
Motor Sequence Learning in Healthy Older Adults Is Not Necessarily Facilitated by Transcranial Direct Current Stimulation (tDCS)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Method and Materials
2.1. Experiment One
2.1.1. Participants
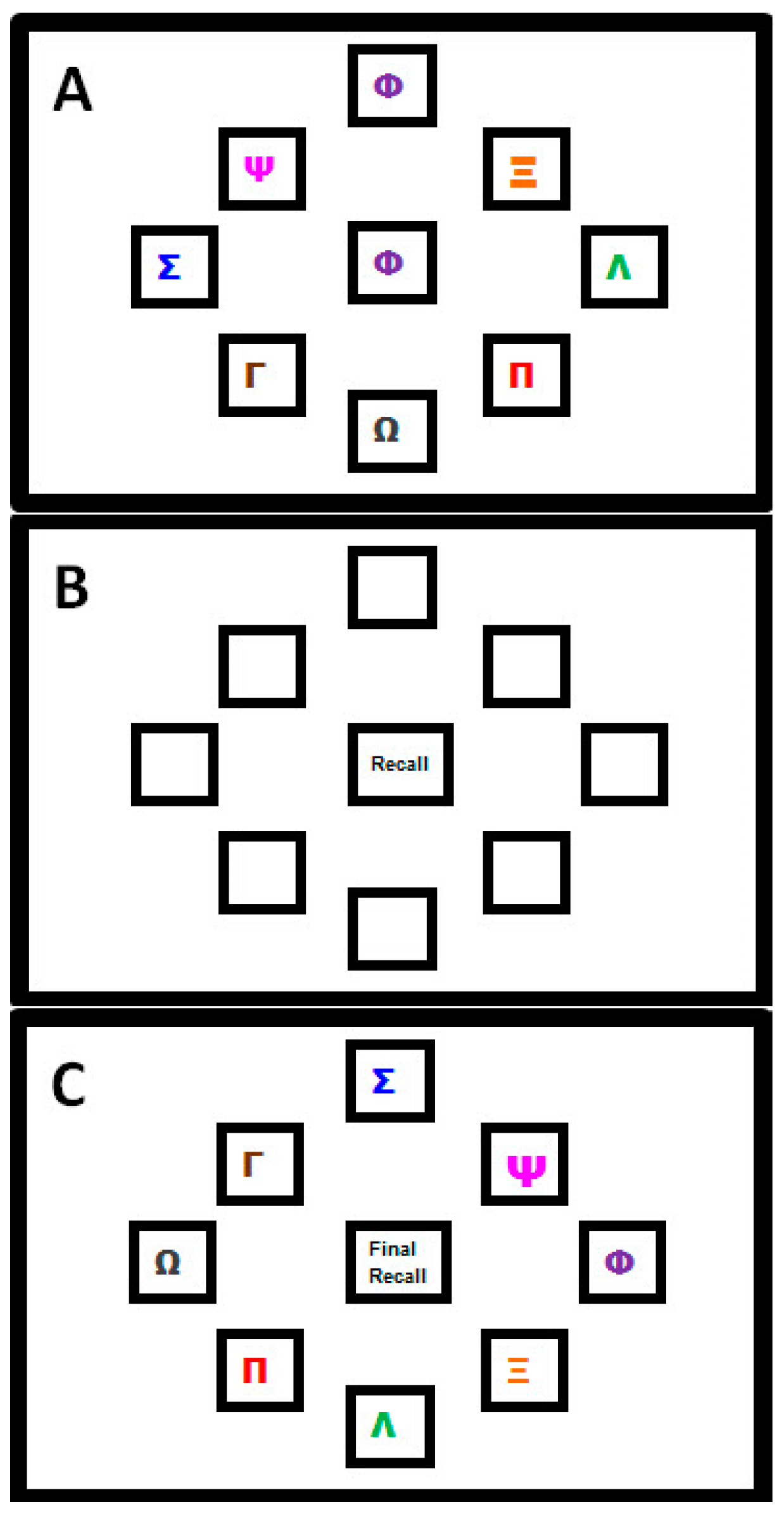
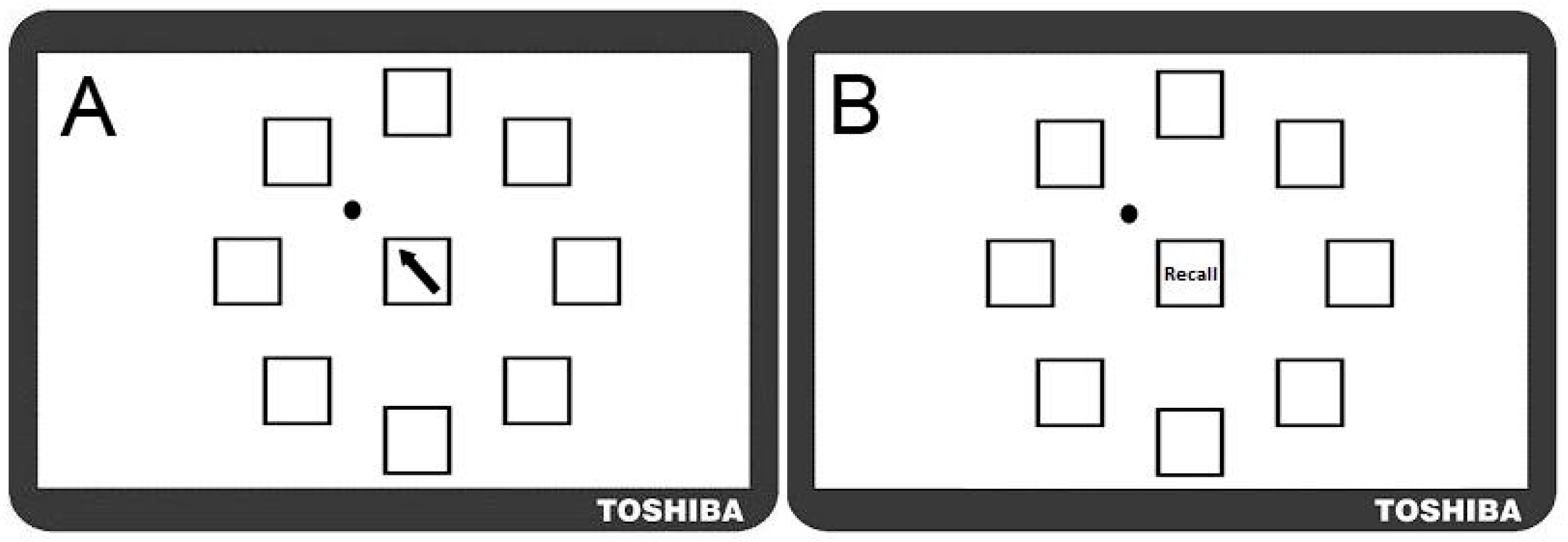
2.1.2. Motor Sequence Learning Task
2.1.3. Procedure for Transcranial Direct Current Stimulation (tDCS)
2.1.4. Analysis
- (i)
- Sequence Learning Measure: Number of moves recalled in the correct sequential order (i.e., Correctly Recalled; CR), with a maximum score of 32. Points were not deducted for incorrect moves;
- (ii)
- Movement Speed Measure: Recall Movement Time (MT), the mean time (s) taken to move the mouse from the centre to a target box when recalling the sequence, was taken as an indication of motor performance.
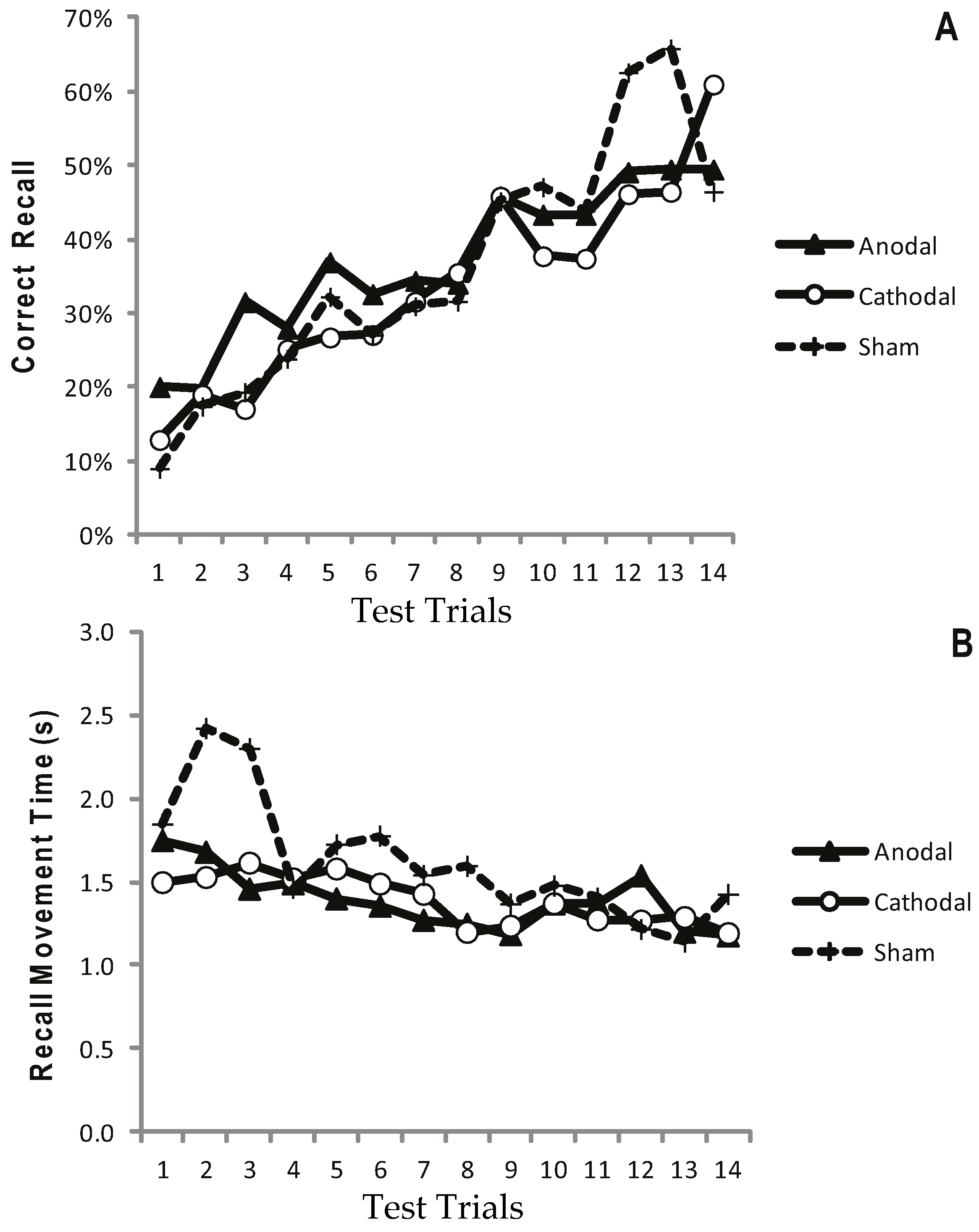
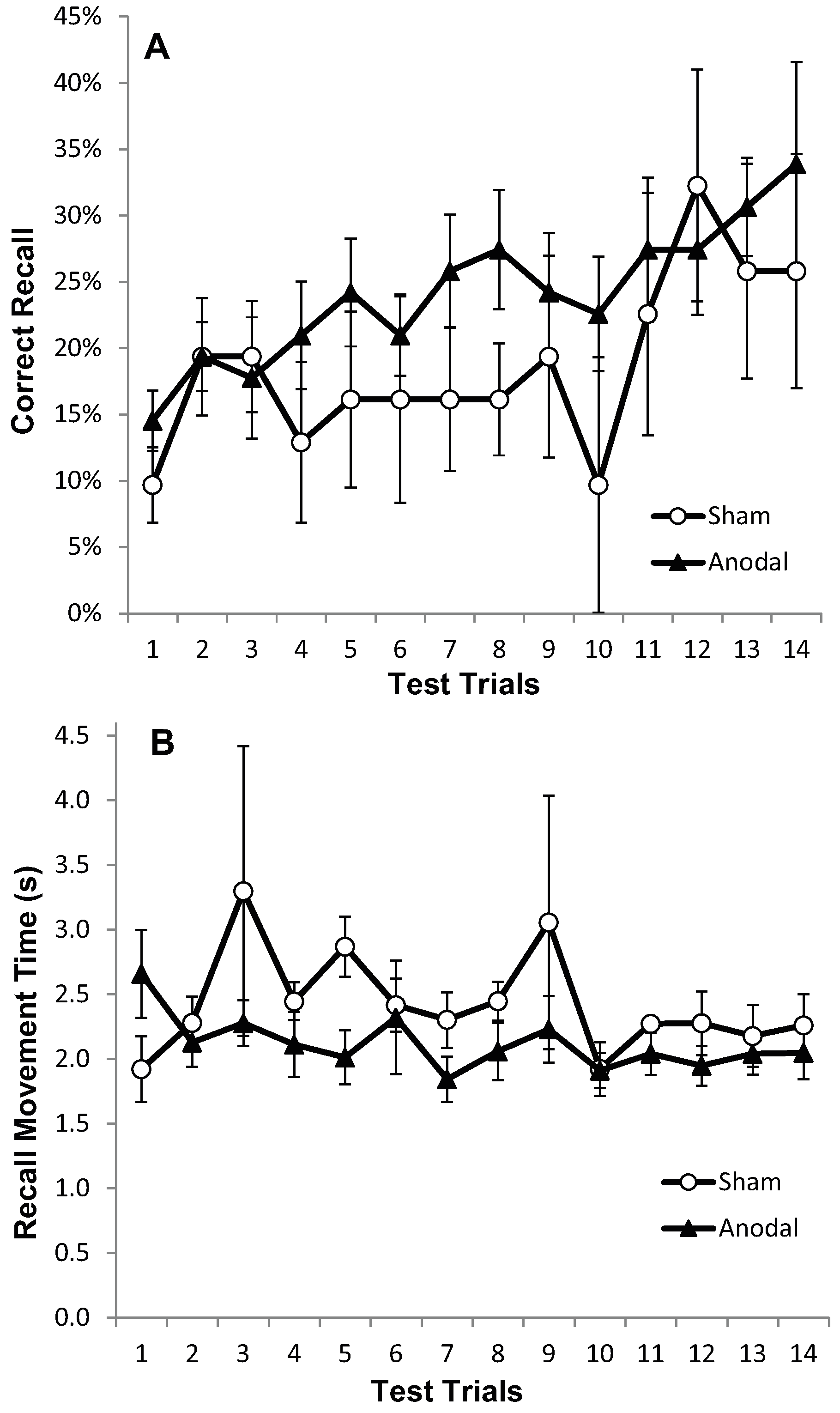
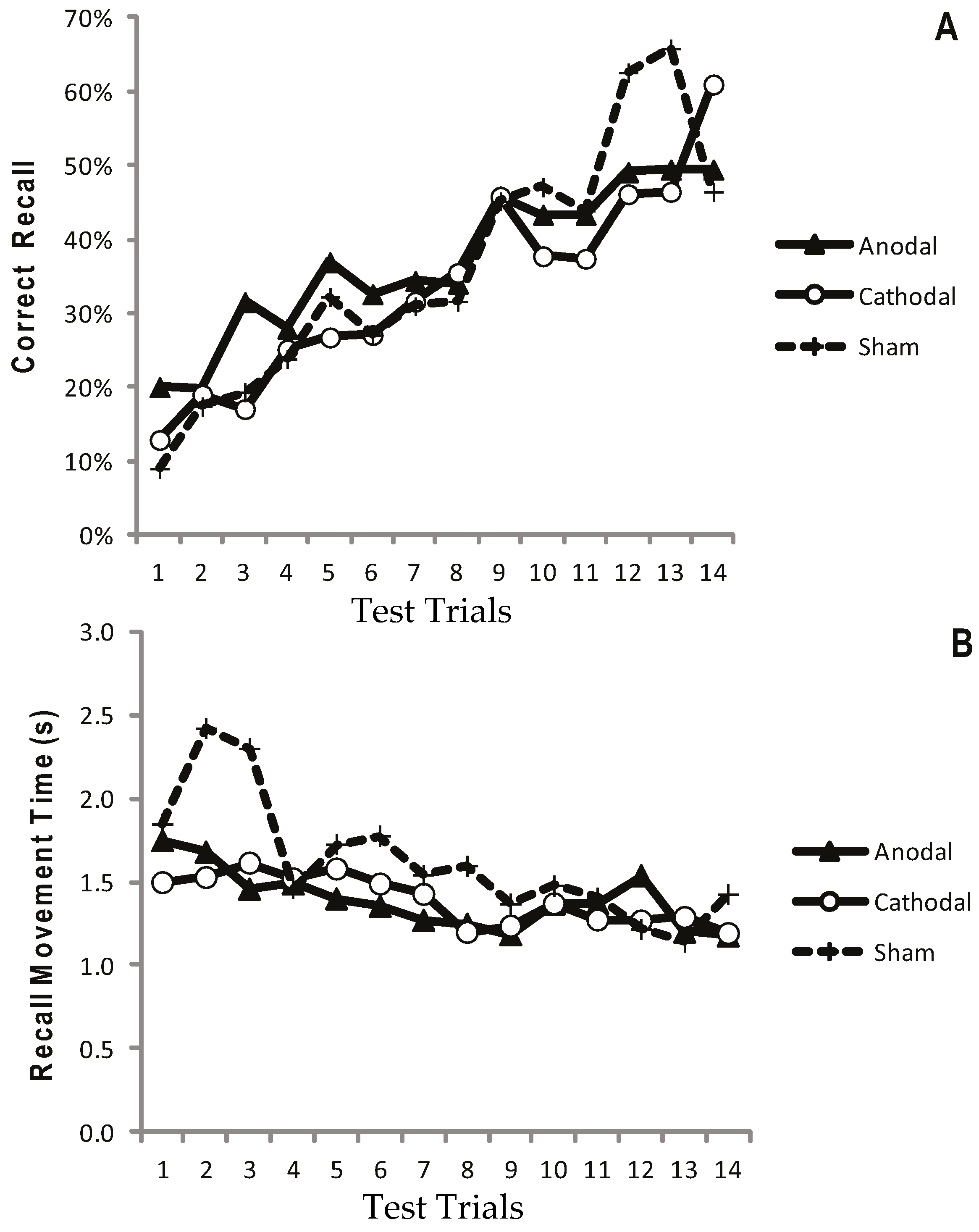
2.1.5. Results: Test Trials
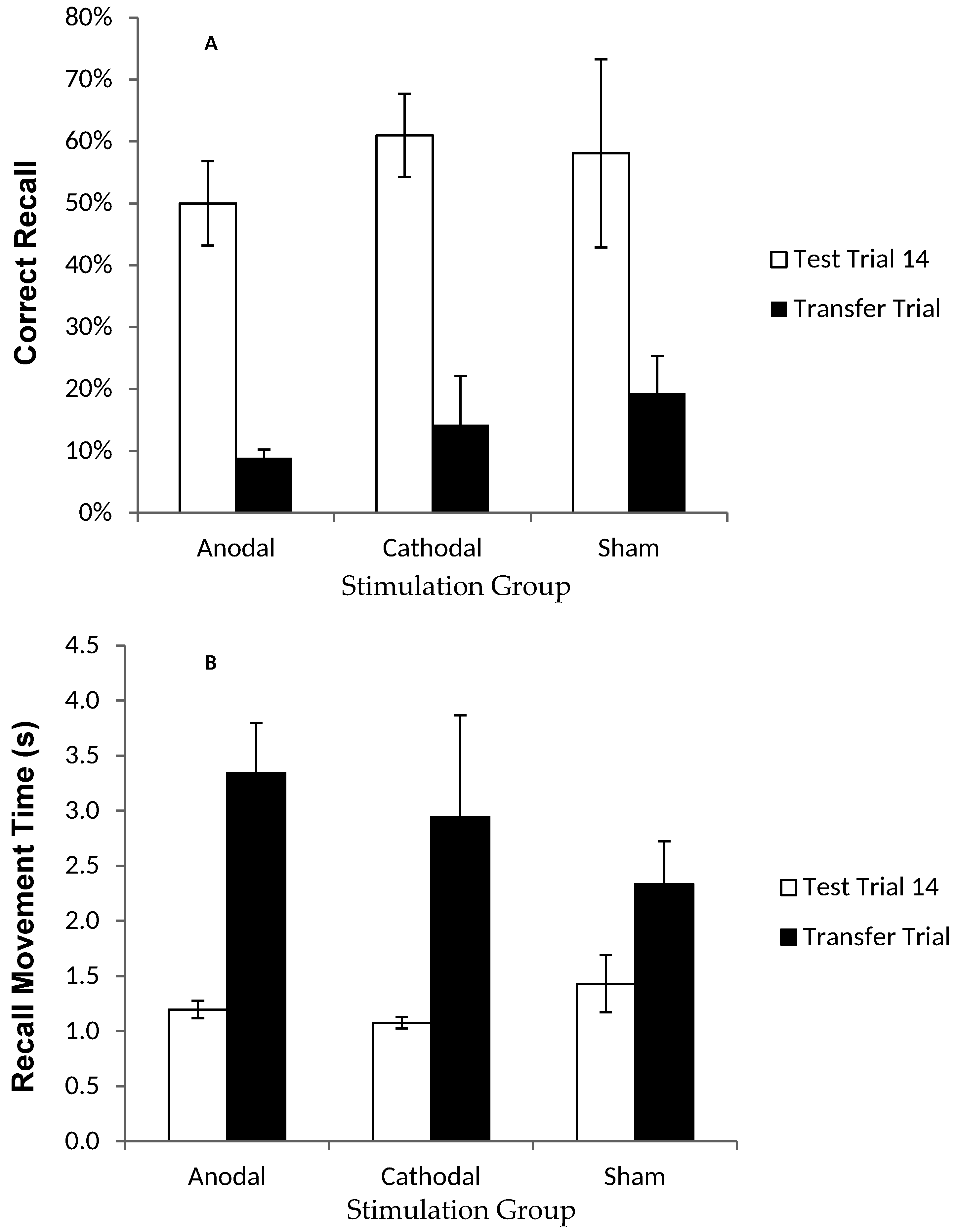
2.1.6. Results: Recall in the Transfer Trial
2.1.7. Discussion
2.2. Experiment Two
2.2.1. Participants
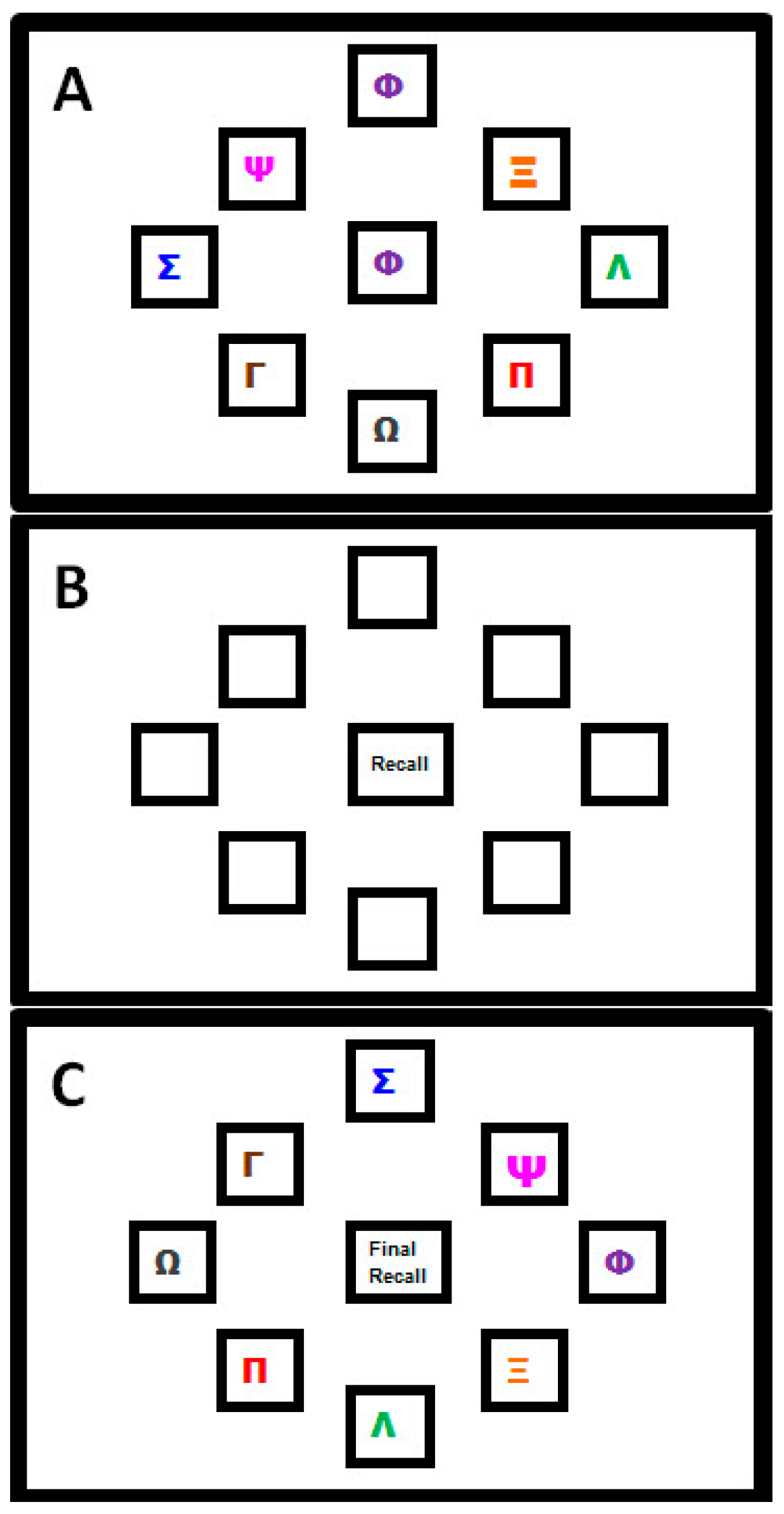
2.2.2. Motor Sequence Learning Task
2.2.3. Procedure for Transcranial Direct Current Stimulation (tDCS)
2.2.4. Analysis
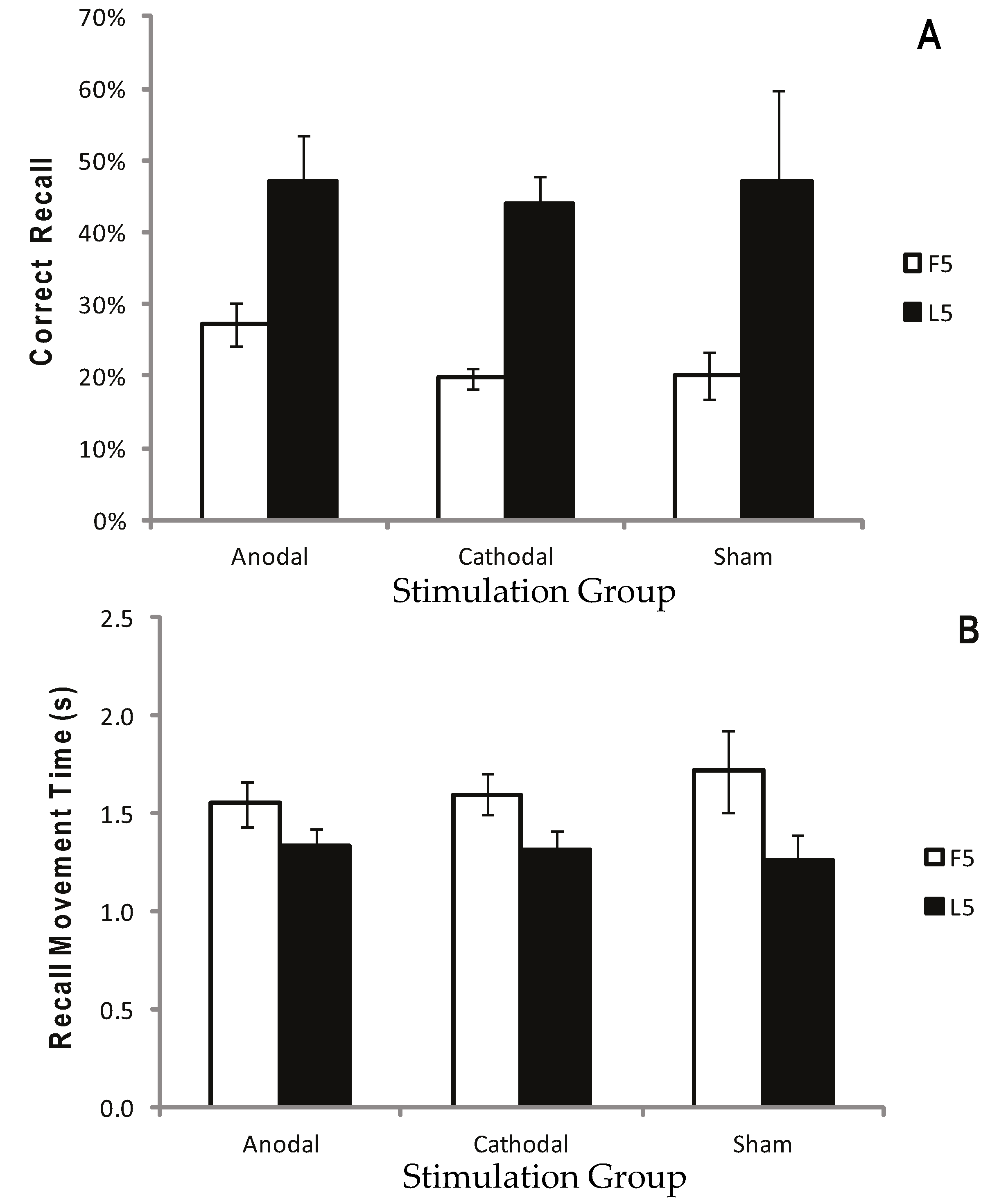
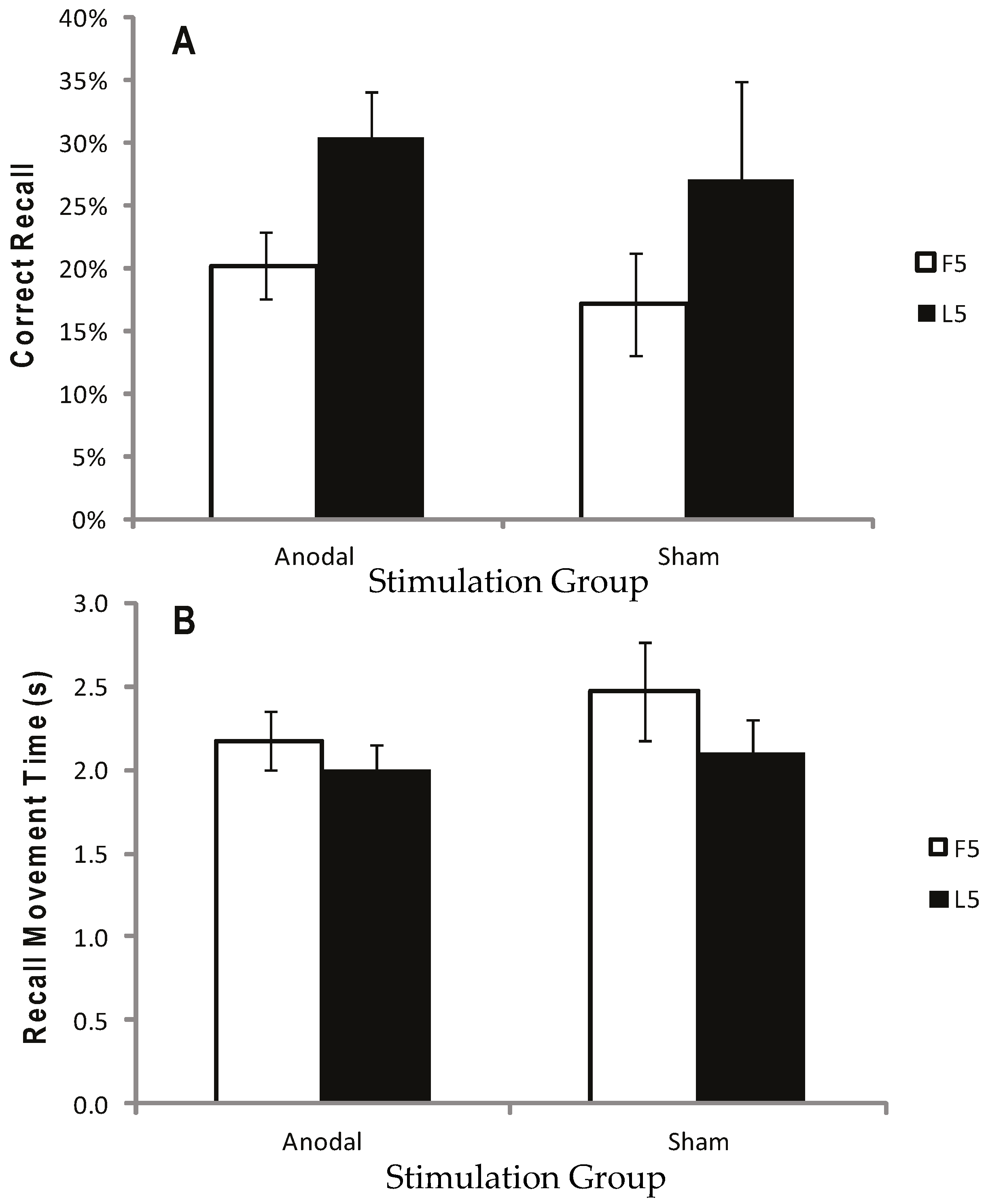
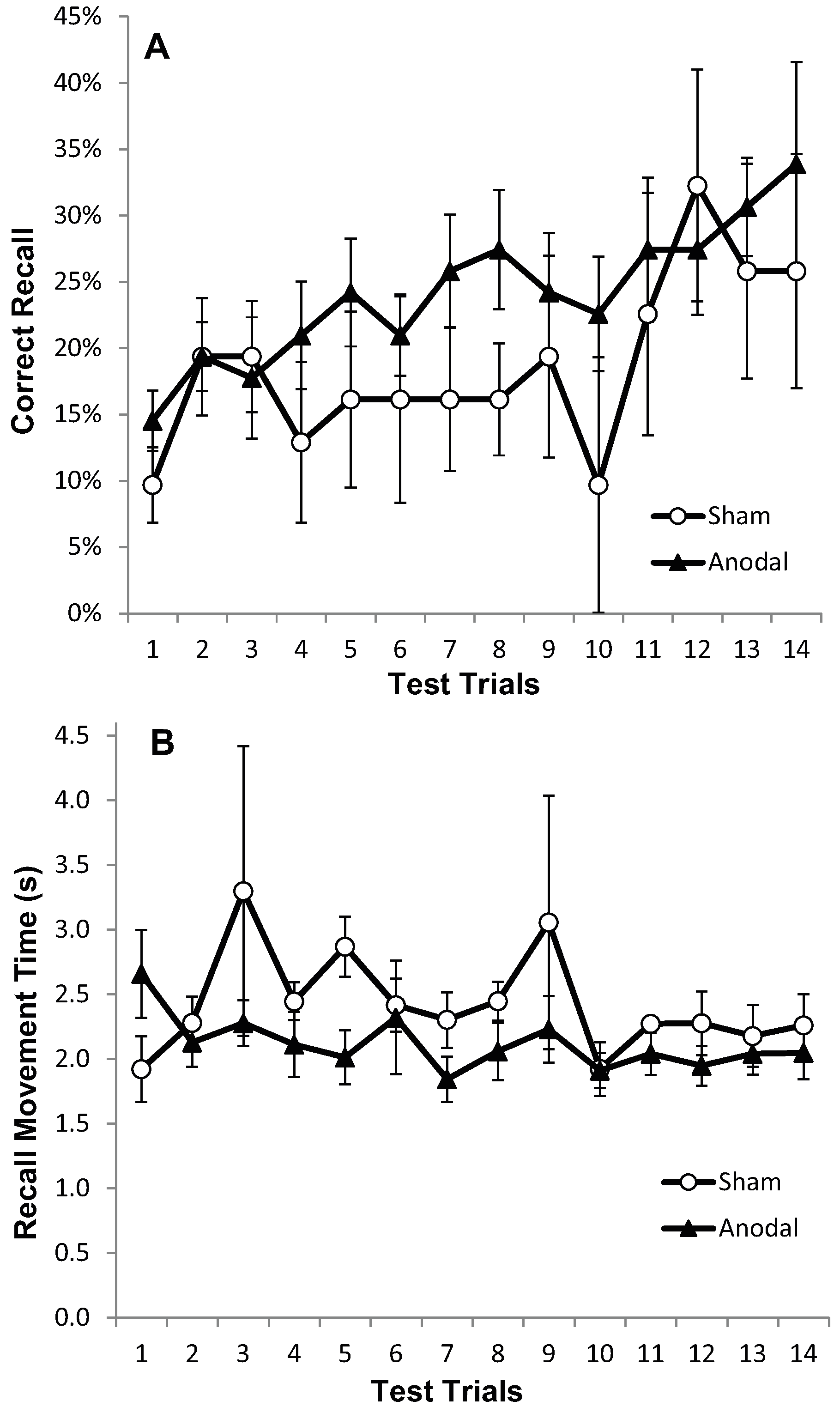
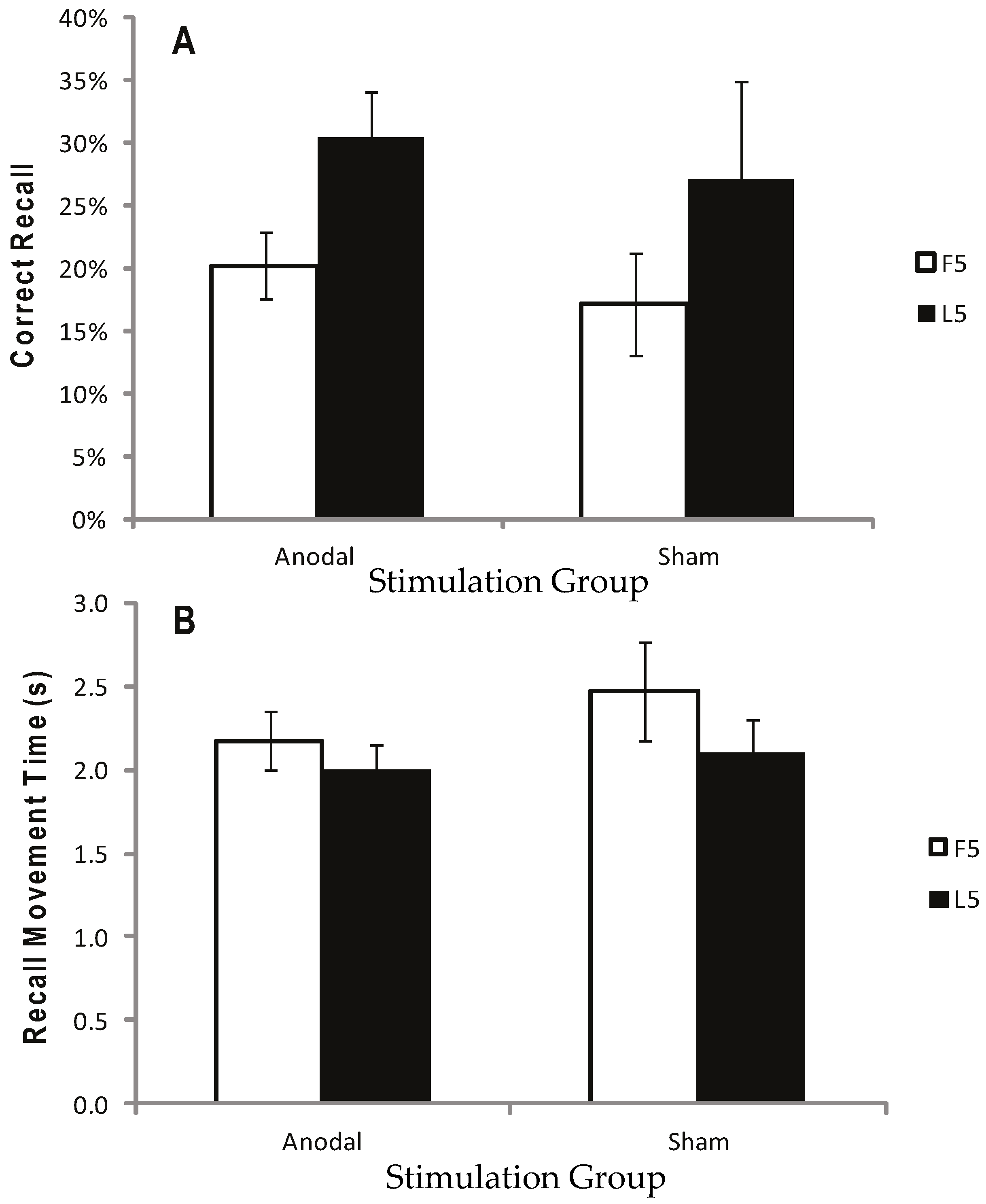
2.2.5. Results
2.2.6. Discussion
3. General Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Barnett, C.H.; Cobbold, A.F. Effects of age upon the mobility of human finger joints. Ann. Rheum. Dis. 1968, 27, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.J.; McComas, A.J.; Petito, F. Physiological changes in ageing muscles. J. Neurol. Neurosurg. Psychiatry 1973, 36, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Delbono, O. Neural control of aging skeletal muscle. Aging Cell 2003, 2, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, J.A.; Larkin, L.M.; Claflin, D.R.; Brooks, S.V. Age-related changes in the structure and function of skeletal muscles. Clin. Exp. Pharmacol. Physiol. 2007, 34, 1091–1096. [Google Scholar] [CrossRef] [PubMed]
- Clark, B.C.; Taylor, J.L. Age-related changes in motor cortical properties and voluntary activation of skeletal muscle. Curr. Aging Sci. 2011, 4, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Wolf, P.A.; D’Agostino, R.B.; Belanger, A.J.; Kannel, W.B. Probability of Stroke: A Risk Profile from the Framingham Study. Stroke 1991, 22, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Kelly-Hayes, M.; Beiser, A.; Kase, C.S.; Scaramucci, A.; D’Agostino, R.B.; Wolf, P.A. The influence of gender and age on disability following ischemic stroke: The Framingham study. J. Stroke Cerebrovasc. Dis. 2003, 12, 119–126. [Google Scholar] [CrossRef]
- Rantenen, T.; Guralnik, J.M.; Foley, D.; Masaki, K.; Leveille, S.; Curb, D.J.; White, L. Midlife hand grip strength as a predictor of old age disability. JAMA 1999, 281, 558–560. [Google Scholar] [CrossRef]
- Giampaoli, S.; Ferrucci, L.; Cecchi, F.; Noce, C.L.; Poce, A.; Dima, F.; Santaquilani, A.; Vescio, M.F.; Menotti, A. Hand-grip strength predicts incident disability in non-disabled older men. Age Ageing 1999, 24, 283–288. [Google Scholar] [CrossRef]
- Katz, S.; Ford, A.B.; Moskowitz, R.W.; Jackson, B.A.; Jaffee, M.W. Studies of illnes in the aged. The index of ADL: A standardized measure of biological and psychological function. JAMA 1963, 185, 914–919. [Google Scholar] [CrossRef] [PubMed]
- Zimerman, M.; Nitsch, M.; Giraux, P.; Gerloff, C.; Cohen, L.G.; Hummel, F.C. Neuroenhancement of the aging brain: Restoring skill acquisition in old subjects. Ann. Neurol. 2013, 73, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Fuortes, M.G.F. Direct Current Stimulation of Motoneurones. J. Physiol. 1954, 126, 494–506. [Google Scholar] [CrossRef] [PubMed]
- Hern, J.E.C.; Landgren, S.; Phillips, C.G.; Porter, R. Selective excitation of corticofungal neurones by surface-anodal stimulation of the baboon’s motor cortex. J. Physiol. 1962, 161, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Bindman, L. Long-lasting changes in level of electrical activity of cerebral cortex produced by polarizing currents. Nature 1962, 196, 584–585. [Google Scholar] [CrossRef] [PubMed]
- Bindman, L.J.; Lippold, C.J.; Redfeard, W.T. The action of brief polarizing currents on the cerebral cortex of the rat (1) during current flow and (2) in the production of long-lasting after-effects. J. Physiol. 1964, 172, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.A.; Imamura, M.; Fregni, F. Updates of the Use of Non-Invasive Brain Stimulation in Physical and Rehabilitation Medicine. J. Rehabil. Med. 2009, 41, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Priori, A.; Beradelli, A.; Rona, S.; Accornero, N.; Manfredi, M. Polarization of the human motor cortex through the scalp. Neuroreport 1998, 9, 2257–2260. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Paulus, W. Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J. Physiol. 2000, 527, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Paulus, W. Sustained excitability elevations induced by transcranial DC motor cortex stimulation in humans. Neurology 2001, 57, 1899–1901. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Nitsche, M.S.; Klein, C.C.; Tergau, F.; Rothwell, J.C.; Paulus, W. Level of action of cathodal DC polarisation induced inhibition of the human motor cortex. Clin. Neurophysiol. 2003, 114, 600–604. [Google Scholar] [CrossRef]
- Stagg, C.J.; O’Shea, J.; Kincses, Z.T.; Woolrich, M.; Matthews, P.M.; Johansen-Berg, H. Modulation of movement-associated cortical activation by transcranial direct current stimulation. Eur. J. Neurosci. 2009, 30, 1412–1423. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.H.; Ahn, S.H.; Byun, W.M.; Kim, C.S.; Lee, M.Y.; Kwon, Y.H. The effect of transcranial direct current stimulation on the cortical activation by motor task in the human brain: An fMRI study. Neurosci. Lett. 2009, 460, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Paulus, W. Transcranial direct current stimulation—Update 2011. Restor. Neurol. Neurosci. 2011, 29, 463–492. [Google Scholar] [PubMed]
- Kwon, H.Y.; Ko, M.; Ahn, S.H.; Kim, Y.; Song, J.C.; Lee, C.; Chang, M.C.; Jang, S.H. Primary motor cortex activation by transcranial direct current stimulation in the human brain. Neurosci. Lett. 2008, 435, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, L.; Koslowsky, M.; Lavidor, M. tDCS polarity effects in motor and cognitive domains: A meta-analytical review. Exp. Brain Res. 2012, 216, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Venkatakrishnan, A.; Sandrini, M. Combining transcranial direct current stimulation and neuroimaging: Novel insights in understanding neuroplasticity. J. Neurophysiol. 2012, 107, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Orban, J.-J.; Civry, O.; Shadmehr, R. Electrifying the motor engram: Effects of tDCS on motor learning and control. Exp. Brain Res. 2014, 232, 3379–3395. [Google Scholar] [CrossRef] [PubMed]
- Boros, K.; Poreisz, C.; Münchau, A.; Paulus, W.; Nitsche, M.A. Premotor transcranial direct current stimulation (tDCS) affects primary motor excitability in humans. Eur. J. Neurosci. 2008, 27, 1292–1300. [Google Scholar] [CrossRef] [PubMed]
- Lang, N.; Siebner, H.R.; Ward, N.S.; Lee, L.; Nitsche, M.A.; Paulus, W.; Rothwell, J.C.; Lemon, R.N.; Frackowiak, R.S. How does transcranial DC stimulation of the primary motor cortex alter regional neuronal activity in the human brain? Eur. J. Neurosci. 2005, 22, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Truong, D.; Minhas, P.; Parra, L.C.; Bikson, M. Inter-individual variation during transcranial direct current stimulation and normalization of dose using MRI-derived computational models. Front. Psychiatry 2012, 3, 91. [Google Scholar] [CrossRef] [PubMed]
- Taub, E.; Uswatte, G.; Pidikiti, R. Constraint-Induced Movement Therapy: A New Family of Techniques with Broad Application to Physical Rehabilitation--A Clinical Review. J. Rehabil. Res. Dev. 1999, 36, 237–251. [Google Scholar] [PubMed]
- Fregni, F.; Boggio, P.S.; Mansur, C.G.; Wagner, T.; Ferreira, M.J.; Lima, M.C.; Rigonatti, S.P.; Marcolin, M.A.; Freedman, S.D.; Nitsche, M.A.; et al. Transcranial direct current stimulation of the unaffected hemisphere in stroke patients. Neuroreport 2005, 16, 1551–1555. [Google Scholar] [CrossRef] [PubMed]
- Boggio, P.S.; Nunes, A.; Rigonatti, S.P.; Nitsche, M.A.; Pascual-Leone, A.; Fregni, F. Repeated sessions of noninvasive brain DC stimulation is associated with motor function improvement in stroke patients. Restor. Neurol. Neurosci. 2007, 25, 123–129. [Google Scholar] [PubMed]
- Tresilian, J. Sensorimotor Control & Learning: An Introduction to the Behavioural Neuroscience of Action; Palgrave Macmillan: New York, NY, USA, 2012. [Google Scholar]
- Raw, R.K.; Allen, R.; Williams, J.H.G.; Mon-Williams, M.; Wilkie, R. Novel movement pattern acquisition as a function of age and hand: Does better performance mean better learning? J. Exp. Psychol. Gen. 2016. submitted. [Google Scholar]
- Zimerman, M.; Heise, K.F.; Hoppe, J.; Cohen, L.G.; Gerloff, C.; Hummel, F.C. Modulation of training by single-session transcranial direct current stimulation to the intact motor cortex enhances motor skill acquisition of the paretic hand. Stroke 2012, 43, 2185–2191. [Google Scholar] [CrossRef] [PubMed]
- Marquez, J.; van Vliet, P.; McElduff, P.; Lagopoulos, J.; Parsons, M. Transcranial direct current stimulation (tDCS): Does it have merit in stroke rehabilitation? A systematic review. Int. J. Stroke 2015, 10, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Kandel, M.; Beis, J.-M.; Le Chapelain, L.; Guesdon, H.; Paysant, J. Non-invasive cerebral stimulation for the upper limb rehabilitation after stroke: A review. Ann. Phys. Rehabil. Med. 2012, 55, 657–680. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.J.; Shuster, M.; O’Hara, E.; Hurley, K.; Middlebrooks, D.; Guilkey, K. A meta-analysis of the efficacy of anodal transcranial direct current stimulation for upper limb motor recovery in stroke survivors. J. Hand Ther. 2013, 26, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Bastani, A.; Jaberzadeh, S. Does anodal transcranial direct current stimulation enhance excitability of the motor cortex and motor function in healthy individuals and subjects with stroke: A systematic review and meta-analysis. Clin. Neurophysiol. 2012, 123, 644–657. [Google Scholar] [CrossRef] [PubMed]
- Elsner, B.; Kugler, J.; Pohl, M.; Mehrholz, J. Transcranial direct current stimulation (tDCS) for improving function and activities of daily living in patients after stroke. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef]
- Boggio, P.S.; Castro, L.O.; Savagim, E.A.; Braite, R.; Cruz, V.C.; Rocha, R.R.; Rigonatti, S.P.; Silva, M.T.; Fregni, F. Enhancement of non-dominant hand motor function by anodal transcranial direct current stimulation. Neurosci. Lett. 2006, 404, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Vines, B.W.; Nair, D.G.; Schlaug, G. Contralateral and Ipsilateral Motor Effects after Transcranial Direct Current Stimulation. Neuroreport 2006, 17, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Cogiamanian, F.; Marceglia, S.; Ardolino, G.; Barbieri, S.; Priori, A. Improved isometric force endurance after transcranial direct current stimulation over the human motor cortical areas. Eur. J. Neurosci. 2007, 26, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Vines, B.W.; Cerruti, C.; Schlaug, G. Dual-Hemisphere tDCS facilitates greater improvements for healthy subjects’ non-dominant hand compared to uni-hemisphere stimulation. BMC Neurosci. 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, A.; Maeoka, H.; Hiyamizu, M.; Shomoto, K.; Morioka, S.; Seki, K. Enhancement of precise hand movements by transcranial direct current stimulation. Neuroreport 2011, 22, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Foerster, A.; Rocha, S.; Wiesiolek, C.; Chagas, A.P.; Machado, G.; Silva, E.; Fregni, F.; Monte-Silva, K. Site-specific effects of mental practice combined with transcranial direct current stimulation on motor learning. Eur. J. Neurosci. 2013, 37, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Convento, S.; Bolognini, N.; Fusaro, M.; Lollo, F.; Vallar, G. Neuromodulation of parietal and motor activity affects motor planning and execution. Cortex 2014, 57, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Hummel, F.C.; Heise, K.; Celnik, P.; Floel, A.; Gerloff, C.; Cohen, L.G. Facilitating skilled right hand function in older subjects by anodal polarization over the left primary cortex. Neurobiol. Aging 2010, 31, 2160–2168. [Google Scholar] [CrossRef] [PubMed]
- Goodwill, A.M.; Reynolds, J.; Daly, R.M.; Kidgell, D.J. Formation of cortical plasticity in older adults following tDCS and motor training. Front. Aging Neurosci. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Parikh, P.J.; Cole, K.J. Effects of transcranial direct current stimulation in combination with motor practice on dexterous grasping and manipulation in healthy older adults. Physiol. Rep. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, B.J.; Bullock, D.; Verwey, W.B.; Averbeck, B.B.; Page, M.P.A. Learning and production of movement sequences: Behavioral, neurophysiological, and modeling perspectives. Hum. Mov. Sci. 2004, 23, 699–746. [Google Scholar] [CrossRef] [PubMed]
- Voelcker-Rehage, C.; Godde, B.; Staudinger, U.M. Physical and motor fitness are both related to cognition in old age. Eur. J. Neurosci. 2010, 31, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Fricke, K.; Henschke, U.; Schlitterlau, A.; Liebetanz, D.; Lang, N.; Hennig, S.; Tergan, F.; Paulus, W. Pharmacological modulation of cortical excitability shifts induced by transcranial direct current stimulation in humans. J. Physiol. 2003, 553, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.K.; Paik, N.-J. Effect of tDCS electrode montage on implicit motor sequence learning in healthy subjects. Exp. Transl. Stroke Med. 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Stagg, C.J.; Jayaram, G.; Pastor, D.; Kincses, Z.T.; Matthews, P.M.; Johansen-Berg, H. Polarity and timing-dependent effects of transcranial direct current stimulation in explicit motor learning. Neuropsychologia 2011, 49, 800–804. [Google Scholar] [CrossRef] [PubMed]
- Horvath, J.C.; Forte, J.D.; Carter, O. Evidence that transcranial direct current stimulation (tDCS) generates little-to-no reliable neurophysiologic effect beyond MEP amplitude modulation in healthy human subjects: A systematic review. Neuropsychologia 2015, 66, 213–236. [Google Scholar] [CrossRef] [PubMed]
- Saiote, C.; Polanía, R.; Rosenberger, K.; Paulus, W.; Antal, A. High-Frequency TRNS Reduces BOLD Activity during Visuomotor Learning. PLoS ONE 2013, 8, e59669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minarik, T.; Sauseng, P.; Dunne, L.; Berger, B.; Sterr, A. Effects of anodal transcranial Direct Current Stimulation on visually guided learning of grip force control. Biology 2015, 4, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Morrell, F. Effect of anodal polarization on the firing pattern of single cortical cells. Ann. N. Y. Acad. Sci. 1961, 92, 860–876. [Google Scholar] [CrossRef] [PubMed]
- Herzfeld, D.J.; Pastor, D.; Haith, A.M.; Rossetti, Y.; Shadmehr, R.; O’Shea, J. Contributions of the cerebellum and the motor cortex to acquisition and retention of motor memories. Neuroimage 2014, 98, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Jebsen, R.H.; Taylor, N.; Trieschmann, R.B.; Trotter, M.J.; Howard, L.A. An objective and Standardized Test of Hand Function. Arch. Phys. Med. Rehabil. 1969, 50, 311–319. [Google Scholar] [PubMed]
- Morgan, M.; Phillips, J.G.; Bradshaw, J.L.; Mattingly, J.B.; Iansek, R.; Bradshaw, J.A. Age-related motor slowness: Simply strategic? J. Gerontol. A. Biol. Sci. 1994, 49, 133–139. [Google Scholar] [CrossRef]
- Welsh, T.N.; Higgins, L.; Elliot, D. Are there age-related differences in learning to optimize speed, accuracy and energy expenditure? Hum. Mov. Sci. 2007, 26, 892–912. [Google Scholar] [CrossRef] [PubMed]
- Raw, R.K.; Kountouriotis, G.K.; Mon-Williams, M.; Wilkie, R.M. Movement control in older adults: Does old age mean middle of the road? J. Exp. Psychol. Hum. Percept. Perform. 2012, 38, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Raw, R.K.; Wilkie, R.; Culmer, P.; Mon-Williams, M. Reduced motor asymmetry in older adults tracing paths. Exp. Brain Res. 2012, 217, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Raw, R.K.; Wilkie, R.; White, A.; Williams, J.H.G.; Mon-Williams, M. Structural learning predicts the ‘goldilocks zone’ needed for getting the measure of manual asymmetries. PLoS ONE 2015, 10, e0128322. [Google Scholar] [CrossRef] [PubMed]
- Nissen, M.J.; Bullemer, P. Attentional Requirements of Learning: Evidence from Performance Measures. Cogn. Psychol. 1987, 19, 1–32. [Google Scholar] [CrossRef]
- Vancleef, K.; Meesen, R.; Swinnen, S.P.; Fujiyama, H. tDCS over left M1 or DLPFC does not improve learning of a bimanual coordination task. Sci. Rep. 2016, 25, 35739. [Google Scholar] [CrossRef] [PubMed]
- Culmer, P.R.; Levesley, M.C.; Mon-Williams, M.; Williams, J.H. A new tool for assessing human movement: The Kinematic Assessment Tool. J. Neurosci. Methods 2009, 184, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Kane, M.J.; Engle, R.W. Working-memory capacity, proactive interference and divided attention: Limits on long-term memory retrieval. J. Exp. Psychol. Learn. Mem. Cogn. 2000, 26, 336–358. [Google Scholar] [CrossRef] [PubMed]
- Lustig, C.; May, C.P.; Hasher, L. Working memory span and the role of proactive interference. J. Exp. Psychol. Gen. 2001, 130, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Voelcker-Rehage, C. Motor-skill learning in older adults—A review of studies on age differences. Eur. Rev. Aging Phys. Act. 2008, 5, 5–16. [Google Scholar] [CrossRef]
- Bishop, N.A.; Lu, T.; Yankner, B.A. Neural Mechanisms of ageing and cognitive decline. Nature 2010, 464, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Reis, J.; Schambra, H.K.; Cohen, L.G.; Buch, E.R.; Fritsch, B.; Zarahn, E.; Celnik, P.A.; Krakauer, J.W. Noninvasive cortical stimulation enhances motor skill acquisition over multiple days through an effect on consolidation. Proc. Natl. Acad. Sci. USA 2009, 106, 1590–1595. [Google Scholar] [CrossRef] [PubMed]
- Tecchio, F.; Zappasodi, F.; Assenza, G.; Tombini, M.; Vollaro, S.; Barbati, G.; Rossini, P.M. Anodal transcranial direct current stimulation enhances procedural consolidation. J. Neurophysiol. 2010, 104, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Price, A.R.; Hamilton, R.H. A Re-evaluation of the Cognitive Effects from Single-session Transcranial Direct Current Stimulation. Brain Stimul. 2015, 8, 663–665. [Google Scholar] [CrossRef] [PubMed]
- Horvath, J.C. New Quantitative Analyses Following Price & Hamilton’s Critique do not Change Original Findings of Horvath et al. Brain Stimul. 2015, 8, 665–666. [Google Scholar] [PubMed]
- Baddeley, A.D. Working memory: Theories, models, and controversies. Annu. Rev. Psychol. 2012, 63, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Grafton, S.T.; Hazeltine, E.; Ivry, R. Functional mapping of sequence learning in normal humans. J. Cogn. Neurosci. 1995, 7, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Kitaguchi, K.; Hikosaka, O. Chunking during human visuomotor sequence learning. Exp. Brain Res. 2003, 152, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Unsworth, N.; Engle, R.W. Individual differences in working memory capacity and learning: Evidence from the serial reaction time task. Mem. Cogn. 2005, 33, 213–220. [Google Scholar] [CrossRef]
- D’Esposito, M.; Aguirre, G.K.; Zarahn, E.; Ballard, D.; Shin, R.K.; Lease, J. Functional MRI studies of spatial and nonspatial working memory. Brain Res. Cogn. Brain Res. 1998, 7, 1–13. [Google Scholar] [CrossRef]
- Fregni, F.; Boggio, P.S.; Nitsche, M.; Bermpohl, F.; Antal, A.; Feredoes, E.; Marcolin, M.A.; Rigonatti, S.P.; Silva, M.T.A.; Paulus, W.; et al. Anodal transcranial direct current stimulation of prefrontal cortex enhances working memory. Exp. Brain Res. 2005, 166, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Ohn, S.H.; Park, C.-L.; Yoo, W.-K.; Ko, M.-H.; Choi, K.P.; Kim, G.-M.; Lee, Y.T.; Kim, Y.-H. Time-dependent effect of transcranial direct current stimulation on the enhancement of working memory. Neuroreport 2008, 19, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, I.H.; Brooks, D.J.; Nixon, P.D.; Frackowiak, R.S.; Passingham, R.E. Motor sequence learning: A study with positron emission tomography. J. Neurosci. 1994, 14, 3775–3790. [Google Scholar] [PubMed]
- Schendan, H.E.; Searl, M.M.; Melrose, R.J.; Stern, C.E. An fMRI study of the role of the medial temporal lobe in implicit and explicit sequence learning. Neuron 2003, 37, 1013–1025. [Google Scholar] [CrossRef]
- De Rover, M.; Pironti, V.A.; McCabe, J.A.; Acosta-Cabronero, J.; Arana, F.S.; Morein-Zamir, S.; Sahakian, B.J. Hippocampal dysfunction in patients with mild cognitive impairment: A functional neuroimaging study of a visuospatial paired associates learning task. Neuropsychologia 2011, 49, 2060–2070. [Google Scholar] [CrossRef] [PubMed]
- Maguire, E.A.; Frith, C.D.; Burgess, N.; Donnett, J.G.; O’Keefe, J. Knowing where things are parahippocampal involvement in encoding object locations in virtual large-scale space. J. Cogn. Neurosci. 1998, 10, 61–76. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.; Nadel, L. The Hippocampus as a Cognitive Map; Oxford University Press: Oxford, UK, 1978. [Google Scholar]
- Tanaka, S.; Hanakawa, T.; Honda, M.; Watanabe, K. Enhancement of pinch force in the lower leg by anodal transcranial direct current stimulation. Exp. Brain Res. 2009, 196, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Sandrini, M.; Cohen, L.G. Modulation of motor learning and memory formation by non-invasive cortical stimulation of the primary motor cortex. Neuropsychol. Rehabil. 2011, 21, 650–675. [Google Scholar] [CrossRef] [PubMed]
- Poreisz, C.; Boros, K.; Antal, A.; Paulus, W. Safety aspects of Transcranial direct current stimulation concerning healthy subjects and patients. Brain Res. Bull. 2007, 72, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Benninger, D.H.; Lomarev, M.; Lopez, G.; Pal, N.; Luckenbaugh, D.A.; Hallett, M. Transcranial Direct Current Stimulation for the Treatment of Focal Hand Dystonia. Mov. Disord. 2011, 26, 1698–1702. [Google Scholar] [CrossRef] [PubMed]
- Buttkus, F.; Weidenmüller, M.; Schneider, S.; Jabusch, H.-C.; Nitsche, M.A.; Paulus, P.; Altenmüller, E. Failure of Cathodal Direct Current Stimulation to improve Fine Motor Control in Musician’s Dystonia. Mov. Disord. 2010, 25, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Korpelainen, J.T.; Sotaniemi, K.A.; Huikuri, H.V.; Myllyla, V.V. Abnormal heart rate variability as a manifestation of autonomic dysfunction in hemispheric brain infarction. Stroke 1996, 27, 2059–2063. [Google Scholar] [CrossRef] [PubMed]
- Clancy, J.A.; Johnson, R.; Raw, R.K.; Deuchars, S.A.; Deuchars, J. Anodal transcranial direct current stimulation (tDCS) over the motor cortex increases sympathetic nerve activity. Brain Stimul. 2014, 7, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Sallustio, F.; Di Legge, S.; Stanzione, P.; Koch, G. Transcranial direct current stimulation of the affected hemisphere does not accelerate recovery of acute patients. Eur. J. Neurol. 2013, 20, 202–204. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.; Hesse, S.; Kroczek, G.; Waldner, A. Transcranial galvanic stimulation (tDCS) + robot-assisted therapy to improve upper limb impairment after stroke: A double-blind placebo controlled trial—preliminary results. Neurorehabil. Neural Repair 2008, 22, 550–551. [Google Scholar]
- Celnik, P.; Paik, N.J.; Vandermeeren, Y.; Dimyan, M.; Leonardo, G.C. Effects of Combined Peripheral Nerve Stimulation and Brain Polarization on Performance of a Motor Sequence Task After Chronic Stroke. Stroke 2009, 40, 1764–1771. [Google Scholar] [CrossRef] [PubMed]
- Hummel, F.C.; Cohen, L.G. Improvement of motor function with noninvasive cortical stimulation in a patient with chronic stroke. Neurorehabil. Neural Repair 2005, 19, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Hummel, F.C.; Celnik, P.; Giraux, P.; Floel, A.; Wu, W.-H.; Gerloff, C.; Cohen, L.G. Effects of non-invasive cortical stimulation on skilled motor function in chronic stroke. Brain 2005, 128, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Hummel, F.C.; Voller, B.; Celnik, P.; Floel, A.; Giraux, P.; Gerloff, C.; Cohen, L.G. Effects of brain polarization on reaction times and pinch force in chronic stroke. BMC Neurosci. 2006, 7. [Google Scholar] [CrossRef] [PubMed]
- Ko, M.H.; Han, S.H.; Park, S.H.; Seo, J.-H.; Kim, Y.-H. Improvement of visual scanning after DC brain polarization of parietal cortex in stroke patients with spatial neglect. Neurosci. Lett. 2008, 448, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Marquez, C.M.S.; Zhang, X.; Swinnen, S.P.; Meesen, R.; Wenderoth, N. Task-specific effect of transcranial direct current stimulation on motor learning. Front. Hum. Neurosci. 2013, 7, 333. [Google Scholar]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raw, R.K.; Allen, R.J.; Mon-Williams, M.; Wilkie, R.M. Motor Sequence Learning in Healthy Older Adults Is Not Necessarily Facilitated by Transcranial Direct Current Stimulation (tDCS). Geriatrics 2016, 1, 32. https://doi.org/10.3390/geriatrics1040032
Raw RK, Allen RJ, Mon-Williams M, Wilkie RM. Motor Sequence Learning in Healthy Older Adults Is Not Necessarily Facilitated by Transcranial Direct Current Stimulation (tDCS). Geriatrics. 2016; 1(4):32. https://doi.org/10.3390/geriatrics1040032
Chicago/Turabian StyleRaw, Rachael K., Richard J. Allen, Mark Mon-Williams, and Richard M. Wilkie. 2016. "Motor Sequence Learning in Healthy Older Adults Is Not Necessarily Facilitated by Transcranial Direct Current Stimulation (tDCS)" Geriatrics 1, no. 4: 32. https://doi.org/10.3390/geriatrics1040032