Sponge Long Non-Coding RNAs Are Expressed in Specific Cell Types and Conserved Networks


1
School of Biological Sciences, University of Queensland, Brisbane, QLD 4072, Australia
2
Department of Medicine, Weill Cornell Medicine, and New York Genome Center, New York, NY 10021, USA
3
Department of Biological Sciences, Carnegie Mellon University, Pittsburgh, PA 15213, USA
*
Author to whom correspondence should be addressed.
Non-Coding RNA 2018, 4(1), 6; https://doi.org/10.3390/ncrna4010006
Submission received: 1 December 2017
/
Revised: 5 February 2018
/
Accepted: 27 February 2018
/
Published: 7 March 2018
(This article belongs to the Special Issue Non-Coding RNAs, from an Evolutionary Perspective)
Abstract
:Although developmental regulation by long non-coding RNAs (lncRNAs) appears to be a widespread feature amongst animals, the origin and level of evolutionary conservation of this mode of regulation remain unclear. We have previously demonstrated that the sponge Amphimedon queenslandica—a morphologically-simple animal—developmentally expresses an array of lncRNAs in manner akin to more complex bilaterians (insects + vertebrates). Here, we first show that Amphimedon lncRNAs are expressed in specific cell types in larvae, juveniles and adults. Thus, as in bilaterians, sponge developmental regulation involves the dynamic, cell type- and context-specific regulation of specific lncRNAs. Second, by comparing gene co-expression networks between Amphimedon queenslandica and Sycon ciliatum—a distantly-related calcisponge—we identify several putative co-expression modules that appear to be shared in sponges; these network-embedded sponge lncRNAs have no discernable sequence similarity. Together, these results suggest sponge lncRNAs are developmentally regulated and operate in conserved gene regulatory networks, as appears to be the case in more complex bilaterians.
Keywords:
evolution; cell type; gene expression; complexity; animals; long non-coding RNAs; development1. Introduction
Animal genomes encode thousands of long non-coding RNAs (lncRNAs) with no apparent protein coding capacity [1,2,3,4,5,6,7,8,9,10,11,12,13,14]. Despite their similarities with protein-coding genes, such as being spliced, transcribed by RNA polymerase II, and possessing 5′-terminal methylguanosine caps and poly(A) 3′-tails [15,16], lncRNAs tend to be expressed in specific cell types [17] and in a more tissue- and developmental stage-restricted manner than protein-coding genes [2,18,19,20,21,22,23,24]. This suggests that animal development requires the fine-scale regulation of expression of specific lncRNAs [25].
However, lncRNAs are rapidly evolving and exhibit poor primary sequence similarity between species. Orthologues are difficult to identify [26], thus precluding a detailed understanding of their evolution in terms of sequence, structure and function. In fact, while the role of lncRNAs in the regulation of developmental gene activity appears to be widespread amongst animals [18,20,22,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41], only a handful of lncRNAs have thus far been shown to possess conserved function(s) in evolutionarily divergent animals, with all functional studies to date being restricted to bilaterians [36,39,42,43,44].
Porifera (sponges) are one of the oldest surviving phyletic lineages of animals, diverging from other animals around 700 Mya [45]. As such, they are an informative lineage for understanding the origin and evolution of animal lncRNAs. Traits shared between sponges and all other animals likely reflect shared inheritance from their last common ancestor (Figure 1A). Sponges consist of four classes—Demospongiae, Calcarea, Homoscleromorpha and Hexactinellida [46,47,48]—of morphologically-simple animals (they lack true gut, nerves and muscles) that share a common body organization and appear to have diverged from each other over 650 Mya [45] (Figure S1). They possess an internal network of canals and ciliated choanocyte chambers lined with epithelial cells, primarily endopinacocytes and choanocytes, and are separated from the external environment by another epithelial layer, the exopinacoderm. Choanocyte chambers pump water through this internal aquiferous canal system, drawing food into the sponge. Between the internal and external epithelial layers is the collagenous mesohyl, which is enriched with multiple types of amoebocytes, including the pluripotent stem cell type—the archeocyte. This juvenile body plan is the outcome of the dramatic reorganization of the radially-symmetrical, bi- or trilayered larva at metamorphosis [49,50,51] (Figure 1B–D).
Despite being one of the simplest animals, sponges developmentally express an array of lncRNAs in manner akin to more morphologically-complex insects and vertebrates [18,20]. These lncRNAs also appear to belong to co-expressed developmental gene modules [18,20], suggesting that complex genome regulation by lncRNAs is an ancient animal trait. However, whether sponge lncRNAs possess other features conserved in bilaterian lncRNAs, including cell type-restricted expression, remains unclear. Here, following our identification and characterization of lncRNAs in the demosponge Amphimedon queenslandica (herein Amphimedon) [20], we analyze spatially- and cell type-restricted expression patterns of a subset of Amphimedon lncRNAs. We also identify a number of modules of co-expressed homologous protein-coding genes and lncRNAs that appear to be conserved between Amphimedon and the calcareous sponge Sycon ciliatum (herein Sycon) [18,20]; these sponges diverged from each other more than 650 Mya [45]. Together, these analyses of sponge lncRNAs further implicate this class of rapidly-evolving non-coding RNAs in the regulation of metazoan development.
2. Results and Discussion
2.1. Sponge lncRNAs Are Enriched in Specific Cell Types
Long non-coding RNAs exhibit cell type-specific restricted expression patterns in bilaterians [2,17,19,21,24,55,56,57]. To assess whether this is the case for Amphimedon lncRNAs and to better understand their putative role(s) in a cellular context, we analyzed cell-type specific CEL-seq (Cell Expression by Linear amplification and sequencing) datasets from three of the main cell-types of sponges–archeocytes, pinacocytes, and choanocytes (Figure 1C,D) [58]. Reads from these datasets were mapped to Amphimedon lncRNA collection (n = 2935) [20] to identify cell-type specific enriched lncRNAs. A set of 684 lncRNAs had detectable expression (CEL-seq normalized count > 0 across the cell-type specific transcriptomes) (Table S1), 136 of which (~20%) were identified as being differentially expressed (Q < 0.05) between the three cell types (Tables S2–S5). Most of these lncRNAs are enriched in choanocytes or archeocytes (57% and 24%, respectively), with only five lncRNAs being pinacocyte-specific (Figure 1E).
We detected no significant structural differences between the 136 cell-type enriched lncRNAs and the remaining ubiquitously expressed lncRNAs (n = 548), with gene length (511 ± 36.28 vs. 451 ± 10.61 (average ± SEM), respectively; Mann-Whitney U test, p-value = 0.096244) and number of exons (1.9 ± 0.09 vs. 1.7 ± 0.05 (average ± SEM), respectively; Mann-Whitney U test, p-value = 0.086181) being similar. Moreover, these two groups of lncRNAs were not significantly different in relation to their positions and direction of transcription with respect to protein-coding genes (i.e., 49.6% vs. 49.2% intergenic co-location; and 20.2% vs. 17.7% having at least one exon that overlaps with an exon of a protein-coding gene on the opposite strand, respectively; Fisher’s exact test, p-value > 0.05 in both cases).
However, of the choanocyte-enriched lncRNAs, AmqTCONS_00001337, AmqTCONS_00001338, and AmqTCONS_00001339 are in close genomic proximity to a cluster of immune-related genes (Tnf receptor-associated factors (TRAFs)) (Figure 1F), which are also differentially enriched in choanocytes [59]. Interestingly, the Tnf receptor-associated factor 4-like belonging to this cluster of immune-related genes (Aqu2.1.23792_001) was also co-expressed with these three lncRNAs (i.e., belonging to the same developmental co-expression module; see Table S7).
Amphimedon possesses nearly 300 genes from the scavenger receptor cysteine-rich domain-containing (SRCR) gene family, many of which are also differentially expressed in choanocytes; these are putatively involved in microbe-associated molecular patterns recognition [59,60,61]. These large complements suggest that this morphologically-simple animal without an apparent adaptive immune system could have the capacity to distinguish and subsequently generate specific responses to foreign and symbiotic bacteria [62]. Consistent with this premise, these three choanocyte-enriched lncRNAs were previously found to be co-expressed with protein-coding genes enriched for scavenger receptor activity [20] and up-regulated when Amphimedon juveniles were exposed to a foreign bacterial suspension belonging to a different sponge species (Rhabdastrella globostellata, Carter 1883) [59]. Together, these findings suggest a putative role for these three lncRNAs in innate immunity in Amphimedon.
Analysis of AmqTCONS_00003141 cell-type expression profile revealed its upregulation in archeocytes and pinacocytes. This cis-antisense lncRNA is also co-expressed with protein-coding genes involved in key intercellular signaling pathways, including the G-protein-coupled receptor Frizzled-B (UniProt: I1G9T3_AMPQE) and TGF-β receptor type-1 (National Center for Biotechnology Information (NCBI) Reference Sequence: XP_011409575.1) [20]. Consistent with this, genes encoding TGF-β, a major immunosuppressive cytokine with a highly-conserved role in metazoan immunity [63,64] and development [65], are also differentially enriched in pinacocytes [59].
2.2. Sponge lncRNAs Show Cell Type-Specific Restricted Expression Patterns
To further validate the cell-type-specific expression of Amphimedon lncRNAs, we selected three independently regulated lncRNA transcripts with different developmental expression profiles for in situ hybridization (ISH) analysis; one upregulated in metamorphosing postlarvae (AmqTCONS_00003141), one upregulated in feeding 3-day old juveniles (AmqTCON_00001029) and one upregulated in larvae (AmqTCONS_00000018) (Figure 2).
AmqTCONS_00003141 is activated 6–7 h after settlement and commencement of metamorphosis, and remains highly expressed as the juvenile body plan is forming (Figure 2A). Its transcripts were localized to subsets of specific internal cells of late postlarvae and juveniles (oscula stage), and not detected in the outer epithelial layer. Specifically, they were detected in a subset of choanocytes comprising newly-formed feeding chambers (Figure 2Ai,Ai’); ISH reveals only a fraction of choanocytes at these stages have detectable expression of AmqTCONS_00003141. Although highly expressed in choanocytes (i.e., belonging to quartile Q4; Table S6), this variability in expression might explain the absence of this transcript in the differentially expressed choanocyte-specific CEL-seq dataset (Table S5). At this stage, choanocytes chambers (as shown in Figure 2Ai,Ai’) typically contain proliferating cells and choanocytes from these chambers can rapidly dedifferentiate into archeocytes [66]. Consistent with some archeocytes being derived from dedifferentiating choanocytes, AmqTCONS_00003141 transcripts were also detected in clusters of archeocytes, which are larger and often form migratory streams (Figure 2Aii,Aii’).
The previously characterized long intergenic ncRNA (lincRNA) AmqTCONS_00001029 is a 526-nt transcript encoded by three exons, expressed from chamber (late postlarval) to adult stages [67] (Figure 2B). In contrast to AmqTCONS_00003141, its transcripts were detected in epithelial cells—endopinacocytes—that line the internal network of canals (Figure 2Bi,Bii’).
The remaining lincRNA (AmqTCONS_00000018), a 959-nt transcript encoded by two exons, was expressed in larval stages right before settlement (Figure 2C). Amphimedon larva has three cells layers; an outer epithelial layer interspersed with globular cells and flask cells, a subepithelial layer composed mostly of large globular, and the inner cell mass [68]. AmqTCONS_00000018 transcripts were detected in subepithelial cells at the boundary between outer cell layer and inner cell mass (Figure 2Ci,Cii).
Together, as in bilaterians [19,21,22,69], Amphimedon lncRNAs are expressed in spatially- and cell type-restricted expression patterns, encompassing multiple cell types, consistent with lncRNA expression during animal development being highly dynamic [18,20] and tightly regulated to a specific developmental and cellular context. Although functional evidence currently is lacking, the restricted expression of these sponge lncRNAs in specific cell types during development, as observed in other animals [70], suggests that these non-coding genes are part of regulatory network(s) in Amphimedon.
2.3. Amphimedon and Sycon lncRNAs Are Co-Expressed with Similar Sets of Protein-Coding Genes
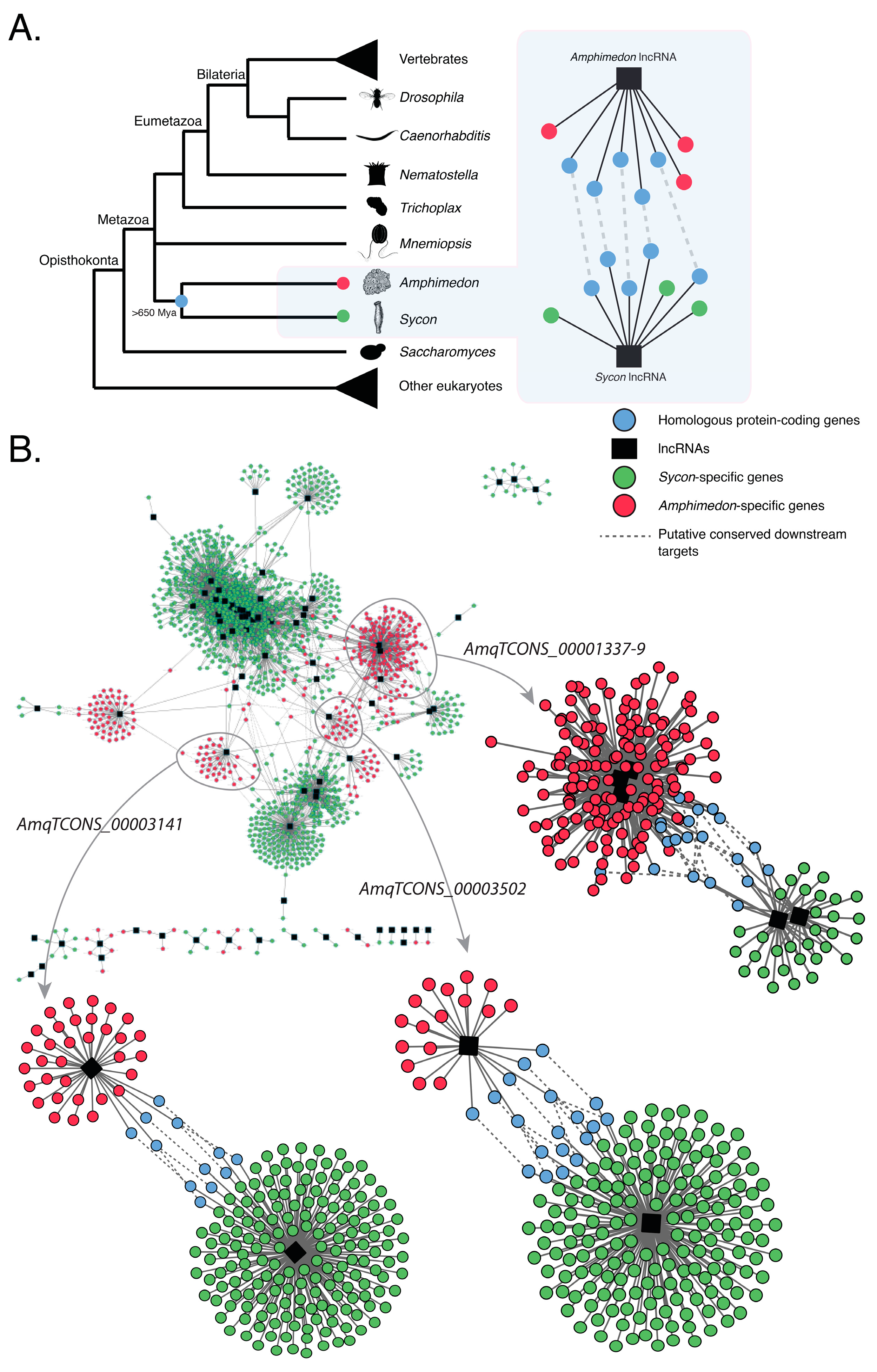
Previous findings have shown that Amphimedon and Sycon lncRNAs are co-expressed with similar sets of protein-coding genes [18,20], suggesting that, despite showing no apparent homology with any known animal lncRNAs, sponge lncRNAs may operate in evolutionarily conserved developmental modules (or networks) (Figure 3A).
To further document this correlation, we focused on the differentially expressed lncRNAs that strongly correlated with the expression profiles of sets of protein-coding genes involved in key animal developmental processes in the sponges Amphimedon and Sycon [18,20]. These two sponges are estimated to have diverged from each other at least 650 Mya [45]. We then constructed co-expression networks (modules), as a proxy for gene regulatory networks, based on these previously identified differentially expressed genes (both coding genes and lncRNAs) in both species [18,20]. These co-expression networks all have lncRNAs in central nodes, suggesting a key regulatory role for these lncRNAs (Figure S2).
Co-expression networks that consist of homologous protein-coding genes between Amphimedon and Sycon and differentially expressed lncRNAs were deemed to be conserved in sponges (Figure 3B). One such example is comprised of either developmental Sycon lncRNAs [18] or Amphimedon lncRNAs AmqTCONS_1337-9, AmqTCONS_3502, and AmqTCONS_0003141 [20], which have no sequence similarity, and a similar set of homologous protein-coding genes (e.g., TGF-β receptor type-1) in both species (Figure 3B; Table 1; Tables S7 and S8).
The co-expression of lncRNAs with homologous coding genes in these sponges suggests these non-coding genes may operate in evolutionarily conserved co-expression networks. Alternatively, given there is no discernible sequence identity between these sponge lncRNAs and currently a lack of functional studies, it is also plausible that these putative co-regulatory networks have evolved independently in Sycon and Amphimedon, with lncRNAs being co-opted separately into homologous protein-coding networks.
3. Conclusions
The dynamic, cell type- and context-specific expression of sponge lncRNAs (this study; [18,20]) is consistent with spatiotemporal expression features of bilaterian lncRNAs also being present in sponges. The expression and possible function of lncRNAs during development can, therefore, be inferred to be present in the last common ancestor of these two lineages. Although currently there is a lack of functional data in sponges, lncRNAs appear to play a role in sponge development by regulating the deployment of various cell differentiation gene batteries as observed in bilaterians [70,71,72,73,74,75]. Given the lack of sequence identity of lncRNAs, it remains unclear if developmental sponge lncRNAs are conserved or independently-evolved. As gene regulatory networks and modules are central for the control and timing of animal development [76,77,78], the finding of similar sets of homologous protein-coding genes co-expressed and, thus possibly co-regulated, with lncRNAs between evolutionarily divergent sponge species, suggests lncRNAs may be playing important roles in these putative conserved gene regulatory networks.
4. Materials and Methods
4.1. Cell-Type Specific Transcriptome Analysis
A total of 39 samples were used from three adult A. queenslandica (5 archeocyte, 5 choanocyte and 3 pinacocyte samples from each adult individual). CEL-seq reads [58] from these samples were mapped back to the A. queenslandica genome [79] using Bowtie2 [80] and the CEL-seq analysis pipeline as previously described [81]. An average of ~9 million reads per sample were obtained, with an average of 60% of the reads mapped onto the Amphimedon genome [79]. All samples with less than 1 million reads formed their own cluster in the preliminary principal component analysis (PCA) using DESeq2 [82] and were therefore discarded from further analyses, resulting in a total of 31 samples (15 archeocyte, 10 choanocyte and 6 pinacocyte samples) used in this study (Table S1). The final counts were analyzed for differential gene expression using DESeq2 [82]. Pairwise comparisons were conducted between each of the three cell types to generate a list of differentially expressed genes for each cell type (Tables S2–S4). A 5% False Discovery Rate cut-off was used to produce the final lists of differentially expressed lncRNAs and protein-coding genes (Table S5). To investigate the full repertoire of lncRNAs expressed (in contrast with differentially expressed) in each cell type, the lncRNAs were also divided into expression quartiles. All the zero count reads were discarded and the median expression value of the non-transformed normalized count values of all samples (from all cell types) were used to calculate the quartile values. These values were used to classify the expression of all the lncRNAs in each cell type into four quartiles, ranging from low (Q1) to highly (Q4) expressed overall (Table S6).
4.2. Gene Isolation and Whole Mount In Situ Hybridization
Amphimedon lncRNA fragments were amplified with gene specific primers, by using complimentary DNA from mixed developmental stages as a polymerase chain reaction (PCR) template. Gene specific primers were as follows: AmqTCONS_00003141_Fw, ATAGGACCCACCCAGTCAAAC and AmqTCONS_00003141_Rev, TTCCTTGTTGTTCCTTGCCCT; AmqTCONS_00001029_Fw, AGA ATTGGCCGTAACAACAAGT and AmqTCONS_00001029_Rev, TCTAAGAAAATCTAAGTTACGTGTACG; AmqTCONS_00000018_Fw, TCCATTCCTATATTTTCCCCTTC and AmqTCONS_00000018_Rev, ATGAGGGTGGGATGATGTGC. The fragments were cloned into pGEM-T Easy (Promega, Fitchburg, Wisconsin, USA) vector using the manufacturer’s protocol and verified by sequencing using M13F and M13R primers. Digoxygenin (DIG)-labelled antisense RNA probes were transcribed from PCR products using DIG RNA Labeling Mix (Roche, Basel, Switzerland) and T7 or SP6 Polymerase (Promega, Fitchburg, Wisconsin, USA) following the manufacturer’s instructions. Whole mount ISH analysis of larval and juvenile gene expression was carried out as described previously [83]. Antisense DIG-labelled riboprobes were hybridized at a final concentration of 1 ng/μL. Whole-mount samples were observed under an Olympus SZX7 or a Nikon eclipse Ti microscope (Olympus Australia Pty Ltd., Mt Waverly, VIC, Australia) and photographed with a Nikon Sight DS-U1 camera (Nikon Australia Pty Ltd., Lidcombe, NSW, Australia).
4.3. Co-Expression Network Analysis
Co-expression networks were constructed based on the previously identified differentially expressed genes (coding genes and lncRNAs) in both Amphimedon [20] and Sycon [18]. Co-expression analysis in both species was performed as previously described (Gaiti et al., 2015). Co-expression networks were visualized using Cytoscape [84]. These networks show genes co-expressed with lncRNAs, where nodes indicate differentially expressed coding-genes, hubs indicate lncRNAs, and edges represent a significant co-expression (both positive ≥ 0.95 and negative ≤ −0.95) (p-value < 0.05). Homology between Sycon and Amphimedon was inferred with BLAST+ (version 2.2.30) [85], using BLASTp (e-value cutoff < 1 × 10−5) against a custom all vs. all database containing all Amphimedon Aqu2.1 peptides [86] and all peptides identified in the Sycon transcriptome [18] using TransDecoder (recommended settings, guided by UniProt and Pfam-A databases) [87]. Putative “evolutionarily conserved” modules were defined as modules containing at least one homologue between species.
4.4. Data Access
Amphimedon cell-type specific CEL-seq datasets can be obtained from NCBI under accession number PRJNA412708 [58]. Amphimedon genome assembly ampQue1 was used throughout the study. Developmental CEL-seq datasets used can be obtained from NCBI Gene Expression Omnibus (GEO) [88] under accession number GSE54364 [89]. The following gene model datasets were used for all analyses. A. queenslandica: Aqu2.1 models [86], lncRNAs [20]. S. ciliatum: coding genes and lncRNAs [18]. The codes used for the gene co-expression analysis are available for download at [20].
Supplementary Materials
The following are available online at https://www.mdpi.com/2311-553X/4/1/6/s1, Figure S1: Schematic representation of the evolutionary relationship among representative sponge species. Blue box highlights the sponge clade. Amphimedon queenslandica and Sycon ciliatum, the sponge species used in this study, are shown in bold red. For detailed phylogenetic relationship analyses of sponges please refer to [46,47,48,52]. Adapted from [20]. Figure S2: Co-expression networks of Amphimedon and Sycon lncRNAs. Co-expression networks are based on the previously identified differentially expressed genes (coding genes and lncRNAs) in Amphimedon [20] and Sycon [18]. Nodes indicate coding-genes, hubs (black) represent lncRNAs, and edges represent significant co-expression (both positive and negative). Table S1: Cell-type specific CEL-seq normalized read counts of Amphimedon lncRNAs and protein-coding genes across all samples. Table S2: Differential gene expression of Amphimedon lncRNAs and protein-coding genes between archeocyte and choanocyte. Only genes that passed our technical cut-off of a 5% False Discovery Rate (FDR) are shown. Table S3: Differential gene expression of Amphimedon lncRNAs and protein-coding genes between archeocyte and pinacocyte. Only genes that passed our technical cut-off of a 5% False Discovery Rate (FDR) are shown. Table S4: Differential gene expression of Amphimedon lncRNAs and protein-coding genes between choanocyte and pinacocyte. Only genes that passed our technical cut-off of a 5% False Discovery Rate (FDR) are shown. Table S5: Final lists of differentially expressed Amphimedon lncRNAs and protein-coding genes for each cell type at a 5% FDR (Q < 0.05). The examples provided in the main text are highlighted in yellow. Table S6: The number of lncRNAs expressed in each cell type for each quarter and combined quarters, using expression quartile values (Q1: 1.45, Q2: 1.95, Q3: 3.11). Table S7: Node tables for the three specific examples of evolutionarily conserved modules of co-expressed coding genes and lncRNAs between sponges shown in Figure 3. Table S8: Edge tables for the three specific examples of evolutionarily conserved modules of co-expressed coding genes and lncRNAs between sponges shown in Figure 3. Nodes are marked as ‘name’ and edges are listed as ‘interaction’.
Supplementary File 1Acknowledgments
We thank Shunsuke Sogabe for help with the cell-type specific transcriptome analysis, and Degnan lab members for constructive discussions. This work was supported by an Australian Research Council grant (FL110100044) to B.M.D.
Author Contributions
F.G., M.T. and B.M.D. designed the study. F.G. and W.L.H. carried out the computational analyses. F.G. performed the gene isolation and whole mount ISH. F.G. wrote the manuscript with critical comments provided by M.T. and B.M.D. All authors discussed the results, commented and approved the final manuscript.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Bertone, P.; Stolc, V.; Royce, T.E.; Rozowsky, J.S.; Urban, A.E.; Zhu, X.; Rinn, J.L.; Tongprasit, W.; Samanta, M.; Weissman, S.; et al. Global identification of human transcribed sequences with genome tiling arrays. Science 2004, 306, 2242–2246. [Google Scholar] [CrossRef] [PubMed]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [PubMed]
- Carninci, P.; Kasukawa, T.; Katayama, S.; Gough, J.; Frith, M.C.; Maeda, N.; Oyama, R.; Ravasi, T.; Lenhard, B.; Wells, C.; et al. The transcriptional landscape of the mammalian genome. Science 2005, 309, 1559–1563. [Google Scholar] [PubMed]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long non-coding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Russell, P.; Ingolia, N.T.; Weissman, J.S.; Lander, E.S. Ribosome profiling provides evidence that large noncoding RNAs do not encode proteins. Cell 2013, 154, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Housman, G.; Ulitsky, I. Methods for distinguishing between protein-coding and long noncoding RNAs and the elusive biological purpose of translation of long noncoding RNAs. Biochim. Biophys. Acta Gene Regul. Mech. 2016, 1859, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Ingolia, N.T.; Brar, G.A.; Stern-Ginossar, N.; Harris, M.S.; Talhouarne, G.J.S.; Jackson, S.E.; Wills, M.R.; Weissman, J.S. Ribosome profiling reveals pervasive translation outside of annotated protein-coding genes. Cell Rep. 2014, 8, 1365–1379. [Google Scholar] [CrossRef] [PubMed]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermuller, J.; Hofacker, I.L.; et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Rivea Morales, D.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, J.; Uszczynska-Ratajczak, B.; Carbonell, S.; Pérez-Lluch, S.; Abad, A.; Davis, C.; Gingeras, T.R.; Frankish, A.; Harrow, J.; Guigo, R.; et al. High-throughput annotation of full-length long noncoding RNAs with capture long-read sequencing. Nat. Genet. 2017, 49, 1731. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; RIKEN Genome Exploration Research Group Phase I & II Team. Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs. Nature 2002, 420, 563–573. [Google Scholar] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long non-coding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Ravasi, T.; Suzuki, H.; Pang, K.C.; Katayama, S.; Furuno, M.; Okunishi, R.; Fukuda, S.; Ru, K.; Frith, M.C.; Gongora, M.M.; et al. Experimental validation of the regulated expression of large numbers of non-coding RNAs from the mouse genome. Genome Res. 2006, 16, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Nowakowski, T.J.; Pollen, A.A.; Lui, J.H.; Horlbeck, M.A.; Attenello, F.J.; He, D.; Weissman, J.S.; Kriegstein, A.R.; Diaz, A.A.; et al. Single-cell analysis of long non-coding RNAs in the developing human neocortex. Genome Biol. 2016, 17, 67. [Google Scholar] [CrossRef] [PubMed]
- Bråte, J.; Adamski, M.; Neumann, R.S.; Shalchian-Tabrizi, K.; Adamska, M. Regulatory RNA at the root of animals: Dynamic expression of developmental lincRNAs in the calcisponge Sycon ciliatum. Proc. R. Soc. Lond. B Biol. Sci. 2015, 282, 20151746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabili, M.N.; Dunagin, M.C.; McClanahan, P.D.; Biaesch, A.; Padovan-Merhar, O.; Regev, A.; Rinn, J.L.; Raj, A. Localization and abundance analysis of human lncRNAs at single-cell and single-molecule resolution. Genome Biol. 2015, 16, 20. [Google Scholar] [CrossRef] [PubMed]
- Gaiti, F.; Fernandez-Valverde, S.L.; Nakanishi, N.; Calcino, A.D.; Yanai, I.; Tanurdzic, M.; Degnan, B.M. Dynamic and widespread lncRNA expression in a sponge and the origin of animal complexity. Mol. Biol. Evol. 2015, 32, 2367–2382. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Dinger, M.E.; Sunkin, S.M.; Mehler, M.F.; Mattick, J.S. Specific expression of long noncoding RNAs in the mouse brain. Proc. Natl. Acad. Sci. USA 2008, 105, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Pauli, A.; Valen, E.; Lin, M.F.; Garber, M.; Vastenhouw, N.L.; Levin, J.Z.; Fan, L.; Sandelin, A.; Rinn, J.L.; Regev, A.; et al. Systematic identification of long non-coding RNAs expressed during zebrafish embryogenesis. Genome Res. 2012, 22, 577–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponjavic, J.; Oliver, P.L.; Lunter, G.; Ponting, C.P. Genomic and transcriptional co-localization of protein-coding and long non-coding RNA pairs in the developing brain. PLoS Genet. 2009, 5, e1000617. [Google Scholar] [CrossRef] [PubMed]
- Zappulo, A.; van den Bruck, D.; Ciolli Mattioli, C.; Franke, V.; Imami, K.; McShane, E.; Moreno-Estelles, M.; Calviello, L.; Filipchyk, A.; Peguero-Sanchez, E.; et al. RNA localization is a key determinant of neurite-enriched proteome. Nat. Commun. 2017, 8, 583. [Google Scholar] [CrossRef] [PubMed]
- Pauli, A.; Rinn, J.L.; Schier, A.F. Non-coding RNAs as regulators of embryogenesis. Nat. Rev. Genet. 2011, 12, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Ulitsky, I. Evolution to the rescue: Using comparative genomics to understand long non-coding RNAs. Nat. Rev. Genet. 2016, 17, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.B.; Boley, N.; Eisman, R.; May, G.E.; Stoiber, M.H.; Duff, M.O.; Booth, B.W.; Wen, J.; Park, S.; Suzuki, A.M.; et al. Diversity and dynamics of the Drosophila transcriptome. Nature 2014, 512, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zhang, Y.; Zhang, X.; Jia, S.; Chen, S.; Kang, L. Genome-wide identification and developmental expression profiling of long noncoding RNAs during Drosophila metamorphosis. Sci. Rep. 2016, 6, 23330. [Google Scholar] [CrossRef] [PubMed]
- Forouzmand, E.; Owens, N.D.L.; Blitz, I.L.; Paraiso, K.D.; Khokha, M.K.; Gilchrist, M.J.; Xie, X.; Cho, K.W.Y. Developmentally regulated long non-coding RNAs in Xenopus tropicalis. Dev. Biol. 2017, 426, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Morlighem, J.-É.R.L.; Cai, J.; Liao, Q.; Perez, C.D.; Gomes, P.B.; Guo, M.; Rádis-Baptista, G.; Lee, S.M.-Y. Identification of long non-coding RNAs in two anthozoan species and their possible implications for coral bleaching. Sci. Rep. 2017, 7, 5333. [Google Scholar] [CrossRef] [PubMed]
- Jayakodi, M.; Jung, J.W.; Park, D.; Ahn, Y.-J.; Lee, S.-C.; Shin, S.-Y.; Shin, C.; Yang, T.-J.; Kwon, H.W. Genome-wide characterization of long intergenic non-coding RNAs (lincRNAs) provides new insight into viral diseases in honey bees Apis cerana and Apis mellifera. BMC Genom. 2015, 16, 680. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, A.M.; Waterhouse, R.M.; Muskavitch, M.A.T. Long non-coding RNA discovery across the genus anopheles reveals conserved secondary structures within and beyond the Gambiae complex. BMC Genom. 2015, 16, 337. [Google Scholar] [CrossRef] [PubMed]
- Mu, C.; Wang, R.; Li, T.; Li, Y.; Tian, M.; Jiao, W.; Huang, X.; Zhang, L.; Hu, X.; Wang, S.; et al. Long non-coding RNAs (lncRNAs) of sea cucumber: Large-scale prediction, expression profiling, non-coding network construction, and lncRNA-microRNA-gene interaction analysis of lncRNAs in Apostichopus japonicus and Holothuria glaberrima during LPS challenge and radial organ complex regeneration. Mar. Biotechnol. 2016, 18, 485–499. [Google Scholar] [PubMed]
- Nam, J.W.; Bartel, D.P. Long noncoding RNAs in C. elegans. Genome Res. 2012, 22, 2529–2540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Necsulea, A.; Soumillon, M.; Warnefors, M.; Liechti, A.; Daish, T.; Zeller, U.; Baker, J.C.; Grutzner, F.; Kaessmann, H. The evolution of lncRNA repertoires and expression patterns in tetrapods. Nature 2014, 505, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Zhang, Q.C.; Georgiev, P.; Ilik, I.A.; Akhtar, A.; Chang, H.Y. Rapid evolutionary turnover underlies conserved lncRNA–genome interactions. Genes Dev. 2016, 30, 191–207. [Google Scholar] [CrossRef] [PubMed]
- Sauvageau, M.; Goff, L.A.; Lodato, S.; Bonev, B.; Groff, A.F.; Gerhardinger, C.; Sanchez-Gomez, D.B.; Hacisuleyman, E.; Li, E.; Spence, M.; et al. Multiple knockout mouse models reveal lincRNAs are required for life and brain development. eLife 2013, 2, e01749. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.H.; Au, K.F.; Yablonovitch, A.L.; Wills, A.E.; Chuang, J.; Baker, J.C.; Wong, W.H.; Li, J.B. RNA sequencing reveals a diverse and dynamic repertoire of the Xenopus tropicalis transcriptome over development. Genome Res. 2013, 23, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Ulitsky, I.; Shkumatava, A.; Jan, C.H.; Sive, H.; Bartel, D.P. Conserved function of lincRNAs in vertebrate embryonic development despite rapid sequence evolution. Cell 2011, 147, 1537–1550. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Cheng, T.; Liu, C.; Liu, D.; Zhang, Q.; Long, R.; Zhao, P.; Xia, Q. Systematic identification and characterization of long non-coding RNAs in the silkworm, Bombyx mori. PLoS ONE 2016, 11, e0147147. [Google Scholar] [CrossRef] [PubMed]
- Young, R.S.; Marques, A.C.; Tibbit, C.; Haerty, W.; Bassett, A.R.; Liu, J.L.; Ponting, C.P. Identification and properties of 1119 candidate lincRNA loci in the Drosophila melanogaster genome. Genome Biol. Evol. 2012, 4, 427–442. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.; Mahadevaiah, S.K.; Khil, P.; Sangrithi, M.N.; Royo, H.; Duckworth, J.; McCarrey, J.R.; VandeBerg, J.L.; Renfree, M.B.; Taylor, W.; et al. Rsx is a metatherian RNA with Xist-like properties in X-chromosome inactivation. Nature 2012, 487, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Heard, E.; Mongelard, F.; Arnaud, D.; Chureau, C.; Vourc’h, C.; Avner, P. Human XIST yeast artificial chromosome transgenes show partial X inactivation center function in mouse embryonic stem cells. Proc. Natl. Acad. Sci. USA 1999, 96, 6841–6846. [Google Scholar] [CrossRef] [PubMed]
- Migeon, B.R.; Kazi, E.; Haisley-Royster, C.; Hu, J.; Reeves, R.; Call, L.; Lawler, A.; Moore, C.S.; Morrison, H.; Jeppesen, P. Human X inactivation center induces random X chromosome inactivation in male transgenic mice. Genomics 1999, 59, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Erwin, D.H.; Laflamme, M.; Tweedt, S.M.; Sperling, E.A.; Pisani, D.; Peterson, K.J. The Cambrian conundrum: Early divergence and later ecological success in the early history of animals. Science 2011, 334, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Gazave, E.; Lapébie, P.; Ereskovsky, A.V.; Vacelet, J.; Renard, E.; Cárdenas, P.; Borchiellini, C. No longer Demospongiae: Homoscleromorpha formal nomination as a fourth class of Porifera. Hydrobiologia 2012, 687, 3–10. [Google Scholar] [CrossRef]
- Hill, M.S.; Hill, A.L.; Lopez, J.; Peterson, K.J.; Pomponi, S.; Diaz, M.C.; Thacker, R.W.; Adamska, M.; Boury-Esnault, N.; Cardenas, P.; et al. Reconstruction of family-level phylogenetic relationships within Demospongiae (Porifera) using nuclear encoded housekeeping genes. PLoS ONE 2013, 8, e50437. [Google Scholar] [CrossRef] [PubMed]
- Worheide, G.; Dohrmann, M.; Erpenbeck, D.; Larroux, C.; Maldonado, M.; Voigt, O.; Borchiellini, C.; Lavrov, D.V. Deep phylogeny and evolution of sponges (phylum Porifera). Adv. Mar. Biol. 2012, 61, 1–78. [Google Scholar] [PubMed]
- Degnan, B.M.; Adamska, M.; Richards, G.S.; Larroux, C.; Leininger, S.; Bergum, B.; Calcino, A.; Taylor, K.; Nakanishi, N.; Degnan, S.M. Porifera. In Evolutionary Developmental Biology of Invertebrates 1: Introduction, Non-Bilateria, Acoelomorpha, Xenoturbellida, Chaetognatha; Wanninger, A., Ed.; Springer: Vienna, Austria, 2015; pp. 65–106. [Google Scholar]
- Ereskovsky, A.V. The Comparative Embryology of Sponges; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2010. [Google Scholar]
- Leys, S.P.; Degnan, B.M. Embryogenesis and metamorphosis in a haplosclerid demosponge: Gastrulation and transdifferentiation of larval ciliated cells to choanocytes. Invertebr. Biol. 2002, 121, 171–189. [Google Scholar] [CrossRef]
- Simion, P.; Philippe, H.; Baurain, D.; Jager, M.; Richter, D.J.; Di Franco, A.; Roure, B.; Satoh, N.; Quéinnec, É.; Ereskovsky, A.; et al. A large and consistent phylogenomic dataset supports sponges as the sister group to all other animals. Curr. Biol. 2017, 27, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Whelan, N.V.; Kocot, K.M.; Moroz, T.P.; Mukherjee, K.; Williams, P.; Paulay, G.; Moroz, L.L.; Halanych, K.M. Ctenophore relationships and their placement as the sister group to all other animals. Nat. Ecol. Evol. 2017, 1, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, R.M.; Haussler, D.; Kent, W.J. The UCSC genome browser and associated tools. Brief. Bioinform. 2013, 14, 144–161. [Google Scholar] [CrossRef] [PubMed]
- Chodroff, R.A.; Goodstadt, L.; Sirey, T.M.; Oliver, P.L.; Davies, K.E.; Green, E.D.; Molnar, Z.; Ponting, C.P. Long noncoding RNA genes: Conservation of sequence and brain expression among diverse amniotes. Genome Boil. 2010, 11, R72. [Google Scholar] [CrossRef] [PubMed]
- Hezroni, H.; Koppstein, D.; Schwartz, M.G.; Avrutin, A.; Bartel, D.P.; Ulitsky, I. Principles of long noncoding RNA evolution derived from direct comparison of transcriptomes in 17 species. Cell Rep. 2015, 11, 1110–1122. [Google Scholar] [CrossRef] [PubMed]
- Washietl, S.; Kellis, M.; Garber, M. Evolutionary dynamics and tissue specificity of human long noncoding RNAs in six mammals. Genome Res. 2014, 24, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Sogabe, S. The Biology of Choanocytes and Choanocyte Chambers and Their Role in the Sponge Stem Cell System. Ph.D. Thesis, School of Biological Sciences, The University of Queensland, Brisbane, Australia, 2017. [Google Scholar]
- Yuen, B. Deciphering the Genomic Tool-Kit Underlying Animal-Bacteria Interactions: Insights through the Demosponge Amphimedon queenslandica. Ph.D. Thesis, School of Biological Sciences, The University of Queensland, Brisbane, Australia, 2016. [Google Scholar]
- Ryu, T.; Seridi, L.; Moitinho-Silva, L.; Oates, M.; Liew, Y.J.; Mavromatis, C.; Wang, X.; Haywood, A.; Lafi, F.F.; Kupresanin, M.; et al. Hologenome analysis of two marine sponges with different microbiomes. BMC Genom. 2016, 17, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuen, B.; Bayes, J.M.; Degnan, S.M. The characterization of sponge NLRs provides insight into the origin and evolution of this innate immune gene family in animals. Mol. Biol. Evol. 2014, 31, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Degnan, S.M. The surprisingly complex immune gene repertoire of a simple sponge, exemplified by the NLR genes: A capacity for specificity? Dev. Comp. Immunol. 2015, 48, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Detournay, O.; Schnitzler, C.E.; Poole, A.; Weis, V.M. Regulation of cnidarian–dinoflagellate mutualisms: Evidence that activation of a host TGF-β innate immune pathway promotes tolerance of the symbiont. Dev. Comp. Immunol. 2012, 38, 525–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, C.J.C.; Smyth, D.J.; Dresser, D.W.; Maizels, R.M. TGF-β in tolerance, development and regulation of immunity. Cell. Immunol. 2016, 299, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Adamska, M.; Degnan, S.M.; Green, K.M.; Adamski, M.; Craigie, A.; Larroux, C.; Degnan, B.M. Wnt and TGF-beta expression in the sponge Amphimedon queenslandica and the origin of metazoan embryonic patterning. PLoS ONE 2007, 2, e1031. [Google Scholar] [CrossRef] [PubMed]
- Sogabe, S.; Nakanishi, N.; Degnan, B.M. The ontogeny of choanocyte chambers during metamorphosis in the demosponge Amphimedon queenslandica. EvoDevo 2016, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Gaiti, F.; Jindrich, K.; Fernandez-Valverde, S.L.; Roper, K.E.; Degnan, B.M.; Tanurdzic, M. Landscape of histone modifications in a sponge reveals the origin of animal cis-regulatory complexity. eLife 2017, 6, e22194. [Google Scholar] [CrossRef] [PubMed]
- Richards, G.S.; Degnan, B.M. The expression of Delta ligands in the sponge Amphimedon queenslandica suggests an ancient role for Notch signaling in metazoan development. EvoDevo 2012, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Goff, L.A.; Groff, A.F.; Sauvageau, M.; Trayes-Gibson, Z.; Sanchez-Gomez, D.B.; Morse, M.; Martin, R.D.; Elcavage, L.E.; Liapis, S.C.; Gonzalez-Celeiro, M.; et al. Spatiotemporal expression and transcriptional perturbations by long non-coding RNAs in the mouse brain. Proc. Natl. Acad. Sci. USA 2015, 112, 6855–6862. [Google Scholar] [CrossRef] [PubMed]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Chalei, V.; Sansom, S.N.; Kong, L.; Lee, S.; Montiel, J.F.; Vance, K.W.; Ponting, C.P. The long non-coding RNA Dali is an epigenetic regulator of neural differentiation. eLife 2014, 3, e04530. [Google Scholar] [CrossRef] [PubMed]
- Dinger, M.E.; Amaral, P.P.; Mercer, T.R.; Pang, K.C.; Bruce, S.J.; Gardiner, B.B.; Askarian-Amiri, M.E.; Ru, K.; Solda, G.; Simons, C.; et al. Long non-coding RNAs in mouse embryonic stem cell pluripotency and differentiation. Genome Res. 2008, 18, 1433–1445. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Donaghey, J.; Carey, B.W.; Garber, M.; Grenier, J.K.; Munson, G.; Young, G.; Lucas, A.B.; Ach, R.; Bruhn, L.; et al. lincRNAs act in the circuitry controlling pluripotency and differentiation. Nature 2011, 477, 295–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Lu, J.Y.; Liu, L.; Yin, Y.; Chen, C.; Han, X.; Wu, B.; Xu, R.; Liu, W.; Yan, P.; et al. Divergent lncRNAs regulate gene expression and lineage differentiation in pluripotent cells. Cell Stem Cell 2016, 18, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Yan, P.; Lu, J.; Song, G.; Zhu, Y.; Li, Z.; Zhao, Y.; Shen, B.; Huang, X.; Zhu, H.; et al. Opposing roles for the lncRNA Haunt and its genomic locus in regulating HoxA gene activation during embryonic stem cell differentiation. Cell Stem Cell 2015, 16, 504–516. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.H.; Erwin, D.H. Gene regulatory networks and the evolution of animal body plans. Science 2006, 311, 796–800. [Google Scholar] [CrossRef] [PubMed]
- Erwin, D.H.; Davidson, E.H. The evolution of hierarchical gene regulatory networks. Nat. Rev. Genet. 2009, 10, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Peter, I.S.; Davidson, E.H. Evolution of gene regulatory networks controlling body plan development. Cell 2011, 144, 970–985. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 2010, 466, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Hashimshony, T.; Wagner, F.; Sher, N.; Yanai, I. CEL-Seq: Single-cell RNA-Seq by multiplexed linear amplification. Cell Rep. 2012, 2, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Larroux, C.; Fahey, B.; Adamska, M.; Richards, G.S.; Gauthier, M.; Green, K.; Lovas, E.; Degnan, B.M. Whole-mount in situ hybridization in Amphimedon. Cold Spring Harbor Protoc. 2008. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Fernandez-Valverde, S.L.; Calcino, A.D.; Degnan, B.M. Deep developmental transcriptome sequencing uncovers numerous new genes and enhances gene annotation in the sponge Amphimedon queenslandica. BMC Genom. 2015, 16, 387. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-Seq: Reference generation and analysis with Trinity. Nat. Protoc. 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Anavy, L.; Levin, M.; Khair, S.; Nakanishi, N.; Fernandez-Valverde, S.L.; Degnan, B.M.; Yanai, I. BLIND ordering of large-scale transcriptomic developmental timecourses. Development 2014, 141, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Amphimedon queenslandica long non-coding RNAs (lncRNAs) are enriched in specific cell types. (A) Phylogenetic tree of selected species with well-described genomes. Yellow background highlights the animal kingdom. The position of Amphimedon queenslandica and Mnemiopsis leidyi is indicated as a polytomy given the current debate on the branching order of poriferan and ctenophore lineages [52,53]; (B) Schematic representation of Amphimedon queenslandica life cycle. Larvae (oval shaped, 400–600 µm long) emerge from maternal brood chambers and then swim in the water column before they develop competence to settle and initiate metamorphosis into a juvenile. The juvenile body plan, which displays the hallmarks of the adult body plan, including an aquiferous system with canals, choanocytes chambers, and oscula, is the outcome of the dramatic reorganization of the radially-symmetrical, bi- or trilayered larva. This juvenile will then grow and mature into a benthic adult (ranging from 10–30 cm3) [51]; (C) Diagram of a juvenile sponge body plan. Water flows into the internal aquiferous system via the ostium and out via the osculum. The mesohyl is shown in blue and populated by archeocytes and other cell types, including sclerocytes and spherulous cells. Adapted from [49]; (D) Optical section of a 3-day-old Amphimedon queenslandica juvenile showing internal morphology and some cell types. Archeocyte (a), choanocyte chamber (cc), endopinacoderm (en), exopinacoderm (ex), ostium (o), osculum (os), sclerocyte (s), spicule (sp), and spherulous cell (sph). Scale bar: 10 μm. Adapted from [49]; (E) Venn diagram denoting the proportion of differentially expressed lncRNAs detected in each of the three cell-type specific transcriptome datasets; (F) Subset of Amphimedon lncRNAs (AmqTCONS_00001337-9) are in close genomic proximity to a cluster of immune-related genes. Coding genes (purple) and long non-coding RNAs (blue) are shown, along with signal coverage tracks showing CEL-seq expression. A grey scale indicates CEL-seq (Cell Expression by Linear amplification and sequencing) expression level: white (no-expression); black (highest expression). The shaded purple area represents the cluster of immune-related genes [Tnf receptor-associated factors (TRAFs)]. Figure was generated using a local instance of the UCSC genome browser [54].
Figure 1.
Amphimedon queenslandica long non-coding RNAs (lncRNAs) are enriched in specific cell types. (A) Phylogenetic tree of selected species with well-described genomes. Yellow background highlights the animal kingdom. The position of Amphimedon queenslandica and Mnemiopsis leidyi is indicated as a polytomy given the current debate on the branching order of poriferan and ctenophore lineages [52,53]; (B) Schematic representation of Amphimedon queenslandica life cycle. Larvae (oval shaped, 400–600 µm long) emerge from maternal brood chambers and then swim in the water column before they develop competence to settle and initiate metamorphosis into a juvenile. The juvenile body plan, which displays the hallmarks of the adult body plan, including an aquiferous system with canals, choanocytes chambers, and oscula, is the outcome of the dramatic reorganization of the radially-symmetrical, bi- or trilayered larva. This juvenile will then grow and mature into a benthic adult (ranging from 10–30 cm3) [51]; (C) Diagram of a juvenile sponge body plan. Water flows into the internal aquiferous system via the ostium and out via the osculum. The mesohyl is shown in blue and populated by archeocytes and other cell types, including sclerocytes and spherulous cells. Adapted from [49]; (D) Optical section of a 3-day-old Amphimedon queenslandica juvenile showing internal morphology and some cell types. Archeocyte (a), choanocyte chamber (cc), endopinacoderm (en), exopinacoderm (ex), ostium (o), osculum (os), sclerocyte (s), spicule (sp), and spherulous cell (sph). Scale bar: 10 μm. Adapted from [49]; (E) Venn diagram denoting the proportion of differentially expressed lncRNAs detected in each of the three cell-type specific transcriptome datasets; (F) Subset of Amphimedon lncRNAs (AmqTCONS_00001337-9) are in close genomic proximity to a cluster of immune-related genes. Coding genes (purple) and long non-coding RNAs (blue) are shown, along with signal coverage tracks showing CEL-seq expression. A grey scale indicates CEL-seq (Cell Expression by Linear amplification and sequencing) expression level: white (no-expression); black (highest expression). The shaded purple area represents the cluster of immune-related genes [Tnf receptor-associated factors (TRAFs)]. Figure was generated using a local instance of the UCSC genome browser [54].
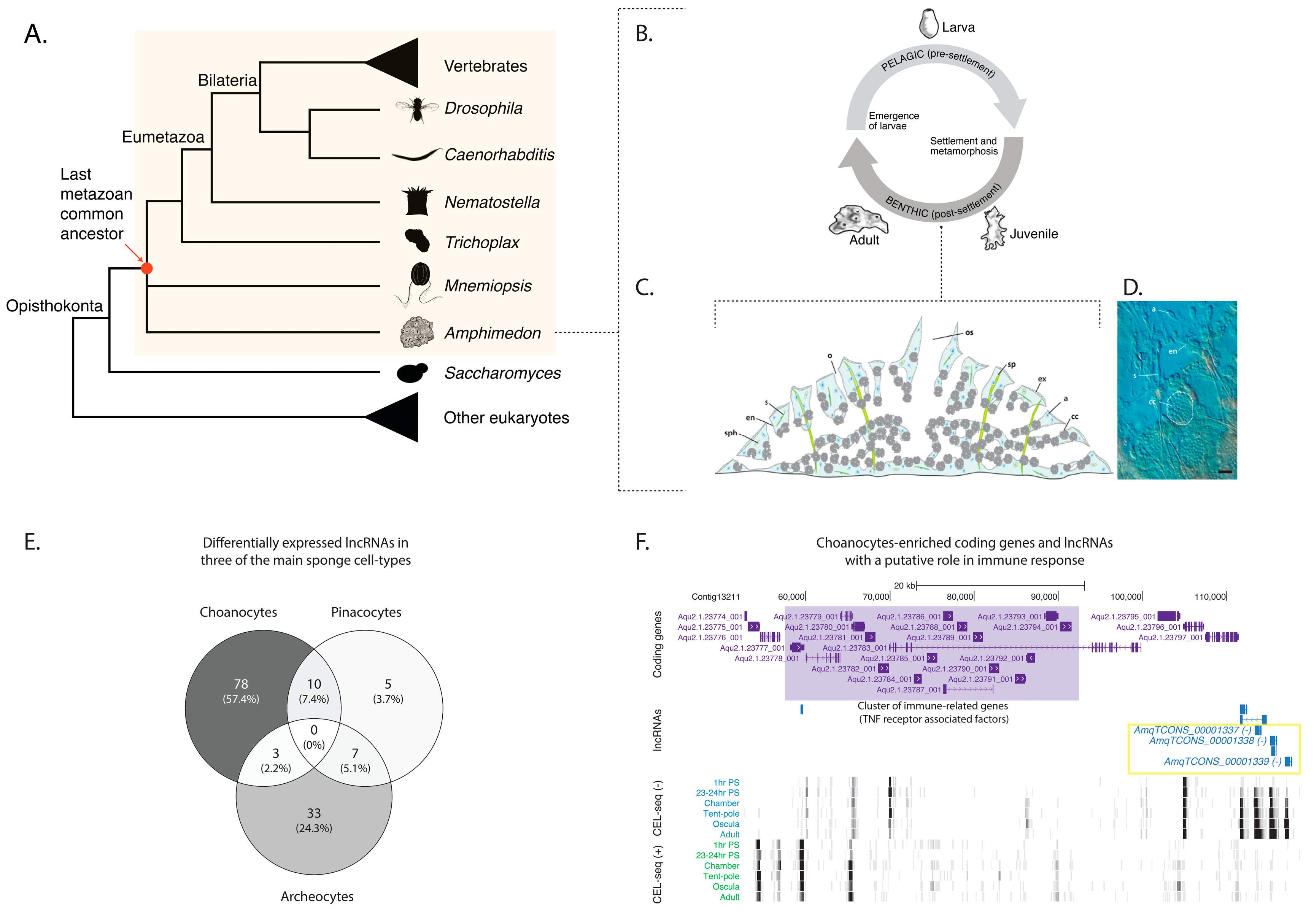
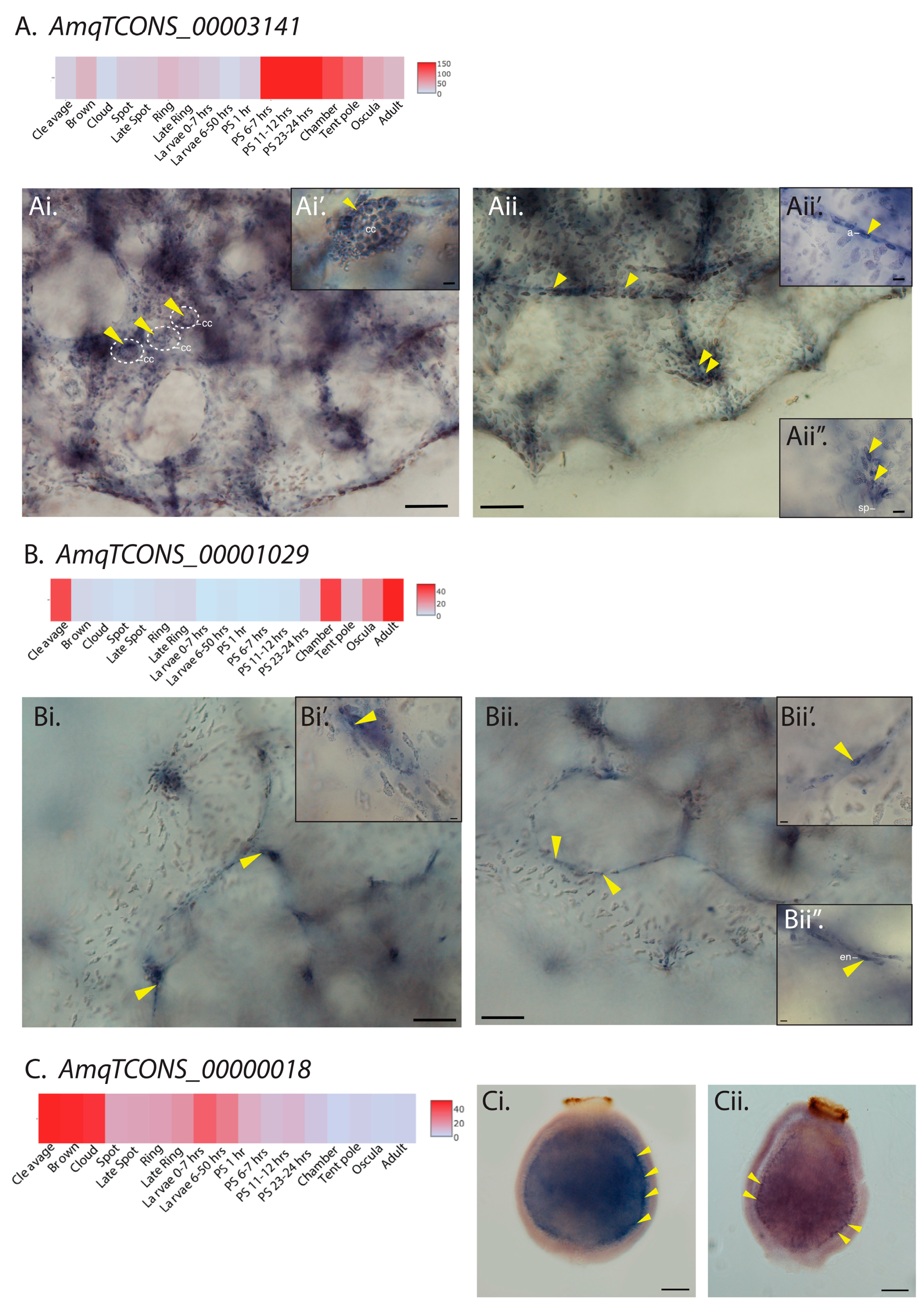
Figure 2.
Cell type-specific restricted expression patterns of three candidate lncRNAs. (A) Heatmap representation of developmental expression of cis-antisense lncRNA AmqTCONS_00003141. (Ai,Aii) Whole mount in situ hybridization (ISH) of AmqTCONS_00003141 to oscula stage juveniles; micrographs are views of the apical (top) side. Yellow arrowheads in (Ai,Ai’) show AmqTCONS_00003141-expressing choanocytes (cc) in chambers. Yellow arrowheads in (Aii) show clusters of AmqTCONS_00003141-expressing cells at the apex of tent-pole like structures, visible here as a vertically oriented cluster of spicules (sp) and associated cells (Aii’’), and aligned in streams running under the body surface. In both cases, these appear to be archeocytes (a) (Aii’). (B) Heatmap representation of developmental expression of lincRNA AmqTCONS_00001029. (Bi,Bii) Whole mount ISH of AmqTCONS_00001029 to oscula stage juveniles; micrographs are views of the apical (top) side. Yellow arrowheads in (Bi,Bi’) indicate tent-pole like structures where there is strong expression of AmqTCONS_00001029. Yellow arrowheads in (Bii) indicate epithelial AmqTCONS_00001029-expressing endopinacocytes (en) (Bii’,Bii’’) that line the internal network of canals. (C) Heatmap representation of expression of the lincRNA AmqTCONS_00000018. (Ci,Cii) Whole mount ISH of larvae labeled with antisense riboprobe and viewed from the lateral side; anterior down. Yellow arrowheads show AmqTCONS_00000018 expression in subepithelial cells at the boundary between outer cell layer and inner cell mass. Scale bars: 50 μm (Ai,Aii,Bi,Bii,Ci,Cii), 5 μm (insets in Ai,Aii,Bi,Bii).
Figure 2.
Cell type-specific restricted expression patterns of three candidate lncRNAs. (A) Heatmap representation of developmental expression of cis-antisense lncRNA AmqTCONS_00003141. (Ai,Aii) Whole mount in situ hybridization (ISH) of AmqTCONS_00003141 to oscula stage juveniles; micrographs are views of the apical (top) side. Yellow arrowheads in (Ai,Ai’) show AmqTCONS_00003141-expressing choanocytes (cc) in chambers. Yellow arrowheads in (Aii) show clusters of AmqTCONS_00003141-expressing cells at the apex of tent-pole like structures, visible here as a vertically oriented cluster of spicules (sp) and associated cells (Aii’’), and aligned in streams running under the body surface. In both cases, these appear to be archeocytes (a) (Aii’). (B) Heatmap representation of developmental expression of lincRNA AmqTCONS_00001029. (Bi,Bii) Whole mount ISH of AmqTCONS_00001029 to oscula stage juveniles; micrographs are views of the apical (top) side. Yellow arrowheads in (Bi,Bi’) indicate tent-pole like structures where there is strong expression of AmqTCONS_00001029. Yellow arrowheads in (Bii) indicate epithelial AmqTCONS_00001029-expressing endopinacocytes (en) (Bii’,Bii’’) that line the internal network of canals. (C) Heatmap representation of expression of the lincRNA AmqTCONS_00000018. (Ci,Cii) Whole mount ISH of larvae labeled with antisense riboprobe and viewed from the lateral side; anterior down. Yellow arrowheads show AmqTCONS_00000018 expression in subepithelial cells at the boundary between outer cell layer and inner cell mass. Scale bars: 50 μm (Ai,Aii,Bi,Bii,Ci,Cii), 5 μm (insets in Ai,Aii,Bi,Bii).
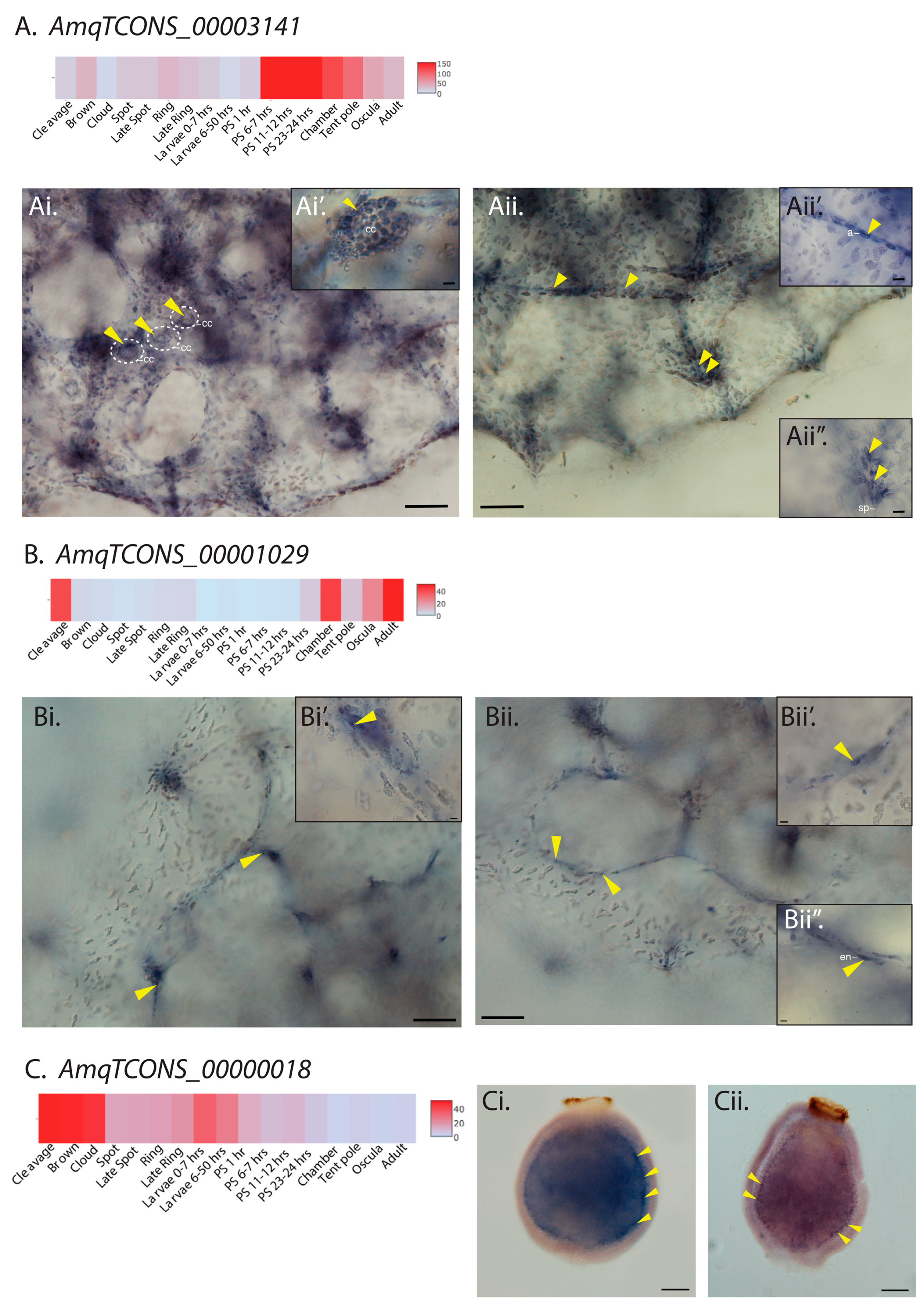
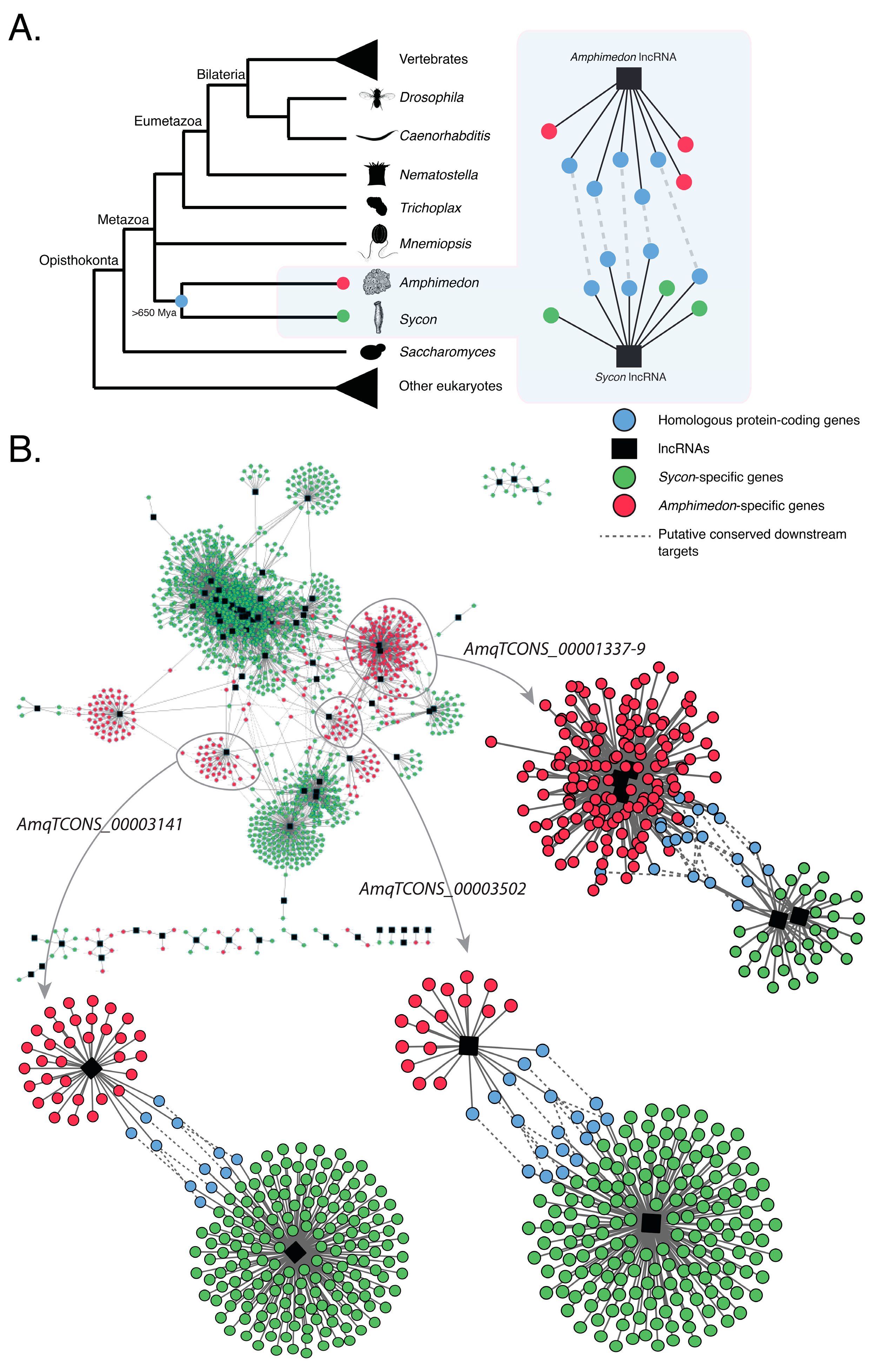
Figure 3.
Putative evolutionarily conserved modules of co-expressed protein-coding genes and lncRNAs in the sponges Amphimedon queenslandica and Sycon ciliatum. (A) A conceptual model of how evolutionarily conserved networks of co-expressed homologous genes and lncRNAs can be inferred across divergent sponge lineages, despite the lack of lncRNA sequence conservation; (B) Co-expression networks based on differentially expressed protein-coding and lncRNA genes in Amphimedon [20] and Sycon [18]. Nodes indicate differentially expressed coding-genes, hubs (black) represent differentially expressed lncRNAs, and edges represent significant co-expression (both positive and negative). Amphimedon-specific genes are shown in red. Sycon-specific genes are shown in green. Conserved homologous genes shared between Amphimedon and Sycon are shown in blue. See Tables S7 and S8 for the complete edge and node lists of genes, and [20] for the developmental expression profiles of AmqTCONS_1337-9, AmqTCONS_3502, and AmqTCONS_0003141 and their co-expressed protein-coding genes.
Figure 3.
Putative evolutionarily conserved modules of co-expressed protein-coding genes and lncRNAs in the sponges Amphimedon queenslandica and Sycon ciliatum. (A) A conceptual model of how evolutionarily conserved networks of co-expressed homologous genes and lncRNAs can be inferred across divergent sponge lineages, despite the lack of lncRNA sequence conservation; (B) Co-expression networks based on differentially expressed protein-coding and lncRNA genes in Amphimedon [20] and Sycon [18]. Nodes indicate differentially expressed coding-genes, hubs (black) represent differentially expressed lncRNAs, and edges represent significant co-expression (both positive and negative). Amphimedon-specific genes are shown in red. Sycon-specific genes are shown in green. Conserved homologous genes shared between Amphimedon and Sycon are shown in blue. See Tables S7 and S8 for the complete edge and node lists of genes, and [20] for the developmental expression profiles of AmqTCONS_1337-9, AmqTCONS_3502, and AmqTCONS_0003141 and their co-expressed protein-coding genes.

{kind=link}
{kind=link}
{kind=link}
Table 1.
List of conserved homologous genes shared between Amphimedon queenslandica (Aqu2.1 prefix) and Sycon ciliatum (scigt prefix) for the three examples of putative evolutionarily conserved modules of co-expressed coding genes and lncRNAs. See Tables S7 and S8 for the complete edge and node lists of genes.
Table 1.
List of conserved homologous genes shared between Amphimedon queenslandica (Aqu2.1 prefix) and Sycon ciliatum (scigt prefix) for the three examples of putative evolutionarily conserved modules of co-expressed coding genes and lncRNAs. See Tables S7 and S8 for the complete edge and node lists of genes.
| AmqTCONS_00003141 | |
| Homologous gene pairs | Description |
| scigt010895-Aqu2.1.43387_001 | mitochondrial dicarboxylate carrier |
| scigt017797-Aqu2.1.41074_001 | protein disulfide-isomerase a5-like |
| scigt001771-Aqu2.1.30885_001 | sh3 and px domain-containing protein 2a-like |
| scigt016036-Aqu2.1.36626_001 | adp-ribosylation factor gtpase-activating protein 2-like |
| scigt018255-Aqu2.1.30885_001 | sh3 and px domain-containing protein 2a-like |
| scigt000612-Aqu2.1.41568_001 | tgf-beta receptor type-1 |
| scigt008994-Aqu2.1.41568_001 | tgf-beta receptor type-1 |
| AmqTCONS_00001337-9 | |
| Homologous gene pairs | Description |
| scigt017951-Aqu2.1.43947_001 | arylsulfatase b-like |
| scigt017951-Aqu2.1.24502_001 | arylsulfatase b-like |
| scigt017951-Aqu2.1.39727_001 | arylsulfatase |
| scigt017951-Aqu2.1.41029_001 | arylsulfatase |
| scigt017951-Aqu2.1.37909_001 | sulfatase |
| scigt014545-Aqu2.1.37909_001 | sulfatase |
| scigt014545-Aqu2.1.41029_001 | arylsulfatase |
| scigt014545-Aqu2.1.39727_001 | arylsulfatase |
| scigt017997-Aqu2.1.32274_001 | usherin |
| scigt020120-Aqu2.1.28087_001 | lysosomal alpha-glucosidase-like isoform x2 |
| scigt020423-Aqu2.1.35119_001 | filamin-c-like isoform x3 |
| scigt000557-Aqu2.1.32241_001 | myosin-i heavy chain |
| scigt008273-Aqu2.1.36394_001 | deleted in malignant brain tumors 1 |
| scigt017951-Aqu2.1.42755_001 | arylsulfatase b-like |
| AmqTCONS_00003502 | |
| Homologous gene pairs | Description |
| scigt000138-Aqu2.1.44676_001 | actin family protein |
| scigt001771-Aqu2.1.38758_001 | tyrosine-protein kinase lck |
| scigt005362-Aqu2.1.44676_001 | actin family protein |
| scigt004922-Aqu2.1.40987_001 | unconventional myosin-viia |
| scigt008792-Aqu2.1.24982_001 | adenylyl cyclase-associated protein 1 |
| scigt012572-Aqu2.1.40987_001 | unconventional myosin-viia |
| scigt014349-Aqu2.1.32914_001 | pleckstrin homology domain-containing family g member 1-like |
| scigt016045-Aqu2.1.28519_001 | ap-2 complex subunit alpha-1-like |
| scigt020995-Aqu2.1.43989_001 | protein plant cadmium resistance 3-like |
| scigt021992-Aqu2.1.44676_001 | actin family protein |
| scigt022018-Aqu2.1.44676_001 | actin family protein |
| scigt025009-Aqu2.1.40987_001 | unconventional myosin-viia |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gaiti, F.; Hatleberg, W.L.; Tanurdžić, M.; Degnan, B.M. Sponge Long Non-Coding RNAs Are Expressed in Specific Cell Types and Conserved Networks. Non-Coding RNA 2018, 4, 6. https://doi.org/10.3390/ncrna4010006
AMA Style
Gaiti F, Hatleberg WL, Tanurdžić M, Degnan BM. Sponge Long Non-Coding RNAs Are Expressed in Specific Cell Types and Conserved Networks. Non-Coding RNA. 2018; 4(1):6. https://doi.org/10.3390/ncrna4010006
Chicago/Turabian StyleGaiti, Federico, William L. Hatleberg, Miloš Tanurdžić, and Bernard M. Degnan. 2018. "Sponge Long Non-Coding RNAs Are Expressed in Specific Cell Types and Conserved Networks" Non-Coding RNA 4, no. 1: 6. https://doi.org/10.3390/ncrna4010006
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.