor and ligand [183]. Indeed, upon ligand binding, the MET-HGF complex is rapidly internalized and MET become polyubiquitinated by Cbl. Cbl is an E3 ubiquitin ligase that associate in the juxtamembrane region of MET upon Y1003 phosphorylation and is phosphorylated upon HGF stimulation. A complex comprising Cbl, the adaptor CIN85 and endophilin promotes the import of MET in multivescicolar bodies trough clatrin coated pits. In this process Cbl is not a mere negative regulator of MET as it also acts as a scaffold for effectors recruitment in endosomes [184]. From the multivescicular body the receptor/ligand complex eventually recycles to the cell surface or is degraded in the lysosomes [185,186]. To the control of ubiquitinated-MET recycling cooperates Hrs, an early endosomal protein that is rapidly tyrosine-phosphorylated in cells stimulated with growth factors. Hrs couples with Stam to constitute the ESCRT complex that controls the initial selection of ubiquitinated proteins into clathrin-coated microdomains of early endosomes. Hrs also promotes receptor traffic toward multivesicular bodies/lysosomes by interacting with sorting nexin1 [187,188]. Conversely, proteasome activity is required for MET internalization and only indirectly for its degradation [189].
4. Signaling Integration by Met Multi-Receptor Complexes
MET has shown the remarkable property to associate with other signaling molecules to form complexes with several other receptors, effectively working as platforms for signal integration and amplification.
MET selectively associates with α6β4 integrin at the plasma membrane contributing to promote invasive growth, independently from laminin binding. CD151, a transmembrane protein of the tetraspanin family is a critical components of the complexes between MET and β4 integrin [190]. Following MET activation, α6β4 is tyrosine phosphorylated and combines with SHC, PI3K and SRC, generating an additional signaling platform that potentiates HGF-triggered activation of Ras- and PI3K-dependent pathways [191], promoting invasion [192] and anchorage independent growth [193]. This association between MET and integrins seems not to be an isolated instance as fibronectin binding to α5β1 integrin leads to a direct association of α5-integrin with MET, activating it in a HGF independent manner and promoting activation of SRC and FAK [194].
Semaphorins are cell surface and soluble signals that control directed migration and axonal guidance by binding to plexins receptors. The SEMA domain in the extracellular part of MET has strong homology to both plexins and semaphorins [195], suggesting a possible interaction between the two receptor families. Indeed Plexin B1 and MET associate in a complex and activation of Plexin B1 by SEMA 4D stimulates the tyrosine kinase activity of METMET, resulting in tyrosine phosphorylation of both receptors and downstream signaling [196,197,198]. Similarly to Plexins also Neuropilins acts as receptors for semaphorins and Neuropilin-1 associates with MET promoting its HGF induced activation and cell invasiveness [199]. Furthermore both Neuropilin-1 and Neuropilin-2 bind HGF, potentially acting as MET co-receptors [200].
A CD44 isoform containing variant exon v6 sequences (CD44v6) is strictly required for MET activation by HGF/SF, independently from HS modification of CD44. Autophosphorylation of MET requires the formation of a complex formed by HGF, MET and CD44v6. In this complex CD44v6 cytoplasmic tail presents binding motif for ezrin, radixin and moesin (ERM) proteins [201], which are phosphorylated by MET [202]. In this complex association of ERM proteins with CD44v6 and their link to the actin cytoskeleton is absolutely required to mediate the HGF-dependent activation of Ras, indicating a scaffolding function of cytoskeleton in HGF signaling [203]. CD44v6 mediated ERM binding to MET also participates to HGF induced receptor internalization [204]. Of note, CD44v6 is a marker of cancer stem cells, functionally cooperating with MET to promote PI3K dependent metastatic growth [205].
Those studies are just examples of how MET cooperates with a variety of other receptor for signaling integration. Of particular relevance for metabolism is the cooperation with insulin signaling. Indeed, in hepatic cells, MET form a complex with insulin receptor which respond to HGF triggering with trans-phosphorylation of the insulin receptor, recruitment of IRS1/2, stimulation of hepatic glucose uptake and suppression of hepatic glucose output [206]. MET also promotes PI3K dependent glucose uptake and glucose utilization from adipocytes [207], skeletal muscle cells [208] and participates to glucose homeostasis in vivo [206]. This is not an isolated example as MET can also be trans-activated and co-immunoprecipitated by other growth factor receptors such as RON [209,210], EGFR [211], HER2, HER3, and RET [212]. Hetrodimerization between growth factor receptors offers a platform to support signal integration, however in few cases the molecular bases of clustering and their relevance for signaling were not fully understood.
5. Conclusions
Thirty years of studies have explored in depth the signaling pathways promoted by the two simple tyrosines in MET cytoplasmic tail. However, the multiple intersections of those pathways as well as the interactions of MET with other transmembrane transducers point to the necessity of switching the approach from a signaling pathway oriented view to a network approach. In such framework a quantitative evaluation of signaling coupled to mathematical modeling could contribute to clarify how the cells integrates MET signaling in its biological context.
Indeed the fascinating question that drove the HGF-MET research still persists: How does a single factor—single receptor couple exerts a pleiotropy of biological effects in a context dependent manner?
Acknowledgments
Andrea Graziani is supported by AIRC, Italian Association for Cancer Research, (IG 13524 and IG 5392 grants) www.airc.it, and CARIPLO Foundation (2010–0737 grant), www.fondazionecariplo.it.
Gianluca Baldanzi is supported by University Piemonte Orientale (Young Investigators), www.unipmn.it. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Abbreviations
| Short name (used in the manuscript) | Full name (Uniprot) |
| HGF | Hepatocyte growth factor |
| MET | Hepatocyte growth factor receptor |
| MSP | Hepatocyte growth factor-like protein |
| RON | Macrophage-stimulating protein receptor |
| CD44v3 | CD44 antigen including variant exon 3 |
| CD44v6 | CD44 antigen including variant exon 6 |
| InlB | Internalin B |
| GRB2 | growth factor receptor-bound protein 2 |
| PLCγ | 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase gamma |
| PI3K | Phosphatidylinositol 4,5-bisphosphate 3-kinase |
| SHP-2 | tyrosine-protein phosphatase non-receptor type 11 |
| SHC | SHC-transforming protein |
| GAB1 | GRB2-associated binding protein |
| MAPK | mitogen-activated protein kinase |
| SOS | Son of sevenless homolog |
| NCK | cytoplasmic protein NCK |
| CRK-I and CRK-II | adapter molecule crk |
| C3G | Rap guanine nucleotide exchange factor 1 |
| N-WASP | Neural Wiskott-Aldrich syndrome protein |
| PKD | Serine/threonine-protein kinase D |
| AKT | RAC-alpha serine/threonine-protein kinase |
| PKC | Protein kinase C |
| PLD | Phospholipase D |
| PAP | Phosphatidic acid phosphohydrolase |
| JUN | Transcription factor AP-1 |
| FOS | Proto-oncogene c-Fos |
| SRC | proto-oncogene tyrosine-protein kinase SRC |
| FYN | Tyrosine-protein kinase Fyn |
| FAK | Focal adhesion kinase |
| αDGK | Diacylglycerol kinase alpha |
| RhoGDI | Rho GDP-dissociation inhibitor |
| RCP | Rab11 family-interacting protein 1 |
| STAT3 | Signal transducer and activator of transcription 3 |
| SHIP-1 | SH2 domain-containing inositol 5-phosphatases 1 |
| SHIP-2 | SH2 domain-containing inositol 5-phosphatases 2 |
| PLA2 | Phospholipase A2 |
| COX-2 | Prostaglandin G/H synthase 2 |
| CREB | Cyclic AMP-responsive element-binding protein |
| RB | Retinoblastoma-associated protein |
| JNK | Mitogen-activated protein kinase 8 |
| MEK-1 | Dual specificity mitogen-activated protein kinase kinase 1 |
| MEK-2 | Dual specificity mitogen-activated protein kinase kinase 2 |
| ERK1 | Mitogen-activated protein kinase 3 |
| ERK2 | Mitogen-activated protein kinase 1 |
| MEK3 | Dual specificity mitogen-activated protein kinase kinase 3 |
| MEK6 | Dual specificity mitogen-activated protein kinase kinase 6 |
| PKR | Protein kinase R |
| p38α | Mitogen-activated protein kinase 14 |
| CDK1 | Cyclin-dependent kinase 1 |
| Snail | Zinc finger protein SNAI1 |
| Slug | Zinc finger protein SNAI2 |
| Egr1 | early growth response protein 1 |
| Elk-1 | ETS domain-containing protein Elk-1 |
| S6K | Ribosomal protein S6 kinase beta |
| GSK3 | Glycogen synthase kinase-3 |
| Lef | Lymphoid enhancer-binding factor |
| PAK | Serine/threonine-protein kinase PAK 1 |
| ROCK | Rho-associated protein kinase |
| MT1-MMP | Matrix metalloproteinase-14 |
| BAD | Bcl2-associated agonist of cell death |
| Bcl-xL | Bcl-2-like protein 1 (long isoform) |
| Mcl-1 | Induced myeloid leukemia cell differentiation protein Mcl-1 |
| mTOR | Serine/threonine-protein kinase mTOR |
| Mdm2 | E3 ubiquitin-protein ligase Mdm2 |
| TP53 | Cellular tumor antigen p53 |
| MYC | Myc proto-oncogene protein |
| 4E-BP1 | Eukaryotic translation initiation factor 4E-binding protein 1 |
| p16-INK4a | Cyclin-dependent kinase inhibitor 2A |
| p21 | Cyclin-dependent kinase inhibitor 1 |
| PTP-1B | Tyrosine-protein phosphatase non-receptor type 1 |
| RPTP-β | Receptor-type tyrosine-protein phosphatase B |
| LAR | Receptor-type tyrosine-protein phosphatase F |
| CD148 | Receptor-type tyrosine-protein phosphatase eta |
| Cbl | E3 ubiquitin-protein ligase CBL |
| CIN85 | SH3 domain-containing kinase-binding protein 1 |
| Hrs | Hepatocyte growth factor-regulated tyrosine kinase substrate |
| Stam | Signal transducing adapter molecule |
| IRS1 | Insulin receptor substrate 1 |
| IRS2 | Insulin receptor substrate 2 |
| EGFR | Epidermal growth factor receptor |
| HER2 | Receptor tyrosine-protein kinase erbB-2 |
| HER3 | Receptor tyrosine-protein kinase erbB-3 |
| RET | Proto-oncogene tyrosine-protein kinase receptor Ret |
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nakamura, T.; Teramoto, H.; Ichihara, A. Purification and characterization of a growth factor from rat platelets for mature parenchymal hepatocytes in primary cultures. Proc. Natl. Acad. Sci. USA 1986, 83, 6489–6493. [Google Scholar] [CrossRef] [PubMed]
- Stoker, M.; Perryman, M. An epithelial scatter factor released by embryo fibroblasts. J. Cell Sci. 1985, 77, 209–223. [Google Scholar] [PubMed]
- Naldini, L.; Weidner, K.M.; Vigna, E.; Gaudino, G.; Bardelli, A.; Ponzetto, C.; Narsimhan, R.P.; Hartmann, G.; Zarnegar, R.; Michalopoulos, G.K. Scatter factor and hepatocyte growth factor are indistinguishable ligands for the met receptor. EMBO J. 1991, 10, 2867–2878. [Google Scholar] [PubMed]
- Sonnenberg, E.; Meyer, D.; Weidner, K.M.; Birchmeier, C. Scatter factor/hepatocyte growth factor and its receptor, the c-met tyrosine kinase, can mediate a signal exchange between mesenchyme and epithelia during mouse development. J. Cell Biol. 1993, 123, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Nishizawa, T.; Hagiya, M.; Seki, T.; Shimonishi, M.; Sugimura, A.; Tashiro, K.; Shimizu, S. Molecular cloning and expression of human hepatocyte growth factor. Nature 1989, 342, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Hagiya, M.; Shimonishi, M.; Nakamura, T.; Shimizu, S. Organization of the human hepatocyte growth factor-encoding gene. Gene 1991, 102, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Kirchhofer, D.; Yao, X.; Peek, M.; Eigenbrot, C.; Lipari, M.T.; Billeci, K.L.; Maun, H.R.; Moran, P.; Santell, L.; Wiesmann, C.; et al. Structural and functional basis of the serine protease-like hepatocyte growth factor beta-chain in met binding and signaling. J. Biol. Chem. 2004, 279, 39915–39924. [Google Scholar] [CrossRef] [PubMed]
- Stamos, J.; Lazarus, R.A.; Yao, X.; Kirchhofer, D.; Wiesmann, C. Crystal structure of the HGF beta-chain in complex with the sema domain of the met receptor. EMBO J. 2004, 23, 2325–2335. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K.; Tsubouchi, H.; Naka, D.; Takahashi, K.; Okigaki, M.; Arakaki, N.; Nakayama, H.; Hirono, S.; Sakiyama, O. Molecular cloning and sequence analysis of cdna for human hepatocyte growth factor. Biochem. Biophys. Res. Commun. 1989, 163, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, K.; Hagiya, M.; Nishizawa, T.; Seki, T.; Shimonishi, M.; Shimizu, S.; Nakamura, T. Deduced primary structure of rat hepatocyte growth factor and expression of the mrna in rat tissues. Proc. Natl. Acad. Sci. USA 1990, 87, 3200–3204. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, T.; Yuhki, N.; Wang, M.H.; Skeel, A.; Leonard, E.J. Cloning, sequencing, and expression of human macrophage stimulating protein (msp, mst1) confirms msp as a member of the family of kringle proteins and locates the MSP gene on chromosome 3. J. Biol. Chem. 1993, 268, 15461–15468. [Google Scholar] [PubMed]
- Stahl, S.J.; Wingfield, P.T.; Kaufman, J.D.; Pannell, L.K.; Cioce, V.; Sakata, H.; Taylor, W.G.; Rubin, J.S.; Bottaro, D.P. Functional and biophysical characterization of recombinant human hepatocyte growth factor isoforms produced in Escherichia coli. Biochem. J. 1997, 326, 763–772. [Google Scholar] [PubMed]
- Cioce, V.; Csaky, K.G.; Chan, A.M.; Bottaro, D.P.; Taylor, W.G.; Jensen, R.; Aaronson, S.A.; Rubin, J.S. Hepatocyte growth factor (HGF)/nk1 is a naturally occurring hgf/scatter factor variant with partial agonist/antagonist activity. J. Biol. Chem. 1996, 271, 13110–13115. [Google Scholar] [CrossRef] [PubMed]
- Montesano, R.; Soriano, J.V.; Malinda, K.M.; Ponce, M.L.; Bafico, A.; Kleinman, H.K.; Bottaro, D.P.; Aaronson, S.A. Differential effects of hepatocyte growth factor isoforms on epithelial and endothelial tubulogenesis. Cell Growth Differ. 1998, 9, 355–365. [Google Scholar] [PubMed]
- Chan, A.M.; Rubin, J.S.; Bottaro, D.P.; Hirschfield, D.W.; Chedid, M.; Aaronson, S.A. Identification of a competitive HGF antagonist encoded by an alternative transcript. Science 1991, 254, 1382–1385. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, T.; Jakubczak, J.; Vieira, W.; Bottaro, D.P.; Breckenridge, D.; Larochelle, W.J.; Merlino, G. Disassociation of met-mediated biological responses in vivo: The natural hepatocyte growth factor/scatter factor splice variant nk2 antagonizes growth but facilitates metastasis. Mol. Cell. Biol. 2000, 20, 2055–2065. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Nakae, Y.; Sogabe, T.; Ihara, I.; Ueno, S.; Sakai, H.; Inoue, H.; Shimizu, S.; Nakamura, T.; Shimizu, N. Structural study of the n-linked oligosaccharides of hepatocyte growth factor by two-dimensional sugar mapping. J. Biochem. 1993, 114, 76–82. [Google Scholar] [PubMed]
- Fukuta, K.; Matsumoto, K.; Nakamura, T. Multiple biological responses are induced by glycosylation-deficient hepatocyte growth factor. Biochem. J. 2005, 388, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Nawa, K.; Ichihara, A.; Kaise, N.; Nishino, T. Purification and subunit structure of hepatocyte growth factor from rat platelets. FEBS Lett. 1987, 224, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Naka, D.; Ishii, T.; Yoshiyama, Y.; Miyazawa, K.; Hara, H.; Hishida, T.; Kidamura, N. Activation of hepatocyte growth factor by proteolytic conversion of a single chain form to a heterodimer. J. Biol. Chem. 1992, 267, 20114–20119. [Google Scholar] [PubMed]
- Weidner, K.M.; Behrens, J.; Vandekerckhove, J.; Birchmeier, W. Scatter factor: Molecular characteristics and effect on the invasiveness of epithelial cells. J. Cell. Biol. 1990, 111, 2097–2108. [Google Scholar] [CrossRef] [PubMed]
- Mars, W.M.; Zarnegar, R.; Michalopoulos, G.K. Activation of hepatocyte growth factor by the plasminogen activators upa and tpa. Am. J. Pathol. 1993, 143, 949–958. [Google Scholar] [PubMed]
- Miyazawa, K.; Shimomura, T.; Kitamura, A.; Kondo, J.; Morimoto, Y.; Kitamura, N. Molecular cloning and sequence analysis of the cdna for a human serine protease reponsible for activation of hepatocyte growth factor. Structural similarity of the protease precursor to blood coagulation factor XII. J. Biol. Chem. 1993, 268, 10024–10028. [Google Scholar] [PubMed]
- Shimomura, T.; Miyazawa, K.; Komiyama, Y.; Hiraoka, H.; Naka, D.; Morimoto, Y.; Kitamura, N. Activation of hepatocyte growth factor by two homologous proteases, blood-coagulation factor XIIa and hepatocyte growth factor activator. Eur. J. Biochem. 1995, 229, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.L.; Huang, P.Y.; Roller, P.; Cho, E.G.; Park, D.; Dickson, R.B. Matriptase/epithin participates in mammary epithelial cell growth and morphogenesis through hgf activation. Mech. Dev. 2010, 127, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Kirchhofer, D.; Peek, M.; Lipari, M.T.; Billeci, K.; Fan, B.; Moran, P. Hepsin activates pro-hepatocyte growth factor and is inhibited by hepatocyte growth factor activator inhibitor-1b (hai-1b) and hai-2. FEBS Lett. 2005, 579, 1945–1950. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K.; Shimomura, T.; Kitamura, N. Activation of hepatocyte growth factor in the injured tissues is mediated by hepatocyte growth factor activator. J. Biol. Chem. 1996, 271, 3615–3618. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Inoue, H.; Hagiya, M.; Shimizu, S.; Nose, T.; Shimohigashi, Y.; Nakamura, T. Hairpin loop and second kringle domain are essential sites for heparin binding and biological activity of hepatocyte growth factor. J. Biol. Chem. 1994, 269, 1131–1136. [Google Scholar] [PubMed]
- Tajima, H.; Matsumoto, K.; Nakamura, T. Regulation of cell growth and motility by hepatocyte growth factor and receptor expression in various cell species. Exp. Cell Res. 1992, 202, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Lyon, M.; Deakin, J.A.; Gallagher, J.T. The mode of action of heparan and dermatan sulfates in the regulation of hepatocyte growth factor/scatter factor. J. Biol. Chem. 2002, 277, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Lyon, M.; Deakin, J.A.; Rahmoune, H.; Fernig, D.G.; Nakamura, T.; Gallagher, J.T. Hepatocyte growth factor/scatter factor binds with high affinity to dermatan sulfate. J. Biol. Chem. 1998, 273, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Honke, K.; Miyazaki, T.; Matsumoto, K.; Nakamura, T.; Ishizuka, I.; Makita, A. Hepatocyte growth factor specifically binds to sulfoglycolipids. J. Biol. Chem. 1994, 269, 9817–9821. [Google Scholar] [PubMed]
- Chirgadze, D.Y.; Hepple, J.P.; Zhou, H.; Byrd, R.A.; Blundell, T.L.; Gherardi, E. Crystal structure of the nk1 fragment of HGF/SF suggests a novel mode for growth factor dimerization and receptor binding. Nat. Struct. Biol. 1999, 6, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Sakata, H.; Stahl, S.J.; Taylor, W.G.; Rosenberg, J.M.; Sakaguchi, K.; Wingfield, P.T.; Rubin, J.S. Heparin binding and oligomerization of hepatocyte growth factor/scatter factor isoforms. Heparan sulfate glycosaminoglycan requirement for met binding and signaling. J. Biol. Chem. 1997, 272, 9457–9463. [Google Scholar] [CrossRef] [PubMed]
- Tolbert, W.D.; Daugherty, J.; Gao, C.; Xie, Q.; Miranti, C.; Gherardi, E.; Vande Woude, G.; Xu, H.E. A mechanistic basis for converting a receptor tyrosine kinase agonist to an antagonist. Proc. Natl. Acad. Sci. USA 2007, 104, 14592–14597. [Google Scholar] [CrossRef] [PubMed]
- Van der Voort, R.; Taher, T.E.; Wielenga, V.J.; Spaargaren, M.; Prevo, R.; Smit, L.; David, G.; Hartmann, G.; Gherardi, E.; Pals, S.T. Heparan sulfate-modified cd44 promotes hepatocyte growth factor/scatter factor-induced signal transduction through the receptor tyrosine kinase c-met. J. Biol. Chem. 1999, 274, 6499–6506. [Google Scholar]
- Rubin, J.S.; Day, R.M.; Breckenridge, D.; Atabey, N.; Taylor, W.G.; Stahl, S.J.; Wingfield, P.T.; Kaufman, J.D.; Schwall, R.; Bottaro, D.P. Dissociation of heparan sulfate and receptor binding domains of hepatocyte growth factor reveals that heparan sulfate-c-met interaction facilitates signaling. J. Biol. Chem. 2001, 276, 32977–32983. [Google Scholar] [CrossRef] [PubMed]
- Gherardi, E.; Youles, M.E.; Miguel, R.N.; Blundell, T.L.; Iamele, L.; Gough, J.; Bandyopadhyay, A.; Hartmann, G.; Butler, P.J. Functional map and domain structure of met, the product of the c-met protooncogene and receptor for hepatocyte growth factor/scatter factor. Proc. Natl. Acad. Sci. USA 2003, 100, 12039–12044. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.; Goldschneider, I. Cutting edge: Identification of a hybrid cytokine consisting of il-7 and the beta-chain of the hepatocyte growth factor/scatter factor. J. Immunol. 2001, 167, 3550–3554. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, G.A.; Naujokas, M.A.; Park, M. Alternative splicing generates isoforms of the met receptor tyrosine kinase which undergo differential processing. Mol. Cell Biol. 1991, 11, 2962–2970. [Google Scholar] [PubMed]
- Lee, C.C.; Yamada, K.M. Identification of a novel type of alternative splicing of a tyrosine kinase receptor. Juxtamembrane deletion of the c-met protein kinase c serine phosphorylation regulatory site. J. Biol. Chem. 1994, 269, 19457–19461. [Google Scholar] [PubMed]
- Prat, M.; Crepaldi, T.; Gandino, L.; Giordano, S.; Longati, P.; Comoglio, P. C-terminal truncated forms of met, the hepatocyte growth factor receptor. Mol. Cell. Biol. 1991, 11, 5954–5962. [Google Scholar] [PubMed]
- Crepaldi, T.; Prat, M.; Giordano, S.; Medico, E.; Comoglio, P.M. Generation of a truncated hepatocyte growth factor receptor in the endoplasmic reticulum. J. Biol. Chem. 1994, 269, 1750–1755. [Google Scholar] [PubMed]
- Wajih, N.; Walter, J.; Sane, D.C. Vascular origin of a soluble truncated form of the hepatocyte growth factor receptor (c-met). Circ. Res. 2002, 90, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Merlin, S.; Pietronave, S.; Locarno, D.; Valente, G.; Follenzi, A.; Prat, M. Deletion of the ectodomain unleashes the transforming, invasive, and tumorigenic potential of the met oncogene. Cancer Sci. 2009, 100, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Deheuninck, J.; Goormachtigh, G.; Foveau, B.; Ji, Z.; Leroy, C.; Ancot, F.; Villeret, V.; Tulasne, D.; Fafeur, V. Phosphorylation of the met receptor on juxtamembrane tyrosine residue 1001 inhibits its caspase-dependent cleavage. Cell Signal 2009, 21, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Prat, M.; Narsimhan, R.P.; Crepaldi, T.; Nicotra, M.R.; Natali, P.G.; Comoglio, P.M. The receptor encoded by the human c-met oncogene is expressed in hepatocytes, epithelial cells and solid tumors. Int. J. Cancer 1991, 49, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Di Renzo, M.F.; Narsimhan, R.P.; Olivero, M.; Bretti, S.; Giordano, S.; Medico, E.; Gaglia, P.; Zara, P.; Comoglio, P.M. Expression of the met/hgf receptor in normal and neoplastic human tissues. Oncogene 1991, 6, 1997–2003. [Google Scholar]
- Bussolino, F.; Di Renzo, M.F.; Ziche, M.; Bocchietto, E.; Olivero, M.; Naldini, L.; Gaudino, G.; Tamagnone, L.; Coffer, A.; Comoglio, P.M. Hepatocyte growth factor is a potent angiogenic factor which stimulates endothelial cell motility and growth. J. Cell. Biol. 1992, 119, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Nishino, T.; Hisha, H.; Nishino, N.; Adachi, M.; Ikehara, S. Hepatocyte growth factor as a hematopoietic regulator. Blood 1995, 85, 3093–3100. [Google Scholar] [PubMed]
- Taher, T.E.; Tjin, E.P.; Beuling, E.A.; Borst, J.; Spaargaren, M.; Pals, S.T. C-cbl is involved in met signaling in b cells and mediates hepatocyte growth factor-induced receptor ubiquitination. J. Immunol. 2002, 169, 3793–3800. [Google Scholar] [CrossRef] [PubMed]
- Stoker, M.; Gherardi, E.; Perryman, M.; Gray, J. Scatter factor is a fibroblast-derived modulator of epithelial cell mobility. Nature 1987, 327, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Shen, C.; Wang, H.; Shen, H.; Chen, Y.; Nie, A.; Yan, G.; Lu, H.; Liu, Y.; Yang, P. Identification of n-glycosylation sites on secreted proteins of human hepatocellular carcinoma cells with a complementary proteomics approach. J. Proteome Res. 2009, 8, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, G.; Perreault, A.; Schrag, J.D.; Park, M.; Cygler, M.; Gehring, K.; Ekiel, I. Insights into function of PSI domains from structure of the met receptor psi domain. Biochem. Biophys. Res. Commun. 2004, 321, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Niemann, H.H.; Jäger, V.; Butler, P.J.; van den Heuvel, J.; Schmidt, S.; Ferraris, D.; Gherardi, E.; Heinz, D.W. Structure of the human receptor tyrosine kinase met in complex with the listeria invasion protein inlb. Cell 2007, 130, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Komada, M.; Hatsuzawa, K.; Shibamoto, S.; Ito, F.; Nakayama, K.; Kitamura, N. Proteolytic processing of the hepatocyte growth factor/scatter factor receptor by furin. FEBS Lett. 1993, 328, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Ronsin, C.; Muscatelli, F.; Mattei, M.G.; Breathnach, R. A novel putative receptor protein tyrosine kinase of the met family. Oncogene 1993, 8, 1195–1202. [Google Scholar] [PubMed]
- Bottaro, D.P.; Rubin, J.S.; Faletto, D.L.; Chan, A.M.; Kmiecik, T.E.; Vande Woude, G.F.; Aaronson, S.A. Identification of the hepatocyte growth factor receptor as the c-met proto-oncogene product. Science 1991, 251, 802–804. [Google Scholar] [CrossRef] [PubMed]
- Naldini, L.; Vigna, E.; Narsimhan, R.P.; Gaudino, G.; Zarnegar, R.; Michalopoulos, G.K.; Comoglio, P.M. Hepatocyte growth factor (hgf) stimulates the tyrosine kinase activity of the receptor encoded by the proto-oncogene c-met. Oncogene 1991, 6, 501–504. [Google Scholar] [PubMed]
- Shen, Y.; Naujokas, M.; Park, M.; Ireton, K. Inib-dependent internalization of listeria is mediated by the met receptor tyrosine kinase. Cell 2000, 103, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Youles, M.; Holmes, O.; Petoukhov, M.V.; Nessen, M.A.; Stivala, S.; Svergun, D.I.; Gherardi, E. Engineering the nk1 fragment of hepatocyte growth factor/scatter factor as a met receptor antagonist. J. Mol. Biol. 2008, 377, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Basilico, C.; Arnesano, A.; Galluzzo, M.; Comoglio, P.M.; Michieli, P. A high affinity hepatocyte growth factor-binding site in the immunoglobulin-like region of met. J. Biol. Chem. 2008, 283, 21267–21277. [Google Scholar] [CrossRef] [PubMed]
- Kong-Beltran, M.; Stamos, J.; Wickramasinghe, D. The sema domain of met is necessary for receptor dimerization and activation. Cancer Cell 2004, 6, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Komada, M.; Kitamura, N. Regulatory role of major tyrosine autophosphorylation site of kinase domain of c-met receptor (scatter factor/hepatocyte growth factor receptor). J. Biol. Chem. 1994, 269, 16131–16136. [Google Scholar] [PubMed]
- Zhen, Z.; Giordano, S.; Longati, P.; Medico, E.; Campiglio, M.; Comoglio, P.M. Structural and functional domains critical for constitutive activation of the HGF-receptor (met). Oncogene 1994, 9, 1691–1697. [Google Scholar] [PubMed]
- Longati, P.; Bardelli, A.; Ponzetto, C.; Naldini, L.; Comoglio, P.M. Tyrosines1234–1235 are critical for activation of the tyrosine kinase encoded by the met proto-oncogene (HGF receptor). Oncogene 1994, 9, 49–57. [Google Scholar] [PubMed]
- Ferracini, R.; Longati, P.; Naldini, L.; Vigna, E.; Comoglio, P.M. Identification of the major autophosphorylation site of the met/hepatocyte growth factor receptor tyrosine kinase. J. Biol. Chem. 1991, 266, 19558–19564. [Google Scholar] [PubMed]
- Ponzetto, C.; Bardelli, A.; Zhen, Z.; Maina, F.; dalla Zonca, P.; Giordano, S.; Graziani, A.; Panayotou, G.; Comoglio, P.M. A multifunctional docking site mediates signaling and transformation by the hepatocyte growth factor/scatter factor receptor family. Cell 1994, 77, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Bardelli, A.; Longati, P.; Williams, T.A.; Benvenuti, S.; Comoglio, P.M. A peptide representing the carboxyl-terminal tail of the met receptor inhibits kinase activity and invasive growth. J. Biol. Chem. 1999, 274, 29274–29281. [Google Scholar] [CrossRef] [PubMed]
- Fournier, T.M.; Kamikura, D.; Teng, K.; Park, M. Branching tubulogenesis but not scatter of madin-darby canine kidney cells requires a functional grb2 binding site in the met receptor tyrosine kinase. J. Biol. Chem. 1996, 271, 22211–22217. [Google Scholar] [CrossRef] [PubMed]
- Royal, I.; Park, M. Hepatocyte growth factor-induced scatter of madin-darby canine kidney cells requires phosphatidylinositol 3-kinase. J. Biol. Chem. 1995, 270, 27780–27787. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Naujokas, M.A.; Fixman, E.D.; Torossian, K.; Park, M. Tyrosine 1356 in the carboxyl-terminal tail of the hgf/sf receptor is essential for the transduction of signals for cell motility and morphogenesis. J. Biol. Chem. 1994, 269, 29943–29948. [Google Scholar] [PubMed]
- Pelicci, G.; Giordano, S.; Zhen, Z.; Salcini, A.E.; Lanfrancone, L.; Bardelli, A.; Panayotou, G.; Waterfield, M.D.; Ponzetto, C.; Pelicci, P.G. The motogenic and mitogenic responses to HGF are amplified by the shc adaptor protein. Oncogene 1995, 10, 1631–1638. [Google Scholar] [PubMed]
- Maina, F.; Casagranda, F.; Audero, E.; Simeone, A.; Comoglio, P.M.; Klein, R.; Ponzetto, C. Uncoupling of grb2 from the met receptor in vivo reveals complex roles in muscle development. Cell 1996, 87, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Ponzetto, C.; Zhen, Z.; Audero, E.; Maina, F.; Bardelli, A.; Basile, M.L.; Giordano, S.; Narsimhan, R.; Comoglio, P. Specific uncoupling of grb2 from the met receptor. Differential effects on transformation and motility. J. Biol. Chem. 1996, 271, 14119–14123. [Google Scholar] [CrossRef] [PubMed]
- Giordano, S.; Bardelli, A.; Zhen, Z.; Menard, S.; Ponzetto, C.; Comoglio, P.M. A point mutation in the met oncogene abrogates metastasis without affecting transformation. Proc. Natl. Acad. Sci. USA 1997, 94, 13868–13872. [Google Scholar] [CrossRef] [PubMed]
- Bardelli, A.; Longati, P.; Gramaglia, D.; Stella, M.C.; Comoglio, P.M. Gab1 coupling to the hgf/met receptor multifunctional docking site requires binding of grb2 and correlates with the transforming potential. Oncogene 1997, 15, 3103–3111. [Google Scholar] [CrossRef] [PubMed]
- Weidner, K.M.; Di Cesare, S.; Sachs, M.; Brinkmann, V.; Behrens, J.; Birchmeier, W. Interaction between gab1 and the c-met receptor tyrosine kinase is responsible for epithelial morphogenesis. Nature 1996, 384, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.; Holgado-Madruga, M.; Maroun, C.; Fixman, E.D.; Kamikura, D.; Fournier, T.; Charest, A.; Tremblay, M.L.; Wong, A.J.; Park, M. Association of the multisubstrate docking protein gab1 with the hepatocyte growth factor receptor requires a functional grb2 binding site involving tyrosine 1356. J. Biol. Chem. 1997, 272, 20811–20819. [Google Scholar] [CrossRef] [PubMed]
- Maroun, C.R.; Holgado-Madruga, M.; Royal, I.; Naujokas, M.A.; Fournier, T.M.; Wong, A.J.; Park, M. The gab1 ph domain is required for localization of gab1 at sites of cell-cell contact and epithelial morphogenesis downstream from the met receptor tyrosine kinase. Mol. Cell. Biol. 1999, 19, 1784–1799. [Google Scholar] [PubMed]
- Gual, P.; Giordano, S.; Williams, T.A.; Rocchi, S.; van Obberghen, E.; Comoglio, P.M. Sustained recruitment of phospholipase c-gamma to gab1 is required for hgf-induced branching tubulogenesis. Oncogene 2000, 19, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Schaeper, U.; Gehring, N.H.; Fuchs, K.P.; Sachs, M.; Kempkes, B.; Birchmeier, W. Coupling of gab1 to c-met, grb2, and shp2 mediates biological responses. J. Cell. Biol. 2000, 149, 1419–1432. [Google Scholar] [CrossRef] [PubMed]
- Sakkab, D.; Lewitzky, M.; Posern, G.; Schaeper, U.; Sachs, M.; Birchmeier, W.; Feller, S.M. Signaling of hepatocyte growth factor/scatter factor (HGF) to the small gtpase rap1 via the large docking protein gab1 and the adapter protein crkl. J. Biol. Chem. 2000, 275, 10772–10778. [Google Scholar] [CrossRef] [PubMed]
- Lamorte, L.; Royal, I.; Naujokas, M.; Park, M. Crk adapter proteins promote an epithelial-mesenchymal-like transition and are required for hgf-mediated cell spreading and breakdown of epithelial adherens junctions. Mol. Biol. Cell 2002, 13, 1449–1461. [Google Scholar] [CrossRef] [PubMed]
- Maroun, C.R.; Naujokas, M.A.; Holgado-Madruga, M.; Wong, A.J.; Park, M. The tyrosine phosphatase shp-2 is required for sustained activation of extracellular signal-regulated kinase and epithelial morphogenesis downstream from the met receptor tyrosine kinase. Mol. Cell. Biol. 2000, 20, 8513–8525. [Google Scholar] [CrossRef] [PubMed]
- Kodama, A.; Matozaki, T.; Fukuhara, A.; Kikyo, M.; Ichihashi, M.; Takai, Y. Involvement of an shp-2-rho small G protein pathway in hepatocyte growth factor/scatter factor-induced cell scattering. Mol. Biol. Cell 2000, 11, 2565–2575. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Kochhar, K.; Nakamura, T.; Iyer, A. Hepatocyte growth factor-induced signal transduction in two normal mouse epithelial cell lines. Biochem. Mol. Biol. Int. 1995, 36, 465–474. [Google Scholar] [PubMed]
- Kochhar, K.S.; Iyer, A.P. Hepatocyte growth factor induces activation of nck and phospholipase c-gamma in lung carcinoma cells. Cancer Lett. 1996, 104, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Buday, L.; Wunderlich, L.; Tamás, P. The nck family of adapter proteins: Regulators of actin cytoskeleton. Cell Signal 2002, 14, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Abella, J.V.; Vaillancourt, R.; Frigault, M.M.; Ponzo, M.G.; Zuo, D.; Sangwan, V.; Larose, L.; Park, M. The gab1 scaffold regulates rtk-dependent dorsal ruffle formation through the adaptor nck. J. Cell Sci. 2010, 123, 1306–1319. [Google Scholar] [CrossRef] [PubMed]
- Graziani, A.; Gramaglia, D.; Cantley, L.C.; Comoglio, P.M. The tyrosine-phosphorylated hepatocyte growth factor/scatter factor receptor associates with phosphatidylinositol 3-kinase. J. Biol. Chem. 1991, 266, 22087–22090. [Google Scholar] [PubMed]
- Bardelli, A.; Maina, F.; Gout, I.; Fry, M.J.; Waterfield, M.D.; Comoglio, P.M.; Ponzetto, C. Autophosphorylation promotes complex formation of recombinant hepatocyte growth factor receptor with cytoplasmic effectors containing sh2 domains. Oncogene 1992, 7, 1973–1978. [Google Scholar] [PubMed]
- Ponzetto, C.; Bardelli, A.; Maina, F.; Longati, P.; Panayotou, G.; Dhand, R.; Waterfield, M.D.; Comoglio, P.M. A novel recognition motif for phosphatidylinositol 3-kinase binding mediates its association with the hepatocyte growth factor/scatter factor receptor. Mol. Cell. Biol. 1993, 13, 4600–4608. [Google Scholar] [PubMed]
- Derman, M.P.; Cunha, M.J.; Barros, E.J.; Nigam, S.K.; Cantley, L.G. Hgf-mediated chemotaxis and tubulogenesis require activation of the phosphatidylinositol 3-kinase. Am. J. Physiol. 1995, 268, F1211–F1217. [Google Scholar] [PubMed]
- Skouteris, G.G.; Georgakopoulos, E. Hepatocyte growth factor-induced proliferation of primary hepatocytes is mediated by activation of phosphatidylinositol 3-kinase. Biochem. Biophys. Res. Commun. 1996, 218, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Osada, S.; Nakashima, S.; Saji, S.; Nakamura, T.; Nozawa, Y. Hepatocyte growth factor (HGF) mediates the sustained formation of 1,2-diacylglycerol via phosphatidylcholine-phospholipase c in cultured rat hepatocytes. FEBS Lett. 1992, 297, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, A.; Hayashi, N.; Tsubouchi, H.; Tanaka, Y.; Ito, T.; Sasaki, Y.; Fusamoto, H.; Daikuhara, Y.; Kamada, T. Intracellular calcium as a second messenger for human hepatocyte growth factor in hepatocytes. Hepatology 1992, 15, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Machide, M.; Kamitori, K.; Nakamura, Y.; Kohsaka, S. Selective activation of phospholipase c gamma1 and distinct protein kinase c subspecies in intracellular signaling by hepatocyte growth factor/scatter factor in primary cultured rat neocortical cells. J. Neurochem. 1998, 71, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, V.; King, R.J. Pkc, p42/p44 mapk, and p38 mapk are required for hgf-induced proliferation of h441 cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2000, 279, L942–L949. [Google Scholar] [PubMed]
- Adachi, T.; Nakashima, S.; Saji, S.; Nakamura, T.; Nozawa, Y. Phospholipase d activation in hepatocyte growth factor-stimulated rat hepatocytes mediates the expressions of c-jun and c-fos: Involvement of protein tyrosine kinase, protein kinase c, and Ca2+. Hepatology 1996, 24, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, N.; Hung, W.; Tremblay, E.; Saulnier, R.; Elliott, B. C-src kinase activity is required for hepatocyte growth factor-induced motility and anchorage-independent growth of mammary carcinoma cells. J. Biol. Chem. 1998, 273, 33714–33721. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.C.; Chen, Y.L.; Cheng, C.H.; Yu, K.C.; Cary, L.A.; Shu, K.H.; Ho, W.L.; Chen, H.C. Src phosphorylates grb2-associated binder 1 upon hepatocyte growth factor stimulation. J. Biol. Chem. 2003, 278, 44075–44082. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Chan, P.C.; Chen, C.L.; Chen, H.C. Phosphorylation of focal adhesion kinase on tyrosine 194 by met leads to its activation through relief of autoinhibition. Oncogene 2011, 30, 153–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.C.; Chan, P.C.; Tang, M.J.; Cheng, C.H.; Chang, T.J. Tyrosine phosphorylation of focal adhesion kinase stimulated by hepatocyte growth factor leads to mitogen-activated protein kinase activation. J. Biol. Chem. 1998, 273, 25777–25782. [Google Scholar] [CrossRef] [PubMed]
- Baldanzi, G.; Cutrupi, S.; Chianale, F.; Gnocchi, V.; Rainero, E.; Porporato, P.; Filigheddu, N.; van Blitterswijk, W.J.; Parolini, O.; Bussolino, F.; et al. Diacylglycerol kinase-alpha phosphorylation by src on y335 is required for activation, membrane recruitment and hgf-induced cell motility. Oncogene 2008, 27, 942–956. [Google Scholar] [CrossRef] [PubMed]
- Cutrupi, S.; Baldanzi, G.; Gramaglia, D.; Maffè, A.; Schaap, D.; Giraudo, E.; van Blitterswijk, W.; Bussolino, F.; Comoglio, P.M.; Graziani, A. Src-mediated activation of alpha-diacylglycerol kinase is required for hepatocyte growth factor-induced cell motility. EMBO J. 2000, 19, 4614–4622. [Google Scholar] [CrossRef] [PubMed]
- Chianale, F.; Rainero, E.; Cianflone, C.; Bettio, V.; Pighini, A.; Porporato, P.E.; Filigheddu, N.; Serini, G.; Sinigaglia, F.; Baldanzi, G.; et al. Diacylglycerol kinase alpha mediates hgf-induced rac activation and membrane ruffling by regulating atypical pkc and rhogdi. Proc. Natl. Acad. Sci. USA 2010, 107, 4182–4187. [Google Scholar] [CrossRef] [PubMed]
- Chianale, F.; Cutrupi, S.; Rainero, E.; Baldanzi, G.; Porporato, P.E.; Traini, S.; Filigheddu, N.; Gnocchi, V.F.; Santoro, M.M.; Parolini, O.; et al. Diacylglycerol kinase-alpha mediates hepatocyte growth factor-induced epithelial cell scatter by regulating rac activation and membrane ruffling. Mol. Biol. Cell 2007, 18, 4859–4871. [Google Scholar] [CrossRef] [PubMed]
- Rainero, E.; Caswell, P.T.; Muller, P.A.; Grindlay, J.; McCaffrey, M.W.; Zhang, Q.; Wakelam, M.J.; Vousden, K.H.; Graziani, A.; Norman, J.C. Diacylglycerol kinase α controls rcp-dependent integrin trafficking to promote invasive migration. J. Cell. Biol. 2012, 196, 277–295. [Google Scholar] [CrossRef] [PubMed]
- Boccaccio, C.; Andò, M.; Tamagnone, L.; Bardelli, A.; Michieli, P.; Battistini, C.; Comoglio, P.M. Induction of epithelial tubules by growth factor hgf depends on the stat pathway. Nature 1998, 391, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Schaper, F.; Siewert, E.; Gómez-Lechon, M.J.; Gatsios, P.; Sachs, M.; Birchmeier, W.; Heinrich, P.C.; Castell, J. Hepatocyte growth factor/scatter factor (hgf/sf) signals via the stat3/aprf transcription factor in human hepatoma cells and hepatocytes. FEBS Lett. 1997, 405, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Stefan, M.; Koch, A.; Mancini, A.; Mohr, A.; Weidner, K.M.; Niemann, H.; Tamura, T. Src homology 2-containing inositol 5-phosphatase 1 binds to the multifunctional docking site of c-met and potentiates hepatocyte growth factor-induced branching tubulogenesis. J. Biol. Chem. 2001, 276, 3017–3023. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Mancini, A.; El Bounkari, O.; Tamura, T. The sh2-domian-containing inositol 5-phosphatase (ship)-2 binds to c-met directly via tyrosine residue 1356 and involves hepatocyte growth factor (HGF)-induced lamellipodium formation, cell scattering and cell spreading. Oncogene 2005, 24, 3436–3447. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.K.; Sasaki, E.; Halter, F.; Pai, R.; Nakamura, T.; Arakawa, T.; Kuroki, T.; Tarnawski, A.S. Hgf triggers activation of the cox-2 gene in rat gastric epithelial cells: Action mediated through the erk2 signaling pathway. FASEB J. 1999, 13, 2186–2194. [Google Scholar] [PubMed]
- Skouteris, G.G.; Schröder, C.H. Cytosolic phospholipase a2 is activated by the hepatocyte growth factor receptor-kinase in madin darby canine kidney cells. J. Cell. Sci. 1997, 110, 1655–1663. [Google Scholar] [PubMed]
- Ma, P.C.; Tretiakova, M.S.; Nallasura, V.; Jagadeeswaran, R.; Husain, A.N.; Salgia, R. Downstream signalling and specific inhibition of c-met/hgf pathway in small cell lung cancer: Implications for tumour invasion. Br. J. Cancer 2007, 97, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Kaposi-Novak, P.; Lee, J.S.; Gòmez-Quiroz, L.; Coulouarn, C.; Factor, V.M.; Thorgeirsson, S.S. Met-regulated expression signature defines a subset of human hepatocellular carcinomas with poor prognosis and aggressive phenotype. J. Clin. Invest. 2006, 116, 1582–1595. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P. Epithelial-mesenchymal transitions in tumour progression. Nat. Rev. Cancer 2002, 2, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Grotegut, S.; von Schweinitz, D.; Christofori, G.; Lehembre, F. Hepatocyte growth factor induces cell scattering through mapk/egr-1-mediated upregulation of snail. EMBO J. 2006, 25, 3534–3545. [Google Scholar] [CrossRef] [PubMed]
- Leroy, P.; Mostov, K.E. Slug is required for cell survival during partial epithelial-mesenchymal transition of hgf-induced tubulogenesis. Mol. Biol. Cell. 2007, 18, 1943–1952. [Google Scholar] [CrossRef] [PubMed]
- Van Leenders, G.J.; Sookhlall, R.; Teubel, W.J.; de Ridder, C.M.; Reneman, S.; Sacchetti, A.; Vissers, K.J.; van Weerden, W.; Jenster, G. Activation of c-met induces a stem-like phenotype in human prostate cancer. PLoS One 2011, 6, e26753. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.T.; King, K.Y.; Brett, J.O.; Cromie, M.J.; Charville, G.W.; Maguire, K.K.; Brunson, C.; Mastey, N.; Liu, L.; Tsai, C.R.; et al. Mtorc1 controls the adaptive transition of quiescent stem cells from g0 to g(alert). Nature 2014, 510, 393–396. [Google Scholar] [PubMed]
- Medico, E.; Mongiovi, A.M.; Huff, J.; Jelinek, M.A.; Follenzi, A.; Gaudino, G.; Parsons, J.T.; Comoglio, P.M. The tyrosine kinase receptors ron and sea control “scattering” and morphogenesis of liver progenitor cells in vitro. Mol. Biol. Cell 1996, 7, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Grisendi, S.; Arpin, M.; Crepaldi, T. Effect of hepatocyte growth factor on assembly of zonula occludens-1 protein at the plasma membrane. J. Cell. Physiol. 1998, 176, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Hay, E.D.; Zuk, A. Transformations between epithelium and mesenchyme: Normal, pathological, and experimentally induced. Am. J. Kidney Dis. 1995, 26, 678–690. [Google Scholar] [CrossRef] [PubMed]
- Potempa, S.; Ridley, A.J. Activation of both map kinase and phosphatidylinositide 3-kinase by ras is required for hepatocyte growth factor/scatter factor-induced adherens junction disassembly. Mol. Biol. Cell. 1998, 9, 2185–2200. [Google Scholar] [CrossRef] [PubMed]
- Khwaja, A.; Lehmann, K.; Marte, B.M.; Downward, J. Phosphoinositide 3-kinase induces scattering and tubulogenesis in epithelial cells through a novel pathway. J. Biol. Chem. 1998, 273, 18793–18801. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J.; Comoglio, P.M.; Hall, A. Regulation of scatter factor/hepatocyte growth factor responses by ras, rac, and rho in mdck cells. Mol. Cell. Biol. 1995, 15, 1110–1122. [Google Scholar] [PubMed]
- Royal, I.; Fournier, T.M.; Park, M. Differential requirement of grb2 and pi3-kinase in HGF/SF-induced cell motility and tubulogenesis. J. Cell. Physiol. 1997, 173, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.X.; Yu, C.F.; Nickel, C.; Thomas, S.; Cantley, L.G. Hepatocyte growth factor induces ERK-dependent paxillin phosphorylation and regulates paxillin-focal adhesion kinase association. J. Biol. Chem. 2002, 277, 10452–10458. [Google Scholar] [CrossRef] [PubMed]
- Graziani, A.; Gramaglia, D.; dalla Zonca, P.; Comoglio, P.M. Hepatocyte growth factor/scatter factor stimulates the ras-guanine nucleotide exchanger. J. Biol. Chem. 1993, 268, 9165–9168. [Google Scholar] [PubMed]
- Hartmann, G.; Weidner, K.M.; Schwarz, H.; Birchmeier, W. The motility signal of scatter factor/hepatocyte growth factor mediated through the receptor tyrosine kinase met requires intracellular action of ras. J. Biol. Chem. 1994, 269, 21936–21939. [Google Scholar] [PubMed]
- Tanimura, S.; Nomura, K.; Ozaki, K.; Tsujimoto, M.; Kondo, T.; Kohno, M. Prolonged nuclear retention of activated extracellular signal-regulated kinase 1/2 is required for hepatocyte growth factor-induced cell motility. J. Biol. Chem. 2002, 277, 28256–28264. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.Y.; Wong, A.S. Activation of p70s6k induces expression of matrix metalloproteinase 9 associated with hepatocyte growth factor-mediated invasion in human ovarian cancer cells. Endocrinology 2006, 147, 2557–2566. [Google Scholar] [CrossRef] [PubMed]
- Monga, S.P.; Mars, W.M.; Pediaditakis, P.; Bell, A.; Mulé, K.; Bowen, W.C.; Wang, X.; Zarnegar, R.; Michalopoulos, G.K. Hepatocyte growth factor induces wnt-independent nuclear translocation of beta-catenin after met-beta-catenin dissociation in hepatocytes. Cancer Res. 2002, 62, 2064–2071. [Google Scholar] [PubMed]
- Papkoff, J.; Aikawa, M. Wnt-1 and hgf regulate gsk3 beta activity and beta-catenin signaling in mammary epithelial cells. Biochem. Biophys. Res. Commun. 1998, 247, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Abella, J.V.; Parachoniak, C.A.; Sangwan, V.; Park, M. Dorsal ruffle microdomains potentiate met receptor tyrosine kinase signaling and down-regulation. J. Biol. Chem. 2010, 285, 24956–24967. [Google Scholar] [CrossRef] [PubMed]
- Wells, C.M.; Ahmed, T.; Masters, J.R.; Jones, G.E. Rho family gtpases are activated during HGF-stimulated prostate cancer-cell scattering. Cell. Motil. Cytoskeleton 2005, 62, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Royal, I.; Lamarche-Vane, N.; Lamorte, L.; Kaibuchi, K.; Park, M. Activation of cdc42, rac, pak, and rho-kinase in response to hepatocyte growth factor differentially regulates epithelial cell colony spreading and dissociation. Mol. Biol. Cell 2000, 11, 1709–1725. [Google Scholar] [CrossRef] [PubMed]
- Nobes, C.D.; Hall, A. Rho, rac, and cdc42 gtpases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia. Cell 1995, 81, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, T.; Sasaki, T.; Takaishi, K.; Kato, M.; Yaku, H.; Araki, K.; Matsuura, Y.; Takai, Y. Rac p21 is involved in insulin-induced membrane ruffling and rho p21 is involved in hepatocyte growth factor- and 12-O-tetradecanoylphorbol-13-acetate (tpa)-induced membrane ruffling in kb cells. Mol. Cell. Biol. 1994, 14, 2447–2456. [Google Scholar] [CrossRef] [PubMed]
- Takaishi, K.; Sasaki, T.; Kato, M.; Yamochi, W.; Kuroda, S.; Nakamura, T.; Takeichi, M.; Takai, Y. Involvement of rho p21 small gtp-binding protein and its regulator in the HGF-induced cell motility. Oncogene 1994, 9, 273–279. [Google Scholar] [PubMed]
- Cozzolino, M.; Stagni, V.; Spinardi, L.; Campioni, N.; Fiorentini, C.; Salvati, E.; Alemà, S.; Salvatore, A.M. P120 catenin is required for growth factor-dependent cell motility and scattering in epithelial cells. Mol. Biol. Cell. 2003, 14, 1964–1977. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.; Nickel, C.H.; Cantley, L.G.; Bruggeman, L.A.; Bennardo, L.N.; Wang, B. Epha kinase activation regulates HGF-induced epithelial branching morphogenesis. J. Cell. Biol. 2003, 162, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Fukata, Y.; Oshiro, N.; Kinoshita, N.; Kawano, Y.; Matsuoka, Y.; Bennett, V.; Matsuura, Y.; Kaibuchi, K. Phosphorylation of adducin by rho-kinase plays a crucial role in cell motility. J. Cell. Biol. 1999, 145, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Nakamura, T.; Kramer, R.H. Hepatocyte growth factor/scatter factor induces tyrosine phosphorylation of focal adhesion kinase (p125fak) and promotes migration and invasion by oral squamous cell carcinoma cells. J. Biol. Chem. 1994, 269, 31807–31813. [Google Scholar] [PubMed]
- Jiang, W.; Hiscox, S.; Nakamura, T.; Hallett, M.; Puntis, M.; Mansel, R. Hepatocyte growth factor induces tyrosine phosphorylation of focal adhesion kinase (fak) and paxillin and enhances cell-matrix interactions. Oncol. Rep. 1996, 3, 819–823. [Google Scholar] [PubMed]
- Trusolino, L.; Cavassa, S.; Angelini, P.; Andó, M.; Bertotti, A.; Comoglio, P.M.; Boccaccio, C. Hgf/scatter factor selectively promotes cell invasion by increasing integrin avidity. FASEB J. 2000, 14, 1629–1640. [Google Scholar] [CrossRef] [PubMed]
- Trusolino, L.; Serini, G.; Cecchini, G.; Besati, C.; Ambesi-Impiombato, F.S.; Marchisio, P.C.; De Filippi, R. Growth factor-dependent activation of alphavbeta3 integrin in normal epithelial cells: Implications for tumor invasion. J. Cell. Biol. 1998, 142, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Nusrat, A.; Parkos, C.A.; Bacarra, A.E.; Godowski, P.J.; Delp-Archer, C.; Rosen, E.M.; Madara, J.L. Hepatocyte growth factor/scatter factor effects on epithelia. Regulation of intercellular junctions in transformed and nontransformed cell lines, basolateral polarization of c-met receptor in transformed and natural intestinal epithelia, and induction of rapid wound repair in a transformed model epithelium. J. Clin. Invest. 1994, 93, 2056–2065. [Google Scholar] [CrossRef] [PubMed]
- Crepaldi, T.; Pollack, A.L.; Prat, M.; Zborek, A.; Mostov, K.; Comoglio, P.M. Targeting of the sf/hgf receptor to the basolateral domain of polarized epithelial cells. J. Cell. Biol. 1994, 125, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Korhan, P.; Erdal, E.; Kandemiş, E.; Cokaklı, M.; Nart, D.; Yılmaz, F.; Can, A.; Atabey, N. Reciprocal activating crosstalk between c-met and caveolin 1 promotes invasive phenotype in hepatocellular carcinoma. PLoS One 2014, 9, e105278. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; O’Brien, L.E.; Wang, F.; Bourne, H.; Mostov, K.E.; Zegers, M.M. Hepatocyte growth factor switches orientation of polarity and mode of movement during morphogenesis of multicellular epithelial structures. Mol. Biol. Cell. 2003, 14, 748–763. [Google Scholar] [CrossRef] [PubMed]
- Pollack, A.L.; Runyan, R.B.; Mostov, K.E. Morphogenetic mechanisms of epithelial tubulogenesis: Mdck cell polarity is transiently rearranged without loss of cell–cell contact during scatter factor/hepatocyte growth factor-induced tubulogenesis. Dev. Biol. 1998, 204, 64–79. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, L.E.; Tang, K.; Kats, E.S.; Schutz-Geschwender, A.; Lipschutz, J.H.; Mostov, K.E. Erk and mmps sequentially regulate distinct stages of epithelial tubule development. Dev. Cell 2004, 7, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Hamasuna, R.; Kataoka, H.; Moriyama, T.; Itoh, H.; Seiki, M.; Koono, M. Regulation of matrix metalloproteinase-2 (mmp-2) by hepatocyte growth factor/scatter factor (hgf/sf) in human glioma cells: HGF/SF enhances mmp-2 expression and activation accompanying up-regulation of membrane type-1 mmp. Int. J. Cancer 1999, 82, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Keiser, J.A. Hepatocyte growth factor enhances mmp activity in human endothelial cells. Biochem. Biophys. Res. Commun. 2000, 272, 900–905. [Google Scholar] [CrossRef] [PubMed]
- McCawley, L.J.; Li, S.; Wattenberg, E.V.; Hudson, L.G. Sustained activation of the mitogen-activated protein kinase pathway. A mechanism underlying receptor tyrosine kinase specificity for matrix metalloproteinase-9 induction and cell migration. J. Biol. Chem. 1999, 274, 4347–4353. [Google Scholar] [CrossRef] [PubMed]
- Tushir, J.S.; D’Souza-Schorey, C. Arf6-dependent activation of erk and rac1 modulates epithelial tubule development. EMBO J. 2007, 26, 1806–1819. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.H.; Jeffers, M.; Bellacosa, A.; Mitsuuchi, Y.; Vande Woude, G.F.; Testa, J.R. Anti-apoptotic signaling by hepatocyte growth factor/met via the phosphatidylinositol 3-kinase/akt and mitogen-activated protein kinase pathways. Proc. Natl. Acad. Sci. USA 2001, 98, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Chen, S.; You, Z.; Yang, F.; Carey, T.E.; Saims, D.; Wang, C.Y. Hepatocyte growth factor inhibits anoikis in head and neck squamous cell carcinoma cells by activation of ERK and AKT signaling independent of nfkappa b. J. Biol. Chem. 2002, 277, 25203–25208. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Hepatocyte growth factor promotes renal epithelial cell survival by dual mechanisms. Am J. Physiol. 1999, 277, F624–F633. [Google Scholar] [PubMed]
- Schulze-Bergkamen, H.; Brenner, D.; Krueger, A.; Suess, D.; Fas, S.C.; Frey, C.R.; Dax, A.; Zink, D.; Büchler, P.; Müller, M.; et al. Hepatocyte growth factor induces mcl-1 in primary human hepatocytes and inhibits cd95-mediated apoptosis via AKT. Hepatology 2004, 39, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Moumen, A.; Patané, S.; Porras, A.; Dono, R.; Maina, F. Met acts on mdm2 via mtor to signal cell survival during development. Development 2007, 134, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- Fassetta, M.; D’Alessandro, L.; Coltella, N.; di Renzo, M.F.; Rasola, A. Hepatocyte growth factor installs a survival platform for colorectal cancer cell invasive growth and overcomes p38 mapk-mediated apoptosis. Cell Signal 2006, 18, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, I.; de Juan, C.; Nakamura, T.; Benito, M. Growth stimulation of rat fetal hepatocytes in response to hepatocyte growth factor: Modulation of c-myc and c-fos expression. Biochem. Biophys. Res. Commun. 1992, 189, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Lechón, M.J.; Guillén, I.; Ponsoda, X.; Fabra, R.; Trullenque, R.; Nakamura, T.; Castell, J.V. Cell cycle progression proteins (cyclins), oncogene expression, and signal transduction during the proliferative response of human hepatocytes to hepatocyte growth factor. Hepatology 1996, 23, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Shibamoto, S.; Nagamine, K.; Shigemori, I.; Omura, S.; Kitamura, N.; Ito, F. Signaling pathways leading to transcription and translation cooperatively regulate the transient increase in expression of c-fos protein. J. Biol. Chem. 2001, 276, 26077–26083. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Meng, Q.; Laterra, J.J.; Rosen, E.M. Ras effector pathways modulate scatter factor-stimulated nf-kappab signaling and protection against DNA damage. Oncogene 2007, 26, 4774–4796. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Morotti, A.; Ponzetto, C. Activation of nf-kappab is essential for hepatocyte growth factor-mediated proliferation and tubulogenesis. Mol. Cell. Biol. 2002, 22, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Tsukada, Y.; Hara, E.; Kitamura, N.; Tanaka, T. Hepatocyte growth factor induces redistribution of p21(cip1) and p27(kip1) through ERK-dependent p16(ink4a) up-regulation, leading to cell cycle arrest at g1 in hepg2 hepatoma cells. J. Biol. Chem. 2005, 280, 31548–31556. [Google Scholar] [CrossRef] [PubMed]
- Shima, N.; Stolz, D.B.; Miyazaki, M.; Gohda, E.; Higashio, K.; Michalopoulos, G.K. Possible involvement of p21/waf1 in the growth inhibition of hepg2 cells induced by hepatocyte growth factor. J. Cell. Physiol. 1998, 177, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, V.; Paliouras, G.N.; Abella, J.V.; Dubé, N.; Monast, A.; Tremblay, M.L.; Park, M. Regulation of the met receptor-tyrosine kinase by the protein-tyrosine phosphatase 1b and t-cell phosphatase. J. Biol. Chem. 2008, 283, 34374–34383. [Google Scholar] [CrossRef] [PubMed]
- Villa-Moruzzi, E.; Puntoni, F.; Bardelli, A.; Vigna, E.; de Rosa, S.; Comoglio, P.M. Protein tyrosine phosphatase ptp-s binds to the juxtamembrane region of the hepatocyte growth factor receptor met. Biochem. J. 1998, 336, 235–239. [Google Scholar] [PubMed]
- Xu, Y.; Xia, W.; Baker, D.; Zhou, J.; Cha, H.C.; Voorhees, J.J.; Fisher, G.J. Receptor-type protein tyrosine phosphatase beta (rptp-beta) directly dephosphorylates and regulates hepatocyte growth factor receptor (hgfr/met) function. J. Biol. Chem. 2011, 286, 15980–15988. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhou, J.; Carey, T.E.; McHugh, J.B.; Voorhees, J.J.; Fisher, G.J. Receptor-type protein tyrosine phosphatase β regulates met phosphorylation and function in head and neck squamous cell carcinoma. Neoplasia 2012, 14, 1015–1022. [Google Scholar] [PubMed]
- Kulas, D.T.; Goldstein, B.J.; Mooney, R.A. The transmembrane protein-tyrosine phosphatase lar modulates signaling by multiple receptor tyrosine kinases. J. Biol. Chem. 1996, 271, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Machide, M.; Hashigasako, A.; Matsumoto, K.; Nakamura, T. Contact inhibition of hepatocyte growth regulated by functional association of the c-met/hepatocyte growth factor receptor and lar protein-tyrosine phosphatase. J. Biol. Chem. 2006, 281, 8765–8772. [Google Scholar] [CrossRef] [PubMed]
- Palka, H.L.; Park, M.; Tonks, N.K. Hepatocyte growth factor receptor tyrosine kinase met is a substrate of the receptor protein-tyrosine phosphatase dep-1. J. Biol. Chem. 2003, 278, 5728–5735. [Google Scholar] [CrossRef] [PubMed]
- Hashigasako, A.; Machide, M.; Nakamura, T.; Matsumoto, K. Bi-directional regulation of ser-985 phosphorylation of c-met via protein kinase c and protein phosphatase 2a involves c-met activation and cellular responsiveness to hepatocyte growth factor. J. Biol. Chem. 2004, 279, 26445–26452. [Google Scholar] [CrossRef] [PubMed]
- Gandino, L.; Longati, P.; Medico, E.; Prat, M.; Comoglio, P.M. Phosphorylation of serine 985 negatively regulates the hepatocyte growth factor receptor kinase. J. Biol. Chem. 1994, 269, 1815–1820. [Google Scholar] [PubMed]
- Lee, J.H.; Gao, C.F.; Lee, C.C.; Kim, M.D.; vande Woude, G.F. An alternatively spliced form of met receptor is tumorigenic. Exp. Mol. Med. 2006, 38, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Joffre, C.; Barrow, R.; Ménard, L.; Calleja, V.; Hart, I.R.; Kermorgant, S. A direct role for met endocytosis in tumorigenesis. Nat. Cell. Biol. 2011, 13, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Guzman, M.; Larsen, E.; Vuori, K. The proto-oncogene c-cbl is a positive regulator of met-induced map kinase activation: A role for the adaptor protein crk. Oncogene 2000, 19, 4058–4065. [Google Scholar] [CrossRef] [PubMed]
- Petrelli, A.; Gilestro, G.F.; Lanzardo, S.; Comoglio, P.M.; Migone, N.; Giordano, S. The endophilin-cin85-cbl complex mediates ligand-dependent downregulation of c-met. Nature 2002, 416, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Peschard, P.; Ishiyama, N.; Lin, T.; Lipkowitz, S.; Park, M. A conserved dpyr motif in the juxtamembrane domain of the met receptor family forms an atypical c-cbl/cbl-b tyrosine kinase binding domain binding site required for suppression of oncogenic activation. J. Biol. Chem. 2004, 279, 29565–29571. [Google Scholar] [CrossRef] [PubMed]
- Raiborg, C.; Bache, K.G.; Gillooly, D.J.; Madshus, I.H.; Stang, E.; Stenmark, H. Hrs sorts ubiquitinated proteins into clathrin-coated microdomains of early endosomes. Nat. Cell Biol. 2002, 4, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Hammond, D.E.; Carter, S.; McCullough, J.; Urbé, S.; vande Woude, G.; Clague, M.J. Endosomal dynamics of met determine signaling output. Mol. Biol. Cell. 2003, 14, 1346–1354. [Google Scholar] [CrossRef] [PubMed]
- Hammond, D.E.; Urbé, S.; vande Woude, G.F.; Clague, M.J. Down-regulation of met, the receptor for hepatocyte growth factor. Oncogene 2001, 20, 2761–2770. [Google Scholar] [CrossRef] [PubMed]
- Franco, M.; Muratori, C.; Corso, S.; Tenaglia, E.; Bertotti, A.; Capparuccia, L.; Trusolino, L.; Comoglio, P.M.; Tamagnone, L. The tetraspanin cd151 is required for met-dependent signaling and tumor cell growth. J. Biol. Chem. 2010, 285, 38756–38764. [Google Scholar] [CrossRef] [PubMed]
- Trusolino, L.; Bertotti, A.; Comoglio, P.M. A signaling adapter function for alpha6beta4 integrin in the control of HGF-dependent invasive growth. Cell 2001, 107, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Bertotti, A.; Comoglio, P.M.; Trusolino, L. Beta4 integrin is a transforming molecule that unleashes met tyrosine kinase tumorigenesis. Cancer Res. 2005, 65, 10674–10679. [Google Scholar] [CrossRef] [PubMed]
- Bertotti, A.; Comoglio, P.M.; Trusolino, L. Beta4 integrin activates a shp2-src signaling pathway that sustains hgf-induced anchorage-independent growth. J. Cell Biol. 2006, 175, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.K.; Sawada, K.; Tiwari, P.; Mui, K.; Gwin, K.; Lengyel, E. Ligand-independent activation of c-met by fibronectin and α(5)β(1)-integrin regulates ovarian cancer invasion and metastasis. Oncogene 2011, 30, 1566–1576. [Google Scholar] [CrossRef] [PubMed]
- Antipenko, A.; Himanen, J.P.; van Leyen, K.; Nardi-Dei, V.; Lesniak, J.; Barton, W.A.; Rajashankar, K.R.; Lu, M.; Hoemme, C.; Püschel, A.W.; et al. Structure of the semaphorin-3a receptor binding module. Neuron 2003, 39, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Giordano, S.; Corso, S.; Conrotto, P.; Artigiani, S.; Gilestro, G.; Barberis, D.; Tamagnone, L.; Comoglio, P.M. The semaphorin 4d receptor controls invasive growth by coupling with met. Nat. Cell. Biol. 2002, 4, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Conrotto, P.; Corso, S.; Gamberini, S.; Comoglio, P.M.; Giordano, S. Interplay between scatter factor receptors and b plexins controls invasive growth. Oncogene 2004, 23, 5131–5137. [Google Scholar] [CrossRef] [PubMed]
- Conrotto, P.; Valdembri, D.; Corso, S.; Serini, G.; Tamagnone, L.; Comoglio, P.M.; Bussolino, F.; Giordano, S. Sema4d induces angiogenesis through met recruitment by plexin b1. Blood 2005, 105, 4321–4329. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, A.; Götze, T.; Korc, M. Hepatocyte growth factor-mediated cell invasion in pancreatic cancer cells is dependent on neuropilin-1. Cancer Res. 2007, 67, 10309–10316. [Google Scholar] [CrossRef] [PubMed]
- Sulpice, E.; Plouët, J.; Bergé, M.; Allanic, D.; Tobelem, G.; Merkulova-Rainon, T. Neuropilin-1 and neuropilin-2 act as coreceptors, potentiating proangiogenic activity. Blood 2008, 111, 2036–2045. [Google Scholar] [CrossRef] [PubMed]
- Orian-Rousseau, V.; Chen, L.; Sleeman, J.P.; Herrlich, P.; Ponta, H. Cd44 is required for two consecutive steps in HGF/c-met signaling. Genes Dev. 2002, 16, 3074–3086. [Google Scholar] [CrossRef] [PubMed]
- Crepaldi, T.; Gautreau, A.; Comoglio, P.M.; Louvard, D.; Arpin, M. Ezrin is an effector of hepatocyte growth factor-mediated migration and morphogenesis in epithelial cells. J. Cell. Biol. 1997, 138, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Orian-Rousseau, V.; Morrison, H.; Matzke, A.; Kastilan, T.; Pace, G.; Herrlich, P.; Ponta, H. Hepatocyte growth factor-induced ras activation requires erm proteins linked to both cd44v6 and f-actin. Mol. Biol. Cell 2007, 18, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Hasenauer, S.; Malinger, D.; Koschut, D.; Pace, G.; Matzke, A.; von Au, A.; Orian-Rousseau, V. Internalization of met requires the co-receptor cd44v6 and its link to erm proteins. PLoS One 2013, 8, e62357. [Google Scholar] [CrossRef] [PubMed]
- Todaro, M.; Gaggianesi, M.; Catalano, V.; Benfante, A.; Iovino, F.; Biffoni, M.; Apuzzo, T.; Sperduti, I.; Volpe, S.; Cocorullo, G.; et al. Cd44v6 is a marker of constitutive and reprogrammed cancer stem cells driving colon cancer metastasis. Cell Stem Cell 2014, 14, 342–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fafalios, A.; Ma, J.; Tan, X.; Stoops, J.; Luo, J.; Defrances, M.C.; Zarnegar, R. A hepatocyte growth factor receptor (met)-insulin receptor hybrid governs hepatic glucose metabolism. Nat. Med. 2011, 17, 1577–1584. [Google Scholar] [CrossRef] [PubMed]
- Bertola, A.; Bonnafous, S.; Cormont, M.; Anty, R.; Tanti, J.F.; Tran, A.; Le Marchand-Brustel, Y.; Gual, P. Hepatocyte growth factor induces glucose uptake in 3t3-l1 adipocytes through a gab1/phosphatidylinositol 3-kinase/glut4 pathway. J. Biol. Chem. 2007, 282, 10325–10332. [Google Scholar] [CrossRef] [PubMed]
- Perdomo, G.; Martinez-Brocca, M.A.; Bhatt, B.A.; Brown, N.F.; O’Doherty, R.M.; Garcia-Ocaña, A. Hepatocyte growth factor is a novel stimulator of glucose uptake and metabolism in skeletal muscle cells. J. Biol. Chem. 2008, 283, 13700–13706. [Google Scholar] [CrossRef] [PubMed]
- Follenzi, A.; Bakovic, S.; Gual, P.; Stella, M.C.; Longati, P.; Comoglio, P.M. Cross-talk between the proto-oncogenes met and ron. Oncogene 2000, 19, 3041–3049. [Google Scholar] [CrossRef] [PubMed]
- Benvenuti, S.; Lazzari, L.; Arnesano, A.; Li Chiavi, G.; Gentile, A.; Comoglio, P.M. Ron kinase transphosphorylation sustains met oncogene addiction. Cancer Res. 2011, 71, 1945–1955. [Google Scholar] [CrossRef] [PubMed]
- Jo, M.; Stolz, D.B.; Esplen, J.E.; Dorko, K.; Michalopoulos, G.K.; Strom, S.C. Cross-talk between epidermal growth factor receptor and c-met signal pathways in transformed cells. J. Biol. Chem. 2000, 275, 8806–8811. [Google Scholar] [CrossRef] [PubMed]
- Tanizaki, J.; Okamoto, I.; Sakai, K.; Nakagawa, K. Differential roles of trans-phosphorylated egfr, her2, her3, and ret as heterodimerisation partners of met in lung cancer with met amplification. Br. J. Cancer 2011, 105, 807–813. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Baldanzi, G.; Graziani, A. Physiological Signaling and Structure of the HGF Receptor MET. Biomedicines 2015, 3, 1-31. https://doi.org/10.3390/biomedicines3010001
AMA Style
Baldanzi G, Graziani A. Physiological Signaling and Structure of the HGF Receptor MET. Biomedicines. 2015; 3(1):1-31. https://doi.org/10.3390/biomedicines3010001
Chicago/Turabian StyleBaldanzi, Gianluca, and Andrea Graziani. 2015. "Physiological Signaling and Structure of the HGF Receptor MET" Biomedicines 3, no. 1: 1-31. https://doi.org/10.3390/biomedicines3010001