Antifungal Potential of Host Defense Peptide Mimetics in a Mouse Model of Disseminated Candidiasis


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Yeast Strains
2.1.2. MIC and MFC Studies
2.1.3. Mammalian Cell Cytotoxicity Assay
2.1.4. Hemolysis Assay
2.1.5. Yeast Membrane Permeability Assessment
2.2. Efficacy of HDP Mimetics in vivo
2.2.1. Mice
2.2.2. Candida Strains and Inoculum Preparation
2.2.3. Drug Preparation
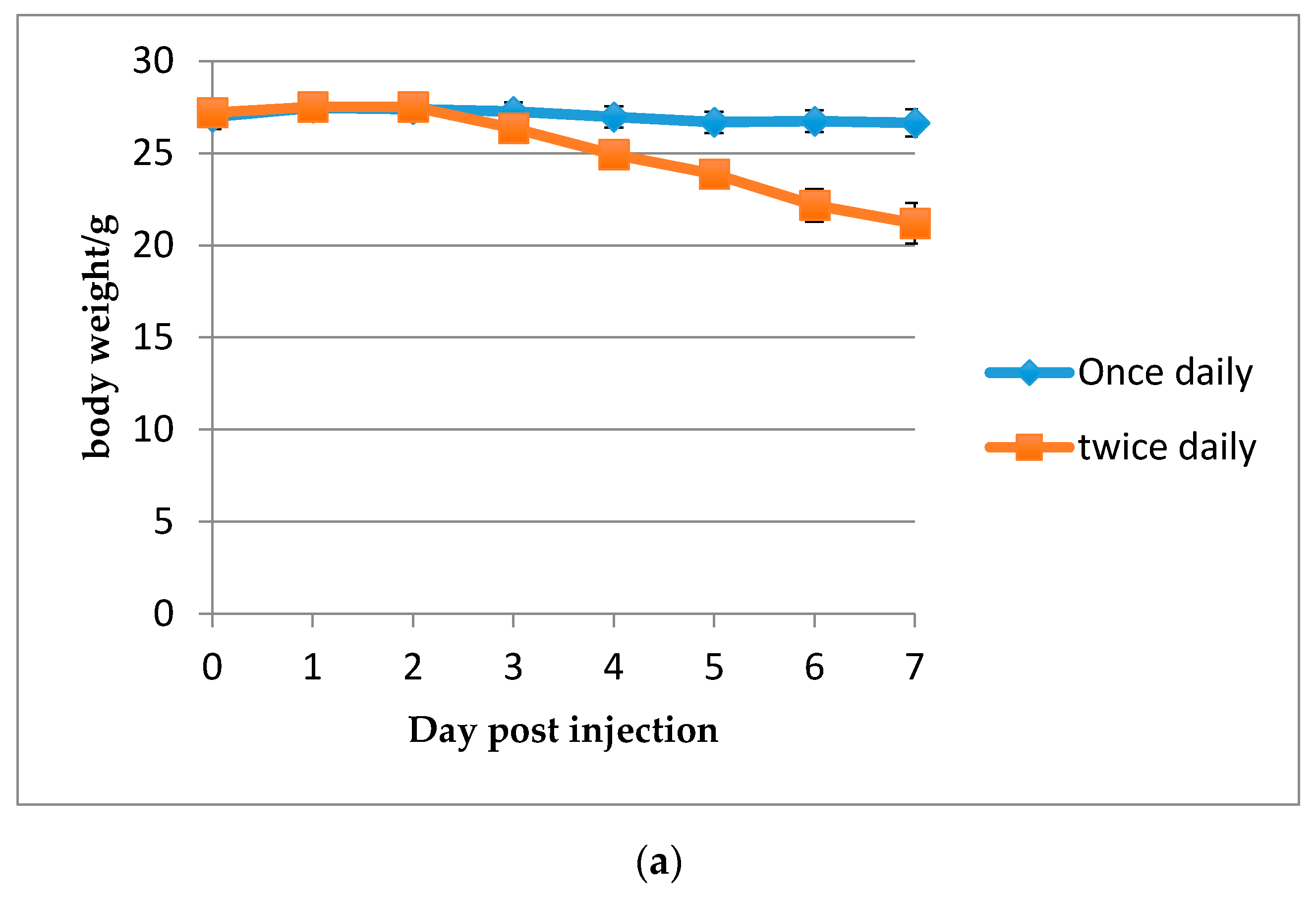
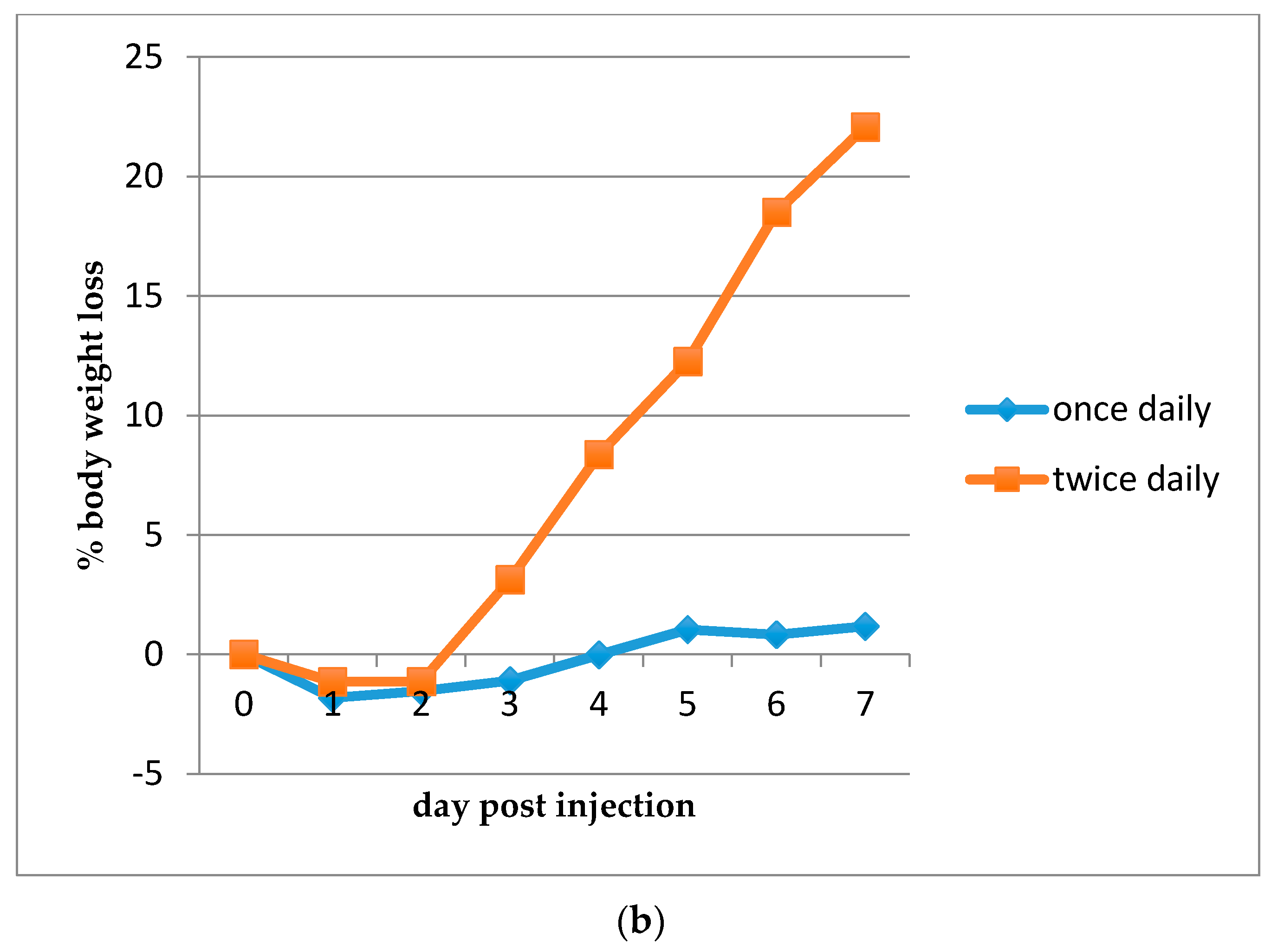
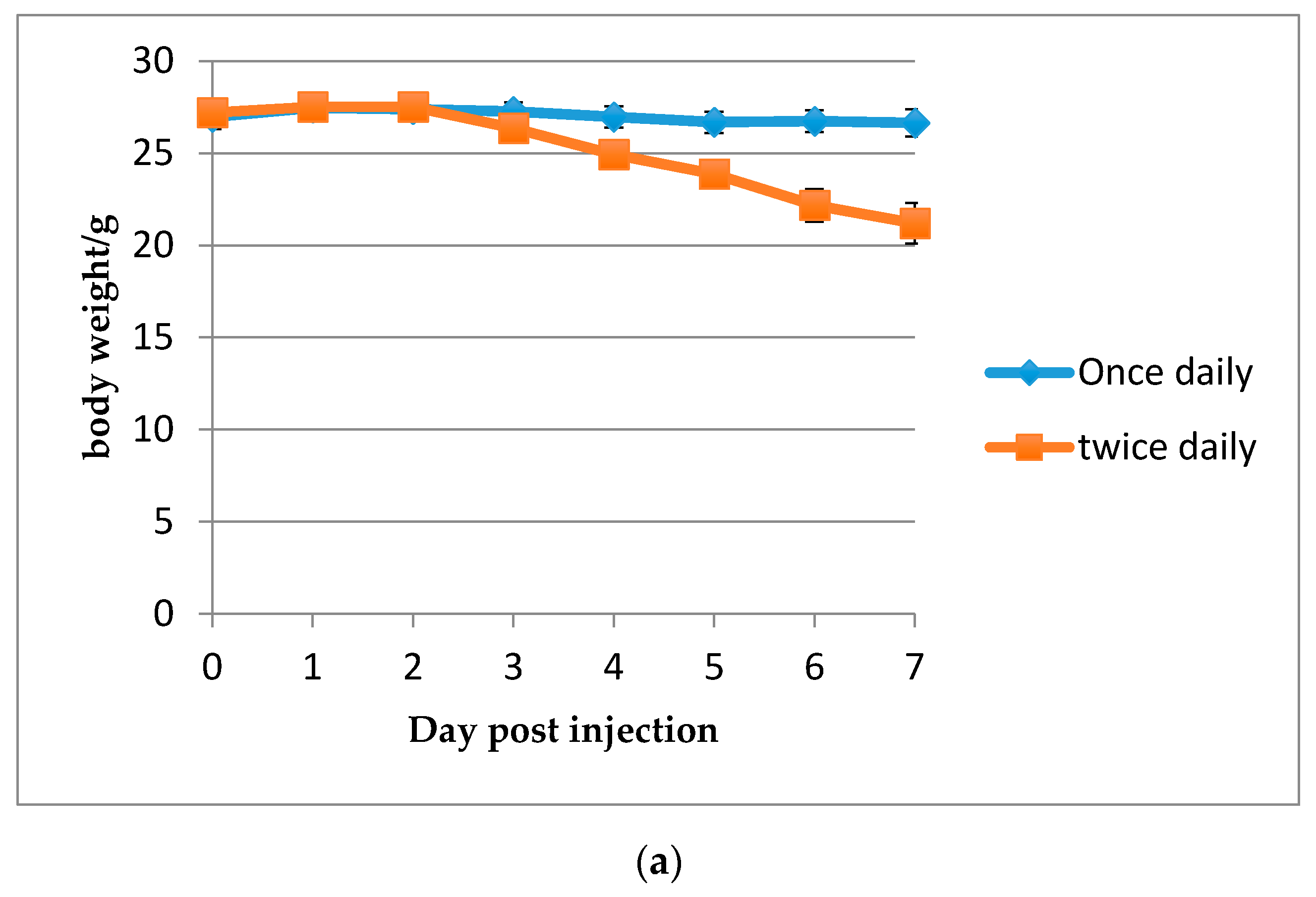
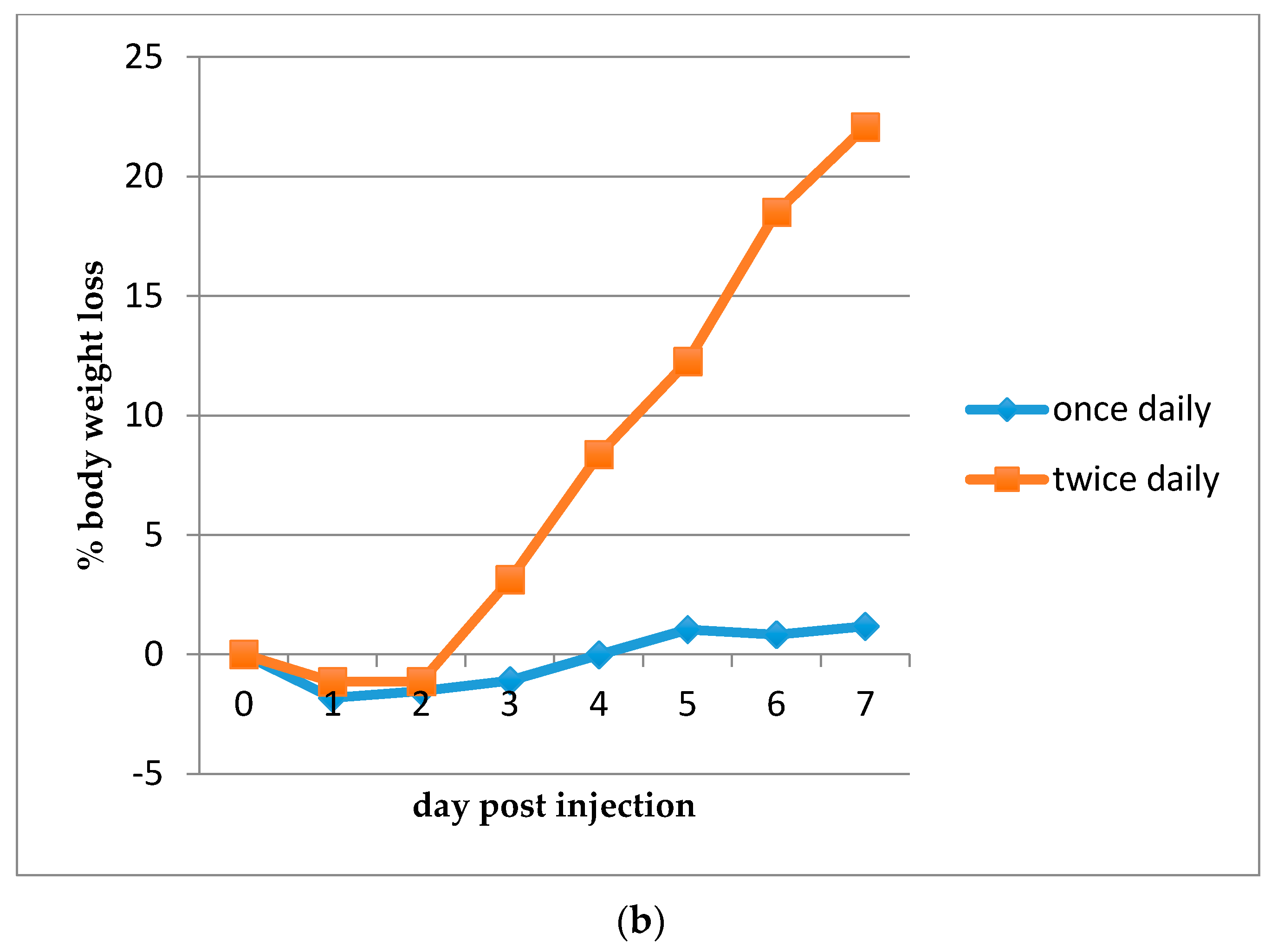
2.2.4. Determination of the Maximum Tolerated Dose (MTD)
2.2.5. Immunosuppressive Mouse Model of Invasive Candidiasis and Comparison of HDP Mimetic Efficacy at 50% MTD
2.2.6. Single Injection Dose Response Study of HDP Mimetics
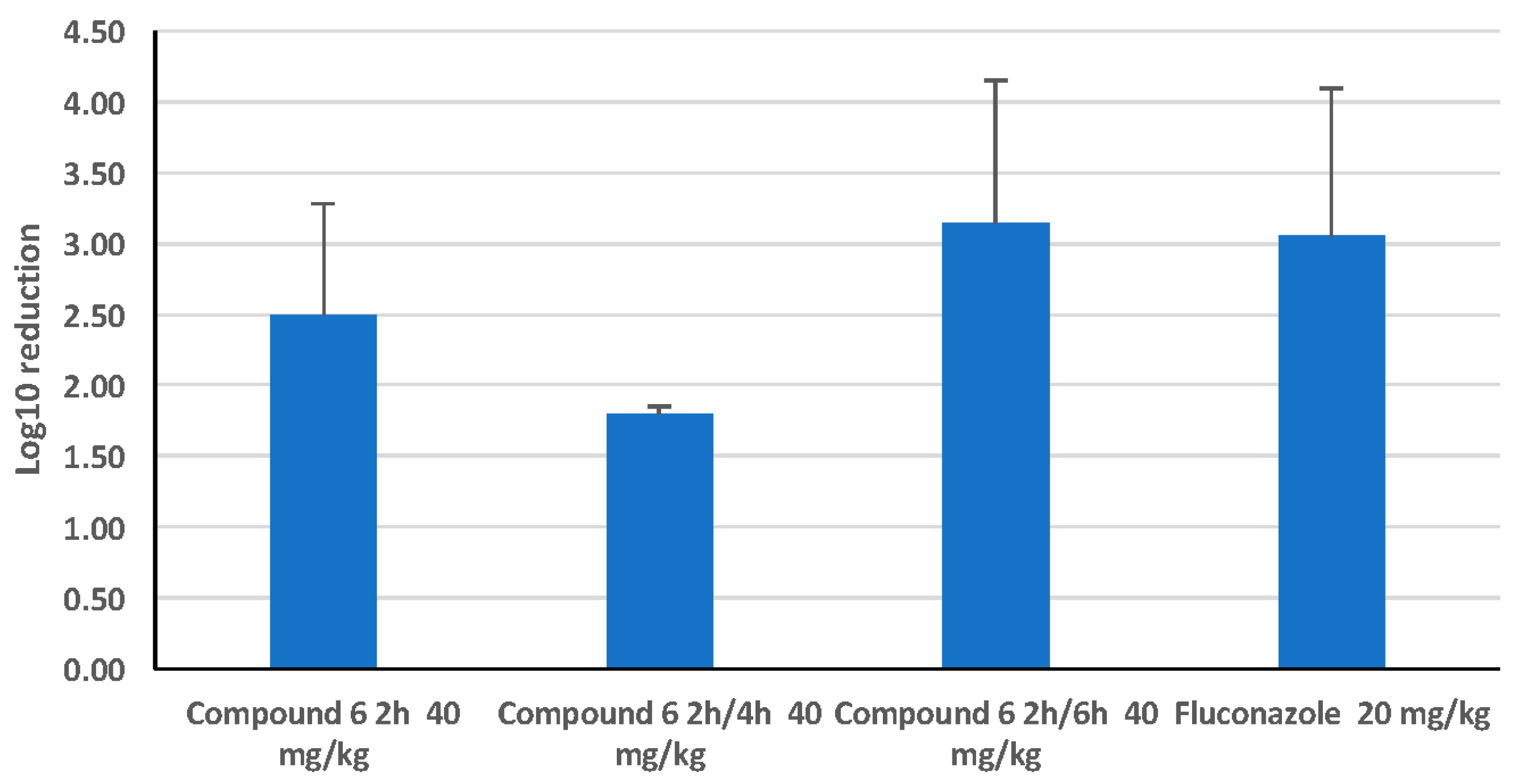
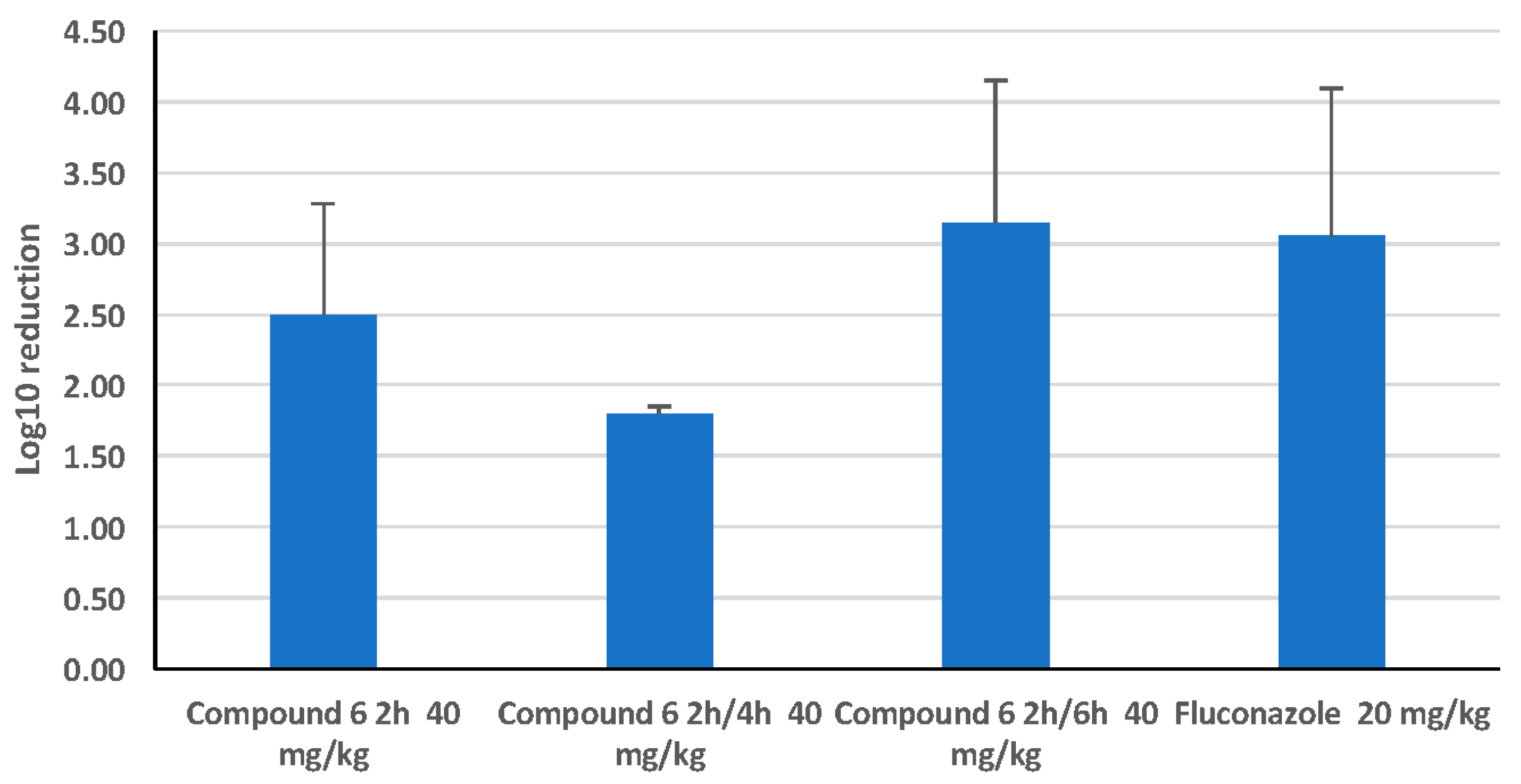
2.2.7. Comparison of a Single Dose with a Split Dose of HDP Mimetic
2.3. Statistical Analysis
2.4. Ethical Statement
3. Results
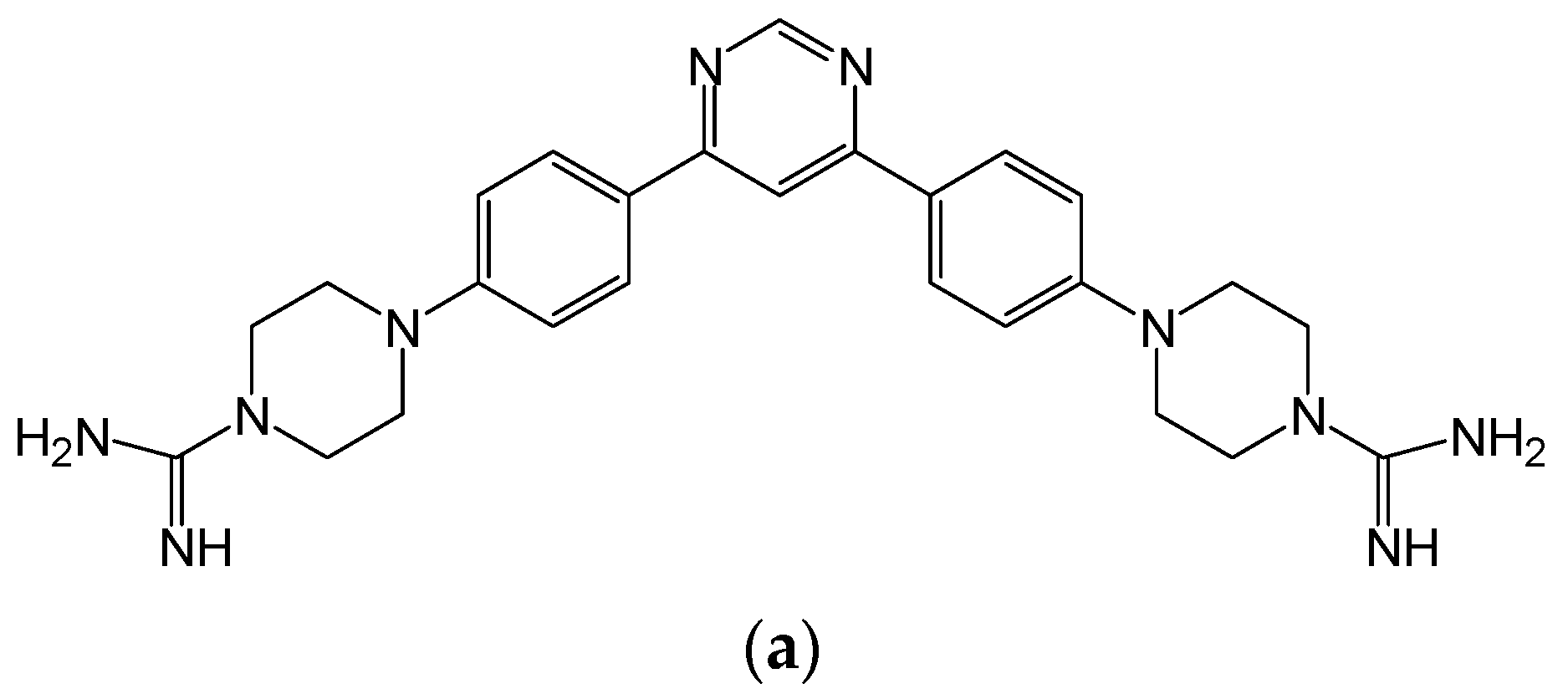
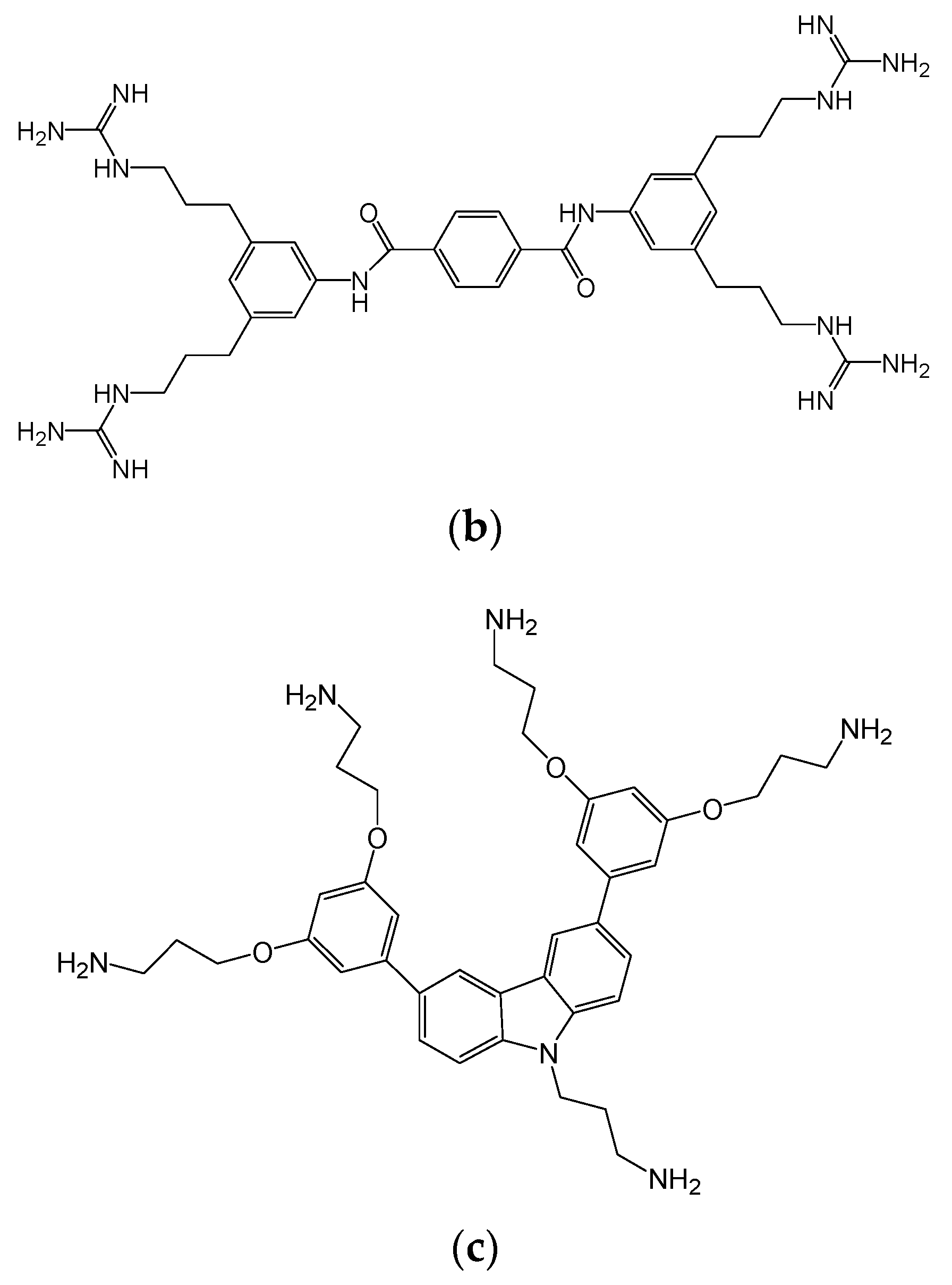
3.1. In Vitro Efficacy of HDP Mimetics against Candida Species
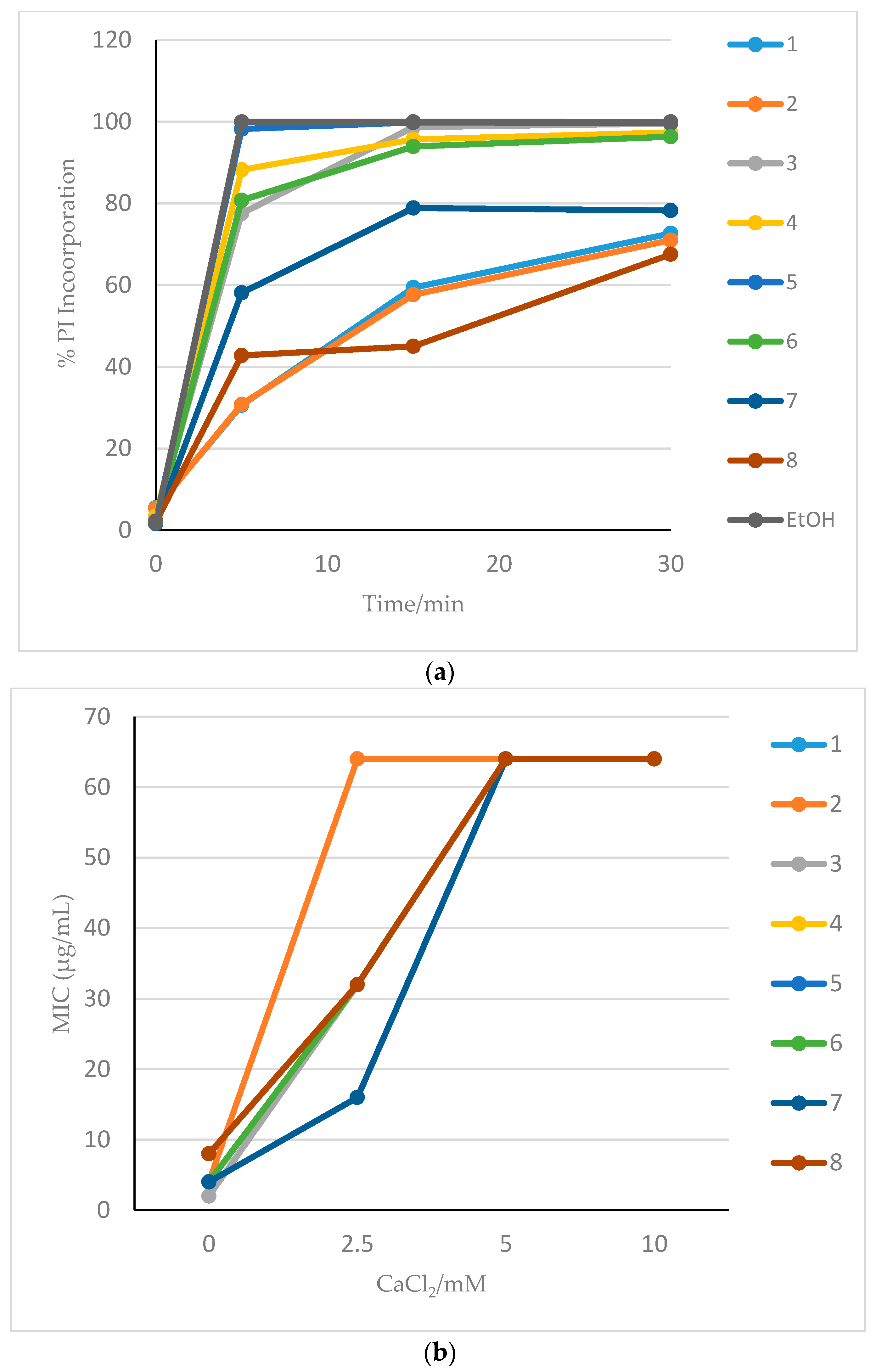
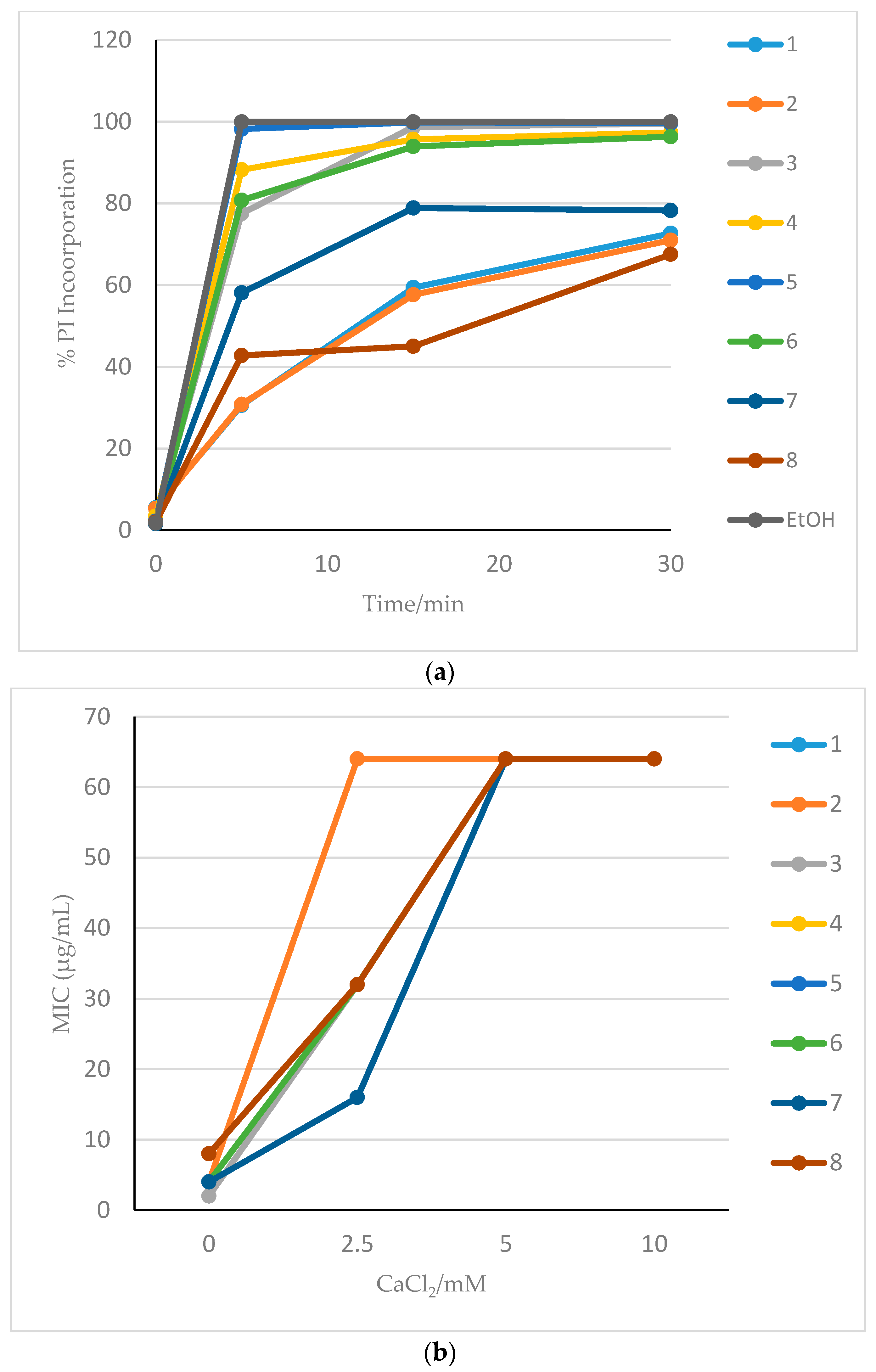
3.2. Membrane Effects of HDP Mimetics on Candida albicans
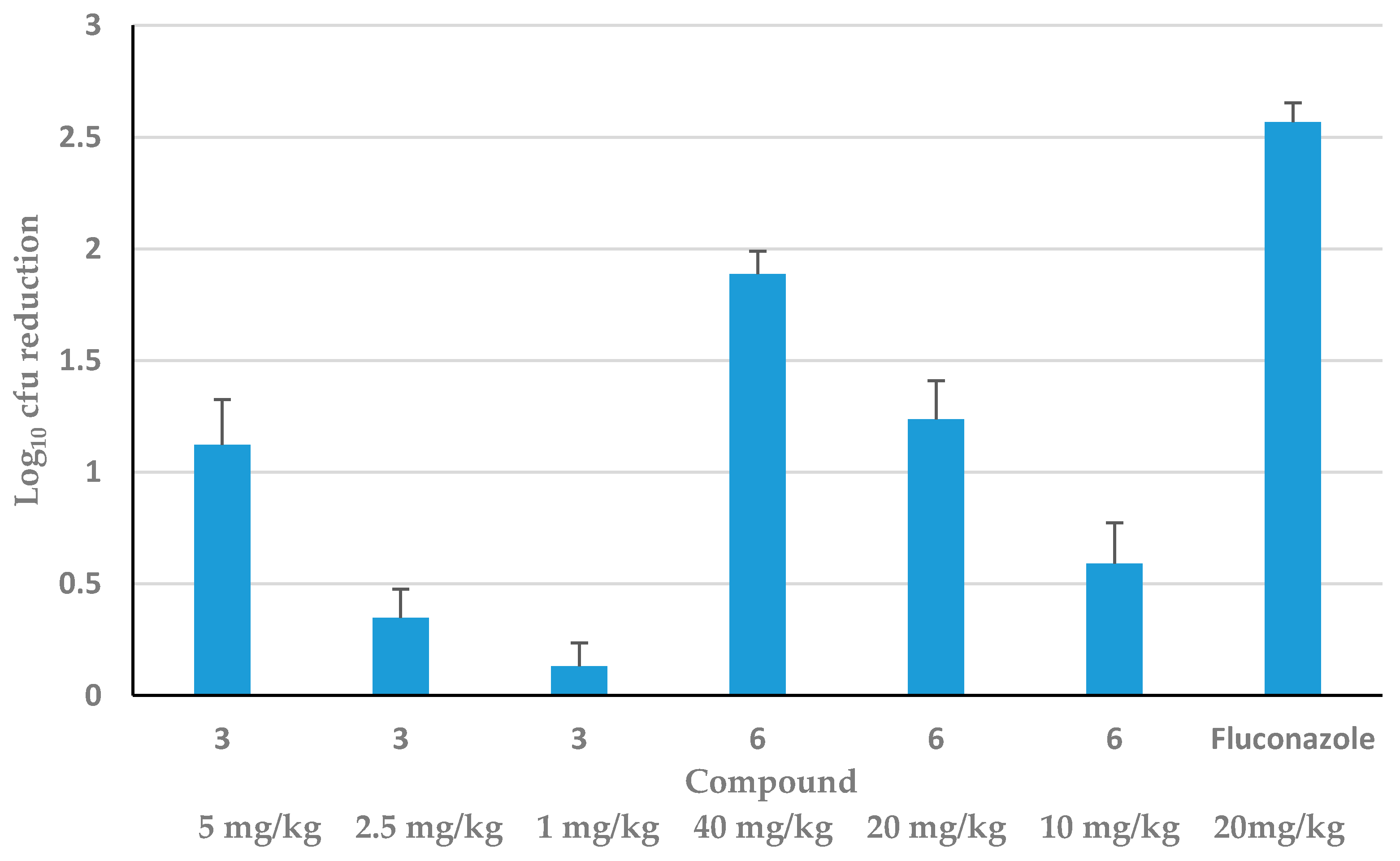
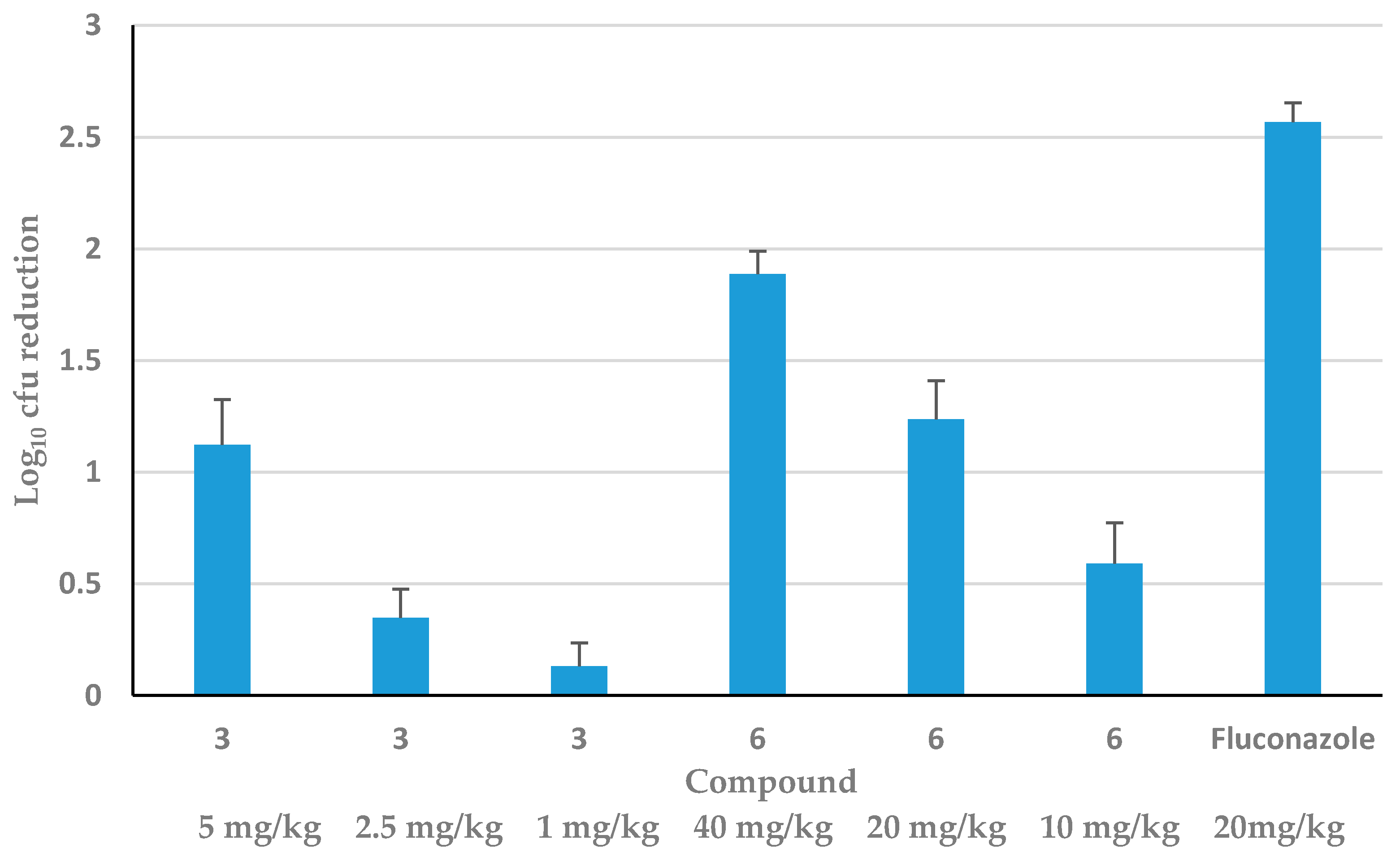
3.3. Efficacy of HDP Mimetics in a Mouse Model of Invasive Candidiasis
4. Discussion
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Bassetti, M.; Righi, E.; Montravers, P.; Cornely, O.A. What has changed in the treatment of invasive candidiasis? A look at the past 10 years and ahead. J. Antimicrob. Chemother. 2018, 73, i14–i25. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, B.J.; Arendrup, M.C. Invasive Candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamoth, F.; Lockhart, S.R.; Berkow, E.L.; Calandra, T. Changes in the epidemiological landscape of invasive candidiasis. J. Antimicrob. Chemother. 2018, 73, i4–i13. [Google Scholar] [CrossRef] [PubMed]
- Pea, F.; Lewis, R.E. Overview of antifungal dosing in invasive candidiasis. J. Antimicrob. Chemother. 2018, 73, i33–i43. [Google Scholar] [PubMed]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole Antifungal Resistance in Candida albicans and Emerging Non-albicans Candida Species. Front. Microbiol. 2016, 7, 2173. [Google Scholar] [CrossRef] [PubMed]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K.O. The roles of antimicrobial peptides in innate host defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef] [PubMed]
- Strom, M.B.; Haug, B.E.; Skar, M.L.; Stensen, W.; Stiberg, T.; Svendsen, J.S. The pharmacophore of short cationic antibacterial peptides. J. Med. Chem. 2003, 46, 1567–1570. [Google Scholar] [CrossRef] [PubMed]
- Mensa, B.; Kim, Y.H.; Choi, S.; Scott, R.; Caputo, G.A.; DeGrado, W.F. Antibacterial mechanism of action of arylamide foldamers. Antimicrob. Agents Chemother. 2011, 55, 5043–5053. [Google Scholar] [CrossRef] [PubMed]
- Tew, G.N.; Scott, R.W.; Klein, M.L.; Degrado, W.F. De novo design of antimicrobial polymers, foldamers, and small molecules: From discovery to practical applications. Acc. Chem. Res. 2010, 43, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Beckloff, N.; Laube, D.; Castro, T.; Furgang, D.; Park, S.; Perlin, D.; Clements, D.; Tang, H.; Scott, R.W.; Tew, G.N.; et al. Activity of an Antimicrobial Peptide Mimetic against Planktonic and Biofilm Cultures of Oral Pathogens. Antimicrob. Agents Chemother. 2007, 51, 4125–4132. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Yamarthy, R.; Felsenstein, S.; Scott, R.W.; Markowitz, K.; Diamond, G. Activity of antimicrobial peptide mimetics in the oral cavity: I. Activity against biofilms of Candida albicans. Mol. Oral Microbiol. 2010, 25, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Ryan, L.K.; Freeman, K.B.; Masso-Silva, J.A.; Falkovsky, K.; Aloyouny, A.; Markowitz, K.; Hise, A.G.; Fatahzadeh, M.; Scott, R.W.; Diamond, G. Activity of potent and selective host defense Peptide mimetics in mouse models of oral candidiasis. Antimicrob. Agents Chemother. 2014, 58, 3820–3827. [Google Scholar] [CrossRef] [PubMed]
- Menzel, L.P.; Chowdhury, H.M.; Masso-Silva, J.A.; Ruddick, W.; Falkovsky, K.; Vorona, R.; Malsbary, A.; Cherabuddi, K.; Ryan, L.K.; DiFranco, K.M.; et al. Potent in vitro and in vivo antifungal activity of a small molecule host defense peptide mimic through a membrane-active mechanism. Sci. Rep. 2017, 7, 4353. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.W.; Tew, G.N. Mimics of Host Defense Proteins; Strategies for Translation to Therapeutic Applications. Curr. Top. Med. Chem. 2017, 17, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Choi, S.; Chen, B.; Doerksen, R.J.; Clements, D.J.; Winkler, J.D.; Klein, M.L.; DeGrado, W.F. Nontoxic membrane-active antimicrobial arylamide oligomers. Angew. Chem. Int. Ed. Engl. 2004, 43, 1158–1162. [Google Scholar] [CrossRef] [PubMed]
- Tew, G.N.; Liu, D.; Chen, B.; Doerksen, R.J.; Kaplan, J.; Carroll, P.J.; Klein, M.L.; DeGrado, W.F. De novo design of biomimetic antimicrobial polymers. Proc. Natl. Acad. Sci. USA 2002, 99, 5110–5414. [Google Scholar] [CrossRef] [PubMed]
- Sanati, H.; Ramos, C.F.; Bayer, A.S.; Ghannoum, M.A. Combination therapy with amphotericin B and fluconazole against invasive candidiasis in neutropenic-mouse and infective-endocarditis rabbit models. Antimicrob. Agents Chemother. 1997, 41, 1345–1348. [Google Scholar] [PubMed]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 62, e1–e50. [Google Scholar] [CrossRef] [PubMed]
- Vande Velde, G.; Kucharikova, S.; Schrevens, S.; Himmelreich, U.; Van Dijck, P. Towards non-invasive monitoring of pathogen-host interactions during Candida albicans biofilm formation using in vivo bioluminescence. Cell. Microbiol. 2014, 16, 115–130. [Google Scholar] [CrossRef] [PubMed]
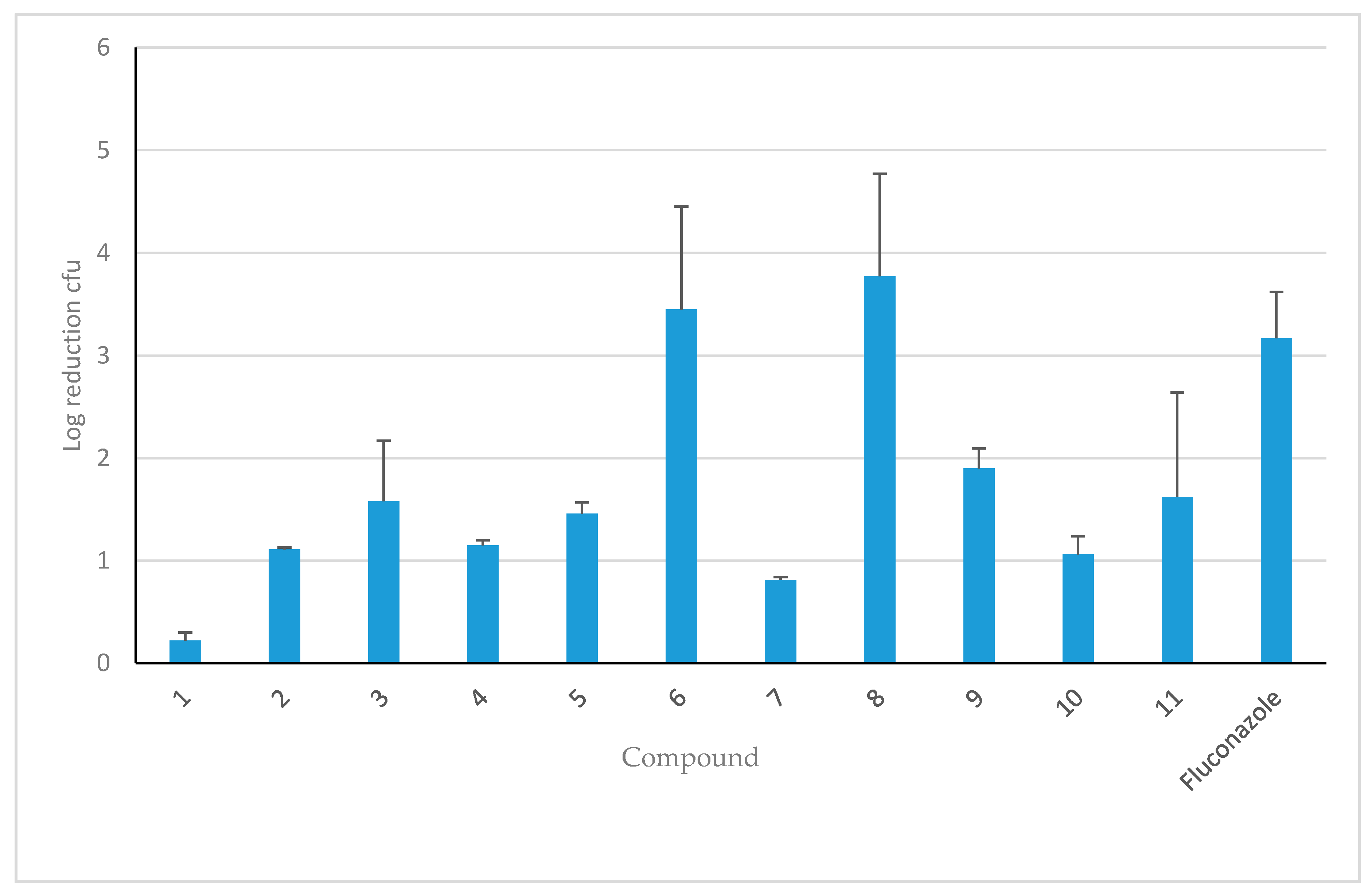

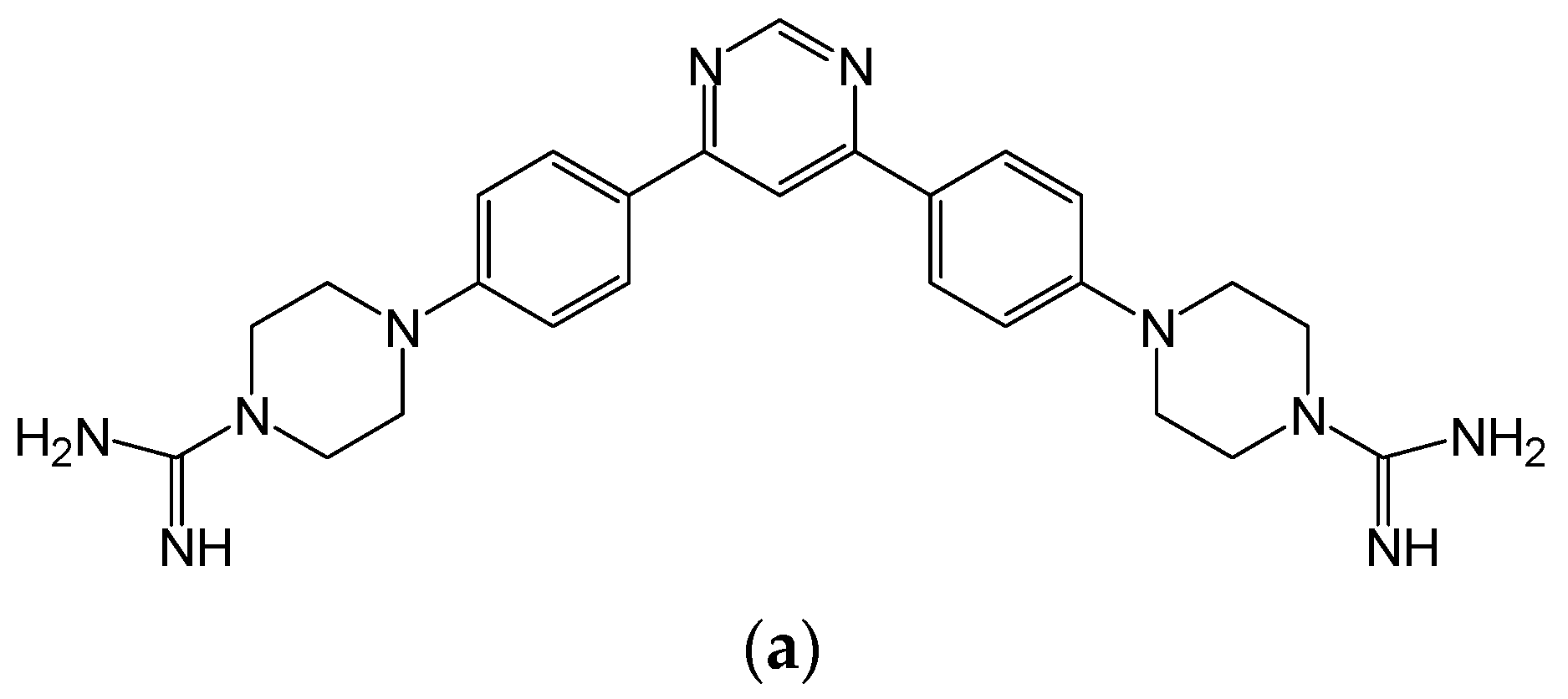{kind=link}
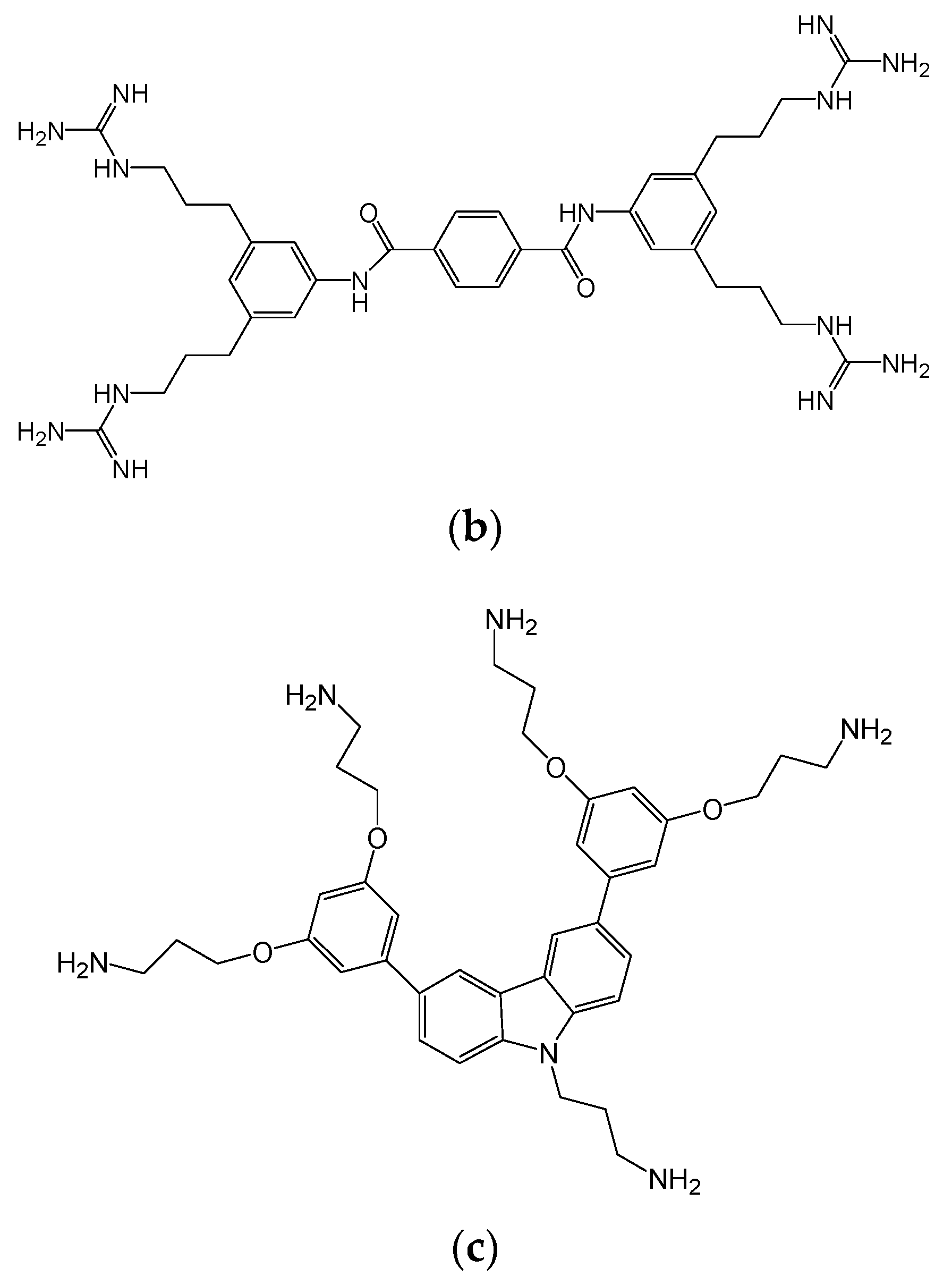{kind=link}
{kind=link}
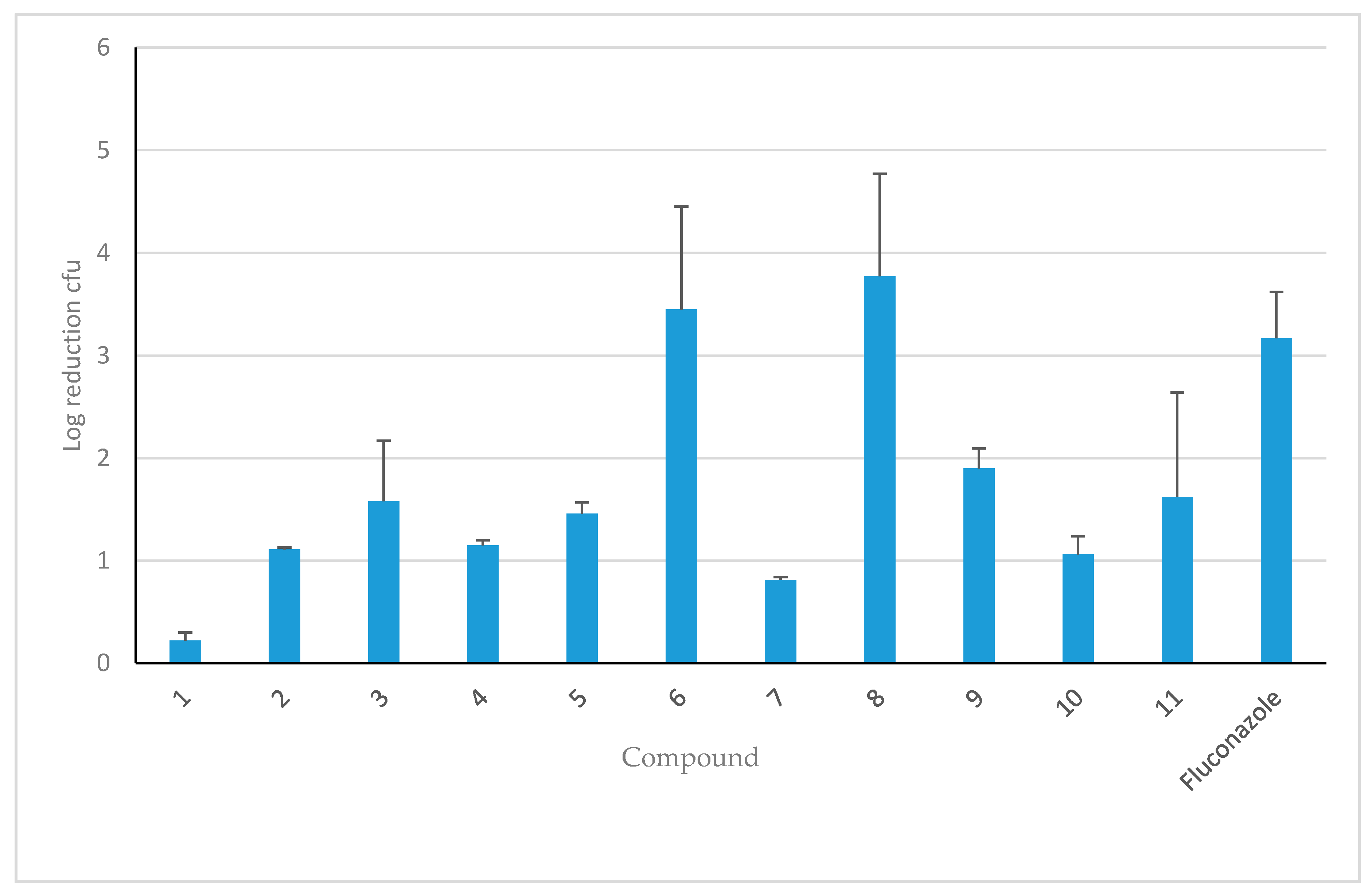{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | Fluconazole | Caspofungin |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Series | Tri-aryl | Bis-amide | Tri-cyclic | Tri-cyclic | Bis-amide | Bis-amide | Bi-aryl | Bis-amide | Bis-amide | Bi-aryl | Bi-aryl | ||
| Activity against C. albicans (μg/mL) | |||||||||||||
| MIC | 4 | 4 | 2 | 4 | 4 | 4 | 4 | 8 | >8 | 0.3 | 8 | 0.25 | 0.25 |
| MFC | 8 | 16 | 8 | 8 | 8 | 8 | 8 | 16 | >8 | 0.78 | ND | 0.5 | |
| MIC+50% Human serum | 4 | 4 | 16 | 4 | 16 | 16 | 16 | 32 | >8 | ND | >8 | ||
| NAC species MIC (μg/mL) | |||||||||||||
| C. tropicalis | 4–8 | 8 | 4 | 2–4 | 2 | 4 | 8 | 4 | 2 | ND | 1 | ||
| C. parapsilosis | 4–8 | 1 | 1 | 2 | 1 | 2 | 2 | 4 | 0.5 | ND | 0.25 | ||
| C. dublinensis | 8 | 32 | 16 | 4 | 2 | 2 | 4 | 4 | ND | ND | ND | ||
| C. glabrata | 4 | 8 | 2 | 2 | 4 | 4 | 4 | 8 | >8 | ND | 1 | ||
| C. krusei | 32 | 8 | 0.50 | 16 | 2 | 2 | 4 | 4 | 1 | ND | 4 | ||
| Fluconazole-resistant NAC Derived from Clinical Isolates from Different Patients: TG-Candida glabrata, CT-Candida tropicalis | MIC (μg/mL) | MFC(μg/mL) |
|---|---|---|
| TG-1 | >64 | >64 |
| TG-3 | >64 | >64 |
| TG-4 | >64 | >64 |
| TG-5 | >64 | >64 |
| TG-6 | >64 | >64 |
| CT-2 | 8 | 32 |
| Candida albicans SC5314 | 4 | 8 |
| Compounds | Cytotoxicity 3T3 EC50 (µM) | Cytotoxicity HepG2 EC50 (µM) | Hemolysis EC50 (µM) |
|---|---|---|---|
| 1 | 311 | ND | >1792 |
| 2 | 358 | ND | >1164 |
| 3 | 478 | 137 | 396.2 |
| 4 | 436 | ND | >1170 |
| 5 | 891 | 315.1 | 1453.5 |
| 6 | >1398.6 | >1398.6 | 1331 |
| 7 | >788.6 | >788.6 | >1577.3 |
| 8 | 390 | 249 | >1272 |
| 9 | 725 | 1456 | 1173 |
| 10 | >1445 | >1445 | >1445 |
| 11 | >1518 | >1518 | >1517 |
| Compounds | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MTD (mg/kg free base) | 10 | 10 | 10 | 20 | 5 | ≥40 | 20 | ≥40 | ≥40 | 10 | 10 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chowdhury, M.H.; Ryan, L.K.; Cherabuddi, K.; Freeman, K.B.; Weaver, D.G.; Pelletier, J.C.; Scott, R.W.; Diamond, G. Antifungal Potential of Host Defense Peptide Mimetics in a Mouse Model of Disseminated Candidiasis. J. Fungi 2018, 4, 30. https://doi.org/10.3390/jof4010030
Chowdhury MH, Ryan LK, Cherabuddi K, Freeman KB, Weaver DG, Pelletier JC, Scott RW, Diamond G. Antifungal Potential of Host Defense Peptide Mimetics in a Mouse Model of Disseminated Candidiasis. Journal of Fungi. 2018; 4(1):30. https://doi.org/10.3390/jof4010030
Chicago/Turabian StyleChowdhury, Mobaswar Hossain, Lisa Kathleen Ryan, Kartikeya Cherabuddi, Katie B. Freeman, Damian G. Weaver, Jeffry C. Pelletier, Richard W. Scott, and Gill Diamond. 2018. "Antifungal Potential of Host Defense Peptide Mimetics in a Mouse Model of Disseminated Candidiasis" Journal of Fungi 4, no. 1: 30. https://doi.org/10.3390/jof4010030