Influence of Iodine Feeding on Microbiological and Physico-Chemical Characteristics and Biogenic Amines Content in a Raw Ewes’ Milk Cheese

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cheese-Making Procedure
2.2. Microbiological Analyses
2.3. Gross Physico-Chemical Composition
2.4. BA Determination
2.5. Statistical Analyses
3. Results
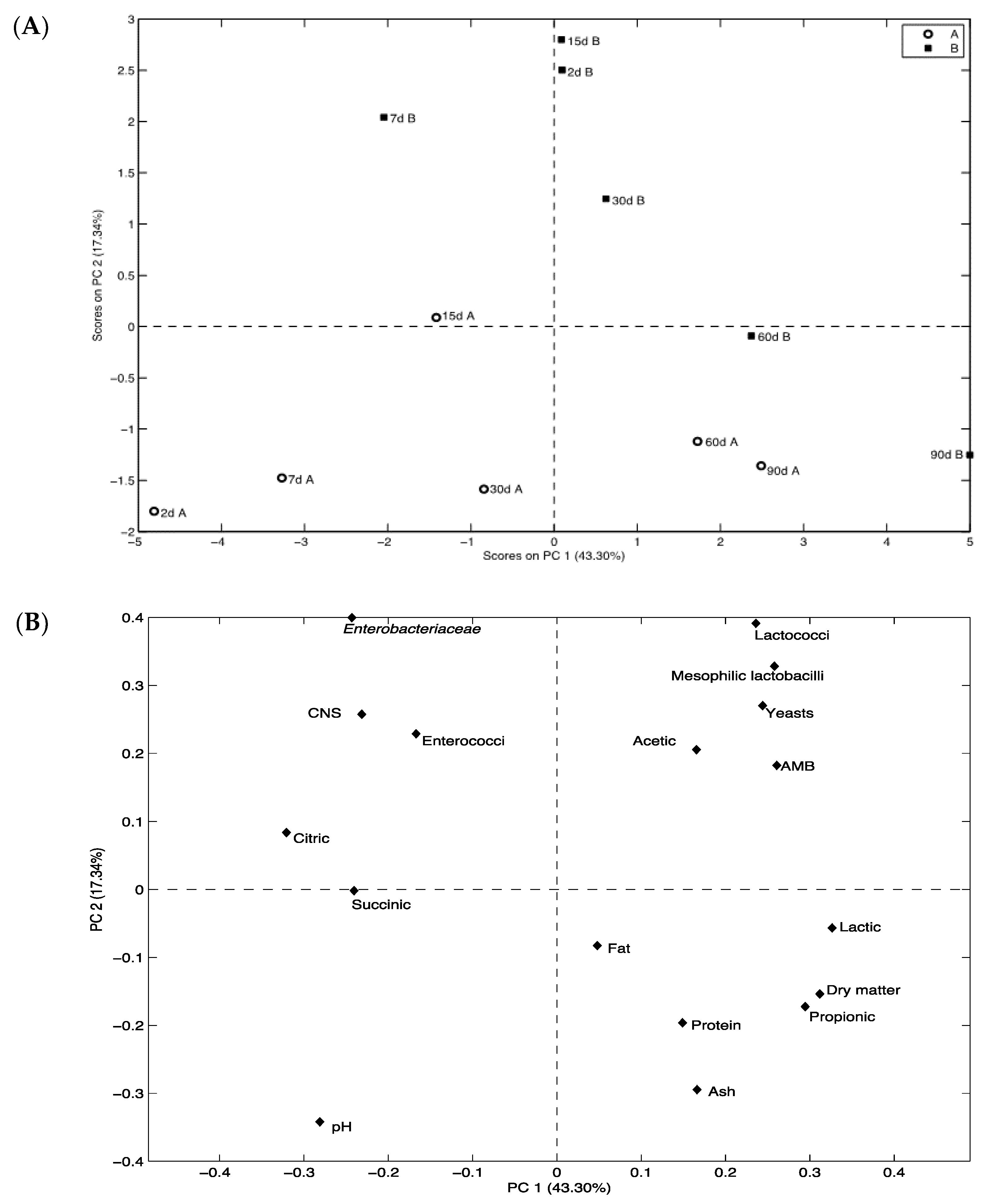
3.1. Microbial Analyses
3.2. Gross Physico-Chemical Composition
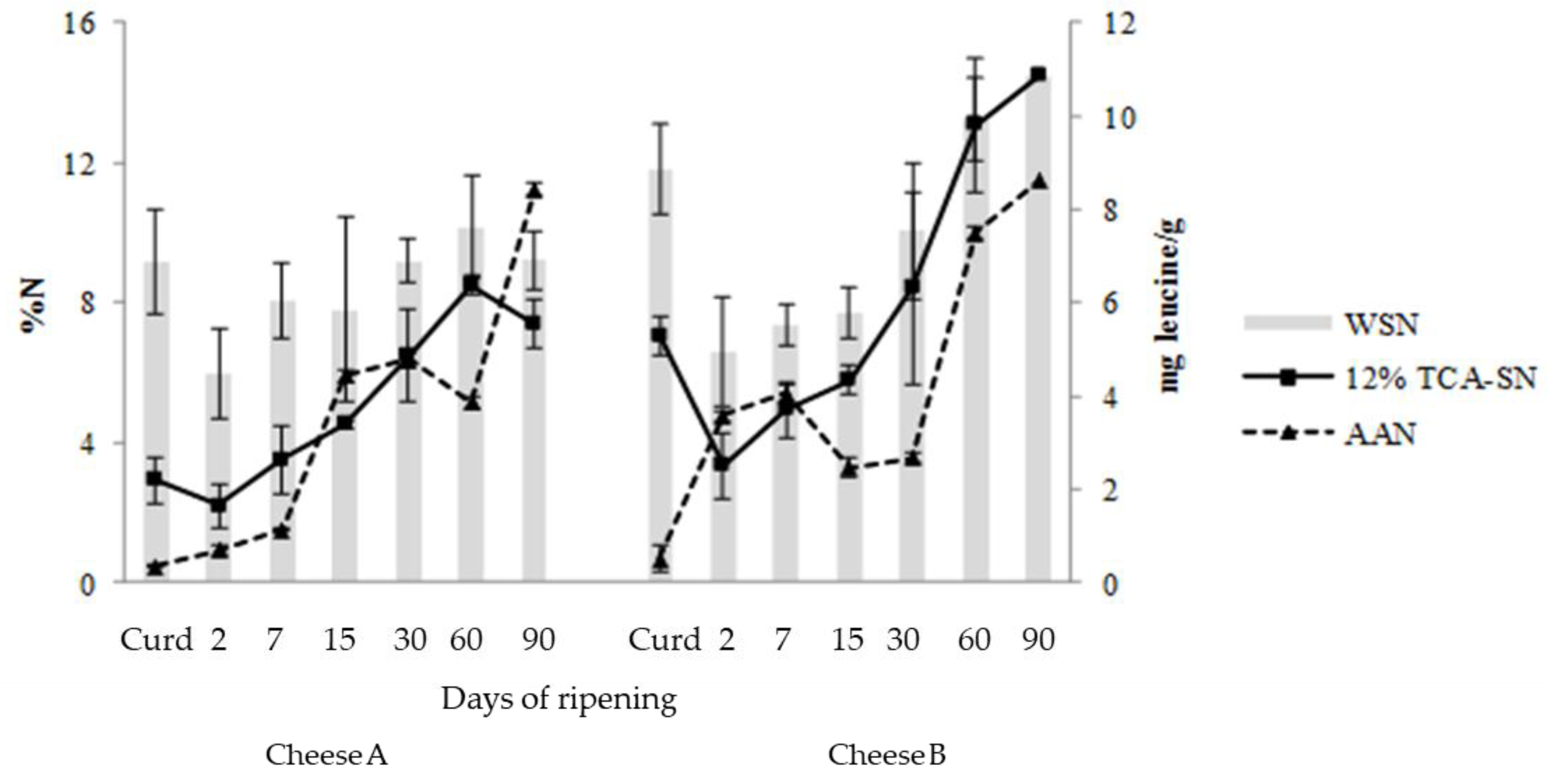
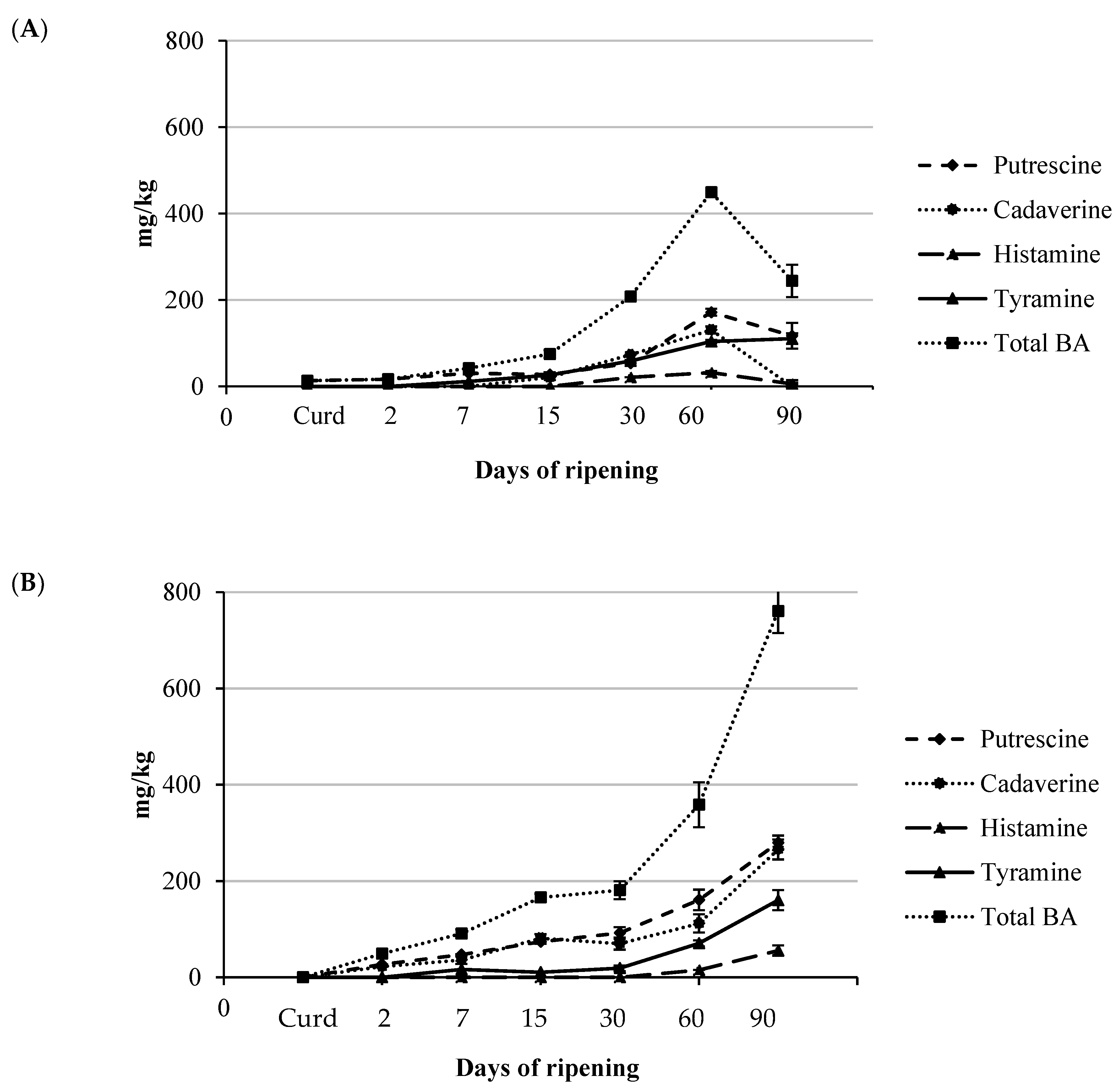
3.3. BA Content
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bader, N.; Möller, U.; Leiterer, M.; Franke, K.; Jahreis, G. Pilot study: Tendency of increasing iodine content in human milk and cow’s milk. Exp. Clin. Endocrinol. Diabetes 2005, 113, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Gaucheron, F. Milk minerals, trace elements, and macroelements. In Milk and Dairy Products in Human Nutrition: Production, Composition and Health; Park, Y.W., Haenlein, G.F.W., Eds.; Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 172–199. ISBN 9781118534168. [Google Scholar]
- Haldimann, M.; Alt, A.; Blank, A.; Blondeau, K. Iodine content of food groups. J. Food Compos. Anal. 2005, 18, 461–471. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Assessment of Iodine Deficiency Disorders and Monitoring Their Elimination, 3rd ed.; World Health Organization (WHO): Geneva, Switzerland, 2007; ISBN 9789241595827. [Google Scholar]
- Knowles, S.O.; Grace, N.D.; Knight, T.W.; McNabba, W.C.; Lee, J. Reasons and means for manipulating the micronutrient composition of milk from grazing dairy cattle. Anim. Feed Sci. Technol. 2006, 131, 154–167. [Google Scholar] [CrossRef]
- Nudda, A.; Battacone, G.; Decandia, M.; Acciaro, M.; Aghini-Lombardi, F.; Frigeri, M.; Pulina, G. The effect of dietary iodine supplementation in dairy goats on milk production traits and milk iodine content. J. Dairy Sci. 2009, 92, 5133–5138. [Google Scholar] [CrossRef] [PubMed]
- Weiss, W.P.; Wyatt, D.J.; Kleinschmit, D.H.; Socha, M.T. Effect of including canola meal and supplemental iodine in diets of dairy cows on short-term changes in iodine concentrations in milk. J. Dairy Sci. 2015, 98, 4841–4849. [Google Scholar] [CrossRef] [PubMed]
- Giello, M.; La Storia, A.; Masucci, F.; Di Francia, A.; Ercolini, D.; Villani, F. Dynamics of bacterial communities during manufacture and ripening of traditional Caciocavallo of Castelfranco cheese in relation to cows’ feeding. Food Microbiol. 2017, 63, 170–177. [Google Scholar] [CrossRef] [PubMed]
- European Commission (EC). No. 1459/2005 of 8 September 2005 amending the conditions for the authorization of a number of feed additives belonging to the group of trace elements. Off. J. Eur. Union 2005, L233, 8–10. [Google Scholar]
- Schirone, M.; Tofalo, R.; Mazzone, G.; Corsetti, A.; Suzzi, G. Biogenic amine content and microbiological profile of Pecorino di Farindola cheese. Food Microbiol. 2011, 28, 128–136. [Google Scholar] [CrossRef] [PubMed]
- International Organization for Standardization (ISO). Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Detection and Enumeration of Escherichia coli O157:H7; ISO 16654; International Organization for Standardization (ISO): Geneva, Switzerland, 2001. [Google Scholar]
- International Organization for Standardization (ISO). Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Detection of Salmonella spp.; ISO 6579; International Organization for Standardization (ISO): Geneva, Switzerland, 2002. [Google Scholar]
- International Organization for Standardization (ISO). Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Detection and Enumeration of Listeria monocytogenes—Part 1: Enumeration Method; ISO 11290-1; International Organization for Standardization (ISO): Geneva, Switzerland, 2005. [Google Scholar]
- Rossi, F.; Torriani, S.; Dellaglio, F. Genus and species-specific PCR-based detection of dairy propionibacteria in environmental samples using primers targeted to the 16S rDNA. Appl. Environ. Microbiol. 1999, 65, 4241–4244. [Google Scholar] [PubMed]
- Tofalo, R.; Schirone, M.; Fasoli, G.; Perpetuini, G.; Patrignani, F.; Manetta, A.C.; Lanciotti, R.; Corsetti, A.; Martino, G.; Suzzi, G. Influence of pig rennet on proteolysis, organic acids content and microbiota of Pecorino di Farindola, a traditional Italian ewe’s raw milk cheese. Food Chem. 2015, 175, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Bouzas, J.; Kantt, C.A.; Bodyfelt, F.; Torres, J.A. Simultaneous determination of sugars and organic acids in Cheddar cheese by high-performance liquid chromatography. J. Food Sci. 1991, 56, 276–278. [Google Scholar] [CrossRef]
- Kuchroo, C.N.; Fox, P.F. Soluble nitrogen in Cheddar cheese: Comparison of extraction procedures. Milchwissenschaft 1982, 37, 331–335. [Google Scholar]
- Polychroniadou, A.; Michaelidou, A.; Paschaloudis, N. Effect of time, temperature and extraction method on the trichloroacetic acid-soluble nitrogen of cheese. Int. Dairy J. 1999, 9, 559–568. [Google Scholar] [CrossRef]
- Folkertsma, B.; Fox, P.F. Use of Cd-ninhydrin reagent to assess proteolysis in cheese during ripening. J. Dairy Res. 1992, 59, 217–224. [Google Scholar] [CrossRef]
- Schirone, M.; Tofalo, R.; Fasoli, G.; Perpetuini, G.; Corsetti, A.; Manetta, A.C.; Ciarrocchi, A.; Suzzi, G. High content of biogenic amines in Pecorino cheeses. Food Microbiol. 2013, 34, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Manzano, M.; Sarais, I.; Stecchini, M.L.; Rondinini, G. Etiology of gas defects in Montasio cheese. Ann. Microbiol. 1990, 40, 255–259. [Google Scholar]
- Centeno, J.A.; Menéndez, S.; Rodriguez-Otero, J.L. Main microbial flora present as natural starters in Cebreiro raw cow’s-milk cheese (Northwest Spain). Int. J. Food Microbiol. 1996, 33, 307–313. [Google Scholar] [CrossRef]
- Quinto, E.; Franco, C.; Rodriguez-Otero, J.L.; Fente, C.; Cepeda, A. Microbiological quality of Cebreiro cheese from Northwest Spain. J. Food Saf. 1994, 14, 1–8. [Google Scholar] [CrossRef]
- Berthier, F.; Beuvier, E.; Dasen, A.; Grappin, R. Origin and diversity of mesophilic lactobacilli in Comté cheese, as revealed by PCR with repetitive and species-specific primers. Int. Dairy J. 2001, 11, 293–305. [Google Scholar] [CrossRef]
- Samelis, J.; Lianou, A.; Kakouri, A.; Delbes, C.; Rogelj, I.; Bogovič-Matijašić, B.O.J.A.N.A.; Montel, M.C. Changes in the microbial composition of raw milk induced by thermization treatments applied prior to traditional Greek hard cheese processing. J. Food Prot. 2009, 72, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Poullet, B.; Huertas, M.; Sánchez, A.; Cáceres, P.; Larriba, G. Main lactic acid bacteria isolated during ripening of Casar de Cáceres cheese. J. Dairy Res. 1993, 60, 123–127. [Google Scholar] [CrossRef]
- Suzzi, G.; Caruso, M.; Gardini, F.; Lombardi, A.; Vannini, L.; Guerzoni, M.E.; Andrighetto, C.; Lanorte, M.T. A survey of enterococci isolated from an artisanal Italian goat’s cheese (semicotto caprino). J. Appl. Microbiol. 2000, 89, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Giraffa, G. Functionality of enterococci in dairy products. Int. J. Food Microbiol. 2003, 88, 215–222. [Google Scholar] [CrossRef]
- Martino, G.P.; Quintana, I.M.; Espariz, M.; Blancato, V.S.; Magni, C. Aroma compounds generation in citrate metabolism of Enterococcus faecium: Genetic characterization of type I citrate gene cluster. Int. J. Food Microbiol. 2016, 218, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Suzzi, G.; Gardini, F. Biogenic amines in dry fermented sausages: A review. Int. J. Food Microbiol. 2003, 88, 41–54. [Google Scholar] [CrossRef]
- Schirone, M.; Tofalo, R.; Visciano, P.; Corsetti, A.; Suzzi, G. Biogenic amines in Italian Pecorino cheese. Front. Microbiol. 2012, 3, 171. [Google Scholar] [CrossRef] [PubMed]
- Tofalo, R.; Fasoli, G.; Schirone, M.; Perpetuini, G.; Pepe, A.; Corsetti, A.; Suzzi, G. The predominance, biodiversity and biotechnological properties of Kluyveromyces marxianus in the production of Pecorino di Farindola cheese. Int. J. Food Microbiol. 2014, 187, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Coloretti, F.; Chiavari, C.; Luise, D.; Tofalo, R.; Fasoli, G.; Suzzi, G.; Grazia, L. Detection and identification of yeasts in natural whey starter for Parmigiano Reggiano cheese-making. Int. Dairy J. 2017, 66, 13–17. [Google Scholar] [CrossRef]
- Moschini, M.; Battaglia, M.; Beone, G.M.; Piva, G.; Masoero, F. Iodine and selenium carry over in milk and cheese in dairy cows: Effect of diet supplementation and milk yield. Animal 2010, 4, 147–155. [Google Scholar] [CrossRef] [PubMed]
- O’Kane, S.M.; Pourshadidi, L.K.; Mulhern, M.S.; Weir, R.R.; Hill, S.; O’Reilly, J.; Kmiotek, D.; Deitrich, C.; Mackle, E.M.; Fitzgerald, E.; et al. The effect of processing and seasonality on the iodine and selenium concentration of cow’s milk produced in Northern Ireland (NI): Implications for population dietary intake. Nutrients 2018, 10, 287. [Google Scholar] [CrossRef] [PubMed]
- Manca, M.G.; Nudda, A.; Rubattu, R.; Boe, R.; Pulina, G. Fatty acid profile of milk fat in goat supplemented with iodized salt. Ital. J. Anim. Sci. 2009, 8 (Suppl. 2), 460. [Google Scholar]
- Secchiari, P.; Mele, M.; Frigeri, M.; Serra, A.; Conte, G.; Pollicardo, A.; Formisano, G.; Alghini-Lombardi, F. Iodine supplementation in dairy ewes nutrition: Effect on milk production traits and milk iodine content. Ital. J. Anim. Sci. 2009, 8 (Suppl. 2), 463. [Google Scholar]
- Fox, P.F.; Law, J.; McSweeney, P.L.H.; Wallace, J. Biochemistry of cheese ripening. In Cheese: Chemistry, Physics and Microbiology. General Aspects, 3rd ed.; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Chapman & Hall: London, UK, 1993; pp. 389–438. ISBN 97800805000942. [Google Scholar]
- Sousa-Gallagher, M.J.; Ardo, Y.; McSweeney, P.L.H. Advances in study of proteolysis during cheese ripening. Int. Dairy J. 2001, 11, 327–345. [Google Scholar] [CrossRef]
- Spano, G.; Russo, P.; Lonvaud-Funel, A.; Lucas, P.; Alexandre, H.; Grandvalet, C.; Coton, E.; Coton, M.; Barnavon, L.; Bach, B.; et al. Biogenic amines in fermented foods. Eur. J. Clin. Nutr. 2010, 64, S95–S100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, M.A.; Moreno-Arribas, M.V. The problem of biogenic amines in fermented foods and the use of potential biogenic amine-degrading microorganisms as a solution. Trends Food Sci. Technol. 2014, 39, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Herrero-Fresno, A.; Martínez, N.; Sánchez-Llana, E.; Díaz, M.; Fernández, M.; Martin, M.C.; Ladero, V.; Alvarez, M.A. Lactobacillus casei strains isolated from cheese reduce biogenic amine accumulation in an experimental model. Int. J. Food Microbiol. 2012, 157, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Ladero, V.; Herrero-Fresno, A.; Martinez, N.; del Río, B.; Linares, D.M.; Fernández, M.; Martin, M.C.; Alvarez, M.A. Genome sequence analysis of the biogenic amine-degrading strain Lactobacillus casei 5b. Genome Announc. 2014, 2, 1–2. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Microbial Groups | Trial | Milk | Curd | 2 Days | 7 Days | 15 Days | 30 Days | 60 Days | 90 Days |
|---|---|---|---|---|---|---|---|---|---|
| Mesophilic lactobacilli | |||||||||
| A | 5.2 (0.02) * | 6.6 (0.006) * | 7.2 (0.01) * | 7.8 (0.002) * | 8.3 (0.02) * | 7.2 (0.009) * | 8.4 (0.006) * | 8.7 (0.001) * | |
| B | 6.8 (0.003) * | 7.9 (0.001) * | 8.6 (0.004) * | 8.5 (0.004) * | 8.8 (0.002) * | 8.5 (0.002) * | 8.1 (0.09) * | 8.7 (0.001) * | |
| Lactococci | |||||||||
| A | 5.4 (0.006) * | 6.1 (0.013) * | 7.5 (0.01) * | 7.6 (0.01) * | 8.1 (0.02) * | 7.9 (0.004) * | 8.7 (0.05) * | 8.5 (0.004) * | |
| B | 6.8 (0.001) * | 8.4 (0.007) * | 8.6 (0.005) * | 8.7 (0.001) * | 8.8 (0.001) * | 8.8 (0.002) * | 8.5 (0.05) * | 8.5 (0.007) * | |
| AMB a | |||||||||
| A | 6.9 (0.001) * | 6.7 (0.001) * | 7.9 (0.001) * | 7.4 (0.007) * | 8.6 (0.004) * | 8.9 (0.001) * | 8.4 (0.001) * | 8.4 (0.006) * | |
| B | 5.5 (0.01) * | 7.0 (0.014) * | 8.7 (0.002) * | 8.8 (0.004) * | 8.9 (0.001) * | 8.9 (0.001) * | 8.9 (0.09) * | 8.7 (0.006) * | |
| Yeasts | |||||||||
| A | - | 4.3 (0.005) * | 3.5 (0.007) * | 4.6 (0.002) * | 4.5 (0.006) * | 4.4 (0.005) * | 4.5 (0.004) | 4.4 (0.007) * | |
| B | - | 3.5 (0.003) * | 4.7 (0.004) * | 4.4 (0.004) * | 5.2 (0.009) * | 5.1 (0.009) * | 5.3 (0.4) | 5.1 (0.003) * | |
| Enterobacteriaceae | |||||||||
| A | 3.4 (0.009) * | 5.4 (0.007) * | 4.7 (0.003) * | 4.3 (0.009) * | 4.5 (0.007) * | 2.6 (0.005) * | 2.4 (0.002) * | 2.4 (0.004) | |
| B | 3.4 (0.001) * | 5.4 (0.005) * | 5.8 (0.002) * | 5.8 (0.001) * | 5.8 (0.001) * | 3.8 (0.002) * | 3.9 (0.003) * | <1 | |
| Enterococci | |||||||||
| A | 2.8 (0.001) * | 4.7 (0.004) * | 6.5 (0.004) * | 5.3 (0.002) * | 4.5 (0.004) * | 4.3 (0.004) * | 5.8 (0.003) * | 5.5 (0.003) * | |
| B | 2.5 (0.006) * | 5.7 (0.002) * | 5.6 (0.005) * | 6.0 (0.001) * | 6.3 (0.002) * | 6.1 (0.001) * | 5.1 (0.07) * | 3.8 (0.001) * | |
| CNS b | |||||||||
| A | 4.9 (0.002) * | 6.4 (0.003) * | 5.4 (0.003) * | 6.5 (0.001) * | 6.5 (0.003) * | 6.7 (0.001) * | 3.1 (0.003) * | 3.4 (0.01) * | |
| B | 6.3 (0.005) * | 6.4 (0.003) * | 7.3 (0.007) * | 6.8 (0.001) * | 5.7 (0.002) * | 5.6 (0.003) * | 5 (1) * | 4.3 (0.01) * | |
| Parameters | Cheese A | Cheese B |
|---|---|---|
| Physico-chemical | ||
| pH | 5.55 ± 0.20 | 5.39 ± 0.10 |
| aw | 0.98 ± 0.01 | 0.97 ± 0.01 |
| % Dry matter | 67.68 ± 4.80 | 66.15 ± 2.23 |
| % Fat 1 | 47.03 ± 3.13 | 51.69 ± 1.73 |
| % Protein 1 | 44.12 ± 6.68 | 40.92 ± 7.54 |
| % Ash 1 | 8.85 ± 0.26 | 7.39 ± 0.47 |
| Organic acids | ||
| Citric acid | 0.5 ± 0.1 | 0.40 ± 0.07 |
| Succinic acid | 0.10 ± 0.05 | 0.21 ± 0.03 |
| Lactic acid | 35 ± 4 | 36 ± 4 |
| Acetic acid | 0.80 ± 0.05 | 0.90 ± 0.06 |
| Propionic acid | 0.04 ± 0.03 | 0.13 ± 0.06 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schirone, M.; Tofalo, R.; Perpetuini, G.; Manetta, A.C.; Di Gianvito, P.; Tittarelli, F.; Battistelli, N.; Corsetti, A.; Suzzi, G.; Martino, G. Influence of Iodine Feeding on Microbiological and Physico-Chemical Characteristics and Biogenic Amines Content in a Raw Ewes’ Milk Cheese. Foods 2018, 7, 108. https://doi.org/10.3390/foods7070108
Schirone M, Tofalo R, Perpetuini G, Manetta AC, Di Gianvito P, Tittarelli F, Battistelli N, Corsetti A, Suzzi G, Martino G. Influence of Iodine Feeding on Microbiological and Physico-Chemical Characteristics and Biogenic Amines Content in a Raw Ewes’ Milk Cheese. Foods. 2018; 7(7):108. https://doi.org/10.3390/foods7070108
Chicago/Turabian StyleSchirone, Maria, Rosanna Tofalo, Giorgia Perpetuini, Anna Chiara Manetta, Paola Di Gianvito, Fabrizia Tittarelli, Noemi Battistelli, Aldo Corsetti, Giovanna Suzzi, and Giuseppe Martino. 2018. "Influence of Iodine Feeding on Microbiological and Physico-Chemical Characteristics and Biogenic Amines Content in a Raw Ewes’ Milk Cheese" Foods 7, no. 7: 108. https://doi.org/10.3390/foods7070108