Straw Wine Melanoidins as Potential Multifunctional Agents: Insight into Antioxidant, Antibacterial, and Angiotensin-I-Converting Enzyme Inhibition Effects



Abstract
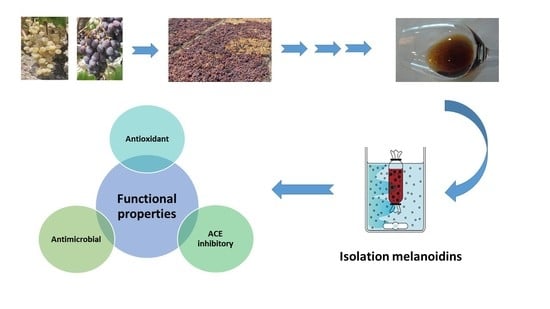
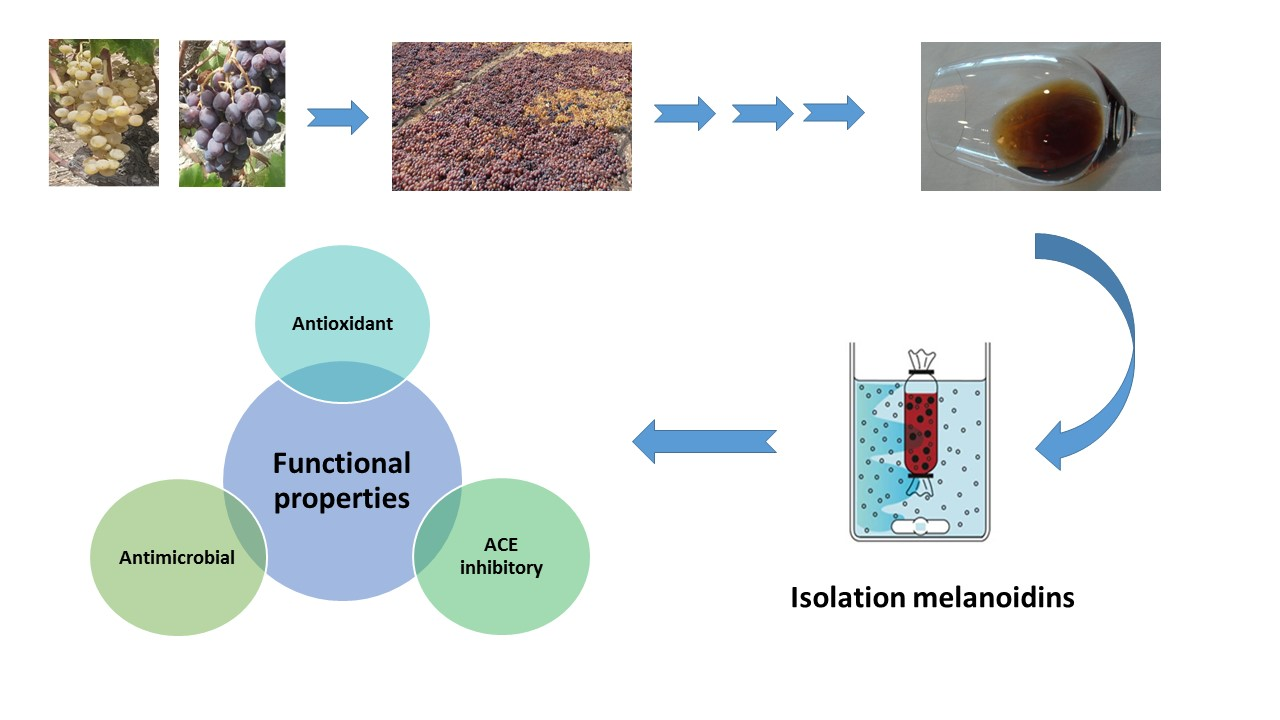
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Samples
2.2. Isolation of Melanoidin Fractions
2.3. Spectroscopic Analysis
2.4. Determination of Total Phenolic Groups Content
2.5. Determination of Total Sugar Content
2.6. Determination of DPPH Radical Scavenging Activity
2.7. Determination of Ferric Reducing Antioxidant Power (FRAP)
2.8. Determination of Antimicrobial Potency
2.9. Determination of Angiotensin-I Converting Enzyme (ACE) Inhibitory Activity
2.10. Statistical Analysis
3. Results and Discussion
3.1. Chemical Characterization of Commandaria Melanoidins
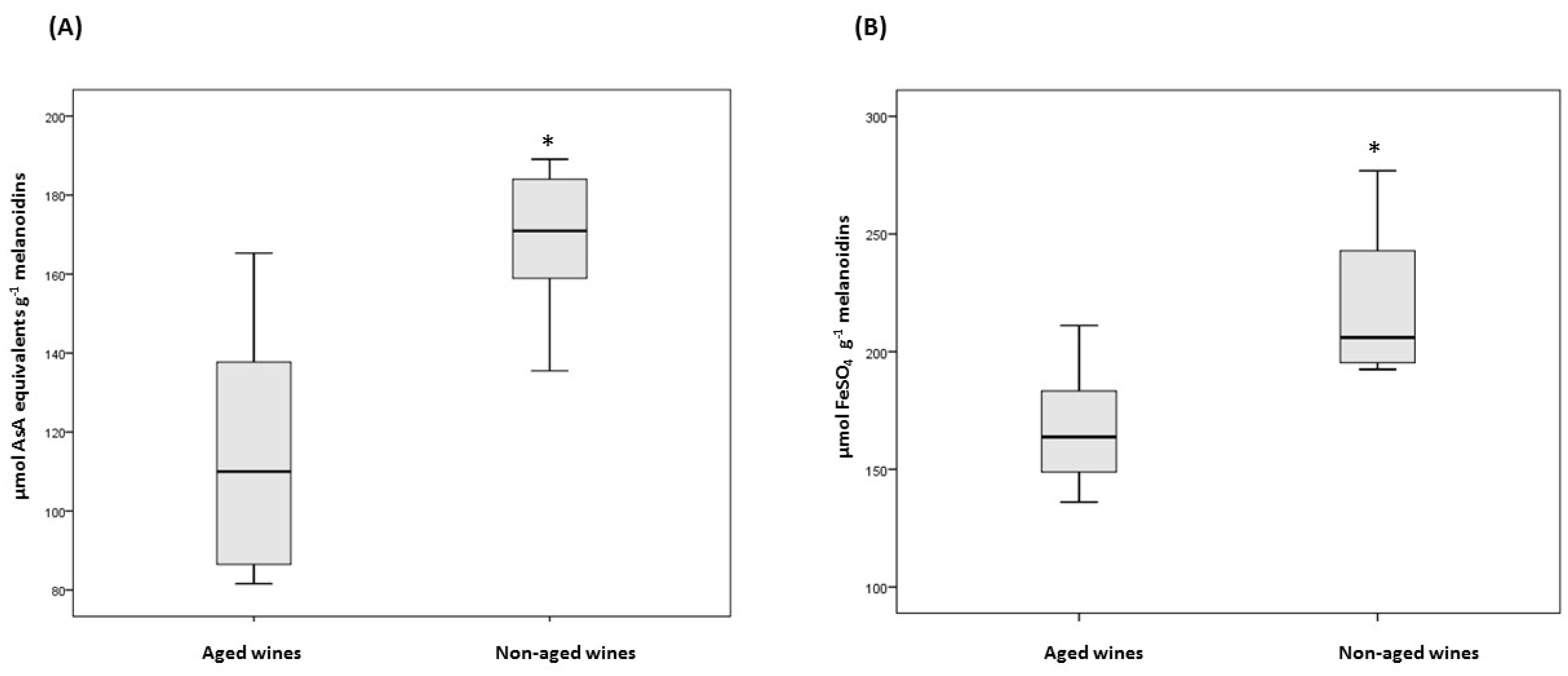
3.2. Antioxidant Activity of Commandaria Melanoidins
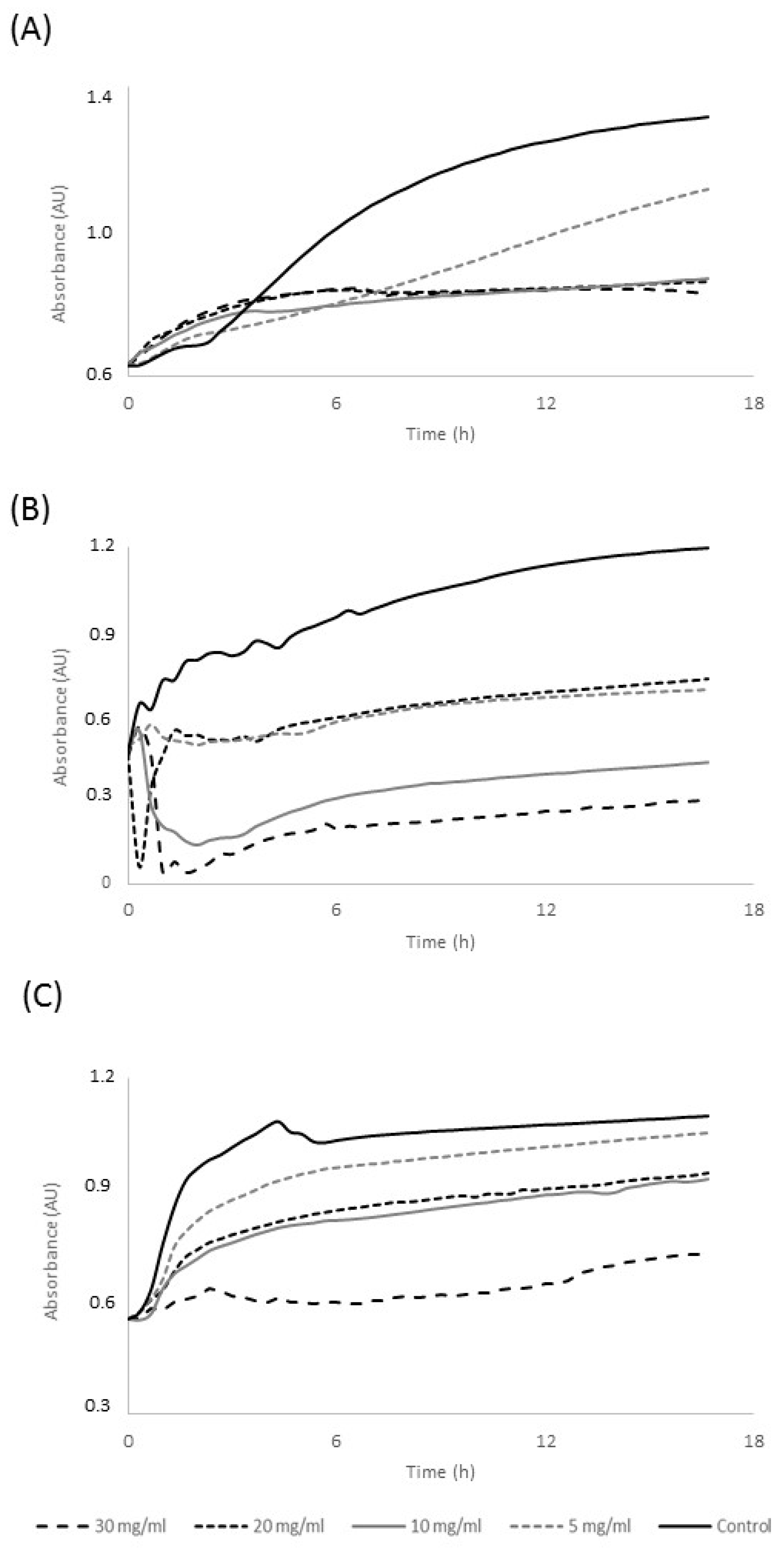
3.3. Antimicrobial Potency of Commandaria Melanoidins
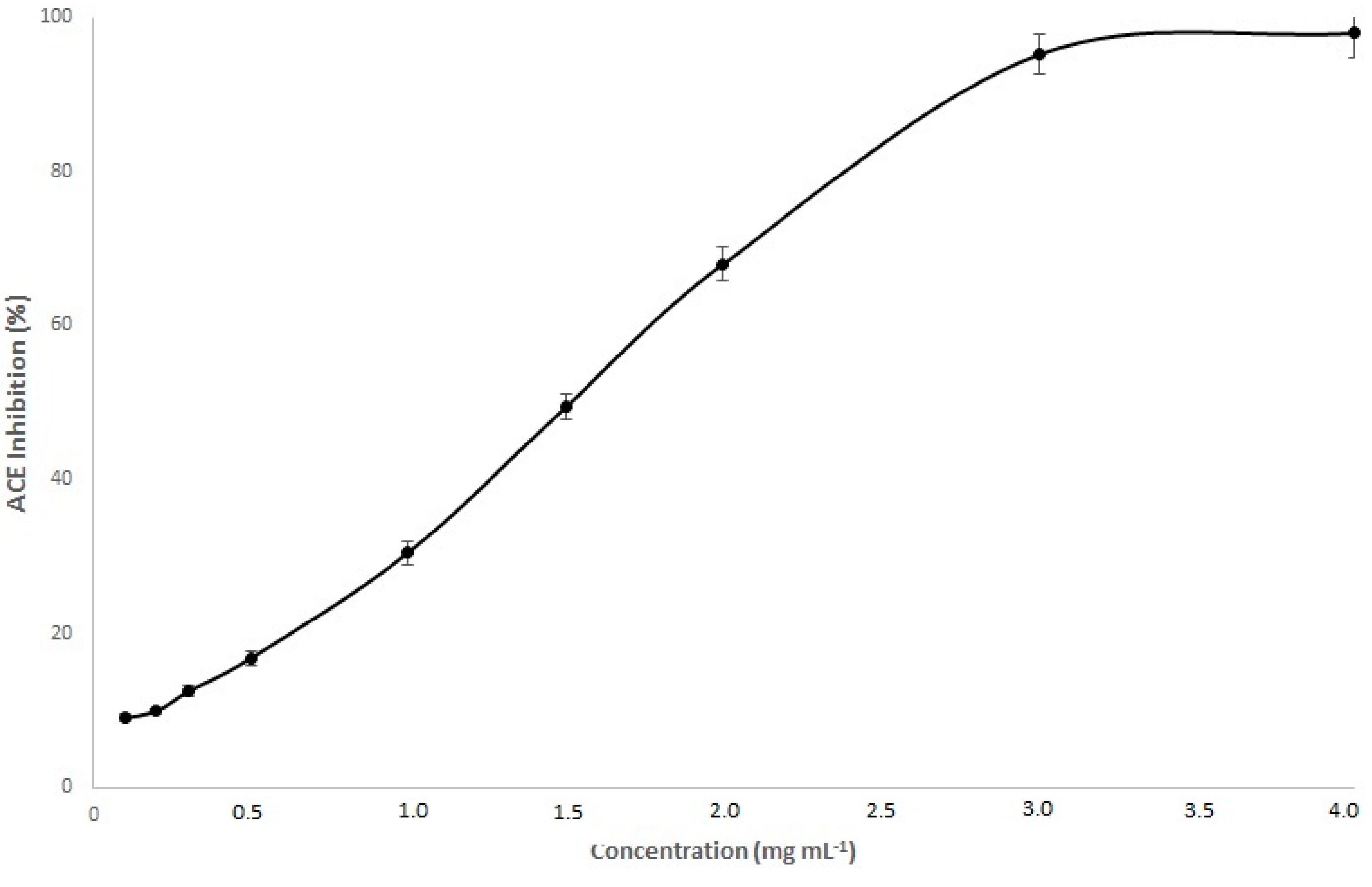
3.4. ACE Inhibitory Effect of Commandaria Melanoidins
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Echavarría, A.P.; Pagán, J.; Ibarz, A. Melanoidins formed by Maillard reaction in food and their biological activity. Food Eng. Rev. 2012, 4, 203–223. [Google Scholar] [CrossRef]
- Morales, F.J.; Somoza, V.; Fogliano, V. Physiological relevance of dietary melanoidins. Amino Acids 2012, 42, 1097–1109. [Google Scholar] [CrossRef] [PubMed]
- Mesías, M.; Delgado-Andrade, C. Melanoidins as a potential functional food ingredient. Curr. Opin. Food Sci. 2017, 14, 37–42. [Google Scholar] [CrossRef]
- Bekedam, E.K.; Loots, M.J.; Schols, H.A.; Van Boekel, M.A.J.S.; Smit, G. Roasting effects on formation mechanisms of coffee brew melanoidins. J. Agric. Food Chem. 2008, 56, 7138–7145. [Google Scholar] [CrossRef] [PubMed]
- Summa, C.; Raposo, F.C.; McCourt, J.; Scalzo, R.L.; Wagner, K.H.; Elmadfa, I.; Anklam, E. Effect of roasting on the radical scavenging activity of cocoa beans. Eur. Food Res. Technol. 2006, 222, 368–375. [Google Scholar] [CrossRef]
- Rivero, D.; Pérez-Magariño, S.; González-Sanjosé, M.L.; Valls-Belles, V.; Codoñer, P.; Muñiz, P. Inhibition of induced DNA oxidative damage by beers: Correlation with the content of polyphenols and melanoidins. J. Agric. Food Chem. 2005, 53, 3637–3642. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Heras, M.; González-Sanjosé, M.L. Binding capacity of brown pigments present in special Spanish sweet wines. LWT Food Sci. Technol. 2009, 42, 1729–1737. [Google Scholar] [CrossRef]
- Rufián-Henares, J.A.; Morales, F.J. Functional properties of melanoidins: In vitro antioxidant, antimicrobial and antihypertensive activities. Food Res. Int. 2007, 40, 995–1002. [Google Scholar] [CrossRef] [Green Version]
- Rufián-Henares, J.A.; De La Cueva, S.P. Antimicrobial activity of coffee melanoidins—A study of their metal-chelating properties. J. Agric. Food Chem. 2009, 57, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, D.; Elena, V.; Angela, C. Effect of dietary melanoidins on lipid peroxidation during simulated gastric digestion: Their possible role in the prevention of oxidative damage. J. Agric. Food Chem. 2010, 58, 2513–2519. [Google Scholar] [CrossRef] [PubMed]
- Cossu, A.; Posadino, A.M.; Giordo, R.; Emanueli, C.; Sanguinetti, A.M.; Piscopo, A.; Poiana, M.; Capobianco, G.; Piga, A.; Pintus, G. Apricot melanoidins prevent oxidative endothelial cell death by counteracting mitochondrial oxidation and membrane depolarization. PLoS ONE 2012, 7, e48817. [Google Scholar] [CrossRef] [PubMed]
- Rufián-Henares, J.A.; Morales, F.J. Angiotensin-I converting enzyme inhibitory activity of coffee melanoidins. J. Agric. Food Chem. 2007, 55, 1480–1485. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, R.C.; Fogliano, V. Bread crust melanoidins as potential prebiotic ingredients. Mol. Nutr. Food Res. 2005, 49, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, D.; Bellesia, A. The gastro-intestinal tract as the major site of biological action of dietary melanoidins. Amino Acids 2015, 47, 1077–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastoriza, S.; Rufián-Henares, J.A. Contribution of melanoidins to the antioxidant capacity of the Spanish diet. Food Chem. 2014, 164, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Di Mattia, C.; Sacchetti, G.; Seghetti, L.; Piva, A.; Mastrocola, D. “Vino cotto” composition and antioxidant activity as affected by non enzymatic browning. Ital. J. Food Sci. 2002, 19, 413–424. [Google Scholar]
- Rivero-Pérez, M.D.; Pérez-Magariño, S.; González-San, J.M.L. Role of melanoidins in sweet wines. Anal. Chim. Acta 2002, 458, 169–175. [Google Scholar] [CrossRef]
- Constantinou, S.; Gomez-Caravaca, A.M.; Goulas, V.; Segura-Carretero, A.; Manganaris, G.A. Metabolic fingerprinting of must obtained from sun-dried grapes of two indigenous Cypriot cultivars destined for the production of “Commandaria”: A protected destignation of origin product. Food Res. Int. 2007, 100, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Constantinou, S.; Gomez-Caravaca, A.M.; Goulas, V.; Segura-Carretero, A.; Koundouras, S.; Manganaris, G.A. The impact of postharvest dehydration methods on qualitative attributes and chemical composition of ‘Xynisteri’ grape (Vitis vinifera) must. Postharvest Biol. Technol. 2018, 135, 114–122. [Google Scholar] [CrossRef]
- Rufián-Henares, J.A.; Morales, F.J. Microtiter plate-based assay for screening antimicrobial activity of melanoidins against E. coli and S. aureus. Food Chem. 2008, 111, 1069–1074. [Google Scholar] [CrossRef]
- Hwang, I.G.; Kim, H.Y.; Woo, K.S.; Lee, J.; Jeong, H.S. Biological activities of Maillard reaction products (MRPs) in a sugar-amino acid model system. Food Chem. 2011, 126, 221–227. [Google Scholar] [CrossRef]
- Moreno-Montoro, M.; Olalla-Herrera, M.; Gimenez-Martinez, R.; Navarro-Alarcon, M.; Rufian-Henares, J.A. Phenolic compounds and antioxidant activity of Spanish commercial grape juices. J. Food Comp. Anal. 2015, 38, 19–26. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Ferreira, A.C.S.; De Freitas, V.; Silva, A.M.S. Oxidation mechanisms occurring in wines. Food Res. Int. 2011, 44, 1115–1126. [Google Scholar] [CrossRef] [Green Version]
- Sacchetti, G.; Maietti, S.; Muzzoli, M.; Scaglianti, M.; Manfredini, S.; Radice, M.; Bruni, R. Comparative evaluation of 11 essential oils of different origin as functional antioxidants, antiradicals and antimicrobials in foods. Food Chem. 2005, 91, 621–632. [Google Scholar] [CrossRef]
- Orphanides, A.; Goulas, V.; Gekas, V. Effect of drying method on the phenolic content and antioxidant capacity of spearmint. Czech J. Food Sci. 2013, 31, 509–513. [Google Scholar] [CrossRef] [Green Version]
- Je, J.Y.; Kim, S.K. Chitosan derivatives killed bacteria by disrupting the outer and inner membrane. J. Agric. Food Chem. 2006, 54, 6629–6633. [Google Scholar] [CrossRef] [PubMed]
- Mosquito, S.; Ochoa, T.J.; Cok, J.; Cleary, T.G. Effect of bovine lactoferrin in Salmonella ser. Typhimurium infection in mice. Biometals 2010, 23, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Zhang, Y.; Ling, J.; Ma, J.; Huang, L.; Zhang, L. An in vitro study on the effects of nisin on the antibacterial activities of 18 antibiotics against Enterococcus faecalis. PLoS ONE 2014, 9, e89209. [Google Scholar] [CrossRef] [PubMed]
- Galanis, S.A.; Spyroulias, G.A.; Pierattelli, R.; Tzakos, A.; Troganis, A.; Gerothanassis, I.P.; Pairas, G.; Manessi-Zoupa, E.; Cordopatis, P. Zinc binding in peptide models of angiotensin-I converting enzyme active sites studied through H-NMR and chemical shift perturbation mapping. Biopolymers 2003, 69, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Tzakos, A.G.; Naqvi, N.; Comporozos, K.; Pierattelli, R.; Theodorou, V.; Gerothanassis, I.P. The molecular basis for the selection of captopril cis and trans conformations by angiotensin I converting enzyme. Bioorg. Med. Chem. Lett. 2006, 16, 5084–5087. [Google Scholar] [CrossRef] [PubMed]
- Galaris, D.; Barbouti, A.; Korantzopoulos, P. Oxidative stress in hepatic ischemia-reperfusion injury: The role of antioxidants and iron chelating compounds. Curr. Pharm. Des. 2006, 12, 2875–2890. [Google Scholar] [CrossRef] [PubMed]
- Kellici, T.F.; Liapakis, G.; Tzakos, A.; Mavromoustakos, T. Pharmaceutical compositions for antihypertensive treatments: A patent review. Expert Opin. Ther. Pat. 2015, 25, 1305–1317. [Google Scholar] [PubMed]
- Wang, H.Y.; Qian, H.; Yao, W.R. Melanoidins produced by the Maillard reaction: Structure and biological activity. Food Chem. 2011, 128, 573–584. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wines | A345 | A420 | A440 | Sugar Contents (mg GE g−1 m †) | Phenolic Groups Contents (mg GAE g−1 m ‡) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Range | Mean | Range | Mean | Range | Mean | Range | Mean | Range | |
| Aged | 0.441 * | 0.313–0.525 | 0.193 * | 0.132–0.234 | 0.168 * | 0.116–0.208 | 509.8 | 452.2–550.7 | 84.3 | 64.7–99.7 |
| Non-aged | 0.238 * | 0.180–0.311 | 0.099 * | 0.066–0.140 | 0.091 * | 0.061–0.130 | 520.5 | 462.6–612.2 | 114.7 | 84.0–165.2 |
| All | 0.340 | 0.180–0.525 | 0.146 | 0.066–0.234 | 0.130 | 0.061–0.208 | 515.1 | 452.2–612.2 | 99.5 | 64.7–165.2 |
| Microorganism | MIC (mg mL−1) | |
|---|---|---|
| Aged Wines | Non-Aged Wines | |
| Listeria monocytogenes | 5 | 5 |
| Salmonella Enteritidis | 5 | 5 |
| Escherichia coli | 10 | 10 |
| Wines | Mean (%) | Range (%) |
|---|---|---|
| Aged | 58.4 | 54.4–62.1 |
| Non-aged | 74.4 | 70.7–79.8 |
| All | 66.4 | 54.4–79.8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goulas, V.; Nicolaou, D.; Botsaris, G.; Barbouti, A. Straw Wine Melanoidins as Potential Multifunctional Agents: Insight into Antioxidant, Antibacterial, and Angiotensin-I-Converting Enzyme Inhibition Effects. Biomedicines 2018, 6, 83. https://doi.org/10.3390/biomedicines6030083
Goulas V, Nicolaou D, Botsaris G, Barbouti A. Straw Wine Melanoidins as Potential Multifunctional Agents: Insight into Antioxidant, Antibacterial, and Angiotensin-I-Converting Enzyme Inhibition Effects. Biomedicines. 2018; 6(3):83. https://doi.org/10.3390/biomedicines6030083
Chicago/Turabian StyleGoulas, Vlasios, Demetra Nicolaou, George Botsaris, and Alexandra Barbouti. 2018. "Straw Wine Melanoidins as Potential Multifunctional Agents: Insight into Antioxidant, Antibacterial, and Angiotensin-I-Converting Enzyme Inhibition Effects" Biomedicines 6, no. 3: 83. https://doi.org/10.3390/biomedicines6030083