
A Matter of the Heart: The African Clawed Frog Xenopus as a Model for Studying Vertebrate Cardiogenesis and Congenital Heart Defects

Abstract
:1. Advantages of Xenopus as a Model Organism
2. Tool Kit for Studying Heart Development and Diseases in Xenopus
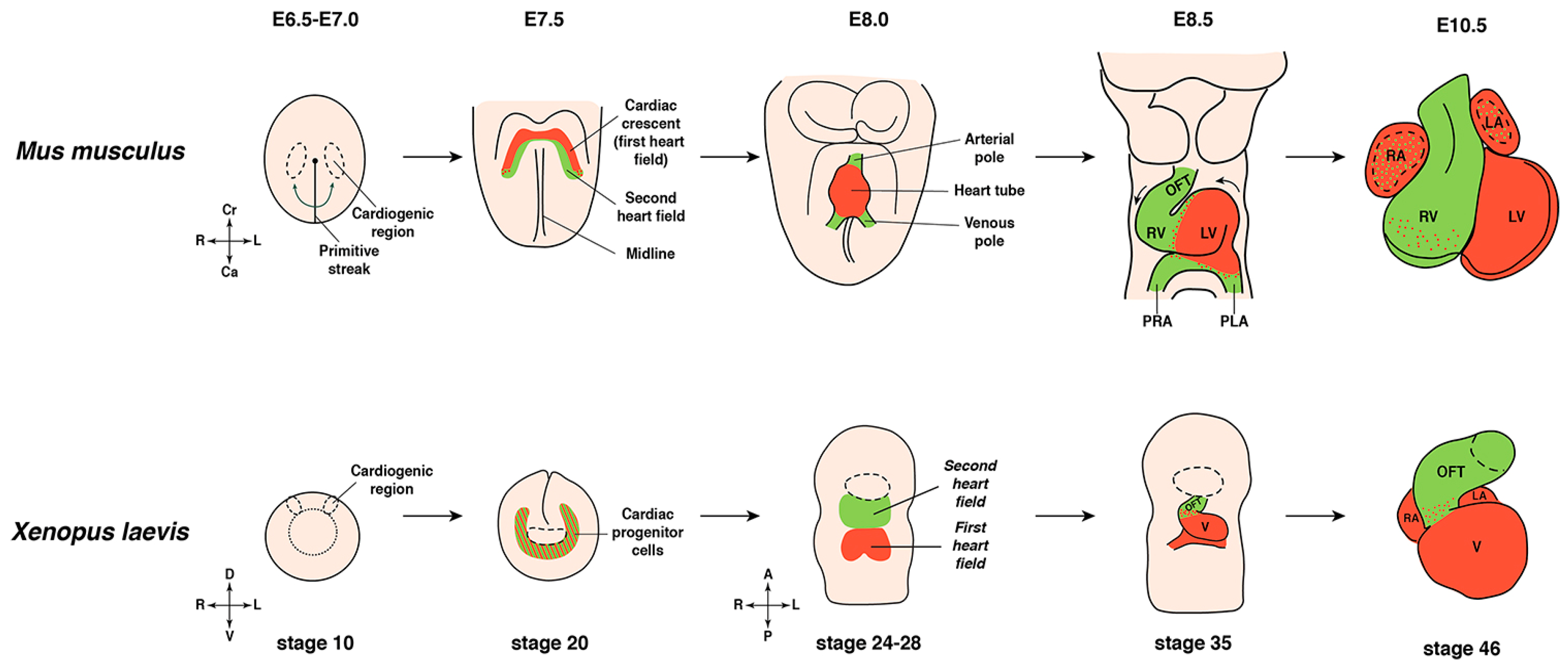
3. Xenopus as a Model for Vertebrate Cardiogenesis
4. Xenopus Models for Human Congenital Heart Defects
5. Modeling GWAS Candidates Associated with CHDs in Xenopus
Acknowledgments
Author Contributions
Conflicts of Interest
Human and Animal Rights and Informed Consent
Abbreviations
| ASD | atrial septal defects |
| CHARGE | Coloboma, Heart defects, choanal Atresia, Retarded growth and development |
| CHD | congenital heart defects |
| CRISPR | clustered regulatory inter-spaced short palindromic repeats |
| DMZ | dorsal marginal zone |
| FHF | first heart field |
| HLHS | hypoplastic left heart syndrome |
| HOS | Holt-Oram syndrome |
| LVOT | left ventricular outflow tract |
| MO | morpholino oligonucleotides |
| OFT | outflow tract |
| SHF | second heart field |
| VMZ | ventral marginal zone |
| VSD | ventricular septal defects |
References
- Wheeler, G.N.; Brandli, A.W. Simple vertebrate models for chemical genetics and drug discovery screens: Lessons from zebrafish and xenopus. Dev. Dyn. 2009, 238, 1287–1308. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, S.M.; Gull, M.; Brandli, A.W. Engineering Xenopus embryos for phenotypic drug discovery screening. Adv. Drug Deliv. Rev. 2014, 69–70, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Blitz, I.L.; Andelfinger, G.; Horb, M.E. Germ layers to organs: Using xenopus to study “later” development. Semin. Cell Dev. Biol. 2006, 17, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Nieuwkoop, P.D.; Faber, J. Normal Table of Xenopus Laevis (Daudin); a Systematical and Chronological Survey of the Development From the Fertilized Egg Till the End of Metamorphosis; North-Holland Publishing Co.: Amsterdam, The Netherlands, 1956; p. 243. [Google Scholar]
- Dale, L.; Slack, J.M. Fate map for the 32-cell stage of Xenopus laevis. Development 1987, 99, 527–551. [Google Scholar] [PubMed]
- Keller, R.E. Vital dye mapping of the gastrula and neurula of Xenopus laevis: II. Prospective areas and morphogenetic movements of the deep layer. Dev. Biol. 1976, 51, 118–137. [Google Scholar] [CrossRef]
- Moody, S.A. Fates of the blastomeres of the 32-cell-stage Xenopus embryo. Dev. Biol. 1987, 122, 300–319. [Google Scholar] [CrossRef]
- Gessert, S.; Kuhl, M. Comparative gene expression analysis and fate mapping studies suggest an early segregation of cardiogenic lineages in Xenopus laevis. Dev. Biol. 2009, 334, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Cleaver, O.; Krieg, P.A. Expression from DNA injected into Xenopus embryos. Methods Mol. Biol. 1999, 127, 133–153. [Google Scholar] [PubMed]
- Tandon, P.; Showell, C.; Christine, K.; Conlon, F.L. Morpholino injection in xenopus. Methods Mol. Biol. 2012, 843, 29–46. [Google Scholar] [PubMed]
- Summerton, J. Morpholino antisense oligomers: The case for an RNase H-independent structural type. Biochim. Biophys. Acta 1999, 1489, 141–158. [Google Scholar] [CrossRef]
- Summerton, J.E. Morpholino, sirna, and s-DNA compared: Impact of structure and mechanism of action on off-target effects and sequence specificity. Curr. Top Med. Chem. 2007, 7, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.; De Robertis, E.M.; Wallingford, J.B.; Niehrs, C. Morpholinos: Antisense and sensibility. Dev. Cell 2015, 35, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Kaltenbrun, E.; Tandon, P.; Amin, N.M.; Waldron, L.; Showell, C.; Conlon, F.L. Xenopus: An emerging model for studying congenital heart disease. Birth Defects Res. A 2011, 91, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Warkman, A.S.; Krieg, P.A. Xenopus as a model system for vertebrate heart development. Semin. Cell Dev. Biol. 2007, 18, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Kontarakis, Z.; Gerri, C.; Nolte, H.; Holper, S.; Kruger, M.; Stainier, D.Y. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature 2015, 524, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Kok, F.O.; Shin, M.; Ni, C.W.; Gupta, A.; Grosse, A.S.; van Impel, A.; Kirchmaier, B.C.; Peterson-Maduro, J.; Kourkoulis, G.; Male, I.; et al. Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish. Dev. Cell 2015, 32, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Merker, S.; Stainier, D.Y. Out with the old, in with the new: Reassessing morpholino knockdowns in light of genome editing technology. Development 2014, 141, 3103–3104. [Google Scholar] [CrossRef] [PubMed]
- Eisen, J.S.; Smith, J.C. Controlling morpholino experiments: Don't stop making antisense. Development 2008, 135, 1735–1743. [Google Scholar] [CrossRef] [PubMed]
- Afouda, B.A.; Hoppler, S. Xenopus explants as an experimental model system for studying heart development. Trends Cardiovasc. Med. 2009, 19, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Afouda, B.A.; Martin, J.; Liu, F.; Ciau-Uitz, A.; Patient, R.; Hoppler, S. GATA transcription factors integrate wnt signalling during heart development. Development 2008, 135, 3185–3190. [Google Scholar] [CrossRef] [PubMed]
- Foley, A.C.; Mercola, M. Heart induction by wnt antagonists depends on the homeodomain transcription factor hex. Genes dev. 2005, 19, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Schneider, V.A.; Mercola, M. Wnt antagonism initiates cardiogenesis in Xenopus laevis. Genes Dev. 2001, 15, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Sater, A.K.; Jacobson, A.G. The role of the dorsal lip in the induction of heart mesoderm in Xenopus laevis. Development 1990, 108, 461–470. [Google Scholar] [PubMed]
- Pandur, P.; Lasche, M.; Eisenberg, L.M.; Kuhl, M. Wnt-11 activation of a non-canonical wnt signalling pathway is required for cardiogenesis. Nature 2002, 418, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Etkin, L.D.; Pearman, B. Distribution, expression and germ line transmission of exogenous DNA sequences following microinjection into Xenopus laevis eggs. Development 1987, 99, 15–23. [Google Scholar] [PubMed]
- Takagi, C.; Sakamaki, K.; Morita, H.; Hara, Y.; Suzuki, M.; Kinoshita, N.; Ueno, N. Transgenic Xenopus laevis for live imaging in cell and developmental biology. Dev. Growth Differ. 2013, 55, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, D.; Marfo, C.A.; Li, D.; Lane, M.; Khokha, M.K. Crispr/cas9: An inexpensive, efficient loss of function tool to screen human disease genes in xenopus. Dev. Biol. 2015, 408, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Tandon, P.; Conlon, F.; Furlow, J.D.; Horb, M.E. Expanding the genetic toolkit in xenopus: Approaches and opportunities for human disease modeling. Dev. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, H.L.; Escalera, R.B., 2nd; Patel, S.S.; Wedemeyer, E.W.; Volk, K.A.; Lohr, J.L.; Reinking, B.E. Echocardiographic assessment of cardiac morphology and function in xenopus. Comp. Med. 2010, 60, 107–113. [Google Scholar] [PubMed]
- Sater, A.K.; Jacobson, A.G. The specification of heart mesoderm occurs during gastrulation in Xenopus laevis. Development 1989, 105, 821–830. [Google Scholar] [PubMed]
- Kolker, S.J.; Tajchman, U.; Weeks, D.L. Confocal imaging of early heart development in Xenopus laevis. Dev. Biol. 2000, 218, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Mohun, T.J.; Leong, L.M.; Weninger, W.J.; Sparrow, D.B. The morphology of heart development in Xenopus laevis. Dev. Biol. 2000, 218, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Nascone, N.; Mercola, M. An inductive role for the endoderm in xenopus cardiogenesis. Development 1995, 121, 515–523. [Google Scholar] [PubMed]
- Lawson, K.A.; Meneses, J.J.; Pedersen, R.A. Clonal analysis of epiblast fate during germ layer formation in the mouse embryo. Development 1991, 113, 891–911. [Google Scholar] [PubMed]
- Tam, P.P.; Parameswaran, M.; Kinder, S.J.; Weinberger, R.P. The allocation of epiblast cells to the embryonic heart and other mesodermal lineages: The role of ingression and tissue movement during gastrulation. Development 1997, 124, 1631–1642. [Google Scholar] [PubMed]
- Buckingham, M.; Meilhac, S.; Zaffran, S. Building the mammalian heart from two sources of myocardial cells. Nat. Rev. Genet. 2005, 6, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Engleka, K.A.; Manderfield, L.J.; Brust, R.D.; Li, L.; Cohen, A.; Dymecki, S.M.; Epstein, J.A. Islet1 derivatives in the heart are of both neural crest and second heart field origin. Circ. Res. 2012, 110, 922–926. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, T.X.; Lee, K.J.; Chien, K.R. Positional specification of ventricular myosin light chain 2 expression in the primitive murine heart tube. Proc. Natl. Acad. Sci. USA 1993, 90, 5157–5161. [Google Scholar] [CrossRef] [PubMed]
- Meilhac, S.M.; Esner, M.; Kelly, R.G.; Nicolas, J.F.; Buckingham, M.E. The clonal origin of myocardial cells in different regions of the embryonic mouse heart. Dev. Cell 2004, 6, 685–698. [Google Scholar] [CrossRef]
- Kelly, R.G.; Brown, N.A.; Buckingham, M.E. The arterial pole of the mouse heart forms from fgf10-expressing cells in pharyngeal mesoderm. Dev. Cell 2001, 1, 435–440. [Google Scholar] [CrossRef]
- Kelly, R.G.; Buckingham, M.E. The anterior heart-forming field: Voyage to the arterial pole of the heart. Trends Genet. 2002, 18, 210–216. [Google Scholar] [CrossRef]
- Zaffran, S.; Kelly, R.G.; Meilhac, S.M.; Buckingham, M.E.; Brown, N.A. Right ventricular myocardium derives from the anterior heart field. Circ. Res. 2004, 95, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Brade, T.; Gessert, S.; Kuhl, M.; Pandur, P. The amphibian second heart field: Xenopus islet-1 is required for cardiovascular development. Dev. Biol. 2007, 311, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Pandur, P.; Sirbu, I.O.; Kuhl, S.J.; Philipp, M.; Kuhl, M. Islet1-expressing cardiac progenitor cells: A comparison across species. Dev. Genes Evol. 2013, 223, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, F.; Bundschu, K.; Kuhl, S.J.; Kuhl, M. Tbx5 overexpression favors a first heart field lineage in murine embryonic stem cells and in Xenopus laevis embryos. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2011, 240, 2634–2645. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.P. Patterning the vertebrate heart. Nat. Rev. Genet. 2002, 3, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Laugwitz, K.L.; Moretti, A.; Caron, L.; Nakano, A.; Chien, K.R. Islet1 cardiovascular progenitors: A single source for heart lineages? Development 2008, 135, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Abu-Issa, R.; Kirby, M.L. Heart field: From mesoderm to heart tube. Annu. Rev. Cell Dev. Biol. 2007, 23, 45–68. [Google Scholar] [CrossRef] [PubMed]
- Lohr, J.L.; Yost, H.J. Vertebrate model systems in the study of early heart development: Xenopus and zebrafish. Am. J. Med. Genet. 2000, 97, 248–257. [Google Scholar] [CrossRef]
- Kowalski, W.J.; Pekkan, K.; Tinney, J.P.; Keller, B.B. Investigating developmental cardiovascular biomechanics and the origins of congenital heart defects. Front. Physiol. 2014, 5, 408. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.; Samtani, R.; Dhanantwari, P.; Lee, E.; Yamada, S.; Shiota, K.; Donofrio, M.T.; Leatherbury, L.; Lo, C.W. A detailed comparison of mouse and human cardiac development. Pediatr. Res. 2014, 76, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Schoenwolf, G.C.; Bleyl, S.B.; Brauer, P.R.; Francis-West, P.H. Development of the Heart. In Larsen’s Human Embryology, 5th ed.; Churchill Livingstone, an imprint of Elsevier Inc.: Philadelphia, PA, USA, 2015; Chapter 12; pp. 267–303. [Google Scholar]
- Martinsen, B.J. Reference guide to the stages of chick heart embryology. Dev. Dyn. 2005, 233, 1217–1237. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D. Genetic regulation of cardiogenesis and congenital heart disease. Annu. Rev. Pathol. 2006, 1, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.I. Incidence of congenital heart disease: II. Prenatal incidence. Pediatr. Cardiol. 1995, 16, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.I. Incidence of congenital heart disease: I. Postnatal incidence. Pediatr. Cardiol. 1995, 16, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.W.; Silberbach, G.M.; Kavanaugh-McHugh, A.; Cottrill, C.; Zhang, Y.; Riggs, S.; Smalls, O.; Johnson, M.C.; Watson, M.S.; Seidman, J.G.; et al. Mutations in the cardiac transcription factor nkx2.5 affect diverse cardiac developmental pathways. J. Clin. Investig. 1999, 104, 1567–1573. [Google Scholar] [CrossRef] [PubMed]
- Schott, J.J.; Benson, D.W.; Basson, C.T.; Pease, W.; Silberbach, G.M.; Moak, J.P.; Maron, B.J.; Seidman, C.E.; Seidman, J.G. Congenital heart disease caused by mutations in the transcription factor nkx2–5. Science 1998, 281, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Durocher, D.; Charron, F.; Warren, R.; Schwartz, R.J.; Nemer, M. The cardiac transcription factors Nkx2–5 and GATA-4 are mutual cofactors. EMBO J. 1997, 16, 5687–5696. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Shioi, T.; Kasahara, H.; Jobe, S.M.; Wiese, R.J.; Markham, B.E.; Izumo, S. The cardiac tissue-restricted homeobox protein Csx/Nkx2.5 physically associates with the zinc finger protein GATA4 and cooperatively activates atrial natriuretic factor gene expression. Mol. Cell. Biol. 1998, 18, 3120–3129. [Google Scholar] [CrossRef] [PubMed]
- Pehlivan, T.; Pober, B.R.; Brueckner, M.; Garrett, S.; Slaugh, R.; Van Rheeden, R.; Wilson, D.B.; Watson, M.S.; Hing, A.V. Gata4 haploinsufficiency in patients with interstitial deletion of chromosome region 8p23.1 and congenital heart disease. Am. J. Med. Genet. 1999, 83, 201–206. [Google Scholar] [CrossRef]
- Sepulveda, J.L.; Belaguli, N.; Nigam, V.; Chen, C.Y.; Nemer, M.; Schwartz, R.J. GATA-4 and Nkx-2.5 coactivate Nkx-2 DNA binding targets: Role for regulating early cardiac gene expression. Mol. Cell. Biol. 1998, 18, 3405–3415. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, H.L.; Sutherland, L.; Kolker, S.J.; Welp, C.; Tajchman, U.; Desmarais, V.; Weeks, D.L. Transient early embryonic expression of Nkx2-5 mutations linked to congenital heart defects in human causes heart defects in Xenopus laevis. Dev. Dyn. 2007, 236, 2475–2484. [Google Scholar] [CrossRef] [PubMed]
- Haworth, K.E.; Kotecha, S.; Mohun, T.J.; Latinkic, B.V. GATA4 and GATA5 are essential for heart and liver development in Xenopus embryos. BMC Dev. Biol. 2008, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Peterkin, T.; Gibson, A.; Patient, R. Redundancy and evolution of gata factor requirements in development of the myocardium. Dev. Biol. 2007, 311, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Maitra, M.; Schluterman, M.K.; Nichols, H.A.; Richardson, J.A.; Lo, C.W.; Srivastava, D.; Garg, V. Interaction of Gata4 and Gata6 with Tbx5 is critical for normal cardiac development. Dev. Biol. 2009, 326, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Basson, C.T.; Cowley, G.S.; Solomon, S.D.; Weissman, B.; Poznanski, A.K.; Traill, T.A.; Seidman, J.G.; Seidman, C.E. The clinical and genetic spectrum of the holt-oram syndrome (heart-hand syndrome). N. Engl. J. Med. 1994, 330, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Cross, S.J.; Ching, Y.H.; Li, Q.Y.; Armstrong-Buisseret, L.; Spranger, S.; Lyonnet, S.; Bonnet, D.; Penttinen, M.; Jonveaux, P.; Leheup, B.; et al. The mutation spectrum in holt-oram syndrome. J. Med. Genet. 2000, 37, 785–787. [Google Scholar] [CrossRef] [PubMed]
- Holt, M.; Oram, S. Familial heart disease with skeletal malformations. Br. Heart J. 1960, 22, 236–242. [Google Scholar] [CrossRef] [PubMed]
- McDermott, D.A.; Bressan, M.C.; He, J.; Lee, J.S.; Aftimos, S.; Brueckner, M.; Gilbert, F.; Graham, G.E.; Hannibal, M.C.; Innis, J.W.; et al. Tbx5 genetic testing validates strict clinical criteria for holt-oram syndrome. Pediatr. Res. 2005, 58, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.D.; Martz, S.N.; Binder, O.; Goetz, S.C.; Price, B.M.; Smith, J.C.; Conlon, F.L. Tbx5 and tbx20 act synergistically to control vertebrate heart morphogenesis. Development 2005, 132, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Goetz, S.C.; Brown, D.D.; Conlon, F.L. Tbx5 is required for embryonic cardiac cell cycle progression. Development 2006, 133, 2575–2584. [Google Scholar] [CrossRef] [PubMed]
- Horb, M.E.; Thomsen, G.H. Tbx5 is essential for heart development. Development 1999, 126, 1739–1751. [Google Scholar] [PubMed]
- Liu, J.J.; Fan, L.L.; Chen, J.L.; Tan, Z.P.; Yang, Y.F. A novel variant in tbx20 (p.D176n) identified by whole-exome sequencing in combination with a congenital heart disease related gene filter is associated with familial atrial septal defect. J. Zhejiang Univ. Sci. B 2014, 15, 830–837. [Google Scholar] [CrossRef] [PubMed]
- Kirk, E.P.; Sunde, M.; Costa, M.W.; Rankin, S.A.; Wolstein, O.; Castro, M.L.; Butler, T.L.; Hyun, C.; Guo, G.; Otway, R.; et al. Mutations in cardiac t-box factor gene tbx20 are associated with diverse cardiac pathologies, including defects of septation and valvulogenesis and cardiomyopathy. Am. J. Hum. Genet. 2007, 81, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Shen, A.; Li, X.; Jiao, W.; Zhang, X.; Li, Z. T-box transcription factor tbx20 mutations in chinese patients with congenital heart disease. Eur. J. Med. Genet. 2008, 51, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Mohapatra, B.; Akasaka, T.; Liu, J.; Ocorr, K.; Towbin, J.A.; Bodmer, R. Transcription factor neuromancer/tbx20 is required for cardiac function in drosophila with implications for human heart disease. Proc. Natl. Acad. Sci. USA 2008, 105, 19833–19838. [Google Scholar] [CrossRef] [PubMed]
- Posch, M.G.; Gramlich, M.; Sunde, M.; Schmitt, K.R.; Lee, S.H.; Richter, S.; Kersten, A.; Perrot, A.; Panek, A.N.; Al Khatib, I.H.; et al. A gain-of-function tbx20 mutation causes congenital atrial septal defects, patent foramen ovale and cardiac valve defects. J. Med. Genet. 2010, 47, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Hammer, S.; Toenjes, M.; Lange, M.; Fischer, J.J.; Dunkel, I.; Mebus, S.; Grimm, C.H.; Hetzer, R.; Berger, F.; Sperling, S. Characterization of tbx20 in human hearts and its regulation by tfap2. J. Cell Biochem. 2008, 104, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Stennard, F.A.; Costa, M.W.; Elliott, D.A.; Rankin, S.; Haast, S.J.; Lai, D.; McDonald, L.P.; Niederreither, K.; Dolle, P.; Bruneau, B.G.; et al. Cardiac t-box factor tbx20 directly interacts with nkx2–5, gata4, and gata5 in regulation of gene expression in the developing heart. Dev. Biol. 2003, 262, 206–224. [Google Scholar] [CrossRef]
- Baldini, A. Digeorge syndrome: An update. Curr. Opin. Cardiol. 2004, 19, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Momma, K. Cardiovascular anomalies associated with chromosome 22q11.2 deletion syndrome. Am. J. Cardiol. 2010, 105, 1617–1624. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, H.; Srivastava, D. Unraveling the genetic and developmental mysteries of 22q11 deletion syndrome. Trends Mol. Med. 2003, 9, 383–389. [Google Scholar] [CrossRef]
- Yagi, H.; Furutani, Y.; Hamada, H.; Sasaki, T.; Asakawa, S.; Minoshima, S.; Ichida, F.; Joo, K.; Kimura, M.; Imamura, S.; et al. Role of tbx1 in human del22q11.2 syndrome. Lancet 2003, 362, 1366–1373. [Google Scholar] [CrossRef]
- Ataliotis, P.; Ivins, S.; Mohun, T.J.; Scambler, P.J. XTbx1 is a transcriptional activator involved in head and pharyngeal arch development in Xenopus laevis. Dev. Dyn. 2005, 232, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.J.; Ataliotis, P.; Kotecha, S.; Towers, N.; Sparrow, D.B.; Mohun, T.J. The mlc1v gene provides a transgenic marker of myocardium formation within developing chambers of the xenopus heart. Dev. Dyn. 2005, 232, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Grossfeld, P.D.; Mattina, T.; Lai, Z.; Favier, R.; Jones, K.L.; Cotter, F.; Jones, C. The 11q terminal deletion disorder: A prospective study of 110 cases. Am. J. Med. Genet. A 2004, 129A, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, P.; Hauge, M.; Henningsen, K.; Hobolth, N.; Mikkelsen, M.; Philip, J. An (11;21) translocation in four generations with chromosome 11 abnormalities in the offspring. A clinical, cytogenetical, and gene marker study. Hum. Hered. 1973, 23, 568–585. [Google Scholar] [CrossRef] [PubMed]
- Penny, L.A.; Dell’Aquila, M.; Jones, M.C.; Bergoffen, J.; Cunniff, C.; Fryns, J.P.; Grace, E.; Graham, J.M., Jr.; Kousseff, B.; Mattina, T.; et al. Clinical and molecular characterization of patients with distal 11q deletions. Am. J. Hum. Genet. 1995, 56, 676–683. [Google Scholar] [PubMed]
- Tunnacliffe, A.; Jones, C.; Le Paslier, D.; Todd, R.; Cherif, D.; Birdsall, M.; Devenish, L.; Yousry, C.; Cotter, F.E.; James, M.R. Localization of jacobsen syndrome breakpoints on a 40-mb physical map of distal chromosome 11q. Genome Res. 1999, 9, 44–52. [Google Scholar] [PubMed]
- Ye, M.; Coldren, C.; Liang, X.; Mattina, T.; Goldmuntz, E.; Benson, D.W.; Ivy, D.; Perryman, M.B.; Garrett-Sinha, L.A.; Grossfeld, P. Deletion of ets-1, a gene in the jacobsen syndrome critical region, causes ventricular septal defects and abnormal ventricular morphology in mice. Hum. Mol. Genet. 2010, 19, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Nie, S.; Bronner, M.E. Dual developmental role of transcriptional regulator ets1 in xenopus cardiac neural crest vs. Heart mesoderm. Cardiovasc. Res. 2015, 106, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Lalani, S.R.; Ware, S.M.; Wang, X.; Zapata, G.; Tian, Q.; Franco, L.M.; Jiang, Z.; Bucasas, K.; Scott, D.A.; Campeau, P.M.; et al. Mctp2 is a dosage-sensitive gene required for cardiac outflow tract development. Hum. Mol. Genet. 2013, 22, 4339–4348. [Google Scholar] [CrossRef] [PubMed]
- Davenport, S.L.; Hefner, M.A.; Mitchell, J.A. The spectrum of clinical features in charge syndrome. Clin. Genet. 1986, 29, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Aramaki, M.; Udaka, T.; Kosaki, R.; Makita, Y.; Okamoto, N.; Yoshihashi, H.; Oki, H.; Nanao, K.; Moriyama, N.; Oku, S.; et al. Phenotypic spectrum of charge syndrome with chd7 mutations. J. Pediatr. 2006, 148, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Jongmans, M.C.; Admiraal, R.J.; van der Donk, K.P.; Vissers, L.E.; Baas, A.F.; Kapusta, L.; van Hagen, J.M.; Donnai, D.; de Ravel, T.J.; Veltman, J.A.; et al. Charge syndrome: The phenotypic spectrum of mutations in the chd7 gene. J. Med. Genet. 2006, 43, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Lalani, S.R.; Safiullah, A.M.; Fernbach, S.D.; Harutyunyan, K.G.; Thaller, C.; Peterson, L.E.; McPherson, J.D.; Gibbs, R.A.; White, L.D.; Hefner, M.; et al. Spectrum of chd7 mutations in 110 individuals with charge syndrome and genotype-phenotype correlation. Am. J. Hum. Genet. 2006, 78, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Sanlaville, D.; Etchevers, H.C.; Gonzales, M.; Martinovic, J.; Clement-Ziza, M.; Delezoide, A.L.; Aubry, M.C.; Pelet, A.; Chemouny, S.; Cruaud, C.; et al. Phenotypic spectrum of charge syndrome in fetuses with chd7 truncating mutations correlates with expression during human development. J. Med. Genet. 2006, 43, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Vissers, L.E.; van Ravenswaaij, C.M.; Admiraal, R.; Hurst, J.A.; de Vries, B.B.; Janssen, I.M.; van der Vliet, W.A.; Huys, E.H.; de Jong, P.J.; Hamel, B.C.; et al. Mutations in a new member of the chromodomain gene family cause charge syndrome. Nat. Genet. 2004, 36, 955–957. [Google Scholar] [CrossRef] [PubMed]
- Wincent, J.; Holmberg, E.; Stromland, K.; Soller, M.; Mirzaei, L.; Djureinovic, T.; Robinson, K.; Anderlid, B.; Schoumans, J. Chd7 mutation spectrum in 28 swedish patients diagnosed with charge syndrome. Clin. Genet. 2008, 74, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, R.; Chen, D.A.; Rada-Iglesias, A.; Zhang, J.; Xiong, Y.; Helms, J.; Chang, C.P.; Zhao, Y.; Swigut, T.; Wysocka, J. Chd7 cooperates with pbaf to control multipotent neural crest formation. Nature 2010, 463, 958–962. [Google Scholar] [CrossRef] [PubMed]
- Schulz, Y.; Wehner, P.; Opitz, L.; Salinas-Riester, G.; Bongers, E.M.; van Ravenswaaij-Arts, C.M.; Wincent, J.; Schoumans, J.; Kohlhase, J.; Borchers, A.; et al. Chd7, the gene mutated in charge syndrome, regulates genes involved in neural crest cell guidance. Hum. Genet. 2014, 133, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- McPherson, R.; Pertsemlidis, A.; Kavaslar, N.; Stewart, A.; Roberts, R.; Cox, D.R.; Hinds, D.A.; Pennacchio, L.A.; Tybjaerg-Hansen, A.; Folsom, A.R.; et al. A common allele on chromosome 9 associated with coronary heart disease. Science 2007, 316, 1488–1491. [Google Scholar] [CrossRef] [PubMed]
- Helgadottir, A.; Thorleifsson, G.; Manolescu, A.; Gretarsdottir, S.; Blondal, T.; Jonasdottir, A.; Jonasdottir, A.; Sigurdsson, A.; Baker, A.; Palsson, A.; et al. A common variant on chromosome 9p21 affects the risk of myocardial infarction. Science 2007, 316, 1491–1493. [Google Scholar] [CrossRef] [PubMed]
- Samani, N.J.; Erdmann, J.; Hall, A.S.; Hengstenberg, C.; Mangino, M.; Mayer, B.; Dixon, R.J.; Meitinger, T.; Braund, P.; Wichmann, H.E.; et al. Genomewide association analysis of coronary artery disease. N. Engl. J. Med. 2007, 357, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, N.C.; Azimi Nehzad, M.; El Shamieh, S.; Stathopoulou, M.G.; Visvikis-Siest, S. Cardiovascular diseases and genome-wide association studies. Clin. Chim. Acta 2011, 412, 1697–1701. [Google Scholar] [CrossRef] [PubMed]
- Fakhro, K.A.; Choi, M.; Ware, S.M.; Belmont, J.W.; Towbin, J.A.; Lifton, R.P.; Khokha, M.K.; Brueckner, M. Rare copy number variations in congenital heart disease patients identify unique genes in left-right patterning. Proc. Natl. Acad. Sci. USA 2011, 108, 2915–2920. [Google Scholar] [CrossRef] [PubMed]
- Greenway, S.C.; Pereira, A.C.; Lin, J.C.; DePalma, S.R.; Israel, S.J.; Mesquita, S.M.; Ergul, E.; Conta, J.H.; Korn, J.M.; McCarroll, S.A.; et al. De novo copy number variants identify new genes and loci in isolated sporadic tetralogy of fallot. Nat. Genet. 2009, 41, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.; Choi, M.; Wakimoto, H.; Ma, L.; Jiang, J.; Overton, J.D.; Romano-Adesman, A.; Bjornson, R.D.; Breitbart, R.E.; Brown, K.K.; et al. De novo mutations in histone-modifying genes in congenital heart disease. Nature 2013, 498, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Grammer, T.C.; Liu, K.J.; Mariani, F.V.; Harland, R.M. Use of large-scale expression cloning screens in the Xenopus laevis tadpole to identify gene function. Dev. Biol. 2000, 228, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Lundby, A.; Rossin, E.J.; Steffensen, A.B.; Acha, M.R.; Newton-Cheh, C.; Pfeufer, A.; Lynch, S.N.; Consortium, Q.T.I.I.G.; Olesen, S.P.; Brunak, S.; et al. Annotation of loci from genome-wide association studies using tissue-specific quantitative interaction proteomics. Nat. Methods 2014, 11, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Boskovski, M.T.; Yuan, S.; Pedersen, N.B.; Goth, C.K.; Makova, S.; Clausen, H.; Brueckner, M.; Khokha, M.K. The heterotaxy gene galnt11 glycosylates notch to orchestrate cilia type and laterality. Nature 2013, 504, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.; Ehret, G.B.; Rice, K.; Verwoert, G.C.; Launer, L.J.; Dehghan, A.; Glazer, N.L.; Morrison, A.C.; Johnson, A.D.; Aspelund, T.; et al. Genome-wide association study of blood pressure and hypertension. Nat. Genet. 2009, 41, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, F.; Isono, M.; Katsuya, T.; Yamamoto, K.; Yokota, M.; Sugiyama, T.; Nabika, T.; Fujioka, A.; Ohnaka, K.; Asano, H.; et al. Blood pressure and hypertension are associated with 7 loci in the japanese population. Circulation 2010, 121, 2302–2309. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, M.S.; Christine, K.S.; Amin, N.M.; Dorr, K.M.; Kushner, E.J.; Bautch, V.L.; Taylor, J.M.; Conlon, F.L. Casz1 promotes vascular assembly and morphogenesis through the direct regulation of an egfl7/rhoa-mediated pathway. Dev. Cell 2013, 25, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Christine, K.S.; Conlon, F.L. Vertebrate castor is required for differentiation of cardiac precursor cells at the ventral midline. Dev. Cell 2008, 14, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Amin, N.M.; Gibbs, D.; Conlon, F.L. Differential regulation of casz1 protein expression during cardiac and skeletal muscle development. Dev. Dyn. 2014, 243, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Sojka, S.; Amin, N.M.; Gibbs, D.; Christine, K.S.; Charpentier, M.S.; Conlon, F.L. Congenital heart disease protein 5 associates with casz1 to maintain myocardial tissue integrity. Development 2014, 141, 3040–3049. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Cardiovascular Event | Developmental Stage | ||||
|---|---|---|---|---|---|
| Danio Rerio | Xenopus Laevis | Mus Musculus | Gallus Gallus | Homo Sapiens | |
| cardiac progenitors | 5 hpf | NF10 | E6.5 | HH4 | CS7 |
| heart field specification | 12 hpf | NF12–NF14 | E7.0–E7.5 | HH5 | CS8 |
| migration to ventral midline | 12–19 hpf | NF12–NF13 | E7.5 | HH7–HH8 | CS9 |
| primary heart tube formation | 21–24 hpf | NF31–NF33 | E8.0 | HH9 | CS10 |
| onset of coordinated muscle contraction | 22 hpf | NF35 | E8.5 | HH10 | CS10 |
| cardiac looping | 30–36 hpf | NF33–NF36 | E8.5–E10.5 | HH10–HH24 | CS13–CS17 |
| onset of blood flow | 30 hpf | NF35 | E8.5 | HH10 | CS11 |
| chamber formation | 30 hpf | NF39–NF40 | E9.5–E12.5 | HH16/17 | CS12–CS15 |
| onset of ventricular trabeculaetion | 48 hpf | NF41 | E9.5 | HH16 | CS11 |
| valvulogenesis | 48 hpf | NF41–NF44 | E9.5 | HH21–HH36 | CS15–CS18 |
| atrial septation | none | NF44–NF45 | E10.0–E14.5 | HH16–HH36 | CS14–CS18 |
| ventricular septation | none | none | E9.0–E14.0 | HH17–HH34 | CS16–CS22 |
| Disease | Affected Genes | Xenopus Model | Cardiovascular Phenotype |
|---|---|---|---|
| Atrial Septal Defects (ASD) | gata4 | LOF | looping defects |
| nkx2-5 | GOF | cardiac conduction defects, ASD | |
| Axonfeld-Reiger Syndrome | pitx2 | GOF, LOF | looping defects and ASD |
| CHARGE Syndrome | chd7 | GOF, LOF | neural cest migration and OFT defects |
| DiGeorge Syndrome | tbx1 | GOF | looping defects |
| Holt-Oram Syndrome | tbx5 | GOF, LOF | looping defects, reduced cardiomyocytes |
| Jacobsen Syndrome | ets1 | LOF | OFT and aortic arch formation defects |
| LVOT obstructive defects, Hypoplastic left heart syndrome | mctp2 | GOF, LOF | looping defects, OFT defects |
| Tetralogy of Fallot | tbx20 | LOF | looping defects, reduced cardiomyocytes |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hempel, A.; Kühl, M. A Matter of the Heart: The African Clawed Frog Xenopus as a Model for Studying Vertebrate Cardiogenesis and Congenital Heart Defects. J. Cardiovasc. Dev. Dis. 2016, 3, 21. https://doi.org/10.3390/jcdd3020021
Hempel A, Kühl M. A Matter of the Heart: The African Clawed Frog Xenopus as a Model for Studying Vertebrate Cardiogenesis and Congenital Heart Defects. Journal of Cardiovascular Development and Disease. 2016; 3(2):21. https://doi.org/10.3390/jcdd3020021
Chicago/Turabian StyleHempel, Annemarie, and Michael Kühl. 2016. "A Matter of the Heart: The African Clawed Frog Xenopus as a Model for Studying Vertebrate Cardiogenesis and Congenital Heart Defects" Journal of Cardiovascular Development and Disease 3, no. 2: 21. https://doi.org/10.3390/jcdd3020021