Zebrafish as a Model for Developmental Neurotoxicity Assessment: The Application of the Zebrafish in Defining the Effects of Arsenic, Methylmercury, or Lead on Early Neurodevelopment


Abstract
:1. Introduction

2. Zebrafish as a Model for DNT
2.1. General Strengths of the Developmental Zebrafish Model System
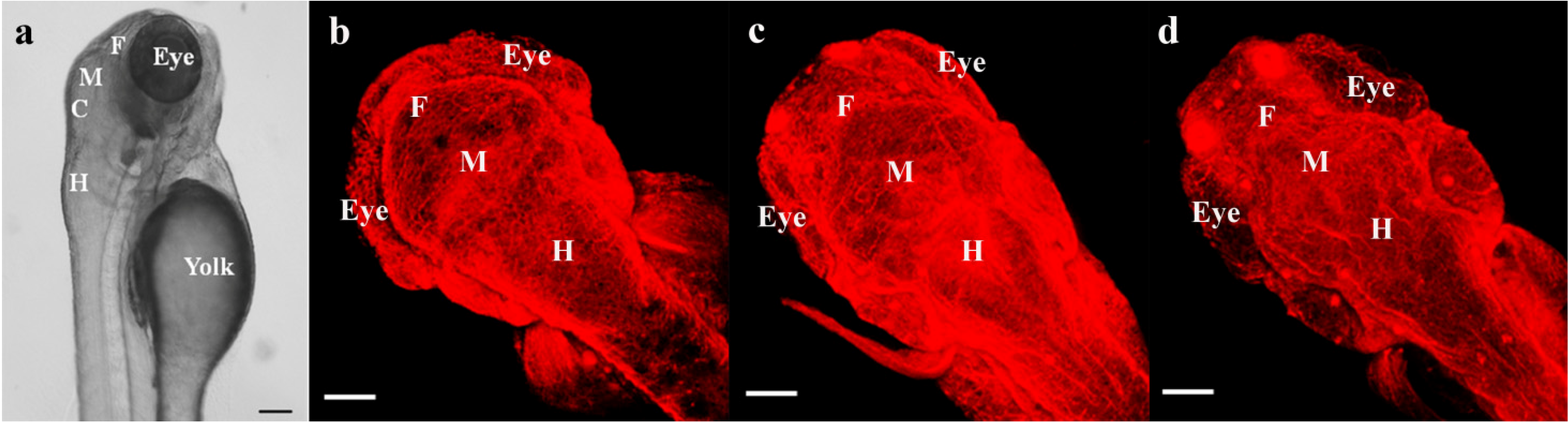
2.2. Comparison of Mammalian and Zebrafish CNS Development
2.3. Blood-Brain Barrier (BBB) in the Zebrafish
3. DNT of Arsenic, Methylmercury, and Lead
3.1. Arsenic

{kind=link}
| Substance | Endpoints | In Vivo Model | Concentration a | Exposure Period b | Key Observations c | References |
|---|---|---|---|---|---|---|
| Arsenic | Axonal/nerve growth | Zebrafish | 2 mM | 4–48 hpf | Altered axon outgrowth in the brain and nerve growth in the spinal cord | Li et al. [120] |
| Behavioral alteration | Rat | 13.6 mg/L (≈0.1 mM) | GD0–PND21 | Delayed behavioral development (reflex responses) | Luo et al. [116] | |
| 10 mg/L (≈0.08 mM) or above | GD6–PND42 | Altered reflex responses or learning/memory behaviors | Xi et al. [118] | |||
| Zebrafish | 2 mM or above | 4–30 hpf | Decrease in reflexive movement frequency under light stimulation | Li et al. [120] | ||
| MeHg | Transcriptomic endpoint | Rat | 0.1 mg/kg (≈0.5 µM) or above | GD6–PND10 | Altered expression of genes related to brain development functional cluster in female offspring brain | Radonjic et al. [121] |
| Mouse | 4 mg/kg (≈0.02 mM) | for 9 weeks including gestation period and 2 weeks post-partum | Altered expression of genes related to functional classes of cell morphology/function, growth factor activity, or receptor binding in pup brain | Jayashankar et al. [122] | ||
| 2.6 mg/kg (≈0.01 mM) | for 8 weeks including gestation period and 2 weeks post-partum | Altered expression of genes enriched in cell proliferation or stress response functions in pup brain | Jayashankar et al. [123] | |||
| 1.5 mg/kg (≈0.007 mM) or above | for 11 weeks including gestation period and 2 weeks post-partum | Exposure to MeHg chloride or MeHg cysteine altered expression of genes involved in functional clusters of immunoglobulin, metal/zinc binding, or methylation in pup brain | Glover et al. [124] | |||
| Zebrafish | 60 µg/L (≈0.3 µM) | 48–72 hpf | Altered expression of clusters of genes involved in apoptosis, oxidative stress response, transcriptional elongation, or DNA repair | Ho et al. [125] | ||
| Behavioral alteration | Rat | 0.5 mg/kg (≈0.002 mM) | GD7–PND21 | Altered vertical activity in 2-month old female, but not in male rats | Cauli et al. [126] | |
| Mouse | 1.5 mg/kg (≈0.007 mM) | for 11 weeks including gestation period and 2 weeks post-partum | Altered open field activity in pups exposed to MeHg chloride, but not MeHg cysteine | Glover et al. [124] | ||
| Pb | Transcriptomic endpoint | Mouse | 0.1 mM | GD8–PND21 | Altered expression of genes related to signal transduction pathway in female pup brain | Kasten-Jolly et al. [127] |
| Zebrafish | 100 ppb (≈0.5 µM) | 2–16 cell stage −72 hpf | Altered global expression of genes related to neurological development, functioning, or diseases | Peterson et al. [128] | ||
| Axonal/nerve growth | Zebrafish | 100 ppb (≈0.5 µM) | ≈2–≈36 hpf | Decreased density of axon tracts | Zhang et al. [70] | |
| Behavioral alteration | Rat | 5 mg/L (≈0.02 mM) | GD0–60 days of age in offspring | Increased locomotor activity | Luo et al. [129] | |
| 2.84 mg/mL (≈14 mM) | GD1–PND24 | Maternal Pb exposure did not induce anxiety-related behavioral change in pups | Molina et al. [130] | |||
| Zebrafish | 10 nM or above | <2–24 hpf | Altered startling behavior in response to tapping stimulation | Rice et al. [131] | ||
| 0.1 mg/L (≈0.5 µM) or above | ≈6–8 to 20–30 hpf | Altered spontaneous movement | Chen et al. [132] | |||
| 0.025 mg/L (≈0.1 µM) or above | 6–96 hpf | Altered swimming activity in response to light stimulation | ||||
| 6–120 hpf | Altered swimming activity under light or dark condition | |||||
| 0.2 mM | 0–144 hpf | Altered spontaneous swimming activity | Dou and Zhang [133] |
3.2. Methylmercury
3.3. Lead
3.4. Mixtures
4. DNT Studies Using the Zebrafish Model System
4.1. Application of the Zebrafish Model in Various Chemical-Induced DNT Studies
4.2. The Zebrafish as a Potential Tool for Chemical-Induced DNT Screening
4.3. DNT Studies of Arsenic, Methylmercury, or Lead Using the Zebrafish
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bellinger, D.C. Prenatal Exposures to Environmental Chemicals and Children’s Neurodevelopment: An Update. Saf. Health Work 2013, 4, 1–11. [Google Scholar] [CrossRef]
- Braun, J.M.; Kahn, R.S.; Froehlich, T.; Auinger, P.; Lanphear, B.P. Exposures to environmental toxicants and attention deficit hyperactivity disorder in U.S. children. Environ. Health Perspect. 2006, 114, 1904–1909. [Google Scholar]
- Canfield, R.L.; Henderson, C.R., Jr.; Cory-Slechta, D.A.; Cox, C.; Jusko, T.A.; Lanphear, B.P. Intellectual impairment in children with blood lead concentrations below 10 microg per deciliter. N. Engl. J. Med. 2003, 348, 1517–1526. [Google Scholar]
- Harrington, R.A.; Lee, L.C.; Crum, R.M.; Zimmerman, A.W.; Hertz-Picciotto, I. Prenatal SSRI Use and Offspring with Autism Spectrum Disorder or Developmental Delay. Pediatrics 2014. [Google Scholar] [CrossRef]
- Dobbing, J. Vulnerable Periods in Developing Brain. In Brain, Behaviour, and Iron in the Infant Diet; Dobbing, J., Ed.; Springer: London, UK, 1990; pp. 1–17. [Google Scholar]
- Giussani, D.A. The vulnerable developing brain. Proc. Natl. Acad. Sci. USA 2011, 108, 2641–2642. [Google Scholar] [CrossRef]
- Perera, F.; Herbstman, J. Prenatal environmental exposures, epigenetics, and disease. Reprod. Toxicol. 2011, 31, 363–373. [Google Scholar] [CrossRef]
- Smart, J.; Dobbing, J.; Adlard, B.; Lynch, A.; Sands, J. Vulnerability of developing brain: Relative effects of growth restriction during the fetal and suckling periods on behavior and brain composition of adult rats. J. Nutr. 1973, 103, 1327–1338. [Google Scholar]
- Fagiolini, M.; Jensen, C.L.; Champagne, F.A. Epigenetic influences on brain development and plasticity. Curr. Opin. Neurobiol. 2009, 19, 207–212. [Google Scholar] [CrossRef]
- Bouayed, J.; Desor, F.; Rammal, H.; Kiemer, A.K.; Tybl, E.; Schroeder, H.; Rychen, G.; Soulimani, R. Effects of lactational exposure to benzo[alpha]pyrene (B[alpha]P) on postnatal neurodevelopment, neuronal receptor gene expression and behaviour in mice. Toxicology 2009, 259, 97–106. [Google Scholar] [CrossRef]
- Kleiber, M.L.; Mantha, K.; Stringer, R.L.; Singh, S.M. Neurodevelopmental alcohol exposure elicits long-term changes to gene expression that alter distinct molecular pathways dependent on timing of exposure. J. Neurodev. Disord. 2013, 5, 6. [Google Scholar] [CrossRef]
- Billstedt, E.; Gillberg, I.C.; Gillberg, C. Autism in adults: Symptom patterns and early childhood predictors. Use of the DISCO in a community sample followed from childhood. J. Child Psychol. Psychiatry 2007, 48, 1102–1110. [Google Scholar] [CrossRef]
- Kessler, R.C.; Adler, L.A.; Barkley, R.; Biederman, J.; Conners, C.K.; Faraone, S.V.; Greenhill, L.L.; Jaeger, S.; Secnik, K.; Spencer, T.; et al. Patterns and predictors of attention-deficit/hyperactivity disorder persistence into adulthood: Results from the national comorbidity survey replication. Biol. Psychiatry 2005, 57, 1442–1451. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P.J. Developmental neurotoxicity of industrial chemicals. Lancet 2006, 368, 2167–2178. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P.J. Neurobehavioural effects of developmental toxicity. Lancet Neurol. 2014, 13, 330–338. [Google Scholar] [CrossRef]
- Chen, A.; Dietrich, K.N.; Huo, X.; Ho, S.M. Developmental neurotoxicants in e-waste: An emerging health concern. Environ. Health Perspect. 2011, 119, 431–438. [Google Scholar]
- Mergler, D.; Anderson, H.A.; Chan, L.H.; Mahaffey, K.R.; Murray, M.; Sakamoto, M.; Stern, A.H. Panel on health risks and toxicological effects of methylmercury. Methylmercury exposure and health effects in humans: A worldwide concern. Ambio 2007, 36, 3–11. [Google Scholar] [CrossRef]
- Mukherjee, A.; Sengupta, M.K.; Hossain, M.A.; Ahamed, S.; Das, B.; Nayak, B.; Lodh, D.; Rahman, M.M.; Chakraborti, D. Arsenic contamination in groundwater: A global perspective with emphasis on the Asian scenario. J. Health Popul. Nutr. 2006, 24, 142–163. [Google Scholar]
- Smith, A.H.; Lingas, E.O.; Rahman, M. Contamination of drinking-water by arsenic in Bangladesh: A public health emergency. Bull. World Health Organ. 2000, 78, 1093–1103. [Google Scholar]
- Tong, S.; von Schirnding, Y.E.; Prapamontol, T. Environmental lead exposure: A public health problem of global dimensions. Bull. World Health Organ. 2000, 78, 1068–1077. [Google Scholar]
- Lidsky, T.I.; Schneider, J.S. Lead neurotoxicity in children: Basic mechanisms and clinical correlates. Brain 2003, 126, 5–19. [Google Scholar] [CrossRef]
- Landrigan, P.J. What causes autism? Exploring the environmental contribution. Curr. Opin. Pediatr. 2010, 22, 219–225. [Google Scholar] [CrossRef]
- Roy, A.; Kordas, K.; Lopez, P.; Rosado, J.L.; Cebrian, M.E.; Vargas, G.G.; Ronquillo, D.; Stoltzfus, R.J. Association between arsenic exposure and behavior among first-graders from Torreon, Mexico. Environ. Res. 2011, 111, 670–676. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar]
- Blader, P.; Strähle, U. Zebrafish developmental genetics and central nervous system development. Hum. Mol. Genet. 2000, 9, 945–951. [Google Scholar] [CrossRef]
- Schmidt, R.; Strähle, U.; Scholpp, S. Neurogenesis in zebrafish—From embryo to adult. Neural Dev. 2013. [Google Scholar] [CrossRef]
- Jeong, J.Y.; Kwon, H.B.; Ahn, J.C.; Kang, D.; Kwon, S.H.; Park, J.A.; Kim, K.W. Functional and developmental analysis of the blood-brain barrier in zebrafish. Brain Res. Bull. 2008, 75, 619–628. [Google Scholar] [CrossRef]
- Goldman, D.; Hankin, M.; Li, Z.; Dai, X.; Ding, J. Transgenic zebrafish for studying nervous system development and regeneration. Transgenic Res. 2001, 10, 21–33. [Google Scholar]
- Umans, R.A.; Taylor, M.R. Zebrafish as a model to study drug transporters at the blood-brain barrier. Clin. Pharmacol. Ther. 2012, 92, 567–570. [Google Scholar] [CrossRef]
- Woo, K.; Fraser, S.E. Order and coherence in the fate map of the zebrafish nervous system. Development 1995, 121, 2595–2609. [Google Scholar]
- Dawid, I.B. Developmental biology of zebrafish. Ann. N. Y. Acad. Sci. 2004, 1038, 88–93. [Google Scholar]
- Halpern, M.E.; Thisse, C.; Ho, R.K.; Thisse, B.; Riggleman, B.; Trevarrow, B.; Weinberg, E.S.; Postlethwait, J.H.; Kimmel, C.B. Cell-autonomous shift from axial to paraxial mesodermal development in zebrafish floating head mutants. Development 1995, 4257–4264. [Google Scholar]
- Hanneman, E.; Trevarrow, B.; Metcalfe, W.K.; Kimmel, C.B.; Westerfield, M. Segmental pattern of development of the hindbrain and spinal cord of the zebrafish embryo. Development 1988, 103, 49–58. [Google Scholar]
- Vesterlund, L.; Jiao, H.; Unneberg, P.; Hovatta, O.; Kere, J. The zebrafish transcriptome during early development. BMC Dev. Biol. 2011, 11. [Google Scholar] [CrossRef]
- Weis, J.S. Analysis of the development of the nervous system of the zebrafish, Brachydanio rerio. II. The effect of nerve growth factor and its antiserum on the nervous system of the zebrafish. J. Embryol. Exp. Morphol. 1968, 19, 121–135. [Google Scholar]
- Levin, E.D.; Sledge, D.; Roach, S.; Petro, A.; Donerly, S.; Linney, E. Persistent behavioral impairment caused by embryonic methylphenidate exposure in zebrafish. Neurotoxicol. Teratol. 2011, 33, 668–673. [Google Scholar] [CrossRef]
- Bretaud, S.; Lee, S.; Guo, S. Sensitivity of zebrafish to environmental toxics implicated in Parkinson’s disease. Neurotoxicol. Teratol. 2004, 26, 857–864. [Google Scholar] [CrossRef]
- Carvan, M.J., 3rd; Loucks, E.; Weber, D.N.; Williams, F.E. Ethanol effects on the developing zebrafish: Neurobehavior and skeletal morphogenesis. Neurotoxicol. Teratol. 2004, 26, 757–768. [Google Scholar] [CrossRef]
- Chen, L.; Huang, C.; Hu, C.; Yu, K.; Yang, L.; Zhou, B. Acute exposure to DE-71: Effects on locomotor behavior and developmental neurotoxicity in zebrafish larvae. Environ. Toxicol. Chem. SETAC 2012, 31, 2338–2344. [Google Scholar]
- Chen, L.; Yu, K.; Huang, C.; Yu, L.; Zhu, B.; Lam, P.K.; Lam, J.C.; Zhou, B. Prenatal transfer of polybrominated diphenyl ethers (PBDEs) results in developmental neurotoxicity in zebrafish larvae. Environ. Sci. Technol. 2012, 46, 9727–9734. [Google Scholar] [CrossRef]
- Chen, X.; Huang, C.; Wang, X.; Chen, J.; Bai, C.; Chen, Y.; Chen, X.; Dong, Q.; Yang, D. BDE-47 disrupts axonal growth and motor behavior in developing zebrafish. Aquat. Toxicol. 2012, 120–121, 35–44. [Google Scholar]
- Cowden, J.; Padnos, B.; Hunter, D.; MacPhail, R.; Jensen, K.; Padilla, S. Developmental exposure to valproate and ethanol alters locomotor activity and retino-tectal projection area in zebrafish embryos. Reprod. Toxicol. 2012, 33, 165–173. [Google Scholar] [CrossRef]
- Chen, Q.; Huang, N.N.; Huang, J.T.; Chen, S.; Fan, J.; Li, C.; Xie, F.K. Sodium benzoate exposure downregulates the expression of tyrosine hydroxylase and dopamine transporter in dopaminergic neurons in developing zebrafish. Birth Defects Res. B Dev. Reprod. Toxicol. 2009, 86, 85–91. [Google Scholar] [CrossRef]
- Chow, E.S.; Hui, M.N.; Lin, C.C.; Cheng, S.H. Cadmium inhibits neurogenesis in zebrafish embryonic brain development. Aquat. Toxicol. 2008, 87, 157–169. [Google Scholar] [CrossRef]
- DeMicco, A.; Cooper, K.R.; Richardson, J.R.; White, L.A. Developmental neurotoxicity of pyrethroid insecticides in zebrafish embryos. Toxicol. Sci. 2010, 113, 177–186. [Google Scholar] [CrossRef]
- Eddins, D.; Cerutti, D.; Williams, P.; Linney, E.; Levin, E.D. Zebrafish provide a sensitive model of persisting neurobehavioral effects of developmental chlorpyrifos exposure: Comparison with nicotine and pilocarpine effects and relationship to dopamine deficits. Neurotoxicol. Teratol. 2010, 32, 99–108. [Google Scholar] [CrossRef]
- Fan, C.Y.; Cowden, J.; Simmons, S.O.; Padilla, S.; Ramabhadran, R. Gene expression changes in developing zebrafish as potential markers for rapid developmental neurotoxicity screening. Neurotoxicol. Teratol. 2010, 32, 91–98. [Google Scholar] [CrossRef]
- Flentke, G.R.; Klingler, R.H.; Tanguay, R.L.; Carvan, M.J., 3rd; Smith, S.M. An evolutionarily conserved mechanism of calcium-dependent neurotoxicity in a zebrafish model of fetal alcohol spectrum disorders. Alcohol. Clin. Exp. Res. 2014, 38, 1255–1265. [Google Scholar] [CrossRef]
- Gu, A.; Shi, X.; Yuan, C.; Ji, G.; Zhou, Y.; Long, Y.; Song, L.; Wang, S.; Wang, X. Exposure to fenvalerate causes brain impairment during zebrafish development. Toxicol. Lett. 2010, 197, 188–192. [Google Scholar] [CrossRef]
- Hill, A.; Howard, C.V.; Strähle, U.; Cossins, A. Neurodevelopmental defects in zebrafish (Danio rerio) at environmentally relevant dioxin (TCDD) concentrations. Toxicol. Sci. 2003, 76, 392–399. [Google Scholar] [CrossRef]
- Loucks, E.; Carvan, M.J., 3rd. Strain-dependent effects of developmental ethanol exposure in zebrafish. Neurotoxicol. Teratol. 2004, 26, 745–755. [Google Scholar] [CrossRef]
- McClain, V.; Stapleton, H.M.; Tilton, F.; Gallagher, E.P. BDE 49 and developmental toxicity in zebrafish. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2012, 155, 253–258. [Google Scholar] [CrossRef]
- Muth-Köhne, E.; Wichmann, A.; Delov, V.; Fenske, M. The classification of motor neuron defects in the zebrafish embryo toxicity test (ZFET) as an animal alternative approach to assess developmental neurotoxicity. Neurotoxicol. Teratol. 2012, 34, 413–424. [Google Scholar] [CrossRef]
- Powers, C.M.; Slotkin, T.A.; Seidler, F.J.; Badireddy, A.R.; Padilla, S. Silver nanoparticles alter zebrafish development and larval behavior: Distinct roles for particle size, coating and composition. Neurotoxicol. Teratol. 2011, 33, 708–714. [Google Scholar] [CrossRef]
- Raftery, T.D.; Isales, G.M.; Yozzo, K.L.; Volz, D.C. High-content screening assay for identification of chemicals impacting spontaneous activity in zebrafish embryos. Environ. Sci. Technol. 2014, 48, 804–810. [Google Scholar] [CrossRef]
- Selderslaghs, I.W.; Hooyberghs, J.; Blust, R.; Witters, H.E. Assessment of the developmental neurotoxicity of compounds by measuring locomotor activity in zebrafish embryos and larvae. Neurotoxicol. Teratol. 2013, 37, 44–56. [Google Scholar] [CrossRef]
- Selderslaghs, I.W.; Hooyberghs, J.; de Coen, W.; Witters, H.E. Locomotor activity in zebrafish embryos: A new method to assess developmental neurotoxicity. Neurotoxicol. Teratol. 2010, 32, 460–471. [Google Scholar] [CrossRef]
- Shi, X.; Gu, A.; Ji, G.; Li, Y.; Di, J.; Jin, J.; Hu, F.; Long, Y.; Xia, Y.; Lu, C.; et al. Developmental toxicity of cypermethrin in embryo-larval stages of zebrafish. Chemosphere 2011, 85, 1010–1016. [Google Scholar] [CrossRef]
- Stehr, C.M.; Linbo, T.L.; Incardona, J.P.; Scholz, N.L. The developmental neurotoxicity of fipronil: Notochord degeneration and locomotor defects in zebrafish embryos and larvae. Toxicol. Sci. 2006, 92, 270–278. [Google Scholar] [CrossRef]
- Sun, Y.; Dong, Z.; Khodabakhsh, H.; Chatterjee, S.; Guo, S. Zebrafish chemical screening reveals the impairment of dopaminergic neuronal survival by cardiac glycosides. PLoS One 2012, 7, e35645. [Google Scholar]
- Ton, C.; Lin, Y.; Willett, C. Zebrafish as a model for developmental neurotoxicity testing. Birth Defects Res. A Clin. Mol. Teratol. 2006, 76, 553–567. [Google Scholar]
- Wen, L.; Wei, W.; Gu, W.; Huang, P.; Ren, X.; Zhang, Z.; Zhu, Z.; Lin, S.; Zhang, B. Visualization of monoaminergic neurons and neurotoxicity of MPTP in live transgenic zebrafish. Dev. Biol. 2008, 314, 84–92. [Google Scholar] [CrossRef]
- Xue, J.Y.; Li, X.; Sun, M.Z.; Wang, Y.P.; Wu, M.; Zhang, C.Y.; Wang, Y.N.; Liu, B.; Zhang, Y.S.; Zhao, X.; et al. An assessment of the impact of SiO2 nanoparticles of different sizes on the rest/wake behavior and the developmental profile of zebrafish larvae. Small 2013, 9, 3161–3168. [Google Scholar] [CrossRef]
- Yang, D.; Lauridsen, H.; Buels, K.; Chi, L.H.; la Du, J.; Bruun, D.A.; Olson, J.R.; Tanguay, R.L.; Lein, P.J. Chlorpyrifos-oxon disrupts zebrafish axonal growth and motor behavior. Toxicol. Sci. 2011, 121, 146–159. [Google Scholar] [CrossRef]
- Yen, J.; Donerly, S.; Levin, E.D.; Linney, E.A. Differential acetylcholinesterase inhibition of chlorpyrifos, diazinon and parathion in larval zebrafish. Neurotoxicol. Teratol. 2011, 33, 735–741. [Google Scholar] [CrossRef]
- Zellner, D.; Padnos, B.; Hunter, D.L.; MacPhail, R.C.; Padilla, S. Rearing conditions differentially affect the locomotor behavior of larval zebrafish, but not their response to valproate-induced developmental neurotoxicity. Neurotoxicol. Teratol. 2011, 33, 674–679. [Google Scholar] [CrossRef]
- Smith, L.E.; Carvan, M.J., 3rd; Dellinger, J.A.; Ghorai, J.K.; White, D.B.; Williams, F.E.; Weber, D.N. Developmental selenomethionine and methylmercury exposures affect zebrafish learning. Neurotoxicol. Teratol. 2010, 32, 246–255. [Google Scholar] [CrossRef]
- Weber, D.N.; Connaughton, V.P.; Dellinger, J.A.; Klemer, D.; Udvadia, A.; Carvan, M.J., 3rd. Selenomethionine reduces visual deficits due to developmental methylmercury exposures. Physiol. Behav. 2008, 93, 250–260. [Google Scholar] [CrossRef]
- Wirbisky, S.E.; Weber, G.J.; Lee, J.W.; Cannon, J.R.; Freeman, J.L. Novel dose-dependent alterations in excitatory GABA during embryonic development associated with lead (Pb) neurotoxicity. Toxicol. Lett. 2014, 229, 1–8. [Google Scholar] [CrossRef]
- Zhang, J.; Peterson, S.M.; Weber, G.J.; Zhu, X.; Zheng, W.; Freeman, J.L. Decreased axonal density and altered expression profiles of axonal guidance genes underlying lead (Pb) neurodevelopmental toxicity at early embryonic stages in the zebrafish. Neurotoxicol. Teratol. 2011, 33, 715–320. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Papan, C.; Campos-Ortega, J.A. Region-specific cell clones in the developing spinal cord of the zebrafish. Dev. Genes Evol. 1999, 209, 135–144. [Google Scholar] [CrossRef]
- Papan, C.; Campos-Ortega, J.A. On the formation of the neural keel and neural tube in the zebrafishDanio (Brachydanio) rerio. Roux’s Arch. Dev. Biol. 1994, 203, 178–186. [Google Scholar] [CrossRef]
- Kimmel, C.B. Patterning the brain of the zebrafish embryo. Annu. Rev. Neurosci. 1993, 16, 707–732. [Google Scholar] [CrossRef]
- Ross, L.S.; Parrett, T.; Easter, S.S., Jr. Axonogenesis and morphogenesis in the embryonic zebrafish brain. J. Neurosci. 1992, 12, 467–482. [Google Scholar]
- Wilson, S.W.; Ross, L.S.; Parrett, T.; Easter, S.S., Jr. The development of a simple scaffold of axon tracts in the brain of the embryonic zebrafish, Brachydanio rerio. Development 1990, 108, 121–145. [Google Scholar]
- Myers, P.Z.; Eisen, J.S.; Westerfield, M. Development and axonal outgrowth of identified motoneurons in the zebrafish. J. Neurosci. 1986, 6, 2278–2289. [Google Scholar]
- Pike, S.H.; Melancon, E.F.; Eisen, J.S. Pathfinding by zebrafish motoneurons in the absence of normal pioneer axons. Development 1992, 114, 825–831. [Google Scholar]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar]
- Abbott, N.J.; Ronnback, L.; Hansson, E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef]
- Bernacki, J.; Dobrowolska, A.; Nierwinska, K.; Malecki, A. Physiology and pharmacological role of the blood-brain barrier. Pharmacol. Rep. 2008, 60, 600–622. [Google Scholar]
- Lawther, B.K.; Sajith, K.; Hari, K. Blood–brain barrier. Contin. Educ. Anaesth. Crit. Care Pain 2011, 11, 128–132. [Google Scholar]
- Banks, W.A. Characteristics of compounds that cross the blood-brain barrier. BMC Neurol. 2009. [Google Scholar] [CrossRef]
- Oldendorf, W.H. Lipid solubility and drug penetration of the blood brain barrier. Proc. Soc. Exp. Biol. Med. 1974, 147, 813–815. [Google Scholar] [CrossRef]
- Adinolfi, M. The development of the human blood-CSF-brain barrier. Dev. Med. Child Neurol. 1985, 27, 532–537. [Google Scholar] [CrossRef]
- Johnson, M.H. Functional brain development in humans. Nat. Rev. Neurosci. 2001, 2, 475–483. [Google Scholar]
- Rodier, P.M. Developing brain as a target of toxicity. Environ. Health Perspect. 1995, 103 (Suppl. 6), 73–76. [Google Scholar] [CrossRef]
- Ek, C.J.; Dziegielewska, K.M.; Habgood, M.D.; Saunders, N.R. Barriers in the developing brain and neurotoxicology. Neurotoxicology 2012, 33, 586–604. [Google Scholar] [CrossRef]
- Luissint, A.C.; Artus, C.; Glacial, F.; Ganeshamoorthy, K.; Couraud, P.O. Tight junctions at the blood brain barrier: Physiological architecture and disease-associated dysregulation. Fluids Barriers CNS 2012, 9, 23. [Google Scholar]
- Tam, S.J.; Watts, R.J. Connecting vascular and nervous system development: Angiogenesis and the blood-brain barrier. Annu. Rev. Neurosci. 2010, 33, 379–408. [Google Scholar] [CrossRef]
- Hawkins, B.T.; Davis, T.P. The blood-brain barrier/neurovascular unit in health and disease. Pharmacol. Rev. 2005, 57, 173–185. [Google Scholar] [CrossRef]
- Morita, K.; Sasaki, H.; Furuse, M.; Tsukita, S. Endothelial claudin: Claudin-5/TMVCF constitutes tight junction strands in endothelial cells. J. Cell Biol. 1999, 147, 185–194. [Google Scholar] [CrossRef]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; Furuse, M.; Tsukita, S. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef]
- Fanning, A.S.; Jameson, B.J.; Jesaitis, L.A.; Anderson, J.M. The tight junction protein ZO-1 establishes a link between the transmembrane protein occludin and the actin cytoskeleton. J. Biol. Chem. 1998, 273, 29745–29753. [Google Scholar]
- Furuse, M.; Itoh, M.; Hirase, T.; Nagafuchi, A.; Yonemura, S.; Tsukita, S.; Tsukita, S. Direct association of occludin with ZO-1 and its possible involvement in the localization of occludin at tight junctions. J. Cell Biol. 1994, 127, 1617–1626. [Google Scholar] [CrossRef]
- Stevenson, B.R.; Siliciano, J.D.; Mooseker, M.S.; Goodenough, D.A. Identification of ZO-1: A high molecular weight polypeptide associated with the tight junction (zonula occludens) in a variety of epithelia. J. Cell Biol. 1986, 103, 755–766. [Google Scholar] [CrossRef]
- Abdelilah-Seyfried, S. Claudin-5a in developing zebrafish brain barriers: Another brick in the wall. Bioessays 2010, 32, 768–776. [Google Scholar] [CrossRef]
- Kiener, T.K.; Sleptsova-Friedrich, I.; Hunziker, W. Identification, tissue distribution and developmental expression of tjp1/zo-1, tjp2/zo-2 and tjp3/zo-3 in the zebrafish, Danio rerio. Gene Expr. Patterns 2007, 7, 767–776. [Google Scholar] [CrossRef]
- Xie, J.; Farage, E.; Sugimoto, M.; Anand-Apte, B. A novel transgenic zebrafish model for blood-brain and blood-retinal barrier development. BMC Dev. Biol. 2010, 10, 76. [Google Scholar]
- ZFIN Staff. Curation of Orthology Data. Manually Curated Data. 2003. Available online: http://zfin.org/ (accessed on 1 April 2014).
- Zhang, J.; Piontek, J.; Wolburg, H.; Piehl, C.; Liss, M.; Otten, C.; Christ, A.; Willnow, T.E.; Blasig, I.E.; Abdelilah-Seyfried, S. Establishment of a neuroepithelial barrier by Claudin5a is essential for zebrafish brain ventricular lumen expansion. Proc. Natl. Acad. Sci. USA 2010, 107, 1425–1430. [Google Scholar] [CrossRef]
- Fleming, A.; Diekmann, H.; Goldsmith, P. Functional characterisation of the maturation of the blood-brain barrier in larval zebrafish. PLoS One 2013, 8, e77548. [Google Scholar]
- Hughes, M.F.; Beck, B.D.; Chen, Y.; Lewis, A.S.; Thomas, D.J. Arsenic exposure and toxicology: A historical perspective. Toxicol. Sci. 2011, 123, 305–332. [Google Scholar] [CrossRef]
- Sun, G. Arsenic contamination and arsenicosis in China. Toxicol. Appl. Pharmacol. 2004, 198, 268–271. [Google Scholar] [CrossRef]
- Wang, S.; Mulligan, C.N. Occurrence of arsenic contamination in Canada: Sources, behavior and distribution. Sci. Total Environ. 2006, 366, 701–721. [Google Scholar] [CrossRef]
- Rodriguez-Lado, L.; Sun, G.; Berg, M.; Zhang, Q.; Xue, H.; Zheng, Q.; Johnson, C.A. Groundwater arsenic contamination throughout China. Science 2013, 341, 866–868. [Google Scholar]
- Nordstrom, D.K. Worldwide occurrences of arsenic in ground water. Science 2002, 296, 2143–2145. [Google Scholar] [CrossRef]
- Balazs, C.L.; Morello-Frosch, R.; Hubbard, A.E.; Ray, I. Environmental justice implications of arsenic contamination in California’s San Joaquin Valley: A cross-sectional, cluster-design examining exposure and compliance in community drinking water systems. Environ. Health 2012, 11, 84. [Google Scholar]
- Nahar, M.N.; Inaoka, T.; Fujimura, M. A consecutive study on arsenic exposure and intelligence quotient (IQ) of children in Bangladesh. Environ. Health Prev. Med. 2014, 19, 194–199. [Google Scholar]
- Rosado, J.L.; Ronquillo, D.; Kordas, K.; Rojas, O.; Alatorre, J.; Lopez, P.; Garcia-Vargas, G.; del Carmen Caamano, M.; Cebrian, M.E.; Stoltzfus, R.J. Arsenic exposure and cognitive performance in Mexican schoolchildren. Environ. Health Perspect. 2007, 115, 1371–1375. [Google Scholar] [CrossRef]
- Wang, S.X.; Wang, Z.H.; Cheng, X.T.; Li, J.; Sang, Z.P.; Zhang, X.D.; Han, L.L.; Qiao, X.Y.; Wu, Z.M.; Wang, Z.Q. Arsenic and fluoride exposure in drinking water: Children’s IQ and growth in Shanyin county, Shanxi province, China. Environ. Health Perspect. 2007, 115, 643–647. [Google Scholar] [CrossRef]
- Wasserman, G.A.; Liu, X.; Loiacono, N.J.; Kline, J.; Factor-Litvak, P.; van Geen, A.; Mey, J.L.; Levy, D.; Abramson, R.; Schwartz, A.; et al. A cross-sectional study of well water arsenic and child IQ in Maine schoolchildren. Environ. Health 2014, 13, 23. [Google Scholar]
- Hamadani, J.D.; Tofail, F.; Nermell, B.; Gardner, R.; Shiraji, S.; Bottai, M.; Arifeen, S.E.; Huda, S.N.; Vahter, M. Critical windows of exposure for arsenic-associated impairment of cognitive function in pre-school girls and boys: A population-based cohort study. Int. J. Epidemiol. 2011, 40, 1593–1604. [Google Scholar] [CrossRef]
- Parajuli, R.P.; Fujiwara, T.; Umezaki, M.; Furusawa, H.; Watanabe, C. Home environment and prenatal exposure to lead, arsenic and zinc on the neurodevelopment of six-month-old infants living in Chitwan Valley, Nepal. Neurotoxicol. Teratol. 2014, 41, 89–95. [Google Scholar] [CrossRef]
- Parajuli, R.P.; Fujiwara, T.; Umezaki, M.; Watanabe, C. Association of cord blood levels of lead, arsenic, and zinc with neurodevelopmental indicators in newborns: A birth cohort study in Chitwan Valley, Nepal. Environ. Res. 2013, 121, 45–51. [Google Scholar] [CrossRef]
- Luo, J.; Qiu, Z.; Chen, J.; Zhang, L.; Liu, W.; Tan, Y.; Shu, W. Maternal and early life arsenite exposure impairs neurodevelopment and increases the expression of PSA-NCAM in hippocampus of rat offspring. Toxicology 2013, 311, 99–106. [Google Scholar] [CrossRef]
- Rodriguez, V.M.; Carrizales, L.; Mendoza, M.S.; Fajardo, O.R.; Giordano, M. Effects of sodium arsenite exposure on development and behavior in the rat. Neurotoxicol. Teratol. 2002, 24, 743–750. [Google Scholar]
- Xi, S.; Sun, W.; Wang, F.; Jin, Y.; Sun, G. Transplacental and early life exposure to inorganic arsenic affected development and behavior in offspring rats. Arch. Toxicol. 2009, 83, 549–556. [Google Scholar] [CrossRef]
- Sannadi, S.; Kadeyala, P.K.; Gottipolu, R.R. Reversal effect of monoisoamyl dimercaptosuccinic acid (MiADMSA) for arsenic and lead induced perturbations in apoptosis and antioxidant enzymes in developing rat brain. Int. J. Dev. Neurosci. 2013, 31, 586–597. [Google Scholar] [CrossRef]
- Li, D.; Lu, C.; Wang, J.; Hu, W.; Cao, Z.; Sun, D.; Xia, H.; Ma, X. Developmental mechanisms of arsenite toxicity in zebrafish (Danio rerio) embryos. Aquat. Toxicol. 2009, 91, 229–237. [Google Scholar] [CrossRef]
- Radonjic, M.; Cappaert, N.L.; de Vries, E.F.; de Esch, C.E.; Kuper, F.C.; van Waarde, A.; Dierckx, R.A.; Wadman, W.J.; Wolterbeek, A.P.; Stierum, R.H.; et al. Delay and impairment in brain development and function in rat offspring after maternal exposure to methylmercury. Toxicol. Sci. 2013, 133, 112–124. [Google Scholar] [CrossRef]
- Jayashankar, S.; Glover, C.N.; Folven, K.I.; Brattelid, T.; Hogstrand, C.; Lundebye, A.K. Cerebral gene expression and neurobehavioural responses in mice pups exposed to methylmercury and docosahexaenoic acid through the maternal diet. Environ. Toxicol. Pharmacol. 2012, 33, 26–38. [Google Scholar]
- Jayashankar, S.; Glover, C.N.; Folven, K.I.; Brattelid, T.; Hogstrand, C.; Lundebye, A.K. Cerebral gene expression in response to single or combined gestational exposure to methylmercury and selenium through the maternal diet. Cell Biol. Toxicol. 2011, 27, 181–197. [Google Scholar]
- Glover, C.N.; Zheng, D.; Jayashankar, S.; Sales, G.D.; Hogstrand, C.; Lundebye, A.K. Methylmercury speciation influences brain gene expression and behavior in gestationally-exposed mice pups. Toxicol. Sci. 2009, 110, 389–400. [Google Scholar] [CrossRef]
- Ho, N.Y.; Yang, L.; Legradi, J.; Armant, O.; Takamiya, M.; Rastegar, S.; Strähle, U. Gene responses in the central nervous system of zebrafish embryos exposed to the neurotoxicant methyl mercury. Environ. Sci. Technol. 2013, 47, 3316–3325. [Google Scholar]
- Cauli, O.; Piedrafita, B.; Llansola, M.; Felipo, V. Gender differential effects of developmental exposure to methyl-mercury, polychlorinated biphenyls 126 or 153, or its combinations on motor activity and coordination. Toxicology 2013, 311, 61–68. [Google Scholar]
- Kasten-Jolly, J.; Heo, Y.; Lawrence, D.A. Central nervous system cytokine gene expression: Modulation by lead. J. Biochem. Mol. Toxicol. 2011, 25, 41–54. [Google Scholar] [CrossRef]
- Peterson, S.M.; Zhang, J.; Weber, G.; Freeman, J.L. Global gene expression analysis reveals dynamic and developmental stage-dependent enrichment of lead-induced neurological gene alterations. Environ. Health Perspect. 2011, 119, 615–621. [Google Scholar]
- Luo, M.; Xu, Y.; Cai, R.; Tang, Y.; Ge, M.M.; Liu, Z.H.; Xu, L.; Hu, F.; Ruan, D.Y.; Wang, H.L. Epigenetic histone modification regulates developmental lead exposure induced hyperactivity in rats. Toxicol. Lett. 2014, 225, 78–85. [Google Scholar] [CrossRef]
- Molina, R.M.; Phattanarudee, S.; Kim, J.; Thompson, K.; Wessling-Resnick, M.; Maher, T.J.; Brain, J.D. Ingestion of Mn and Pb by rats during and after pregnancy alters iron metabolism and behavior in offspring. Neurotoxicology 2011, 32, 413–422. [Google Scholar] [CrossRef]
- Rice, C.; Ghorai, J.K.; Zalewski, K.; Weber, D.N. Developmental lead exposure causes startle response deficits in zebrafish. Aquat. Toxicol. 2011, 105, 600–608. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Y.; Liu, W.; Bai, C.; Liu, X.; Liu, K.; Li, R.; Zhu, J.H.; Huang, C. Developmental lead acetate exposure induces embryonic toxicity and memory deficit in adult zebrafish. Neurotoxicol. Teratol. 2012, 34, 581–586. [Google Scholar] [CrossRef]
- Dou, C.; Zhang, J. Effects of lead on neurogenesis during zebrafish embryonic brain development. J. Hazard. Mater. 2011, 194, 277–282. [Google Scholar] [CrossRef]
- Pacyna, E.G.; Pacyna, J.M.; Sundseth, K.; Munthe, J.; Kindbom, K.; Wilson, S.; Steenhuisen, F.; Maxson, P. Global emission of mercury to the atmosphere from anthropogenic sources in 2005 and projections to 2020. Atmos. Environ. 2010, 44, 2487–2499. [Google Scholar] [CrossRef]
- Sheehan, M.C.; Burke, T.A.; Navas-Acien, A.; Breysse, P.N.; McGready, J.; Fox, M.A. Global methylmercury exposure from seafood consumption and risk of developmental neurotoxicity: A systematic review. Bull. World Health Organ. 2014, 92, 254F–269F. [Google Scholar] [CrossRef]
- Wiener, J.G.; Suchanek, T.H. The basis for ecotoxicological concern in aquatic ecosystems contaminated by historical mercury mining. Ecol. Appl. 2008, 18 (8 Suppl.), A3–A11. [Google Scholar] [CrossRef]
- Ekino, S.; Susa, M.; Ninomiya, T.; Imamura, K.; Kitamura, T. Minamata disease revisited: An update on the acute and chronic manifestations of methyl mercury poisoning. J. Neurol. Sci. 2007, 262, 131–144. [Google Scholar] [CrossRef]
- Harada, M. Minamata disease: Methylmercury poisoning in Japan caused by environmental pollution. Crit. Rev. Toxicol. 1995, 25, 1–24. [Google Scholar] [CrossRef]
- Shamlaye, C.; Davidson, P.W.; Myers, G.J. The Seychelles child development study: Two decades of collaboration. Seychelles Med. Dent. J. 2004, 7, 92–98. [Google Scholar]
- Davidson, P.W.; Cory-Slechta, D.A.; Thurston, S.W.; Huang, L.S.; Shamlaye, C.F.; Gunzler, D.; Watson, G.; van Wijngaarden, E.; Zareba, G.; Klein, J.D.; et al. Fish consumption and prenatal methylmercury exposure: Cognitive and behavioral outcomes in the main cohort at 17 years from the Seychelles child development study. Neurotoxicology 2011, 32, 711–717. [Google Scholar] [CrossRef]
- Davidson, P.W.; Myers, G.J.; Cox, C.; Axtell, C.; Shamlaye, C.; Sloane-Reeves, J.; Cernichiari, E.; Needham, L.; Choi, A.; Wang, Y.; et al. Effects of prenatal and postnatal methylmercury exposure from fish consumption on neurodevelopment: Outcomes at 66 months of age in the Seychelles Child Development Study. JAMA 1998, 280, 701–707. [Google Scholar] [CrossRef]
- Myers, G.J.; Davidson, P.W.; Cox, C.; Shamlaye, C.F.; Choisy, O.; Cernichiari, E.; Choi, A.; Sloane-Reeves, J.; Axtell, C.; Gao, P.; et al. The Seychelles child development study: Results and new directions through twenty-nine months. Water Air Soil Pollut. 1997, 97, 53–61. [Google Scholar]
- Myers, G.J.; Davidson, P.W.; Cox, C.; Shamlaye, C.F.; Palumbo, D.; Cernichiari, E.; Sloane-Reeves, J.; Wilding, G.E.; Kost, J.; Huang, L.S.; et al. Prenatal methylmercury exposure from ocean fish consumption in the Seychelles child development study. Lancet 2003, 361, 1686–1692. [Google Scholar] [CrossRef]
- Debes, F.; Budtz-Jorgensen, E.; Weihe, P.; White, R.F.; Grandjean, P. Impact of prenatal methylmercury exposure on neurobehavioral function at age 14 years. Neurotoxicol. Teratol. 2006, 28, 536–547. [Google Scholar] [CrossRef]
- Grandjean, P.; Weihe, P.; White, R.F.; Debes, F.; Araki, S.; Yokoyama, K.; Murata, K.; Sorensen, N.; Dahl, R.; Jorgensen, P.J. Cognitive deficit in 7-year-old children with prenatal exposure to methylmercury. Neurotoxicol. Teratol. 1997, 19, 417–428. [Google Scholar] [CrossRef]
- Grandjean, P.; White, R.F.; Weihe, P.; Jorgensen, P.J. Neurotoxic risk caused by stable and variable exposure to methylmercury from seafood. Ambul. Pediatr. 2003, 3, 18–23. [Google Scholar] [CrossRef]
- Murata, K.; Weihe, P.; Budtz-Jorgensen, E.; Jorgensen, P.J.; Grandjean, P. Delayed brainstem auditory evoked potential latencies in 14-year-old children exposed to methylmercury. J. Pediatr. 2004, 144, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Saint-Amour, D.; Roy, M.S.; Bastien, C.; Ayotte, P.; Dewailly, E.; Despres, C.; Gingras, S.; Muckle, G. Alterations of visual evoked potentials in preschool Inuit children exposed to methylmercury and polychlorinated biphenyls from a marine diet. Neurotoxicology 2006, 27, 567–578. [Google Scholar] [CrossRef]
- Julvez, J.; Debes, F.; Weihe, P.; Choi, A.; Grandjean, P. Sensitivity of continuous performance test (CPT) at age 14 years to developmental methylmercury exposure. Neurotoxicol. Teratol. 2010, 32, 627–632. [Google Scholar] [CrossRef]
- Rice, D.C. Effects of developmental methylmercury exposure on nervous system function in monkeys. In Methylmercury and Neurotoxicity; Ceccatelli, S., Aschner, M., Eds.; Springer Science + Busniess Media, LLC: New York, NY, USA, 2012; Chapter 8; pp. 143–161. [Google Scholar]
- Gunderson, V.M.; Grant, K.S.; Burbacher, T.M.; Fagan, J.F., 3rd; Mottet, N.K. The effect of low-level prenatal methylmercury exposure on visual recognition memory in infant crab-eating macaques. Child Dev. 1986, 57, 1076–1083. [Google Scholar] [CrossRef]
- Gunderson, V.M.; Grant-Webster, K.S.; Burbacher, T.M.; Mottet, N.K. Visual recognition memory deficits in methylmercury-exposed Macaca fascicularis infants. Neurotoxicol. Teratol. 1988, 10, 373–379. [Google Scholar] [CrossRef]
- Burbacher, T.M.; Sackett, G.P.; Mottet, N.K. Methylmercury effects on the social behavior of Macaca fascicularis infants. Neurotoxicol. Teratol. 1990, 12, 65–71. [Google Scholar]
- Rice, D.C.; Gilbert, S.G. Early chronic low-level methylmercury poisoning in monkeys impairs spatial vision. Science 1982, 216, 759–761. [Google Scholar]
- Rice, D.C.; Gilbert, S.G. Effects of developmental exposure to methyl mercury on spatial and temporal visual function in monkeys. Toxicol. Appl. Pharmacol. 1990, 102, 151–163. [Google Scholar] [CrossRef]
- Newland, M.C.; Hoffman, D.J.; Heath, J.C.; Donlin, W.D. Response inhibition is impaired by developmental methylmercury exposure: Acquisition of low-rate lever-pressing. Behav. Brain Res. 2013, 253, 196–205. [Google Scholar] [CrossRef]
- Beyrouty, P.; Stamler, C.J.; Liu, J.N.; Loua, K.M.; Kubow, S.; Chan, H.M. Effects of prenatal methylmercury exposure on brain monoamine oxidase activity and neurobehaviour of rats. Neurotoxicol. Teratol. 2006, 28, 251–259. [Google Scholar] [CrossRef]
- Bjorklund, O.; Kahlstrom, J.; Salmi, P.; Ogren, S.O.; Vahter, M.; Chen, J.F.; Fredholm, B.B.; Dare, E. The effects of methylmercury on motor activity are sex- and age-dependent, and modulated by genetic deletion of adenosine receptors and caffeine administration. Toxicology 2007, 241, 119–133. [Google Scholar] [CrossRef]
- Miyamoto, K.; Nakanishi, H.; Moriguchi, S.; Fukuyama, N.; Eto, K.; Wakamiya, J.; Murao, K.; Arimura, K.; Osame, M. Involvement of enhanced sensitivity of N-methyl-D-aspartate receptors in vulnerability of developing cortical neurons to methylmercury neurotoxicity. Brain Res. 2001, 901, 252–258. [Google Scholar] [CrossRef]
- Onishchenko, N.; Tamm, C.; Vahter, M.; Hokfelt, T.; Johnson, J.A.; Johnson, D.A.; Ceccatelli, S. Developmental exposure to methylmercury alters learning and induces depression-like behavior in male mice. Toxicol. Sci. 2007, 97, 428–437. [Google Scholar] [CrossRef]
- Roegge, C.S.; Morris, J.R.; Villareal, S.; Wang, V.C.; Powers, B.E.; Klintsova, A.Y.; Greenough, W.T.; Pessah, I.N.; Schantz, S.L. Purkinje cell and cerebellar effects following developmental exposure to PCBs and/or MeHg. Neurotoxicol. Teratol. 2006, 28, 74–85. [Google Scholar]
- Bertossi, M.; Girolamo, F.; Errede, M.; Virgintino, D.; Elia, G.; Ambrosi, L.; Roncali, L. Effects of methylmercury on the microvasculature of the developing brain. Neurotoxicology 2004, 25, 849–857. [Google Scholar] [CrossRef]
- Coccini, T.; Roda, E.; Castoldi, A.F.; Poli, D.; Goldoni, M.; Vettori, M.V.; Mutti, A.; Manzo, L. Developmental exposure to methylmercury and 2,2',4,4',5,5'-hexachlorobiphenyl (PCB153) affects cerebral dopamine D1-like and D2-like receptors of weanling and pubertal rats. Arch. Toxicol. 2011, 85, 1281–1294. [Google Scholar] [CrossRef]
- Roda, E.; Coccini, T.; Acerbi, D.; Castoldi, A.; Bernocchi, G.; Manzo, L. Cerebellum cholinergic muscarinic receptor (subtype-2 and -3) and cytoarchitecture after developmental exposure to methylmercury: An immunohistochemical study in rat. J. Chem. Neuroanat. 2008, 35, 285–294. [Google Scholar] [CrossRef]
- Stringari, J.; Nunes, A.K.; Franco, J.L.; Bohrer, D.; Garcia, S.C.; Dafre, A.L.; Milatovic, D.; Souza, D.O.; Rocha, J.B.; Aschner, M.; et al. Prenatal methylmercury exposure hampers glutathione antioxidant system ontogenesis and causes long-lasting oxidative stress in the mouse brain. Toxicol. Appl. Pharmacol. 2008, 227, 147–154. [Google Scholar] [CrossRef]
- Perino, J.; Ernhart, C.B. The relation of subclinical lead level to cognitive and sensorimotor impairment in black preschoolers. J. Learn. Disabil. 1974, 7, 616–620. [Google Scholar] [CrossRef]
- De la Burdé, B.; Choate, M.S. Early asymptomatic lead exposure and development at school age. J. Pediatr. 1975, 87, 638–642. [Google Scholar] [CrossRef]
- Bellinger, D.; Leviton, A.; Waternaux, C.; Needleman, H.; Rabinowitz, M. Longitudinal analyses of prenatal and postnatal lead exposure and early cognitive development. N. Engl. J. Med. 1987, 316, 1037–1043. [Google Scholar]
- Dietrich, K.N.; Krafft, K.M.; Bornschein, R.L.; Hammond, P.B.; Berger, O.; Succop, P.A.; Bier, M. Low-level fetal lead exposure effect on neurobehavioral development in early infancy. Pediatrics 1987, 80, 721–730. [Google Scholar]
- Liu, J.; Gao, D.; Chen, Y.; Jing, J.; Hu, Q.; Chen, Y. Lead exposure at each stage of pregnancy and neurobehavioral development of neonates. Neurotoxicology 2014, 44C, 1–7. [Google Scholar]
- Surkan, P.J.; Zhang, A.; Trachtenberg, F.; Daniel, D.B.; McKinlay, S.; Bellinger, D.C. Neuropsychological function in children with blood lead levels <10 microg/dL. Neurotoxicology 2007, 28, 1170–1177. [Google Scholar]
- Bushnell, P.J.; Bowman, R.E. Persistence of impaired reversal learning in young monkeys exposed to low levels of dietary lead. J. Toxicol. Environ. Health 1979, 5, 1015–1023. [Google Scholar] [CrossRef]
- Bushnell, P.J.; Bowman, R.E. Reversal learning deficits in young monkeys exposed to lead. Pharmacol. Biochem. Behav. 1979, 10, 733–742. [Google Scholar] [CrossRef]
- Levin, E.D.; Schneider, M.L.; Ferguson, S.A.; Schantz, S.L.; Bowman, R.E. Behavioral effects of developmental lead exposure in rhesus monkeys. Dev. Psychobiol. 1988, 21, 371–382. [Google Scholar] [CrossRef]
- Rice, D.C. Behavioral effects of lead: Commonalities between experimental and epidemiologic data. Environ. Health Perspect. 1996, 104 (Suppl. 2), 337–351. [Google Scholar] [CrossRef]
- Laughlin, N.K.; Bushnell, P.J.; Bowman, R.E. Lead exposure and diet: Differential effects on social development in the rhesus monkey. Neurotoxicol. Teratol. 1991, 13, 429–440. [Google Scholar]
- Devi, C.B.; Reddy, G.H.; Prasanthi, R.P.; Chetty, C.S.; Reddy, G.R. Developmental lead exposure alters mitochondrial monoamine oxidase and synaptosomal catecholamine levels in rat brain. Int. J. Dev. Neurosci. 2005, 23, 375–381. [Google Scholar] [CrossRef]
- Jett, D.A.; Beckles, R.A.; Navoa, R.V.; McLemore, G.L. Increased high-affinity nicotinic receptor-binding in rats exposed to lead during development. Neurotoxicol. Teratol. 2002, 24, 805–811. [Google Scholar] [CrossRef]
- Reddy, G.R.; Devi, B.C.; Chetty, C.S. Developmental lead neurotoxicity: Alterations in brain cholinergic system. Neurotoxicology 2007, 28, 402–407. [Google Scholar]
- Wang, X.M.; Liu, W.J.; Zhang, R.; Zhou, Y.K. Effects of exposure to low-level lead on spatial learning and memory and the expression of mGluR1, NMDA receptor in different developmental stages of rats. Toxicol. Ind. Health 2013, 29, 686–696. [Google Scholar] [CrossRef]
- Petit, T.L.; LeBoutillier, J.C.; Brooks, W.J. Altered sensitivity to NMDA following developmental lead exposure in rats. Physiol. Behav. 1992, 52, 687–693. [Google Scholar]
- Schneider, J.S.; Mettil, W.; Anderson, D.W. Differential effect of postnatal lead exposure on gene expression in the hippocampus and frontal cortex. J. Mol. Neurosci. 2012, 47, 76–88. [Google Scholar] [CrossRef]
- Xu, J.; Yan, H.C.; Yang, B.; Tong, L.S.; Zou, Y.X.; Tian, Y. Effects of lead exposure on hippocampal metabotropic glutamate receptor subtype 3 and 7 in developmental rats. J. Negat. Results Biomed. 2009, 8, 5. [Google Scholar] [CrossRef]
- Zito, K.; Scheuss, V. NMDA Receptor Function and Physiological Modulation. In Encyclopedia of Neuroscience; Squire, L.R., Ed.; Academic Press: Oxford, UK, 2009; Volume 6; pp. 1157–1164. [Google Scholar]
- Kim, Y.; Ha, E.H.; Park, H.; Ha, M.; Kim, Y.; Hong, Y.C.; Kim, E.J.; Kim, B.N. Prenatal lead and cadmium co-exposure and infant neurodevelopment at 6 months of age: The mothers and children’s environmental health (MOCEH) study. Neurotoxicology 2013, 35, 15–22. [Google Scholar] [CrossRef]
- Rai, A.; Maurya, S.K.; Khare, P.; Srivastava, A.; Bandyopadhyay, S. Characterization of developmental neurotoxicity of As, Cd, and Pb mixture: Synergistic action of metal mixture in glial and neuronal functions. Toxicol. Sci. 2010, 118, 586–601. [Google Scholar] [CrossRef]
- Wasserman, G.A.; Liu, X.; Parvez, F.; Factor-Litvak, P.; Ahsan, H.; Levy, D.; Kline, J.; van Geen, A.; Mey, J.; Slavkovich, V.; et al. Arsenic and manganese exposure and children’s intellectual function. Neurotoxicology 2011, 32, 450–457. [Google Scholar] [CrossRef]
- Fischer, C.; Fredriksson, A.; Eriksson, P. Coexposure of neonatal mice to a flame retardant PBDE 99 (2,2',4,4',5-pentabromodiphenyl ether) and methyl mercury enhances developmental neurotoxic defects. Toxicol. Sci. 2008, 101, 275–285. [Google Scholar] [CrossRef]
- Vitalone, A.; Catalani, A.; Cinque, C.; Fattori, V.; Matteucci, P.; Zuena, A.R.; Costa, L.G. Long-term effects of developmental exposure to low doses of PCB 126 and methylmercury. Toxicol. Lett. 2010, 197, 38–45. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 2013, 10, 70–86. [Google Scholar]
- Li, X.; Ma, Y.; Li, D.; Gao, X.; Li, P.; Bai, N.; Luo, M.; Tan, X.; Lu, C.; Ma, X. Arsenic impairs embryo development via down-regulating Dvr1 expression in zebrafish. Toxicol. Lett. 2012, 212, 161–168. [Google Scholar] [CrossRef]
- Long, Y.; Li, Q.; Li, J.; Cui, Z. Molecular analysis, developmental function and heavy metal-induced expression of ABCC5 in zebrafish. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2011, 158, 46–55. [Google Scholar] [CrossRef]
- Hassan, S.A.; Moussa, E.A.; Abbott, L.C. The effect of methylmercury exposure on early central nervous system development in the zebrafish (Danio rerio) embryo. J. Appl. Toxicol. 2012, 32, 707–713. [Google Scholar]
- Cuello, S.; Ximenez-Embun, P.; Ruppen, I.; Schonthaler, H.B.; Ashman, K.; Madrid, Y.; Luque-Garcia, J.L.; Camara, C. Analysis of protein expression in developmental toxicity induced by MeHg in zebrafish. Analyst 2012, 137, 5302–5311. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Freeman, J.L. Zebrafish as a model for investigating developmental lead (Pb) neurotoxicity as a risk factor in adult neurodegenerative disease: A mini-review. Neurotoxicology 2014, 43, 57–64. [Google Scholar] [CrossRef]
- Peterson, S.M.; Zhang, J.; Freeman, J.L. Developmental reelin expression and time point-specific alterations from lead exposure in zebrafish. Neurotoxicol. Teratol. 2013, 38, 53–60. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, J.; Freeman, J.L. Zebrafish as a Model for Developmental Neurotoxicity Assessment: The Application of the Zebrafish in Defining the Effects of Arsenic, Methylmercury, or Lead on Early Neurodevelopment. Toxics 2014, 2, 464-495. https://doi.org/10.3390/toxics2030464
Lee J, Freeman JL. Zebrafish as a Model for Developmental Neurotoxicity Assessment: The Application of the Zebrafish in Defining the Effects of Arsenic, Methylmercury, or Lead on Early Neurodevelopment. Toxics. 2014; 2(3):464-495. https://doi.org/10.3390/toxics2030464
Chicago/Turabian StyleLee, Jinyoung, and Jennifer L. Freeman. 2014. "Zebrafish as a Model for Developmental Neurotoxicity Assessment: The Application of the Zebrafish in Defining the Effects of Arsenic, Methylmercury, or Lead on Early Neurodevelopment" Toxics 2, no. 3: 464-495. https://doi.org/10.3390/toxics2030464