Hyperglycemia Alters the Structure and Hemodynamics of the Developing Embryonic Heart

Abstract
:1. Introduction
2. Results
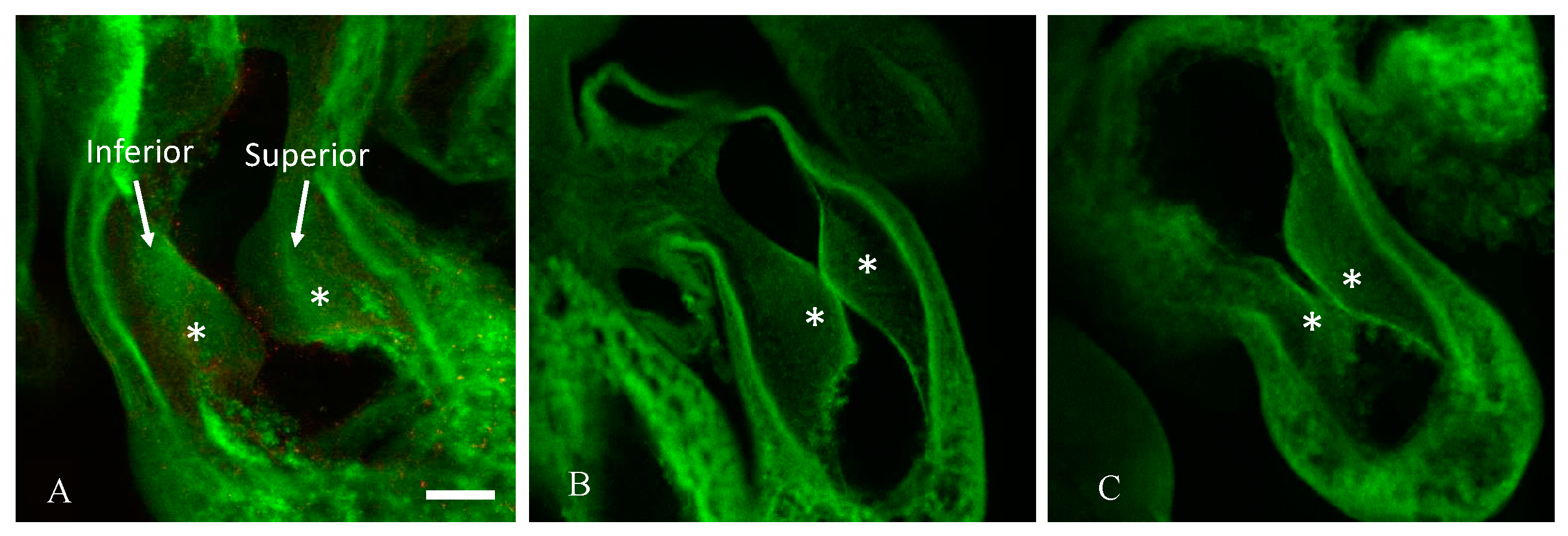
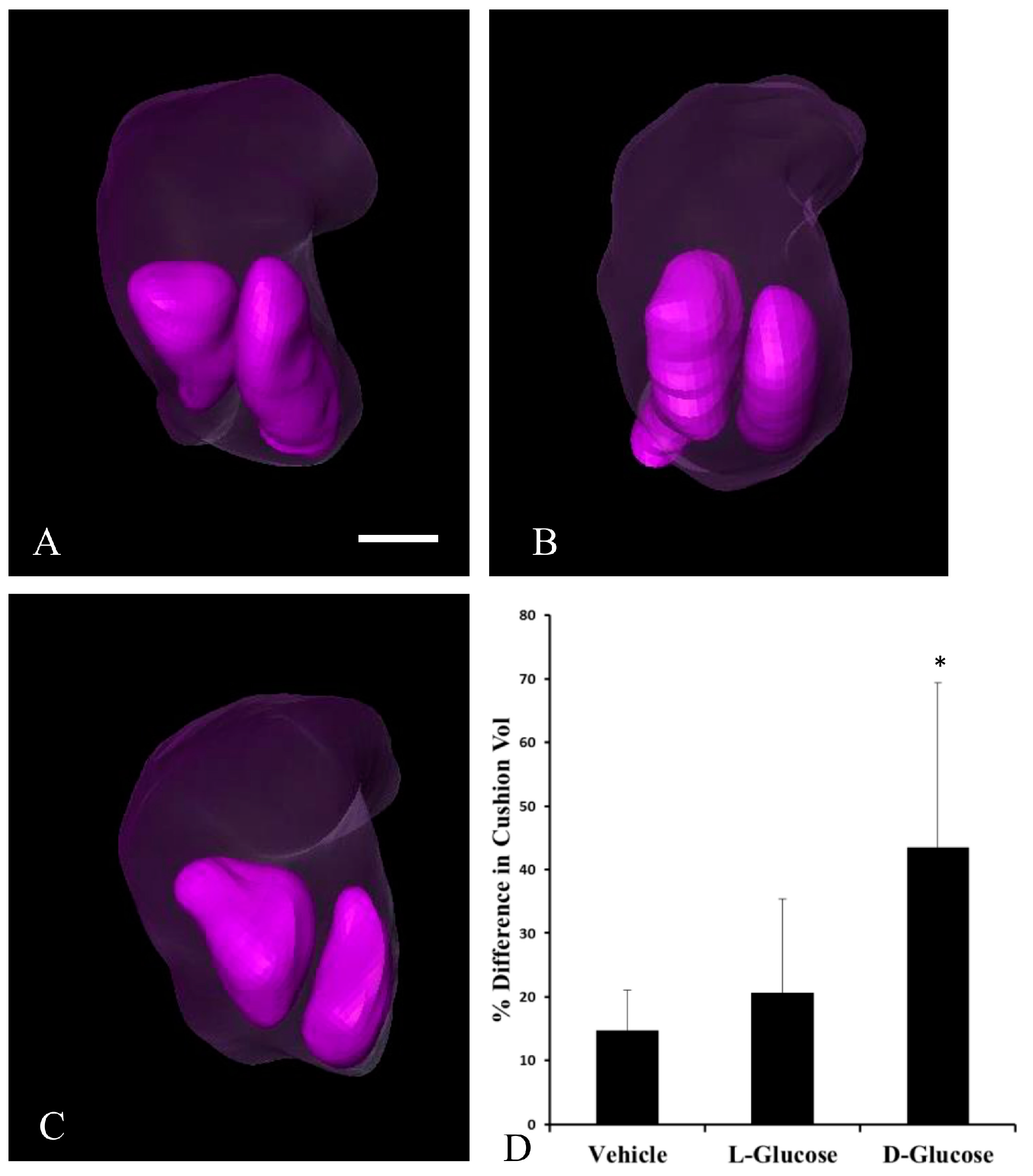
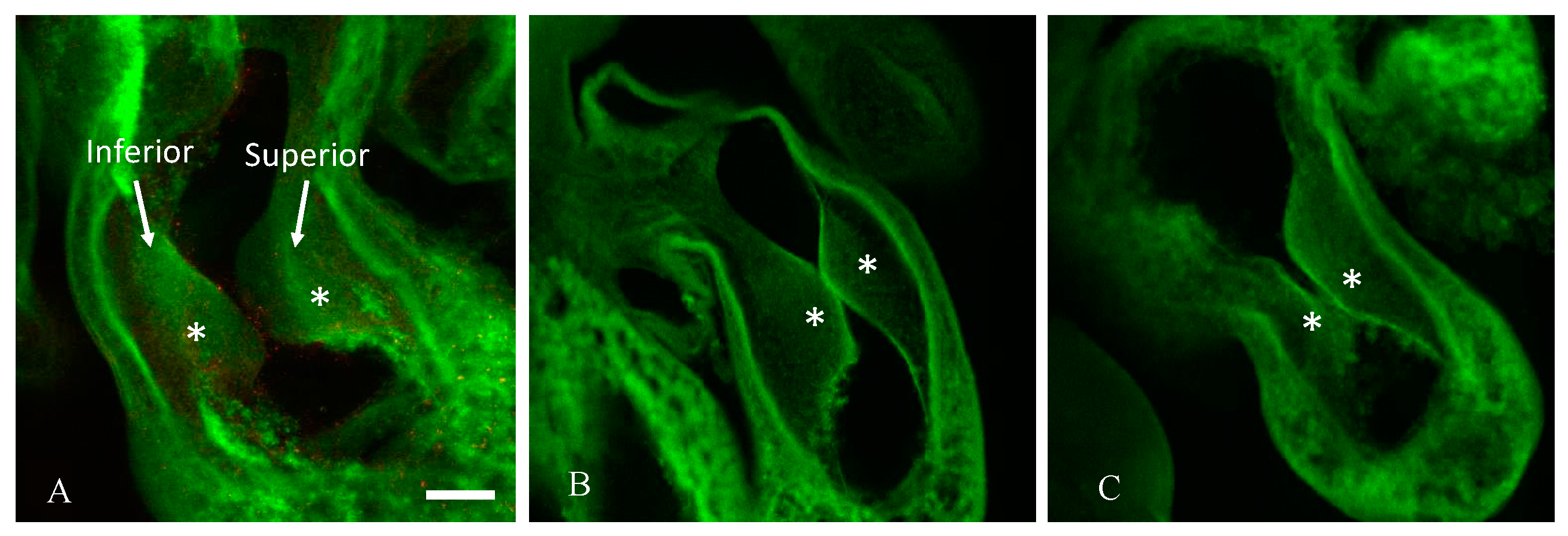
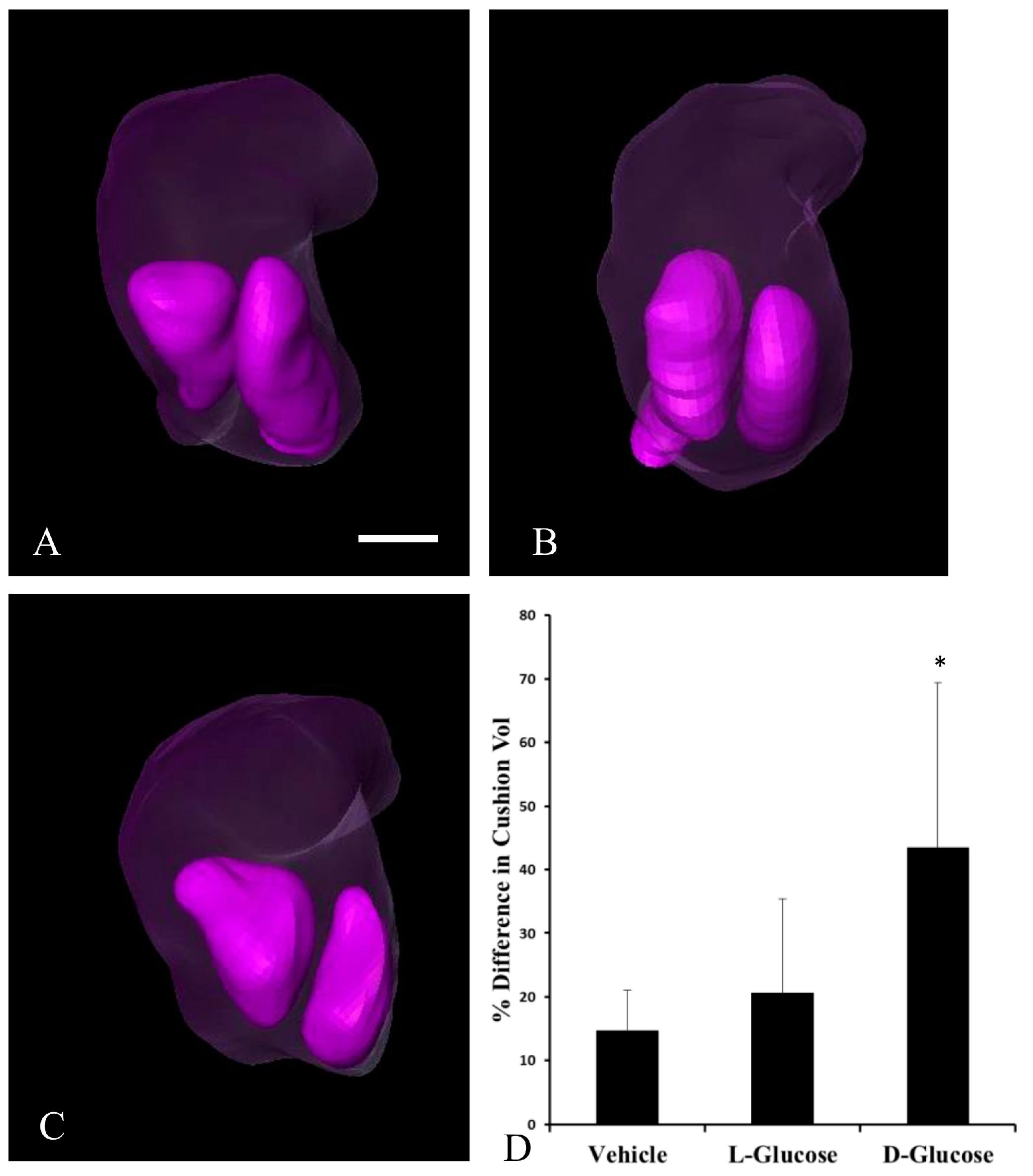
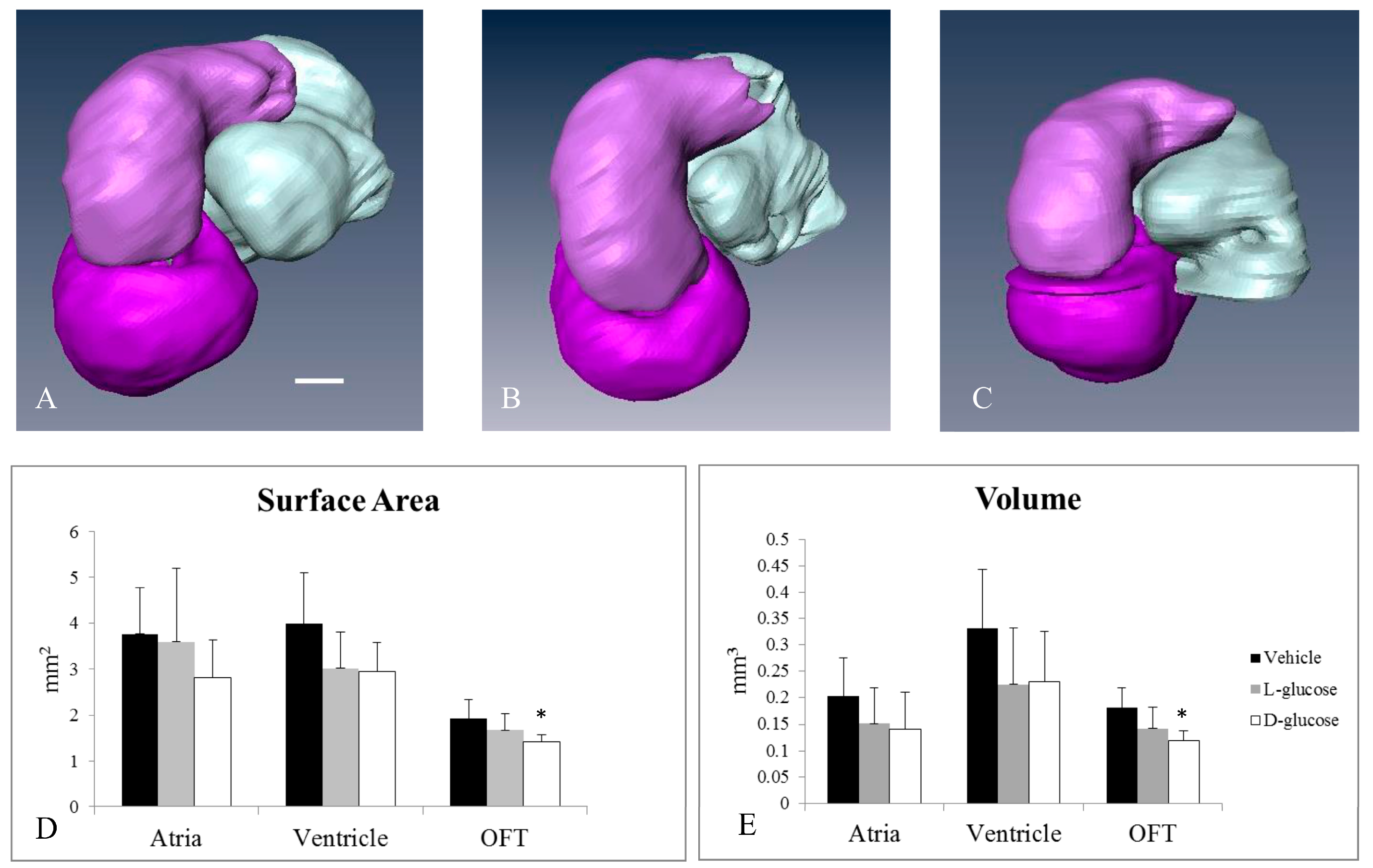
2.1. Hyperglycemia Causes Asymmetric Endocardial Cushions
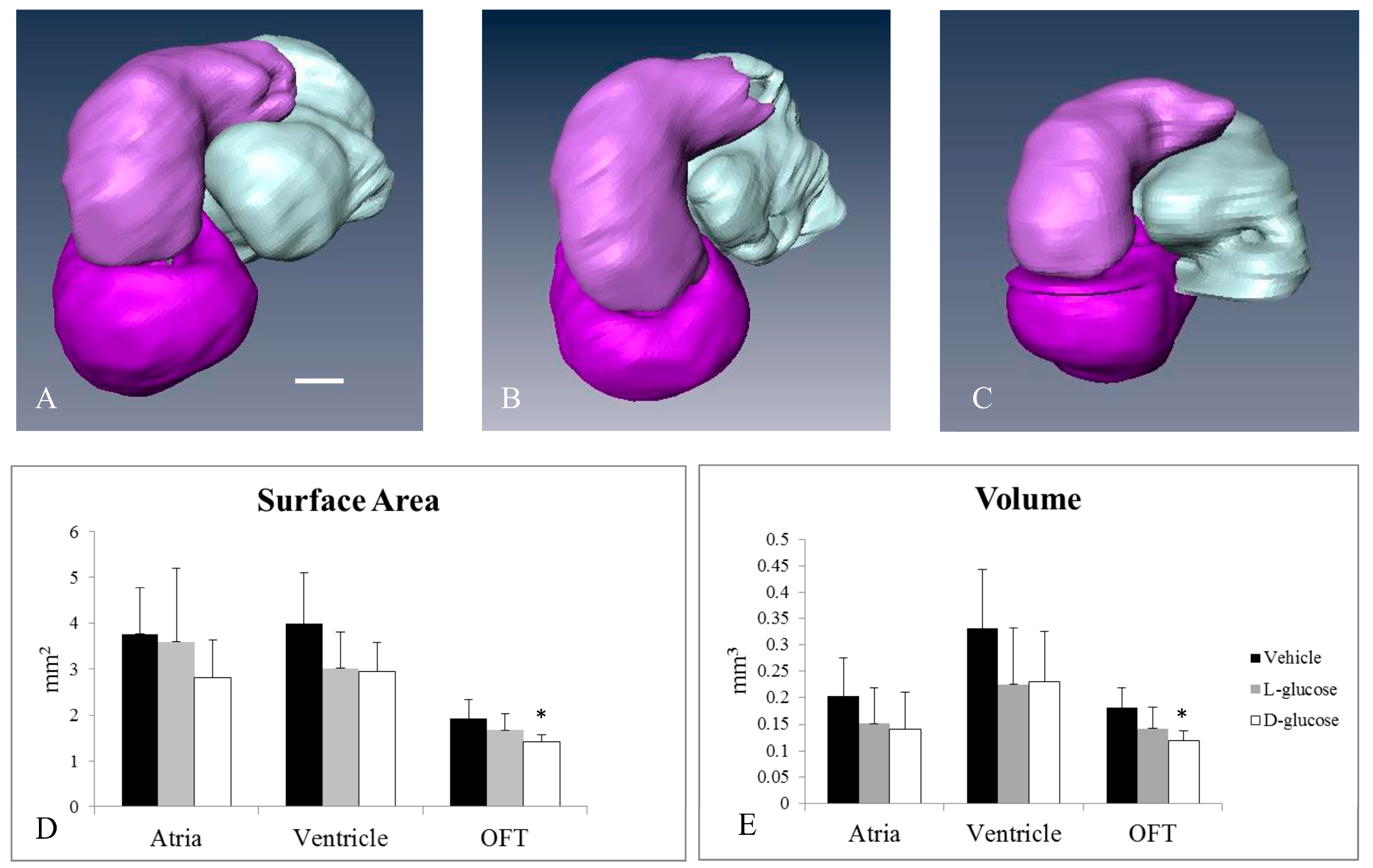
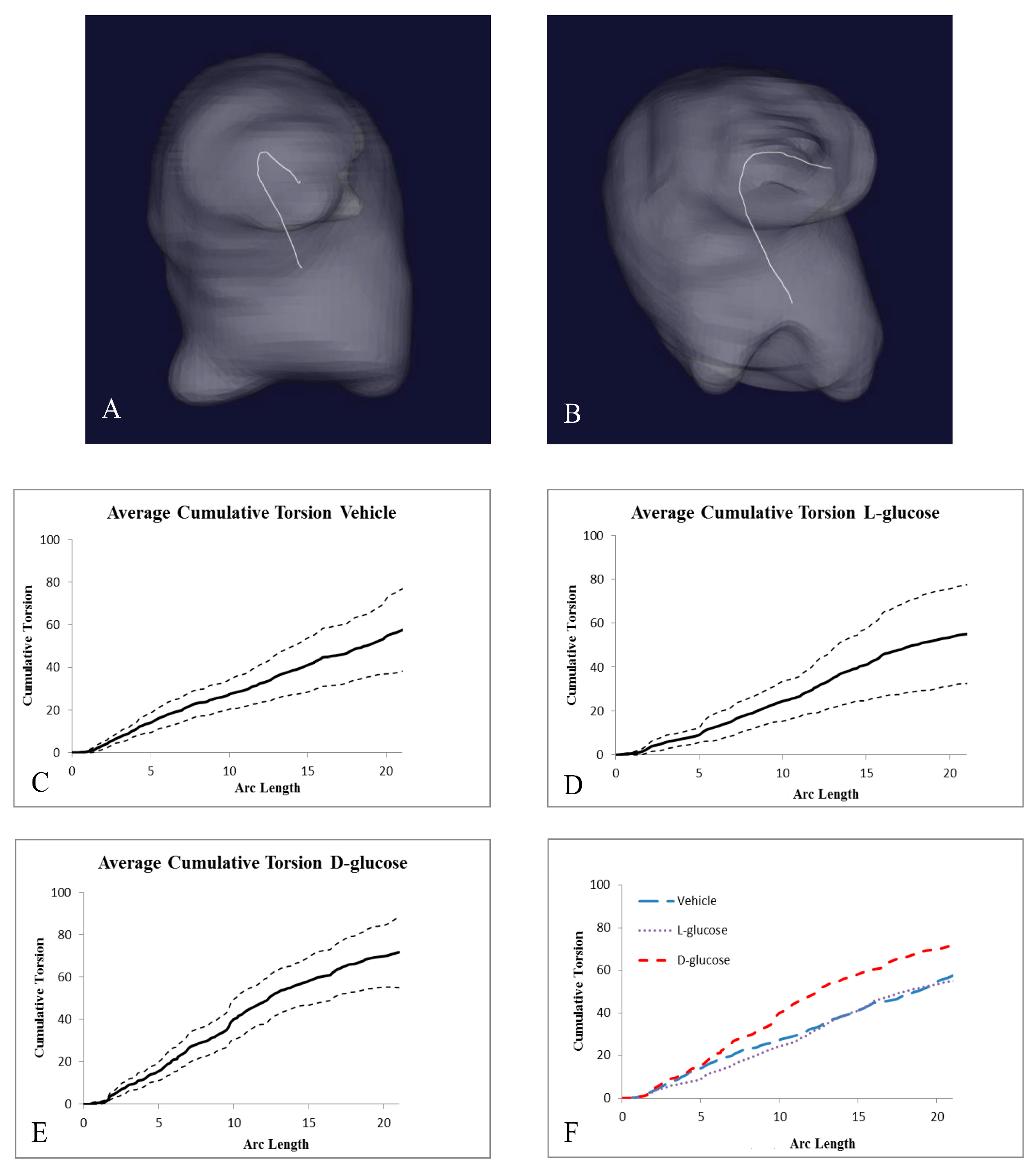
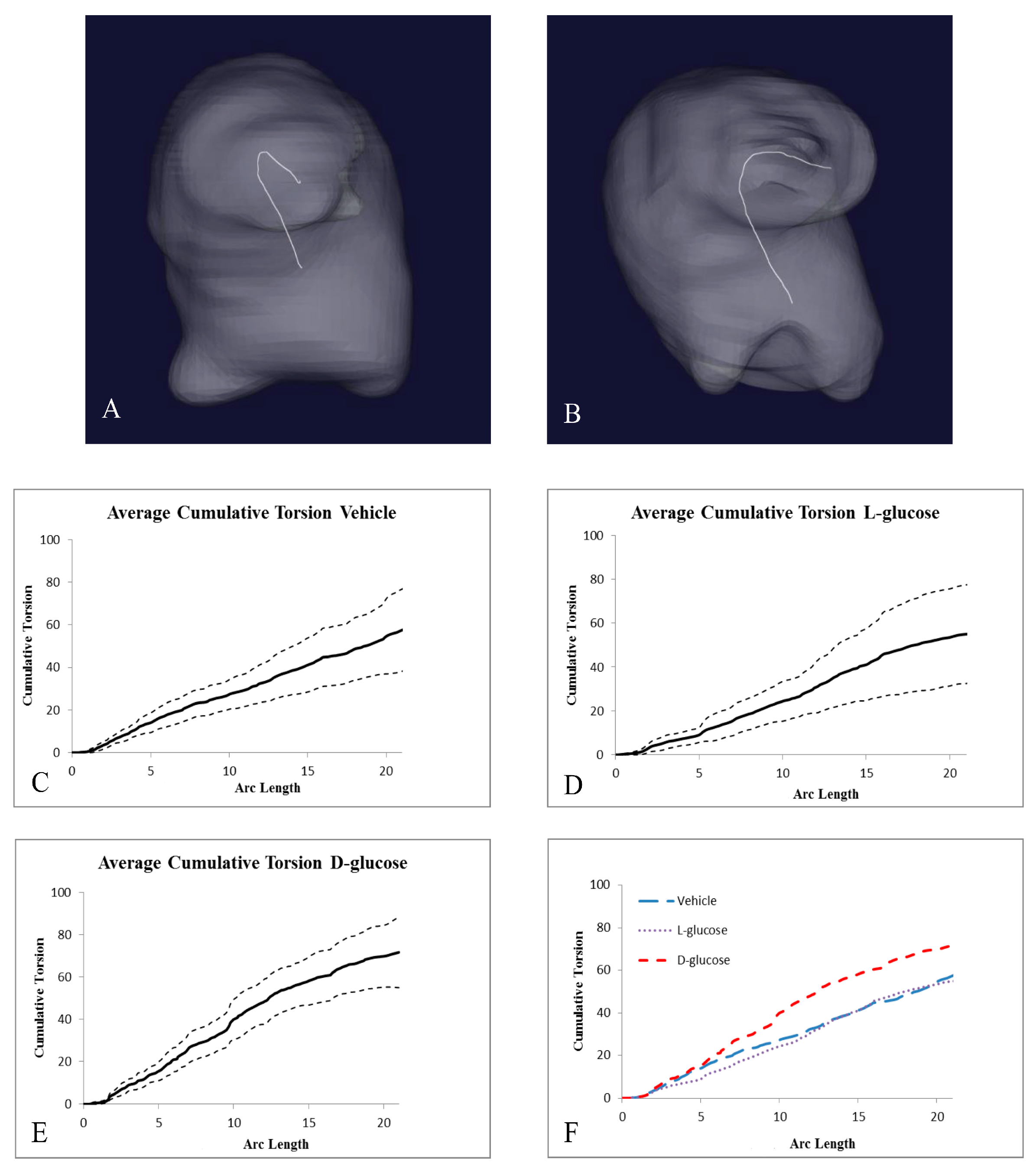
2.2. Hyperglycemia Alters the Curvature of the OFT
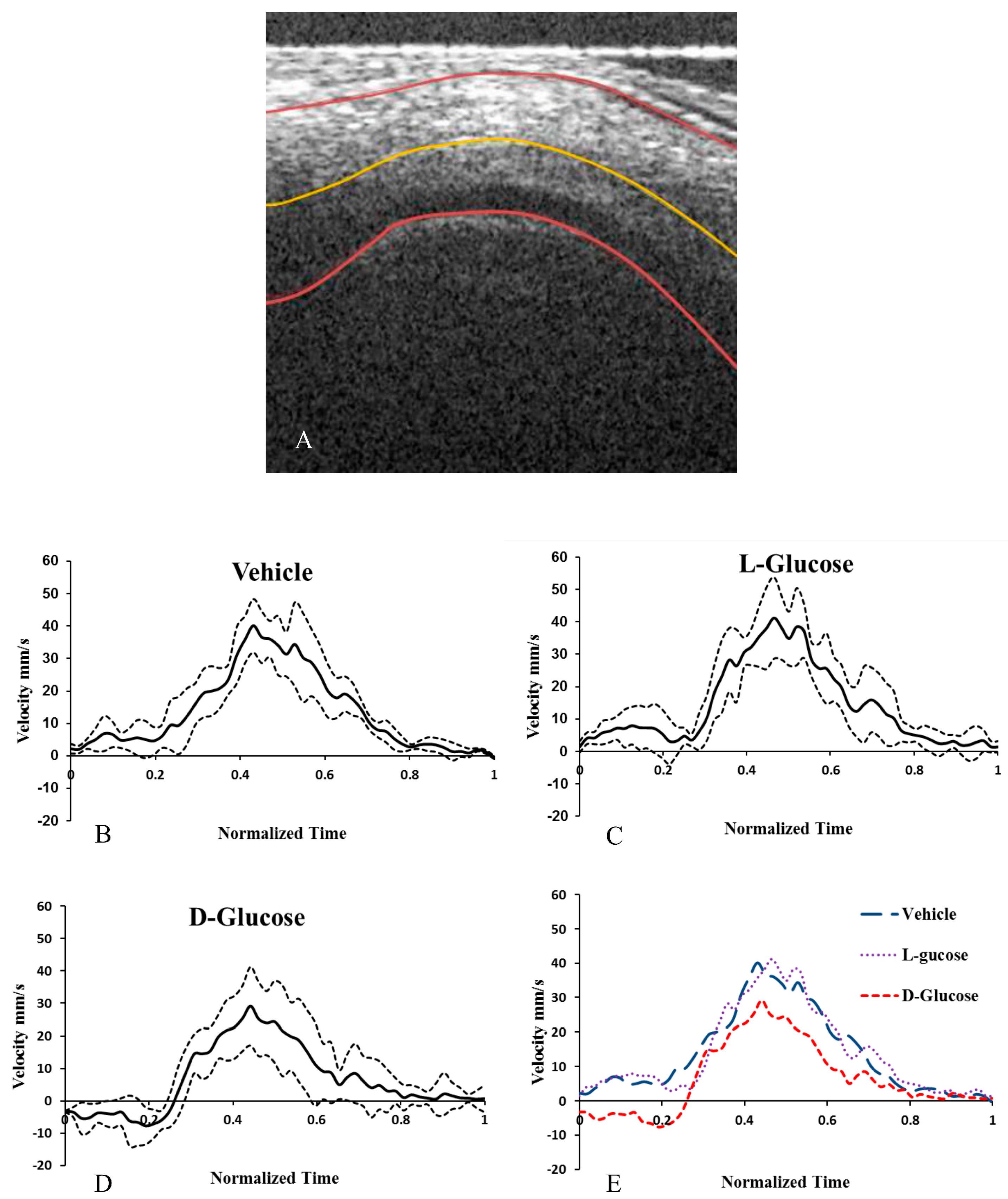
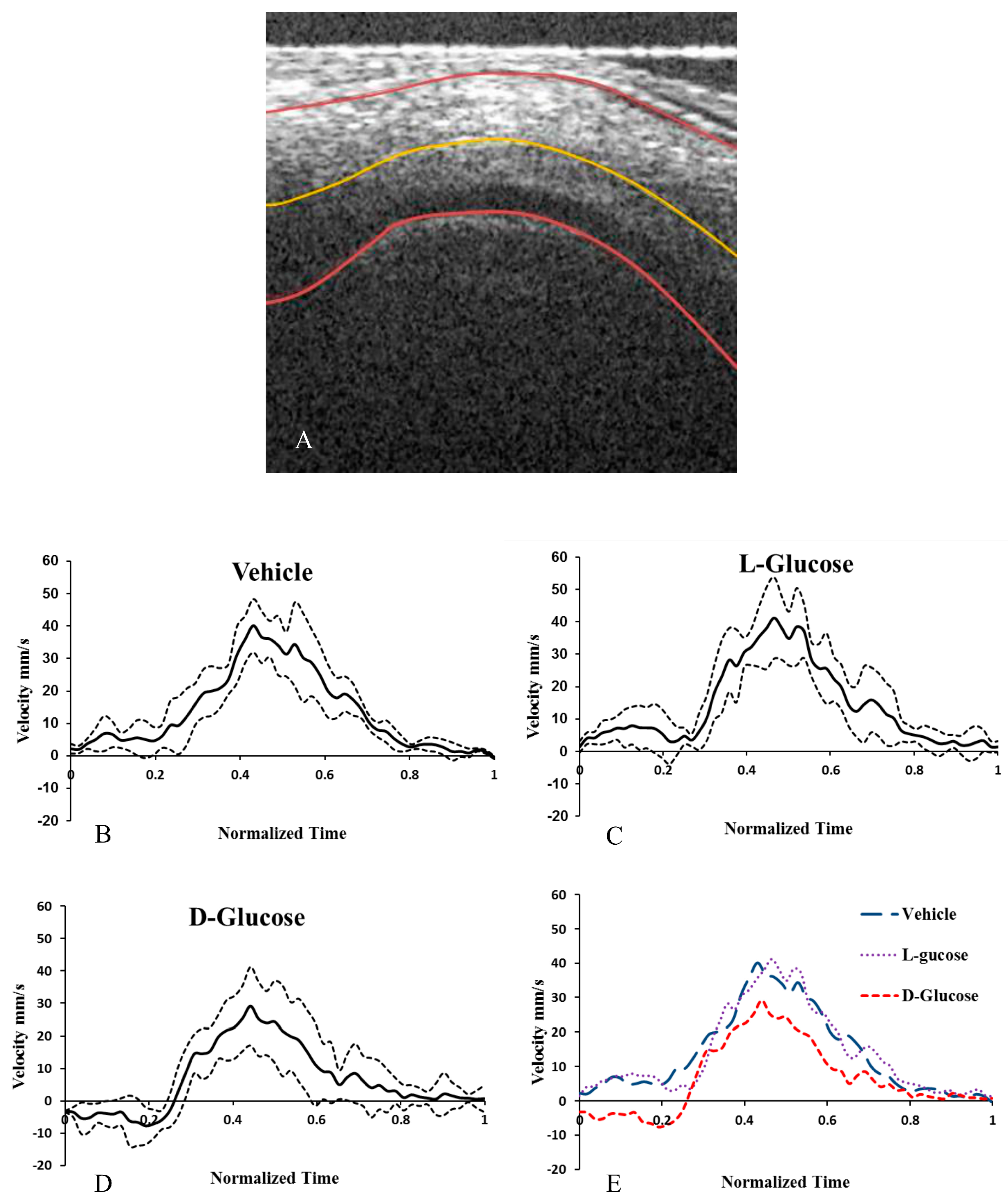
2.3. Hyperglycemia Reduces Altered Blood Flow Velocity in the OFT
3. Discussion
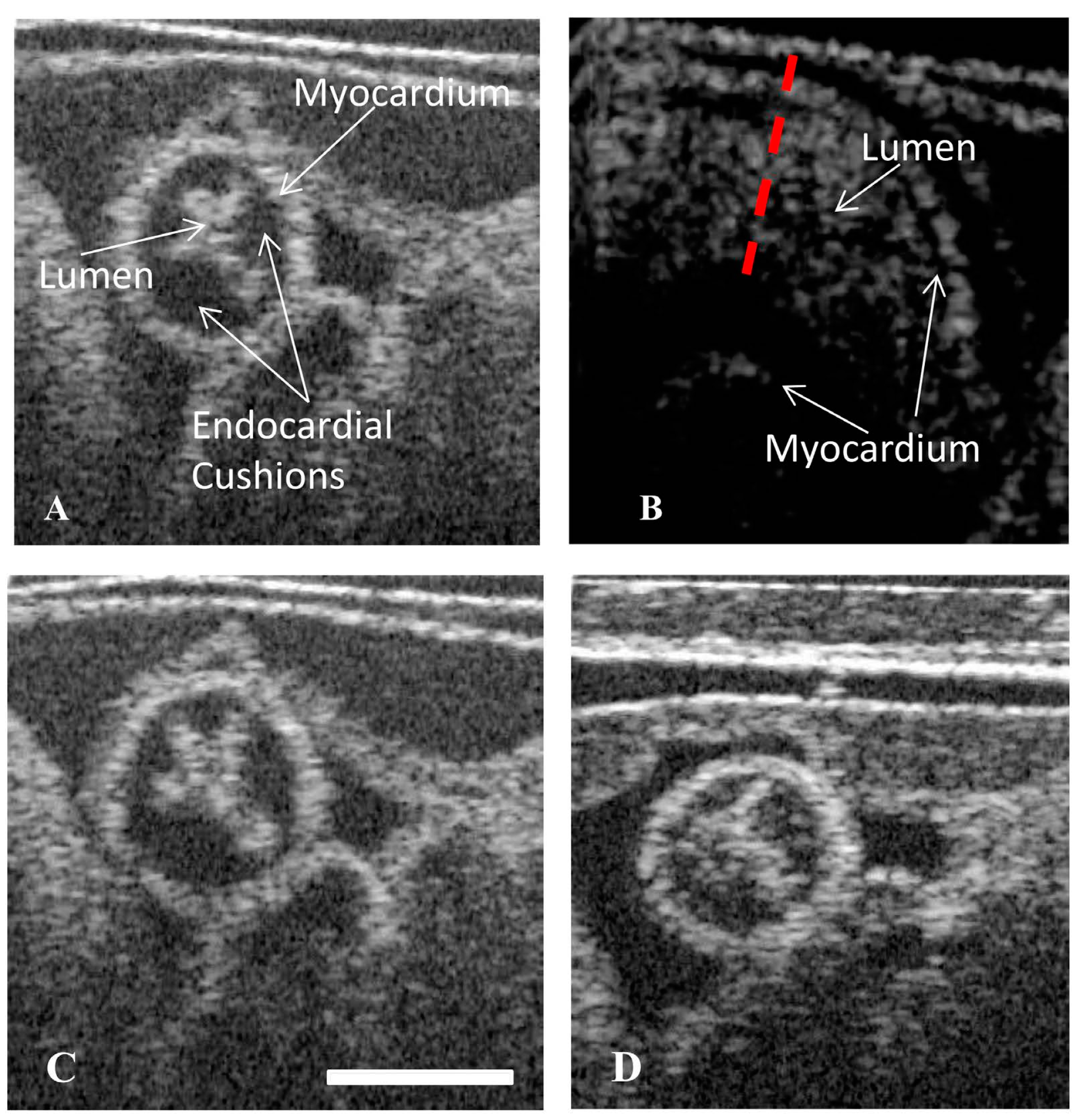
4. Materials and Methods
4.1. Embryo Hyperglycemia
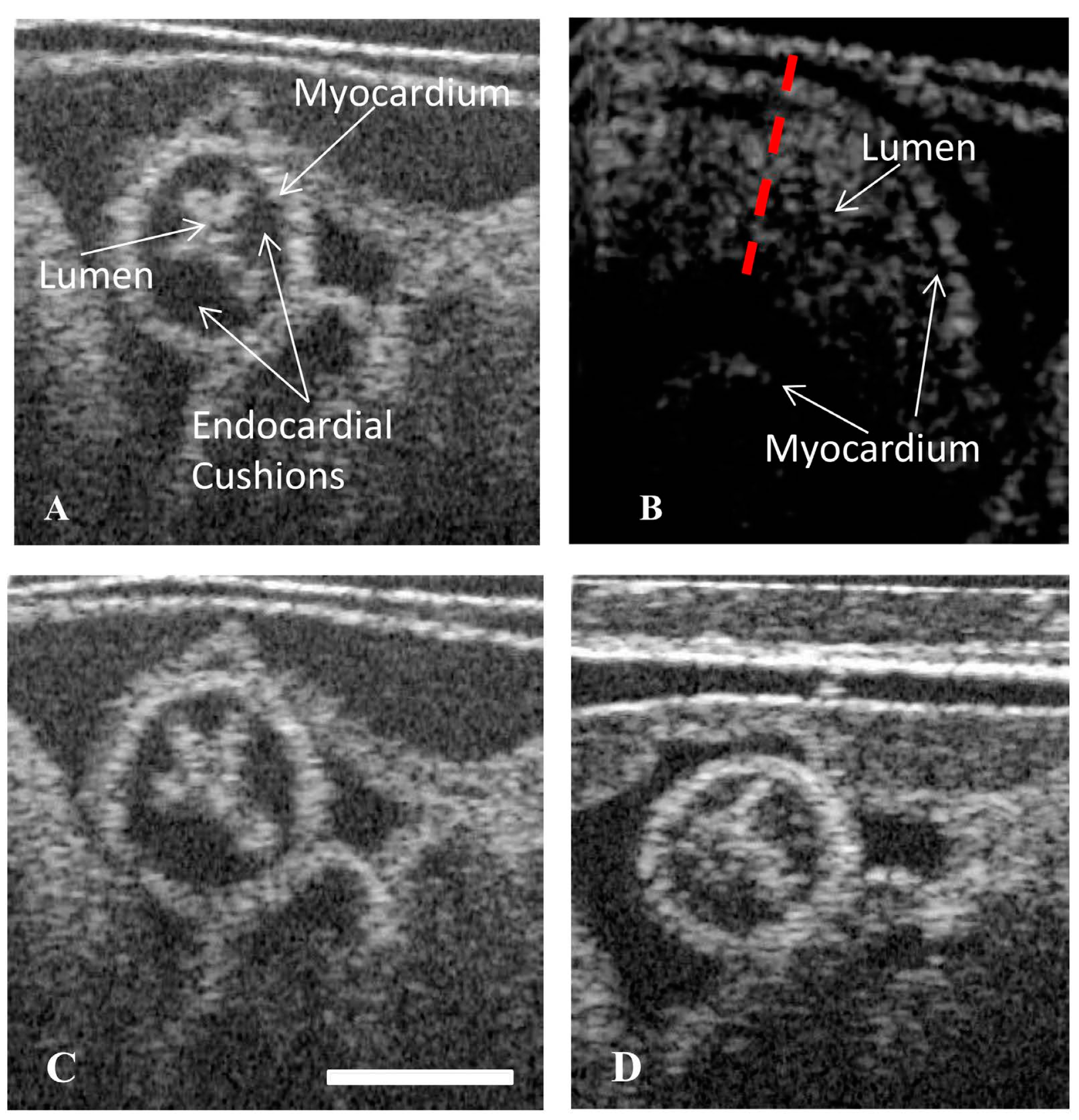
4.2. Dynamic Imaging of the OFT with Optical Coherence Tomography
4.3. Static Structural Imaging of the OFT
4.4. Structural Analysis
4.5. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hornberger, L.K. Maternal Diabetes and the Fetal Heart. Heart 2006, 92, 1019–1021. [Google Scholar] [CrossRef] [PubMed]
- Kousseff, B.G. Gestational Diabetes Mellitus (Class A): A Human Teratogen? Am. J. Med. Genet. Part A 1999, 83, 402–408. [Google Scholar] [CrossRef]
- Kousseff, B.G. Diabetic Embryopathy. Curr. Opin. Pediatr. 1999, 11, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, K.J.; Correa, A.; Feinstein, J.A.; Botto, L.; Britt, A.E.; Daniels, S.R.; Elixson, M.; Warnes, C.A.; Webb, C.L. Noninherited Risk Factors and Congenital Cardiovascular Defects: Current Knowledge—A Scientific Statement from the American Heart Association Council on Cardiovascular Disease in the Young. Circulation 2007, 115, 2995–3014. [Google Scholar] [CrossRef] [PubMed]
- Abu-Daya, A.; Sater, A.K.; Wells, D.E.; Mohun, T.J.; Zimmerman, L.B. Absence of Heartbeat in the Xenopus Tropicalis Mutation Muzak Is Caused by a Nonsense Mutation in Cardiac Myosin myh6. Dev. Biol. 2009, 336, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Hove, J.R.; Köster, R.W.; Forouhar, A.S.; Acevedo-Bolton, G.; Fraser, S.E.; Gharib, M. Intracardiac Fluid Forces Are an Essential Epigenetic Factor for Embryonic Cardiogenesis. Nature 2003, 421, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Culver, J.C.; Dickinson, M.E. The Effects of Hemodynamic Force on Embryonic Development. Microcirculation 2010, 17, 164–178. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.; Rosenquist, G. Spectrum of Cardiovascular Anomalies Following Cardiac Loop Constriction in the Chick Embryo. Birth Defects Orig. Artic. Ser. 1978, 14, 431–432. [Google Scholar] [PubMed]
- Stekelenburg-de Vos, S. Acutely Altered Hemodynamics Following Venous Obstruction in the Early Chick Embryo. J. Exp. Biol. 2003, 206, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Rugonyi, S.; Pentecost, J.O.; Thornburg, K.L. Finite Element Modeling of Blood Flow-Induced Mechanical Forces in the Outflow Tract of Chick Embryonic Hearts. Comput. Struct. 2007, 85, 727–738. [Google Scholar] [CrossRef]
- Liu, A.; Wang, R.K.; Thornburg, K.L.; Rugonyi, S. Dynamic Variation of Hemodynamic Shear Stress on the Walls of Developing Chick Hearts: Computational Models of the Heart Outflow Tract. Eng. Comput. 2009, 25, 73–86. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, A.; Yin, X.; Troyer, A.; Thornburg, K.; Wang, R.K.; Rugonyi, S. Measurement of Absolute Blood Flow Velocity in Outflow Tract of HH18 Chicken Embryo Based on 4D Reconstruction Using Spectral Domain Optical Coherence Tomography. Biomed. Opt. Express 2010, 1, 798–811. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Nickerson, A.; Troyer, A.; Yin, X.; Cary, R.; Thornburg, K.; Wang, R.; Rugonyi, S. Quantifying Blood Flow and Wall Shear Stresses in the Outflow Tract of Chick Embryonic Hearts. Comput. Struct. 2012, 89, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yin, X.; Shi, L.; Liu, A.; Rugonyi, S.; Wang, R. Measurement of Strain and Strain Rate in Embryonic Chick Heart In Vivo Using Spectral Domain Optical Coherence Tomography. Trans. Biomed. Eng. 2011, 58, 2333–2338. [Google Scholar]
- Liu, A.; Wang, R.; Thornburg, K.L.; Rugonyi, S. Efficient Postacquisition Synchronization of 4-D Nongated Cardiac Images Obtained from Optical Coherence Tomography: Application to 4-D Reconstruction of the Chick Embryonic Heart. J. Biomed. Opt. 2015, 14, 44020. [Google Scholar] [CrossRef] [PubMed]
- Rugonyi, S.; Shaut, C.; Liu, A.; Thornburg, K.; Wang, R.K. Changes in Wall Motion and Blood Flow in the Outflow Tract of Chick Embryonic Hearts Observed with Optical Coherence Tomography after Outflow Tract Banding and Vitelline-Vein Ligation. Phys. Med. Biol. 2008, 53, 5077–5091. [Google Scholar] [CrossRef] [PubMed]
- Martinsen, B.J. Reference Guide to the Stages of Chick Heart Embryology. Dev. Dyn. 2005, 233, 1217–1237. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D.; Olson, E.N. A Genetic Blueprint for Cardiac Development. Nature 2000, 407, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Carver, F.M.; Shibley, I.A.; Pennington, J.S.; Pennington, S.N. Differential Expression of Glucose Transporters during Chick Embryogenesis. Cell. Mol. Life Sci. 2001, 58, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, U.J.; Håkan Borg, L.A.; Cederberg, J.; Nordstrand, H.; Simán, C.M.; Wentzel, C.; Wentzel, P. Pathogenesis of Diabetes-Induced Congenital Malformations. Upsala J. Med. Sci. 2000, 105, 53–84. [Google Scholar] [CrossRef] [PubMed]
- Loffredo, C.; Wilson, P.; Ferencz, C. Maternal Diabetes: An Independent Risk Factor for Major Cardiovascular Malformations with Increased Mortality of Affected Infants. Teratology 2001, 64, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Scott-Drechsel, D.E.; Rugonyi, S.; Marks, D.L.; Thornburg, K.L.; Hinds, M.T. Hyperglycemia Slows Embryonic Growth and Suppresses Cell Cycle via Cyclin D1 and p21. Diabetes 2013, 62, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Narchi, H.; Kulaylat, N. Heart Disease in Infants of Diabetic Mothers. Images Paediatr. Cardiol. 2000, 2, 17–23. [Google Scholar] [PubMed]
- Harris, I.S.; Black, B.L. Development of the Endocardium. Pediatr. Cardiol. 2010, 31, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z. Cardiac Malformations and Alteration of TGFbeta Signaling System in Diabetic Embryopathy. Birth Defects Res. B Dev. Reprod. Toxicol. 2010, 89, 97–105. [Google Scholar] [PubMed]
- Molin, D.G.; Roest, P.A.; Nordstrand, H.; Wisse, L.J.; Poelmann, R.E.; Eriksson, U.J.; Groot, G.D.; Adriana, C. Disturbed Morphogenesis of Cardiac Outflow Tract and Increased Rate of Aortic Arch Anomalies in the Offspring of Diabetic Rats. Birth Defects Res. A Clin. Mol. Teratol. 2004, 70, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Svensson, K.; Eriksson, U.J. High Glucose Concentration Inhibits Migration of Rat Cranial Neural Crest Cells in Vitro. Diabetologia 1996, 39, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Reece, A. NIH Public Access. Clin. Lab. Med. 2012, 29, 997–1003. [Google Scholar]
- Morgan, S.C.; Relaix, F.; Sandell, L.L.; Loeken, M.R. Oxidative Stress during Diabetic Pregnancy Disrupts Cardiac Neural Crest Migration and Causes Outflow Tract Defects. Birth Defects Res. Part A Clin. Mol. Teratol. 2008, 82, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Larger, E.; Marre, M.; Corvol, P.; Gasc, J. Chicken Chorioallantoic Membrane Model. Methods Mol. Biol. 2012, 843, 47–57. [Google Scholar] [CrossRef]
- Midgett, M.; Rugonyi, S. Congenital Heart Malformations Induced by Hemodynamic Altering Surgical Interventions. Front. Physiol. 2014, 5, 287. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, K.N.; Spitz, C.; Shekhar, A.; Yalcin, H.C.; Butcher, J.T. Computational Fluid Dynamics of Developing Avian Outflow Tract Heart Valves. Ann. Biomed. Eng. 2012, 40, 2212–2227. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Yin, X.; Shi, L.; Li, P.; Thornburg, K.L.; Wang, R.; Rugonyi, S. Biomechanics of the Chick Embryonic Heart Outflow Tract at HH18 Using 4D Optical Coherence Tomography Imaging and Computational Modeling. PLoS ONE 2012, 7, e40869. [Google Scholar] [CrossRef] [PubMed]
- McCane, D.R.; Hanson, R.L.; Charles, M.A.; Jacobsson, T.H.; Pettitt, D.; Bennett, P.H.; Knowler, W.C. Comparison of Tests for Glycated Haemoglobin and Fasting and Two Hour Plasma Glucose Concentrations as Diagnostic Methods for Diabetes. BMJ 1994, 308, 1323. [Google Scholar] [CrossRef]
- Grout, J. Understanding your lab test results. Diabetes Self-Manag. 2013, 30, 27–30. [Google Scholar] [PubMed]
- Midgett, M.; Goenezen, S.; Rugonyi, S. Blood Flow Dynamics Reflect Degree of Outflow Tract Banding in Hamburger—Hamilton Stage 18 Chicken Embryos Blood Flow Dynamics Reflect Degree of Outflow Tract Banding in Hamburger—Hamilton Stage 18 Chicken Embryos. J. R. Soc. Interface 2014, 11, 20140643. [Google Scholar] [CrossRef] [PubMed]
- Piccinelli, M.; Veneziani, A.; Steinman, D.; Remuzzi, A.; Antiga, L. A Framework for Geometric Analysis of Vascular Structures: Application to Cerebral Aneurysms. IEEE Trans Biomed Eng. 2009, 8, 1141–1155. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Total No. Embryos | Cushion Asymmetry |
|---|---|---|
| Vehicle | 10 | 0 |
| l-glucose | 10 | 1 |
| d-glucose | 10 | 6 * |
| Treatment | Number of Samples | Cumulative Torsion | Average Arc Length |
|---|---|---|---|
| Vehicle | 12 | 57.01 ± 19.19 | 20.84 ± 6.92 |
| l-Glucose | 9 | 55.06 ± 22.58 | 21.15 ± 5.93 |
| d-Glucose | 8 | 71.27 ± 16.09 *+ | 20.75 ± 8.30 |
| Treatment | Average Velocity (mm/s) | Average Peak Velocity (mm/s) | Average Min. Velocity (mm/s) | % of Cycle with Back Flow | Cardiac Period (s) |
|---|---|---|---|---|---|
| Control | 13.9 ± 3.8 | 40.7 ± 12.4 | −0.60 ± 0.79 | 6.3 ± 5.2 | 0.39 ± 0.02 |
| l-Glucose | 17.4 ± 5.2 | 41.5 ± 15.6 | −1.73 ± 3.8 | 2.4 ± 3.7 | 0.406 ± 0.015 |
| d-Glucose | 8.0 ± 3.1*+ | 31.4 ± 8.2 | −12.4 ± 6.9 *+ | 30.1 ± 10.3* | 0.413 ± 0.023 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lawson, T.B.; Scott-Drechsel, D.E.; Chivukula, V.K.; Rugonyi, S.; Thornburg, K.L.; Hinds, M.T. Hyperglycemia Alters the Structure and Hemodynamics of the Developing Embryonic Heart. J. Cardiovasc. Dev. Dis. 2018, 5, 13. https://doi.org/10.3390/jcdd5010013
Lawson TB, Scott-Drechsel DE, Chivukula VK, Rugonyi S, Thornburg KL, Hinds MT. Hyperglycemia Alters the Structure and Hemodynamics of the Developing Embryonic Heart. Journal of Cardiovascular Development and Disease. 2018; 5(1):13. https://doi.org/10.3390/jcdd5010013
Chicago/Turabian StyleLawson, Taylor B., Devon E. Scott-Drechsel, Venkat Keshav Chivukula, Sandra Rugonyi, Kent L. Thornburg, and Monica T. Hinds. 2018. "Hyperglycemia Alters the Structure and Hemodynamics of the Developing Embryonic Heart" Journal of Cardiovascular Development and Disease 5, no. 1: 13. https://doi.org/10.3390/jcdd5010013