Oncolytic Adenoviruses in Gastrointestinal Cancers

1
Department of Medicine, Division of Hematology Oncology, Mayo Clinic Arizona, 13400 E Shea Blvd, Scottsdale, AZ 85205, USA
2
Department of Molecular Medicine, Mayo Clinic, Rochester, MN 55905, USA
3
Center for Individualized Medicine, Mayo Clinic, 200 1st St SW, Rochester, MN 55905, USA
4
Mayo Clinic Cancer Center, 5881 E Mayo Blvd, Phoenix, AZ 85054, USA
*
Author to whom correspondence should be addressed.
Biomedicines 2018, 6(1), 33; https://doi.org/10.3390/biomedicines6010033
Submission received: 31 January 2018
/
Revised: 5 March 2018
/
Accepted: 9 March 2018
/
Published: 11 March 2018
(This article belongs to the Special Issue Adenoviruses: From Virus to Medicine)
Abstract
:Gastrointestinal malignancies are challenging cancers with considerable economic and societal impacts on health care systems worldwide. While advances in surgical approaches have provided benefits to a proportion of patients, only modest improvements have been attained in the treatment of patients with advanced disease, resulting in limited improvement in survival rates in these patients. Oncolytic adenoviruses are being developed to address gastrointestinal malignancies. Each platform has evolved to maximize tumor-cell killing potency while minimizing toxicities. Tumor-specific bioengineered adenoviruses using chimeric promoters, prodrug convertase enzymes, lethal genes, tumor suppressor genes, and pseudo-typed capsids can provide the innovations for eventual success of oncolytic virotherapy. This article will review the developments in adenoviral platforms in the context of specific gastrointestinal cancers. From the bench to the implementation of clinical trials, this review aims to highlight advances in the field from its early days to the current state of affairs as it pertains to the application of adenoviral oncolytic therapy to gastrointestinal cancers.
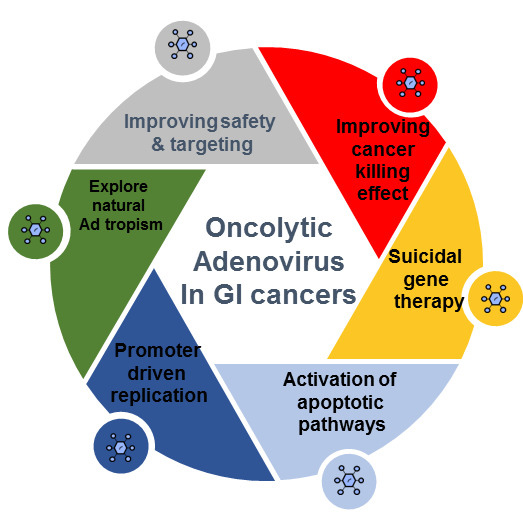
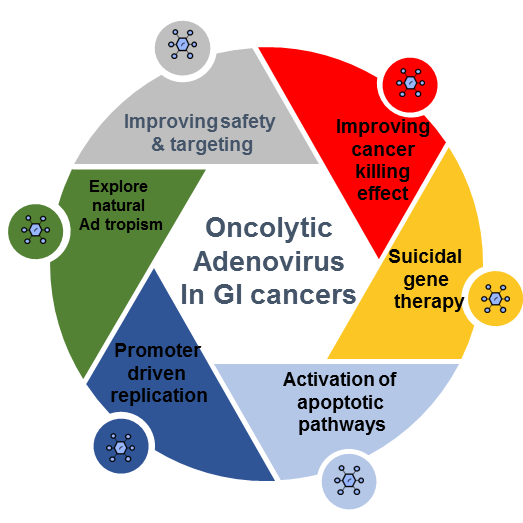
1. Introduction
Gastrointestinal cancers are a significant public health concern worldwide. They have a considerable impact on health economics that permeates multiple aspects of healthcare ranging from screening and prevention to hospice care [1]. In the United States, colorectal cancer is the fourth leading cause of cancer mortality. Pancreatic cancer and hepatocellular cancer also feature in the ten most lethal cancers in the United States [2]. Given the tremendous unmet therapeutic need for these cancers, novel approaches are in imminent need. Oncovirotherapy using adenoviruses (Ads) represents a very attractive anti-cancer therapeutic platform.
This review will focus on the evolution of adenoviral bioengineering and manipulation in esophageal, gastric, pancreatic, liver, biliary, and colorectal cancers. Progress in the field from pre-clinical studies to initial clinical trials will be covered with emphasis on barriers in the area and lessons learned to date. It is important to highlight that gastrointestinal malignancies were grouped as a way to facilitate the understanding of the evolution of the viral vectors per gastrointestinal organ. The cell biology of a vast heterogeneity of tumors in each organ is beyond the scope of this review.
2. Oncolytic Adenoviral Platform Development
Ads are non-enveloped DNA viruses with an icosahedral capsid encompassing a linear duplex genome of ~36 kb. Ads have been found in the majority of vertebrates [3]. Human Ads are ubiquitous in the environment and have been classified into 57 serotypes (Ad1-Ad57) based on cross-susceptibility to neutralizing antibodies and seven subgroups (A–G). Within each subgroup, there are similarities in virulence and tissue tropism [4]. In immunocompetent individuals, human Ad infections are mild, consisting of self-limited respiratory cold-like infections.
Human adenoviruses represent an interesting oncolytic virotherapy platform given their (1) high transduction efficiency in transformed cells (~10,000 viral particles per infected cell), (2) lack of integration into the host genome resulting in a lowered risk of insertional mutagenesis, (3) low seroprevalence with regards to specific serotypes, (4) high fidelity DNA polymerase, which confers relative stability, and (5) ability to attain tumor specificity through substitution of the viral promoter with cancer tissue selective promoters or mutations that enable virus replication to occur preferentially in transformed cells.
Oncolytic virotherapy (OV) is a promising therapeutic platform with applications across a broad array of malignancies. The field of OV has its roots in 1890 when the first reports of spontaneous tumor regression were noted, following an episode of viral illness in a patient with leukemia. Similar observations followed in patients exposed to viral infections such as varicella, hepatitis, and measles in patients with Hodgkin’s disease, resulting in spontaneous regressions. However, these effects were mostly transient in nature. After a period of relative dormancy, a revival in the OV arena over the last 15 years has been incited by the increased understanding of the biology of viruses and advances in synthetic biology and recombinant nucleic acid technology allowing for the synthesis of more potent, selective and safe therapeutic viral vectors. In 2006, the adenovirus H101 was approved for clinical use in China for patients with advanced nasopharyngeal cancer [5]. Thus far, a series of clinical trials using H101 combined with standard chemotherapy demonstrated a better overall response in head and neck cancers [6].
Oncolytic adenoviral (OA) strategies have been evaluated in gastrointestinal (GI) malignancies. Although the initial research was conducted with Ad5, natural tropism and seroprevalence are becoming critical factors for vector success. A number of Ad subgroups can provide natural GI tropism such as the subgroup A (Ad12), and subgroup F (Ad40, Ad 41) [3]. The Ad12 E1B protein had similarities with A-gliadin and was studied as a factor in the development of gluten intolerance [7]. Ad40 and Ad41 are responsible for acute diarrhea and gastroenteritis, mostly in children [8]. These Ads use coxsackie-adenovirus receptors, CAR, and Ad 41 may promote disruption of the enterochromaffin cells and enteric glial cells leading to serotonin release [9]. Other types of natural tropism [3] are described in Table 1.
Additionally, tumor specificity and tropism can be enhanced through promoter driven replication. In GI malignancies, a number of adenoviral tumor-specific promoters have been analyzed to date (Table 2). Each of them will be discussed in the tumor-type-specific sections that follow.
Ads provide the possibility for hexon swapping and fiber pseudotyping among the subgroups, which is attractive toward maximizing targeting and delaying immune neutralization. Given this vast array of Ad serotypes, chimeric capsid investigations are still evolving, especially when considering which combinations would best enhance tissue tropism and provide on-target attachment. A few studies have shown that some chimeras combined with specific molecules could promote a reduction of tropism, which is of significant interest, as it may enhance virus bioavailability and contribute to diminishing liver and reticuloendothelial system sequestration [19].
3. Preclinical Perspectives in Gastrointestinal Cancers
3.1. Esophageal Cancer
Esophageal cancer has an incidence of 17,290 cases a year in the United States (USA), and patients who are inflicted typically experience a reduced quality of life [20]. Although there have been a number of advances in treatment in recent years, the overall mortality remains considerably high. Oncolytic Ads have undergone pre-clinical evaluation in esophageal cancer models (Table 3). One of the first such Ad vectors was designed to induce cell cycle arrest, which enhanced the oncolytic effect, with reduced nuclear factor kappa-B (NF-κB) and maximized apoptosis, primarily in p53 mutant cells [21].
Meanwhile, another study evaluated resistant tumor cells which exhibit properties of cancer stem cells. Radiotherapy resistance is a significant concern in esophageal cancer. The study analyzed an Ad carrying the tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) that preferentially induced apoptosis in radio-resistant cancer cells [22]. Another group investigated the combined effects of chemotherapy and oncovirotherapy using Ad. Four drugs were analyzed in combination with a modified Ad5, which encompassed the deletion of the 55 kDa-encoding E1B region as and a part of the E3 region. The four medications tested were 5-fluorouracil, etoposide, mitomycin C, and cisplatin. 5-Fluorouracil intensified the cell cycle at the S phase and promoted G2/M phase entry. Conversely, cisplatin produced G1 phase arrest. Interestingly, cisplatin action was antagonistic to the Ad vector, as it inhibited the Ad-mediated cell-cycle, not favoring a viral cytopathic effect. As such, cisplatin could lead to cross-resistance with Ad vectors [23].
Furthermore, another group developed the vector Telomelysin (OBP-301), which is an Ad5, with E1A genes under the control of a human telomerase reverse transcriptase (hTERT) promoter. The study achieved selective viral replication in tumor cells that expressed telomerase activity. The pre-clinical efficacy of this vector was found to be encouraging, and its evaluation has now progressed to clinical trials [24]. Along these lines, another group tested the adenoviral vector H101 in esophageal carcinoma [25].
A chimeric vector, using Ad H101 as a packaging template and the Newcastle disease virus (NDV) hemagglutinin-neuraminidase (HN), was tested in animal models. An increase in reactive oxygen species resulted in cytotoxicity and complete curative responses with prolonged survival with the use of intra-tumoral vector inoculation [26]. Despite being attractive tools conceptually, chimeric vectors have demonstrated diminished transduction rates when compared to wild types, resulting in a somewhat tempered enthusiasm for further evaluation.
3.2. Gastric Cancer
Gastric cancer has an incidence of 26,240 cases per year in the USA [20]. Surgery can be curative, but diagnosis in the setting of patients with advanced stage limits the number of patients eligible for surgical therapies. Novel therapeutic approaches are needed for patients with advanced disease.
OV therapy represents a promising platform for use in gastric cancer (Table 4). Enhancements in safety can be achieved by manipulation using Ad tumor-specific promoters, which enable the development of conditionally replicating adenoviruses (CRAds). Midkine (MK) and cyclooxygenase-2 (Cox-2M and Cox-2L) demonstrate high transcriptional activity in cell-lines from gastric cancer and represent promising elements for incorporation into CRAds. A study using an Ad5/3 vector with the Cox-2 promoter (Cox-2CRAd) demonstrated encouraging anti-tumor activity in gastric cancer models [11]. In another study, a CRAd using the CEA promoter was evaluated in gastric cancer cell lines with high CEA and had potent efficacy [17].
Along these lines, a group investigated the Ad Telomelysin (OBP-301), to target quiescent stem-like cells. These cells are in a dormant phase, which makes them resistant to chemotherapies. A strategy to overcome their resistance is to mobilize them into the cell cycle to elicit a treatment response. It has been shown that cell cycle mobilization can be induced by adenoviral infection. This study was important from this perspective, given that mobilization of cells from a quiescent state into the cell-cycle had only been previously achieved in leukemia [14].
Another study established a viral gene therapy approach by creating an Ad with endostatin, which is known to be an inhibitor of angiogenesis [27]. This vector exerted a compounded effect of the wild-type OV and endostatin toward promoting oncolysis. Correspondingly, another study investigated an Ad OV platform under a survivin promoter along with the Hsp-70 chaperone gene. The vector replicated selectively only in survivin-positive gastric cancer cells in mouse models [15].
3.3. Liver and Biliary Cancers
Liver and intrahepatic bile duct cancers have an incidence of approximately 42,220 cases per year in the USA [20]. Hepatocellular carcinoma (HCC) is the most common primary liver cancer [28]. Current therapies for advanced HCC have thus far not resulted in a high proportion of cures. The application of OV is an attractive approach in this setting, and a number of pre-clinical evaluations have been undertaken in this context (Table 5).
Ad CV890 was the first Ad5-specific developed for selectively targeting hepatocellular carcinoma (HCC) by using alpha-fetoprotein (AFP) as a tumor-specific promoter. This study evaluated the ability of the vector to eliminate distant tumor recurrence when used in combination with doxorubicin in animal models. These promising data support further evaluation of this OV in clinical trials [29].
A pioneer Ad in OV, ONYX-015 [30], has been evaluated in HCC. Its safety profile and oncolytic efficacy were encouraging in animal models where the vector was loaded with a murine endostatin gene [31].
Another strategy for enhancing OV and gene therapy delivery is the dual promoter approach. It can enhance tumor specificity as well as vector delivery of gene therapy in HCC. A CRAd system placed the Ad E1B gene under a TP53 gene expressing cassette coupled with hTERT promoter and a hypoxia response element (HRE) promoter. The goal was to foster p53 protein production in a hypoxic microenvironment in telomerase-positive HCC cells. An enhanced oncolytic effect was reported with this dual promoter-enabled construct [32].
Other studies have successfully implemented anti-tumor gene therapy using the Ad OV platform to deliver a tumor suppressor gene, TSLC1, which is lost in many human cancers, including liver cancers, and conserved in normal cells [33].
A parallel strategy to enhance tumor-killing effect was achieved by adding lethal mitochondrial genes to the Ad OV platform to be selectively expressed in tumor cells by the vector. One such vector that has been developed encodes the second mitochondria-derived activator of caspases (SMAC) protein. It demonstrated superior oncolytic potency but had also exhibited cytotoxicity toward normal cells, emphasizing the importance of achieving tumor selectivity when deploying more potent vectors [34].
Besides AFP, transthyretin [35] has also been identified as an Ad promoter for enhancing HCC vector specificity. Recently, GOLPH2 (GP73), a Golgi protein, was also instituted as an HCC-specific adenovirus promoter [13]. A number of hybrid promoters are also being evaluated. An example of this is the HRE-AFP promoter which has been utilized in an Ad platform to deliver melittin, a bee venom toxic peptide that can induce HCC apoptosis. Inhibitory effects of the vector in HCC were reported as a result of a triple killing mechanism targeting AFP-positive cells in a hypoxic tumor microenvironment, and cells with p53 deficiency [16].
Additional layers of safety through the use of microRNAs, such as let-7 to control vector replication and diminish hepatotoxicity, have also been utilized [36]. Recently, an Ad expressing long non-coding RNA that can competitively bind oncogenic miRNAs has achieved reasonable anti-tumor efficacy [37].
Anti-tumor effect and oncolysis have been improved by a number of strategies. SOCS3, suppressor of cytokine signaling 3, can downregulate Cyclin D1 and anti-apoptotic proteins [38]. Similarly, SOCS1 negatively regulates signal transduction and activation of transcription 3 (STAT3) and can be employed to inhibit STAT3 phosphorylation and ultimately downregulate survivin and c-myc [39].
Another strategy to augment OV potency within the tumoral hypoxic microenvironment relies on oxygen-dependent degradation domain-regulated vectors [40]. Similarly, an Ad OV platform using manganese superoxide dismutase has been shown to suppress HCC growth effectively in patient-derived xenografted mice [41].
Recently, the concern regarding the concomitant use of Ad and cisplatin has been overcome by providing XAF1 in a vector platform. XAF1 counters the effects of the inhibitor of apoptosis protein (IAP). This vector enabled enhanced tumor cell apoptosis through activation of the caspase-9/PARP pathway, which ultimately resulted in reduced cisplatin doses [42].
3.4. Pancreatic Cancer
Pancreatic cancer has an incidence of 55,440 cases per year and more than 21,000 deaths each year in the USA [20]. Despite recent advances in systemic therapy with the advent of gemcitabine/nab-paclitaxel and FOLFIRINOX, survival rates have not meaningfully changed, and new approaches for treatment are in imminent need.
Efforts are underway to develop specific OVs for pancreatic cancer (Table 6). An Ad with E1B-55 kDa deletion was one of the first adenoviral vectors to show a response to OV comparable to the other tumors. This vector can selectively replicate in TP53 deficient cells. Additionally, this Ad vector was manipulated to express uracil phosphoribosyl transferase (UPRT), which can enhance therapeutic effects, given that it overcomes 5-fluorouracil resistance [43].
Another gene-therapy-based OV utilized Ad5 carrying suicide genes such as the cytosine deaminase (CD) as well as herpes virus thymidine kinase (HSV TK). This vector was able to improve radiotherapy effect without excessive pancreatic toxicity [44]. Conversely, an Ad carrying the TK gene alone resulted in enhanced survival when used in combination with ganciclovir using an intra-ductal delivery approach in animal models [45].
One approach toward achieving tumor selectivity in pancreatic cancer involves an Ad targeting matrix metalloproteases (MMPs) at the surface entry level and has been shown to reduce metastases with no significant toxicities in vivo for pancreatic cancer models [46].
Likewise, another approach along these lines has employed the use of a SYE ligand, with targeting sequence SYENFSA, acting as a pancreatic cancer-targeting ligand capable of boosting promoter specificity of an Ad. Studies conducted with this ligand have achieved robust transduction efficiency, resulting in potent oncolysis in pancreatic tumors [47].
3.5. Colorectal Cancer
Colon cancer has an incidence of 97,220 cases per year in the USA, and rectal cancer has an incidence of 43,030 [20]. Despite prevention efforts leading to some improvement in survival due to an increase in early detection, it remains a significant cause of cancer morbidity and mortality. OV therapy represents an exciting avenue for novel therapies in colorectal cancers.
Besides achieving tumor specificity, OV platforms for colon cancer have also focused on expressing pro-drug activating enzymes in cancer cells (Table 7). One early study used the enzyme nitroreductase (NTR) in an E1B-55-kDa deleted Ad. This combination elicited enhanced sensitization of colon cancer cells in vitro to the prodrug CB1954, resulting in reduced tumor growth over five weeks in animal models [48]. Another pro-drug activating enzyme that has been used is carboxypeptidase G2 (CPG2), in an Ad OV platform under the control of an hTERT promoter. Following vector delivery, the administered prodrug ZD2767 was converted by CPG2 into a cytotoxic drug and resulted in tumor growth regression or complete tumor eradication in xenografts [49].
Moreover, the previously described Ad Telomelysin (OBP-301) was modified to yield the construct Telomelysin (OBP-405) that contains an RGD motif in the HI loop of the fiber knob. This modification allows for the ability to overcome the limitation of CAR expression in target cells [50].
Congruent with previous OVs using dual gene virotherapy, the study exhibited promising results in colon cancer cells by using manganese superoxide dismutase as a potential tumor suppressor gene along with the tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) gene embedded in an Ad platform with a deleted E1B-55 kD region. The complete elimination of tumors in xenograft models in vivo was reported in [51].
Furthermore, ST13, a colorectal cancer-specific tumor suppressor gene, was inserted into a CRAds under the control of CEA promoter resulting in significant levels of apoptosis in colon cancer cells [18]. Another study used the colon cancer-specific gene TAp63 in an Ad under the survivin promoter. This construct provided selective replication in HCT116 cells with minimal toxicity in L02 cells [52].
Strategies targeting cancer stem cell/stem cell-like markers are an additional promising approach in OV research. One such construct involved the use of an Ad with a CD133-targeting motif resulting in promising efficacy both in vitro and in vivo [53].
4. Clinical Trials and Translational Period of Research
A clinical trial testing Telomelysin in esophageal cancer is eagerly awaited, as the first Ad trial for this type of cancer. As for gastric cancer, some trials have evaluated viral vectors in advanced peritoneal disease, along with ovarian cancer, but none has focused solely on gastric cancer and Ad OV. In the case of pancreatic cancer, primary liver cancers, and colorectal cancer, a number of clinical trials are ongoing (Table 8). Data emerging from these studies will inform the design of the next generation of Ad vectors. It is widely recognized that vector delivery is a challenging step in the eventual application of OV, and advances in this realm are critically needed to address adenoviral liver immune clearance, improve the intra-tumoral viral spread and bystander killing effect, enhance tumor-cell infection, and provide tumor-specific immunity [54].
5. Conclusions
Adenoviral vectors represent a promising platform for cancer therapy in GI cancers as highlighted in this review. A number of exciting strategies applied to OV vector design have facilitated tumor selectivity, potent cytotoxicity, and tumor microenvironment modulation. A broad array of pre-clinical evaluations in GI cancers is showing promise, and a number of these concepts have been carried forward to early phase clinical studies. It is anticipated that future efforts will encompass the study of OV Ads in combination with immunotherapies such as immune checkpoint inhibitors, improvements in the vector delivery/evasion of immune response, and amalgamation with gene editing approaches.
Author Contributions
All authors contributed equally to this review. Raquel Yokoda and Mitesh Borad contributed to the concept and the research, and to writing and editing. Bolni Nagalo contributed to the concept and to writing, proofreading, and editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Douaiher, J.; Ravipati, A.; Grams, B.; Chowdhury, S.; Alatise, O.; Are, C. Colorectal cancer-global burden, trends, and geographical variations. J. Surg. Oncol. 2017, 115, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Howlader, N.N.A.; Krapcho, M.; Miller, D.; Bishop, K.; Kosary, C.L.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D.R.; et al. (Eds.) SEER Cancer Statistics Review 1975–2014; National Cancer Institute: Bethesda, MD, USA, 2017.
- Alemany, R. Oncolytic Adenoviruses in Cancer Treatment. Biomedicines 2014, 2, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.P.; Seto, J.; Liu, E.B.; Dehghan, S.; Hudson, N.R.; Lukashev, A.N.; Ivanova, O.; Chodosh, J.; Dyer, D.W.; Jones, M.S.; et al. Computational analysis of two species C human adenoviruses provides evidence of a novel virus. J. Clin. Microbiol. 2011, 49, 3482–3490. [Google Scholar] [CrossRef] [PubMed]
- Andtbacka, R.H.; Kaufman, H.L.; Collichio, F.; Amatruda, T.; Senzer, N.; Chesney, J.; Delman, K.A.; Spitler, L.E.; Puzanov, I.; Agarwala, S.S.; et al. Talimogene Laherparepvec Improves Durable Response Rate in Patients With Advanced Melanoma. J. Clin. Oncol. 2015, 33, 2780–2788. [Google Scholar] [CrossRef] [PubMed]
- Old, M.O.; Wise-Draper, T.; Wright, C.L.; Kaur, B.; Teknos, T. The current status of oncolytic viral therapy for head and neck cancer. World J. Otorhinolaryngol. Head Neck Surg. 2016, 2, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Lahdeaho, M.L.; Lehtinen, M.; Rissa, H.R.; Hyoty, H.; Reunala, T.; Maki, M. Antipeptide antibodies to adenovirus E1b protein indicate enhanced risk of celiac disease and dermatitis herpetiformis. Int. Arch. Allergy Immunol. 1993, 101, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Operario, D.J.; Platts-Mills, J.A.; Nadan, S.; Page, N.; Seheri, M.; Mphahlele, J.; Praharaj, I.; Kang, G.; Araujo, I.T.; Leite, J.P.G.; et al. Etiology of Severe Acute Watery Diarrhea in Children in the Global Rotavirus Surveillance Network Using Quantitative Polymerase Chain Reaction. J. Infect. Dis. 2017, 216, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Westerberg, S.; Hagbom, M.; Rajan, A.; Loitto, V.; Persson, B.D.; Allard, A.; Nordgren, J.; Sharma, S.; Magnusson, K.E.; Arnberg, N.; et al. Interaction of human enterochromaffin cells with human enteric adenovirus 41 leads to serotonin release and subsequent activation of enteric glia cells. J. Virol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa, T.; Matsuoka, J.; Yamatsuji, T.; Maeda, Y.; Durbin, M.L.; Naomoto, Y. Adenovirus-mediated cancer gene therapy and virotherapy. Int. J. Mol. Med. 2010, 25, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Ono, H.A.; Davydova, J.G.; Adachi, Y.; Takayama, K.; Barker, S.D.; Reynolds, P.N.; Krasnykh, V.N.; Kunisaki, C.; Shimada, H.; Curiel, D.T.; et al. Promoter-controlled infectivity-enhanced conditionally replicative adenoviral vectors for the treatment of gastric cancer. J. Gastroenterol. 2005, 40, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Pin, R.H.; Reinblatt, M.; Fong, Y. Utilizing alpha-fetoprotein expression to enhance oncolytic viral therapy in hepatocellular carcinoma. Ann. Surg. 2004, 240, 659–665. [Google Scholar] [PubMed]
- Wang, Y.; Liu, T.; Huang, P.; Zhao, H.; Zhang, R.; Ma, B.; Chen, K.; Huang, F.; Zhou, X.; Cui, C.; et al. A novel Golgi protein (GOLPH2)-regulated oncolytic adenovirus exhibits potent antitumor efficacy in hepatocellular carcinoma. Oncotarget 2015, 6, 13564–13578. [Google Scholar] [CrossRef] [PubMed]
- Yano, S.; Tazawa, H.; Hashimoto, Y.; Shirakawa, Y.; Kuroda, S.; Nishizaki, M.; Kishimoto, H.; Uno, F.; Nagasaka, T.; Urata, Y.; et al. A genetically engineered oncolytic adenovirus decoys and lethally traps quiescent cancer stem-like cells in S/G2/M phases. Clin. Cancer Res. 2013, 19, 6495–6505. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ji, W.; Hu, H.; Ma, J.; Li, X.; Mei, W.; Xu, Y.; Hu, H.; Yan, Y.; Song, Q.; et al. Survivin promoter-regulated oncolytic adenovirus with Hsp70 gene exerts effective antitumor efficacy in gastric cancer immunotherapy. Oncotarget 2014, 5, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.Y.; Wang, K.L.; Fang, F.F.; Gu, W.; Huang, F.; Wang, F.Z.; Li, B.; Wang, L.N. Triple-controlled oncolytic adenovirus expressing melittin to exert inhibitory efficacy on hepatocellular carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 10403–10411. [Google Scholar] [PubMed]
- Araki, Y.; Fujiwara, H.; Inada, S.; Atsuji, K.; Yamagishi, H. [An antitumor effect of oncolytic adenovirus capable of selectively replicating in CEA-expressing cancer cells and its enhancement by 5-FU]. Gan Kagaku Ryoho 2006, 33, 1754–1755. [Google Scholar]
- Zhou, X.; Xie, G.; Wang, S.; Wang, Y.; Zhang, K.; Zheng, S.; Chu, L.; Xiao, L.; Yu, Y.; Zhang, Y.; et al. Potent and specific antitumor effect for colorectal cancer by CEA and Rb double regulated oncolytic adenovirus harboring ST13 gene. PLoS ONE 2012, 7, e47566. [Google Scholar] [CrossRef] [PubMed]
- Koski, A.; Karli, E.; Kipar, A.; Escutenaire, S.; Kanerva, A.; Hemminki, A. Mutation of the fiber shaft heparan sulphate binding site of a 5/3 chimeric adenovirus reduces liver tropism. PLoS ONE 2013, 8, e60032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. Cancer Stat. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Moriyama, H.; Yasuda, H.; Hara, K.; Maniwa, Y.; Hamada, H.; Yokono, K.; Nagata, M. Modification of the Rb-binding domain of replication-competent adenoviral vector enhances cytotoxicity against human esophageal cancers via NF-κB activity. Hum. Gene Ther. 2007, 18, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Komaki, R.; Wang, L.; Fang, B.; Chang, J.Y. Treatment of radioresistant stem-like esophageal cancer cells by an apoptotic gene-armed, telomerase-specific oncolytic adenovirus. Clin. Cancer Res. 2008, 14, 2813–2823. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Kawamura, K.; Li, Q.; Okamoto, S.; Suzuki, N.; Kobayashi, H.; Liang, M.; Tada, Y.; Tatsumi, K.; Hiroshima, K.; et al. Combinatory cytotoxic effects produced by E1B-55kDa-deleted adenoviruses and chemotherapeutic agents are dependent on the agents in esophageal carcinoma. Cancer Gene Ther. 2010, 17, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T. A novel molecular therapy using bioengineered adenovirus for human gastrointestinal cancer. Acta Med. Okayama 2011, 65, 151–162. [Google Scholar] [PubMed]
- Zheng, H.; Li, M.S.; Zhao, G.Q.; Dong, Z.M. [Effect of CEA gene regulation on the anti-tumor activity of oncolytic adenovirus H101 to esophageal carcinoma]. Zhonghua Zhong Liu Za Zhi 2011, 33, 822–826. [Google Scholar] [PubMed]
- He, D.; Sun, L.; Li, C.; Hu, N.; Sheng, Y.; Chen, Z.; Li, X.; Chi, B.; Jin, N. Anti-tumor effects of an oncolytic adenovirus expressing hemagglutinin-neuraminidase of Newcastle disease virus in vitro and in vivo. Viruses 2014, 6, 856–874. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, M.S.; Boehm, T.; Shing, Y.; Fukai, N.; Vasios, G.; Lane, W.S.; Flynn, E.; Birkhead, J.R.; Olsen, B.R.; Folkman, J. Endostatin: An endogenous inhibitor of angiogenesis and tumor growth. Cell 1997, 88, 277–285. [Google Scholar] [CrossRef]
- Jiang, K.; Al-Diffhala, S.; Centeno, B.A. Primary Liver Cancers-Part 1: Histopathology, Differential Diagnoses, and Risk Stratification. Cancer Control 2018, 25. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yu, D.C.; Chen, Y.; Amin, P.; Zhang, H.; Nguyen, N.; Henderson, D.R. A hepatocellular carcinoma-specific adenovirus variant, CV890, eliminates distant human liver tumors in combination with doxorubicin. Cancer Res 2001, 61, 6428–6436. [Google Scholar] [PubMed]
- Heise, C.; Sampson-Johannes, A.; Williams, A.; McCormick, F.; Von Hoff, D.D.; Kirn, D.H. ONYX-015, an E1B gene-attenuated adenovirus, causes tumor-specific cytolysis and antitumoral efficacy that can be augmented by standard chemotherapeutic agents. Nat. Med. 1997, 3, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Sham, J.; Yang, J.; Su, C.; Xue, H.; Chua, D.; Sun, L.; Zhang, Q.; Cui, Z.; Wu, M.; et al. Potent antitumor efficacy of an E1B 55kDa-deficient adenovirus carrying murine endostatin in hepatocellular carcinoma. Int. J. Cancer 2005, 113, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.C.; Zhang, Q.; Yang, Y.; Lu, M.Q.; Li, H.; Xu, C.; Chen, G.H. p53-expressing conditionally replicative adenovirus CNHK500-p53 against hepatocellular carcinoma in vitro. World J. Gastroenterol. 2007, 13, 683–691. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Lei, W.; Wang, S.; Xiao, R.; Guo, K.; Xia, Y.; Zhou, X.; Zhang, K.; Liu, X.; Wang, Y. Overexpression of tumor suppressor TSLC1 by a survivin-regulated oncolytic adenovirus significantly inhibits hepatocellular carcinoma growth. J. Cancer Res. Clin. Oncol. 2012, 138, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.W.; Zhong, S.Y.; Liu, B.S.; Liu, J.; Cai, R.; Wang, Y.G.; Liu, X.Y.; Qian, C. Enhanced sensitivity of hepatocellular carcinoma cells to chemotherapy with a Smac-armed oncolytic adenovirus. Acta Pharmacol. Sin. 2007, 28, 1996–2004. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.L.; Lee, C.H.; Teo, M.L.; Lin, Y.J.; Huang, Y.S.; Wu, C.L.; Shiau, A.L. Transthyretin-driven oncolytic adenovirus suppresses tumor growth in orthotopic and ascites models of hepatocellular carcinoma. Cancer Sci. 2009, 100, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Lv, S.; Yang, J.; Wang, X.; Hu, H.; Su, C.; Zhou, C.; Li, J.; Huang, Y.; Li, L.; et al. Use of microRNA Let-7 to control the replication specificity of oncolytic adenovirus in hepatocellular carcinoma cells. PLoS ONE 2011, 6, e21307. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Su, Y.; Sun, B.; Ji, W.; Peng, Z.; Xu, Y.; Wu, M.; Su, C. An Artificially Designed Interfering lncRNA Expressed by Oncolytic Adenovirus Competitively Consumes OncomiRs to Exert Antitumor Efficacy in Hepatocellular Carcinoma. Mol. Cancer Ther. 2016, 15, 1436–1451. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.C.; Cao, X.; Gui, J.H.; Zhou, X.M.; Zhong, D.; Yan, Q.L.; Huang, W.D.; Qian, Q.J.; Zhao, F.L.; Liu, X.Y. Augmenting the antitumor effect of TRAIL by SOCS3 with double-regulated replicating oncolytic adenovirus in hepatocellular carcinoma. Hum. Gene Ther. 2011, 22, 1109–1119. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, W.; Wei, X.; Cui, Q.; Lou, W.; Wang, G.; Hu, X.; Qian, C. Potent antitumor activity of oncolytic adenovirus-mediated SOCS1 for hepatocellular carcinoma. Gene Ther. 2013, 20, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, H.; Li, L.; Wu, H.; Wang, C.; Yan, Z.; Wang, Y.; Su, C.; Jin, H.; Zhou, F.; et al. The combination of an oxygen-dependent degradation domain-regulated adenovirus expressing the chemokine RANTES/CCL5 and NK-92 cells exerts enhanced antitumor activity in hepatocellular carcinoma. Oncol. Rep. 2013, 29, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Ma, B.; Wang, Y.; Xiao, R.; Kong, Y.; Zhou, X.; Xia, D. Targeting gene-virus-mediated manganese superoxide dismutase effectively suppresses tumor growth in hepatocellular carcinoma in vitro and in vivo. Cancer Biother. Radiopharm. 2014, 29, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Wang, Y.; Zhou, X.; Huang, P.; Zhang, R.; Liu, T.; Cui, C.; Liu, X.; Wang, Y. Synergistic suppression effect on tumor growth of hepatocellular carcinoma by combining oncolytic adenovirus carrying XAF1 with cisplatin. J. Cancer Res. Clin. Oncol. 2015, 141, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Sunamura, M.; Hamada, H.; Motoi, F.; Oonuma, M.; Abe, H.; Saitoh, Y.; Hoshida, T.; Ottomo, S.; Omura, N.; Matsuno, S. Oncolytic virotherapy as a novel strategy for pancreatic cancer. Pancreas 2004, 28, 326–329. [Google Scholar] [CrossRef] [PubMed]
- Freytag, S.O.; Barton, K.N.; Brown, S.L.; Narra, V.; Zhang, Y.; Tyson, D.; Nall, C.; Lu, M.; Ajlouni, M.; Movsas, B.; et al. Replication-competent adenovirus-mediated suicide gene therapy with radiation in a preclinical model of pancreatic cancer. Mol. Ther. 2007, 15, 1600–1606. [Google Scholar] [CrossRef] [PubMed]
- Jose, A.; Sobrevals, L.; Miguel Camacho-Sanchez, J.; Huch, M.; Andreu, N.; Ayuso, E.; Navarro, P.; Alemany, R.; Fillat, C. Intraductal delivery of adenoviruses targets pancreatic tumors in transgenic Ela-myc mice and orthotopic xenografts. Oncotarget 2013, 4, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Jose, A.; Rovira-Rigau, M.; Luna, J.; Gimenez-Alejandre, M.; Vaquero, E.; Garcia de la Torre, B.; Andreu, D.; Alemany, R.; Fillat, C. A genetic fiber modification to achieve matrix-metalloprotease-activated infectivity of oncolytic adenovirus. J. Control Release 2014, 192, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Hiraoka, N.; Goto, N.; Rin, Y.; Miura, K.; Narumi, K.; Uchida, H.; Tagawa, M.; Aoki, K. A targeting ligand enhances infectivity and cytotoxicity of an oncolytic adenovirus in human pancreatic cancer tissues. J. Control Release 2014, 192, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Green, N.K.; Reynolds, G.M.; Flavell, J.R.; Mautner, V.; Kerr, D.J.; Young, L.S.; Searle, P.F. Enhanced efficacy of Escherichia coli nitroreductase/CB1954 prodrug activation gene therapy using an E1B-55K-deleted oncolytic adenovirus vector. Gene Ther. 2004, 11, 1126–1136. [Google Scholar] [CrossRef] [PubMed]
- Schepelmann, S.; Ogilvie, L.M.; Hedley, D.; Friedlos, F.; Martin, J.; Scanlon, I.; Chen, P.; Marais, R.; Springer, C.J. Suicide gene therapy of human colon carcinoma xenografts using an armed oncolytic adenovirus expressing carboxypeptidase G2. Cancer Res 2007, 67, 4949–4955. [Google Scholar] [CrossRef] [PubMed]
- Taki, M.; Kagawa, S.; Nishizaki, M.; Mizuguchi, H.; Hayakawa, T.; Kyo, S.; Nagai, K.; Urata, Y.; Tanaka, N.; Fujiwara, T. Enhanced oncolysis by a tropism-modified telomerase-specific replication-selective adenoviral agent OBP-405 (‘Telomelysin-RGD’). Oncogene 2005, 24, 3130–3140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gu, J.; Zhao, L.; He, L.; Qian, W.; Wang, J.; Wang, Y.; Qian, Q.; Qian, C.; Wu, J.; et al. Complete elimination of colorectal tumor xenograft by combined manganese superoxide dismutase with tumor necrosis factor-related apoptosis-inducing ligand gene virotherapy. Cancer Res 2006, 66, 4291–4298. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Liu, H.; Zhang, Z.; Basnet, S.; Dai, Z.; Li, S.; Wang, Y.; Xu, B.; Ge, H. A dual-regulated oncolytic adenovirus carrying TAp63 gene exerts potent antitumor effect on colorectal cancer cells. Am. J. Transl. Res. 2017, 9, 2966–2974. [Google Scholar] [PubMed]
- Sato-Dahlman, M.; Miura, Y.; Huang, J.L.; Hajeri, P.; Jacobsen, K.; Davydova, J.; Yamamoto, M. CD133-targeted oncolytic adenovirus demonstrates anti-tumor effect in colorectal cancer. Oncotarget 2017, 8, 76044–76056. [Google Scholar] [CrossRef] [PubMed]
- Yokoda, R.; Nagalo, B.M.; Vernon, B.; Oklu, R.; Albadawi, H.; DeLeon, T.T.; Zhou, Y.; Egan, J.B.; Duda, D.G.; Borad, M.J. Oncolytic virus delivery: From nano-pharmacodynamics to enhanced oncolytic effect. Oncolytic Virother. 2017, 6, 39–49. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
Table 1.
Natural tropism of adenoviral platforms.
| Subgroup | Serotype | Attachment Receptors | Natural Tropism |
|---|---|---|---|
| A | 12 | CAR | Gastrointestinal |
| B1 | 35 | CD46 | Respiratory |
| B2 | 3 | DSG-2 | Renal |
| B3 | 11 | CD46 and DSG-2 | Renal |
| C | 2 and 5 | CAR | Respiratory |
| D | 19 | CAR and sialic acid | Ocular |
| E | 4 | CAR | Respiratory and Ocular |
| F | 40 and 41 | CAR | Gastrointestinal |
| G | 52 | unknown | Gastrointestinal |
CAR: coxsackie-adenovirus receptor; DSG-2: desmoglein-2.
Table 2.
Adenoviral platform and incorporated promoters for gastrointestinal malignancies.
| Virus Promoter | GI Cancer | Ref. |
|---|---|---|
| Cytomegalovirus (CMV) | Ubiquitous | [10] |
| Midkine (MK) | Gastric | [11] |
| Cyclooxygenase-2 (Cox-2M) and (Cox-2L) | Gastric | [11] |
| Alpha-fetoprotein (AFP) | HCC | [12] |
| Golgi protein 73 | HCC | [13] |
| Human telomerase reverse transcriptase (hTERT) | Cancer cells in general | [14] |
| Survivin | Cancer cells in general | [15] |
| Chimeric: Hypoxia-response element (HRE) and Alpha-fetoprotein (AFP) | HCC | [16] |
| Carcinoembryonic antigen (CEA) | Gastric cancer cells and Colon cancer cells | [17,18] |
HCC: Hepatocellular carcinoma; GI: Gastrointestinal.
Table 3.
Preclinical research in esophageal cancer.
| Viral Construct Name | In Vitro Cell Line | In Vivo Model | Vector Modifications | Conclusion | Ref. |
|---|---|---|---|---|---|
| AxE1AdB | EC-GI-10 T.Tn. | CB17 scid mouse CDX Heterotopic subcutanous transplant | E1A gene abolish binding to pRB | Enhanced apoptosis, and cytotoxicity against p53-mutant cells | [21] |
| Ad/TRAIL-E1 | Seg-1 Seg-1 with Radioresistance (R) TE-2, and TE-2R | Nude mice CDX Heterotopic subcutanous transplant | hTERT promoter controlling E1A | Ad/TRAIL-E1 preferentially targeted radioresistant-cells | [22] |
| Ad-delE1B55 | TE-1 TE-2 TE-10 TE-11 YES-2 YES-4 YES-5 YES-6 T. Tn | Nude mice CDX Heterotopic subcutanous transplant | CMV Promoter Deleted a part of E3 region and 55 kDa-encoding E1B region | The combinatory antitumor effect depends on the chemotherapy agent | [23] |
| Telomelysin (OBP-301) | A549 H1299 | Nude mice PDX Orthotopic transplant | hTERT Promoter Deleted a part of E3 region and 55 kDa-encoding E1B region | A substantial anti-tumor effect was achieved when radiation followed the intratumoral injection | [24] |
| Ad-hTERTp-E1a-HN | EC109 | Nude mice CDX Heterotopic subcutaneous transplant | hTERT Promoter Deleted a part of E3 region and 55 kDa-encoding E1B region Expressing HN from NDV | Suppression in tumor volume in both delivery modes IT and IVComplete response to vector IT injection | [26] |
Abbreviations: Scid: severe combined immunodeficient; CDX: cancer cell-line-derived xenotransplant; PDX: patient-derived xenotransplant; pRB: Retinoblastoma protein; CMV: cytomegalovirus; hTERT: human telomerase reverse transcriptase; TRAIL: TNF-related apoptosis-inducing ligand; HN: hemagglutinin-neuraminidase; NDV: Newcastle disease virus; IT: intratumor; IV: intravenously.
Table 4.
Preclinical research in gastric cancer.
| Viral Construct Name | In Vitro Cell Line | In Vivo Model | Vector Modifications | Conclusion | Ref. |
|---|---|---|---|---|---|
| AdCEAp/Rep | MKN-45 MKN-74 | - | CEA promoter | Cytotoxicity against CEA producing cells was dose-dependent | [17] |
| Telomelysin (OBP-301) | MKN-45 MKN-74 | NOD/SCID Mice CDX Heterotopic transplant | hTERT promoter | Cell death in quiescent CD133+ cells | [14] |
| E1B 55-kDa-attenuated Ad | AGS MGc80-3 | C57Bl6/J Mice CDX Heterotopic transplant | E1B 55-kDa-deficient Ad expressing Endostatin | Synergistic effect | [27] |
| AdSurp-Hsp70 | SGC-7901 BCG-823 MNK-45 | Nude mice CDX Heterotopic transplant | Survivin promoter Vector expressing chaperone Hsp-70 | Selective replication in survivin-positive tumor cells | [15] |
Abbreviations: CEA: carcinoembryonic antigen; hTERT: human telomerase reverse Transcriptase; NOD/SCID: non-obese diabetic/severe combined immunodeficient; CDX: cancer cell-line-derived xenotransplant.
Table 5.
Preclinical research in liver cancer.
| Viral Construct Name | In Vitro Cell Line | In Vivo Model | Vector Modifications | Conclusion | Ref. |
|---|---|---|---|---|---|
| CV890 | HepG2 Huh7 Hep3B PLC/PRF/5 SNU449 | Nude mice CDX Heterotopic subcutaneous transplant | AFP TRE to control an artificial E1A-IRES-E1B bicistronic cassette in an adenovirus 5 vector | Volume of distant xenografts dropped below baseline at 4 weeks | [29] |
| CNH500-p53 | Hep3B HepG2 SMMC-7721 | - | hTERT promoter and HRE promoter | Higher oncolytic effect | [32] |
| ZD55-Smac | Bel-7404 SMMC7721 Huh-7 | - | Incorporation of therapeutic gene: ZD-Smac Under CMV promoter | ZD55-Smac was superior to ONYX-015 | [34] |
| AFP-D55-SOC3 | Hep3B PLC HepG2 Huh-7 LM6 BEL7404 | Nude mice CDX Heterotopic subcutaneous transplant | SOCS3 downregulate Cyclin D1 and anti-apoptotic proteins such as XIAP, Survivin, Bcl-xL, and Mcl-1 | Restoration of SOCS3 antagonize HCC therapeutic resistance to TRAIL | [38] |
| SD55-TSLC1 | Huh-7 | Nude mice CDX Heterotopic subcutaneous transplant | Expression of TSLC1 a tumor suppressor gene | Caspase pathways provide antitumor effect | [33] |
| AdCN305-SOCS1 | Bel-7404 Hep3B Huh-7 SMMC7721 | Nude mice CDX Heterotopic subcutaneous transplant | Expression of an SOCS1 a negative regulator of STAT3 | Inhibition of STAT3 phosphorylation and downregulation of survivin, cyclin-D1, Bcl-xL, and C-myc | [39] |
| AdCN205-IL-24-miR-34a | PLC/PRF/5 Huh7 Bel-7404 | Nude mice CDX Heterotopic subcutaneous transplant | Co-expression of miRNA-34a and IL-24 | Complete tumor regression | [36] |
| QG511-HA-Melittin | Hep3B SMMC7721 HepG2 | Nude mice CDX Heterotopic subcutaneous transplant | hybrid promoter, hypoxia-response element and alpha-fetoprotein (HRE)-AFP | Inhibit the growth of HCC xenografts | [16] |
| AdSVPE1a-lncR | Huh-7 HepG2 SMMC7721 Hep3B L02 | Nude mice CDX Heterotopic subcutaneous transplant | Long noncoding RNA expression under a surviving promoter | Competitively consumes OncomiRs (oncogenic miRNAs) promoting tumor shrinkage | [37] |
Abbreviations: CDX: cancer cell-line-derived xenotransplant; hTERT: human telomerase reverse transcriptase; HCC: hepatocellular carcinoma; SOCS1: suppressor of cytokine signaling 1; SOCS3: suppressor of cytokine signaling 3; TRAIL: TNF-related apoptosis-inducing ligand.
Table 6.
Preclinical research in pancreatic cancer.
| Viral Construct Name | In Vitro Cell Line | In Vivo Model | Vector Modifications | Conclusion | Ref. |
|---|---|---|---|---|---|
| Ad5-yCD/mutTKSR39rep-ADP | Panc 1 MiaPaCa-2 | Nude mice CDX Heterotopic subcutaneous transplant; GEMM, CDX Orthotopic transplant | Contains a bacterial cytosine deaminase (CD) and wild-type herpes simplex virus thymidine kinase (HSV-1 TK) gene under CMV promoter | Improved the effectiveness of radiotherapy without excessive toxicity | [44,45] |
| AdTATMMP | RWP1 Panc1 PSC-21 CAF-25 CAF-28 | GEMM CDX orthotopic transplant | AdTATMMP transduction is activated by matrix metalloproteases MMP2 and MMP9 | In comparison to Ad5 wild type, there was increased antitumor activity | [46] |
| AdSur-SYE | AsPC-1 BxPC-3 Panc-1 MIAPaCa-2 | Nude mice CDX Heterotopic subcutaneous transplant | Displays a pancreatic cancer targeting sequence SYENFSA on the fiber knob; survivin promoter | High infectivity in human pancreatic cancer tissues | [47] |
Abbreviations: GEMM: Genetically engineered mouse model; CDX: cancer cell-line-derived xenotransplant.
Table 7.
Preclinical research in colorectal cancer.
| Viral Construct Name | In Vitro Cell Line | In Vivo Model | Vector Modifications | Conclusion | Ref |
|---|---|---|---|---|---|
| CRAd-NTR(PS1217H6) | SW480 | Nude mice CDX Heterotopic subcutaneous transplant | Vector E1B-55-KDa-deleted expressing prodrug-activating enzyme nitroreductase (NTR) | Greater sensitization to the prodrug CB1954 | [48] |
| Telomelysin OBP-405 | SW620 | Nude miceCDX Heterotopic subcutaneous transplant | Vector has mutant fiber containing the RGD peptide, CDCRGDCFC, in the HI loop of the fiber knob | Increased infection property | [50] |
| ZD55-MnSOD ZD55-TRAIL | SW620 | Nude mice CDX Heterotopic Random tumor inoculation | Vector with the E1B 55-kDa gene deletion and expressing Manganese superoxide dismutase (MnSOD) | Effective oncolysis | [51] |
| AdV.hTERT-CPG2 | SW620 SW480 HCT116 LS174T LoVo DLD-1 HT-29 Caco-2 Colo205 | Nude mice CDX Heterotopic subcutaneous transplant | Delivery of gene for the prodrug-activating enzyme carboxypeptidase G2 (CPG2) to tumors | significant bystander effects in vivo | [49] |
| Ad·(ST13)·CEA·E1A(Δ24) | SW620 HT29 HCT116 | Nude mice CDX Heterotopic subcutaneous transplant | Vector with CEA promoter expressing suppression of ST13 | Induced tumor apoptosis through the mitochondrial-mediated apoptosis pathway | [18] |
| Ad-survivin-ZD55-TAp63 | HCT116 | Nude mice CDX Heterotopic subcutaneous transplant | Tap63 expressing cassette in Adenovirus under surviving | in vitro inhibition of cell proliferation | [52] |
| AdML-TYML | LoVo LS174T | Nude mice CDX Heterotopic subcutaneous transplant | A TYMLSRN peptide motif in place of the primary CAR-binding domains in AB-loop of fiber knob | Selective virus for CSC | [53] |
Abbreviations: CDX: cancer cell-line-derived xenotransplant; CEA: carcinoembryonic antigen; CSC: cancer stem-like cells; ST13: tumorigenicity 13.
Table 8.
Clinical trials using adenoviral oncolytic therapy in gastrointestinal cancers.
| GI Cancer | Vector Construct | Phase | Country | Clinicaltrials.gov Number |
|---|---|---|---|---|
| Pancreatic cancer | Ad5-yCD/mutTKSR39rep-hIL12 (Oncolytic adenovirus expressing two suicide genes and human IL-12) | I | USA | NCT03281382 |
| Pancreatic cancer | LOAd703 Oncolytic adenovirus serotype 5/35 encoding TMZ-CD40L and 4-1BBL | I/II | Sweden | NCT03225989 |
| Pancreatic cancer | LOAd703 | I/II | USA | NCT02705196 |
| Pancreatic cancer | VCN-1 expressing PH20 hyaluronidase | I | Spain | NCT02045602 |
| Pancreatic cancer | VCN-1 expressing PH20 hyaluronidase | I | Spain | NCT02045589 |
| Hepatocellular carcinoma | Telomesyn OBP-301 | I/II | Korea & Taiwan | NCT02293850 |
| Hepatocellular carcinoma | Recombinant Ad5 | III | China | NCT01869088 |
| Liver Cancer | Ad5-CMV-p53 | I | USA | NCT00003147 |
| Colorectal cancer | LOAd703 | I/II | Sweden | NCT03225989 |
| Colorectal cancer | Ad11/Ad3 Enadenotucirev (previously ColoAd1) | I/II | Belgium & Spain | NCT02028442 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yokoda, R.T.; Nagalo, B.M.; Borad, M.J. Oncolytic Adenoviruses in Gastrointestinal Cancers. Biomedicines 2018, 6, 33. https://doi.org/10.3390/biomedicines6010033
AMA Style
Yokoda RT, Nagalo BM, Borad MJ. Oncolytic Adenoviruses in Gastrointestinal Cancers. Biomedicines. 2018; 6(1):33. https://doi.org/10.3390/biomedicines6010033
Chicago/Turabian StyleYokoda, Raquel T., Bolni M. Nagalo, and Mitesh J. Borad. 2018. "Oncolytic Adenoviruses in Gastrointestinal Cancers" Biomedicines 6, no. 1: 33. https://doi.org/10.3390/biomedicines6010033
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.