Why Are Weissella spp. Not Used as Commercial Starter Cultures for Food Fermentation?


UMR C-95 QualiSud, Université de La Réunion, CIRAD, Université Montpellier, Montpellier SupAgro, Université d’Avignon et des Pays de Vaucluse, F-97490 Sainte Clotilde, France
*
Author to whom correspondence should be addressed.
Fermentation 2017, 3(3), 38; https://doi.org/10.3390/fermentation3030038
Submission received: 25 June 2017
/
Revised: 10 July 2017
/
Accepted: 14 July 2017
/
Published: 3 August 2017
(This article belongs to the Special Issue Fermentation and Bioactive Metabolites)
Abstract
:Among other fermentation processes, lactic acid fermentation is a valuable process which enhances the safety, nutritional and sensory properties of food. The use of starters is recommended compared to spontaneous fermentation, from a safety point of view but also to ensure a better control of product functional and sensory properties. Starters are used for dairy products, sourdough, wine, meat, sauerkraut and homemade foods and beverages from dairy or vegetal origin. Among lactic acid bacteria, Lactobacillus, Lactococcus, Leuconostoc, Streptococcus and Pediococcus are the majors genera used as starters whereas Weissella is not. Weissella spp. are frequently isolated from spontaneous fermented foods and participate to the characteristics of the fermented product. They possess a large set of functional and technological properties, which can enhance safety, nutritional and sensory characteristics of food. Particularly, Weissella cibaria and Weissella confusa have been described as high producers of exo-polysaccharides, which exhibit texturizing properties. Numerous bacteriocins have been purified from Weissella hellenica strains and may be used as bio-preservative. Some Weissella strains are able to decarboxylate polymeric phenolic compounds resulting in a better bioavailability. Other Weissella strains showed resistance to low pH and bile salts and were isolated from healthy human feces, suggesting their potential as probiotics. Despite all these features, the use of Weissella spp. as commercial starters remained non-investigated. Potential biogenic amine production, antibiotic resistance pattern or infection hazard partly explains this neglecting. Besides, Weissella spp. are not recognized as GRAS (Generally Recognized As Safe). However, Weissella spp. are potential powerful starters for food fermentation as well as Lactococcus, Leuconostoc or Lactobacillus species.
1. Introduction
Fermentation has been applied for thousand years as a method to preserve perishable foods. It seems that fermentation appeared in the period 8000–6000 BC in Iraq with the fabrication of cheese. Later, in the period 4000–2000 BC, Egyptians discovered how to use yeasts to make leavened bread and wine [1]. Nowadays, a large variety of fermented foods are consumed around the world and appreciated for their organoleptic and sensory properties, including cheese, yoghurt, kefir, beer, Kombucha, Pozol, sourdough, Kimchi, olives, sauerkraut, pickles, Tempe, Gari, Fufu or sausages [2]. Whatever the nature of the raw material, fermented foods are obtained either by spontaneous fermentation or by the use of starter cultures. Spontaneous fermentation results from the combined actions of microorganisms originating from the raw materials or the environment, including yeast, fungi and bacteria [3]. Started fermentation consists in inoculating the raw material with a single or a mix of safe microbial strains in order to control and accelerate the fermentation process. These two different processes result in different molecular changes and different sensory properties. Microorganisms used in commercial starter cultures include bacteria, yeasts and molds. Molds are used in the production of meat and cheese products and the species available as commercial starter cultures are Penicillium roqueforti, Penicillium nalgiovense and Penicillium chrysogenum [4]. Yeasts are used in the production of beer, wine, bread and cheese. The species available as commercial starter cultures include Debaryomyces hansenii, Galactomyces geotrichum but mostly Saccharomyces cerevisiae. Among the microorganisms available as starter cultures, lactic acid bacteria (LAB) are the most diverse. They are used to start wine, bread, vegetables, meat and dairy product fermentation.
LAB are a group of gram-positive bacteria, non-spore forming, cocci or rods, that produce lactic acid as the major end-product of carbohydrate fermentation. LAB are extensively involved in spontaneous fermentation and the major genera are Carnobacterium, Enterococcus, Lactobacillus, Lactococcus, Leuconostoc, Oenococcus, Pediococcus, Streptococcus, Tetragenococcus, Vagococcus and Weissella [5]. They play a central role in the fermentation process. Particularly, they cause a rapid acidification of the raw material by the production of lactic acid and produce several other important compounds such as ethanol, carbon dioxide, bacteriocins, exopolysaccharides (EPS), aromatic compounds, vitamins or useful enzymes. Lactic fermented foods have a long and safe history of consumption and have been associated with health benefits, safety, nutritional and sensory properties improvement. For these reasons, LAB have gained interest these last decades and the research for new LAB starters is increasingly developed.
Nowadays, more than 20 LAB species are available as commercial starter cultures. Among those, Lactobacillus plantarum, Lb. delbrueckii, Lb. sakei, Lactococcus lactis, Streptococcus thermophilus, Pediococcus pentosaceus and Leuconostoc mesenteroides are the species mostly used [6,7,8]. They have been used for decades in food fermentation, are recognized as safe and were selected from technological and sensorial assays.
Over the past 20 years, scientists have gained interest for the genus Weissella. Weissella was proposed as a genus in 1993 from re-classified Lactobacillus and Leuconostoc spp. They are frequently detected in many fermented foods [9,10,11,12,13] and participate to the final stage of the fermented product, together with Lactobacillus and Leuconostoc. As other LAB species used as commercial starters, Weissella spp. produce bacteriocins, EPS and hydrolytic enzymes [14]. However, they are still not used as commercial starters in food industries and remain under a laboratory investigation stage.
This review aims to understand the reasons why Weissella spp. are not used as commercial starters in food fermentation. Taxonomy, characteristics and the role of Weissella spp. in spontaneous fermentation were investigated, as well as their functional and technological properties, and possible reasons that would explain the poor investigation of Weissella spp. as starters.
2. Weissella Genus
2.1. Phenotypic and Genetic Characteristics of Weissella spp.
Weissella genus belongs to the Leuconostocaceae family together with the genera Leuconostoc, Oenococcus and Fructobacillus [15]. Weissella genus was proposed in 1993 from previously Leuconostoc and Lactobacillus classified species. Weissella spp. are gram-positive, catalase negative, asporogenous, non-motile, except for W. beninensis, and lack cytochrome [16]. Cells are either short rods or ovoid and occur in pair or in short chains. They are facultative anaerobic with an obligatory fermentative metabolism. Weissella spp. are hetero-fermentative: glucose is fermented via the hexose-monophosphate and phosphoketolase pathways. End products are lactic acid, CO2, ethanol and acetic acid, except for Weissella ceti which does not produce gas [17]. Configuration of lactic acid can be either D(-) or DL. Hydrolysis of arginine depends on the species whereas all Leuconostoc spp. are arginine-negative. Cell-wall peptidoglycan is composed of lysine and the interpeptide bridge contains alanine or serine, except for Weissella kandleri which contains glycine. Growth mainly occurs between 15 °C and 37 °C, but some strains of Weissella cibaria and Weissella confusa are able to grow at 45 °C. The type species is Weissella viridescens. Fusco et al. [18] have drawn a complete description of Weissella spp., except for Weissella bombi and Weissella jogaejeotgali which have been described afterwards.
Distinction between Weissella, Leuconostoc and heterofermentative lactobacilli remains impossible based only on morphological observation. Since they produce gas from carbohydrates, members of the genus Weissella may be distinguished from homofermentative lactobacilli, enterococci, pediococci, lactococci and streptococci. Generally, identification of Weissella spp. is performed with molecular methods, such as 16S rRNA gene sequencing, amplified ribosomal DNA restriction analysis (ARDRA), ribotyping, Matrix-associated laser desorption ionization Time-of-Flight (Maldi-ToF), repetitive element-PCR fingerprinting using (GTG)5-PCR or fluorescent-Amplified Fragment Length Polymorphism (fAFLP) [18].
The genome sequence of 28 strains belonging to 12 species has been published (GenBank). The smallest genomes belong to Weissella halotolerans (1.36 Mb) and W. ceti (1.35 to 1.39 Mb), which share the same taxonomic branch. The largest genomes belong to W. cibaria (2.32 to 2.47 Mb), W. confusa (2.18 to 2.28 Mb), W. jogaejeotgali (2.11 Mb) and Weissella oryzae (2.13 Mb), all these four species having been isolated from fermented foods. The core-proteome of Weissella represented 729 COGs (Clusters of Orthologous Genes), over a pan-proteome of 4712 COGs. Core-proteome is essentially involved in housekeeping but shows a relative lack of shared traits between all species, which might explain the diversity of niches of the species.
2.2. Taxonomical Position and Specific Traits
Collins et al. [19] demonstrated that Lactobacillus confusus, Lactobacillus kandleri, Lactobacillus minor, Lactobacillus viridescens and Lactobacillus halotolerans were phylogenetically related to Leuconoctoc paramesenteroides. Besides, a unique murein type, Lys-Ser/Ala, was reported within Lb. viridescens, Lb. minor and Lb. halotolerans, which was identical to that found in the Lc. paramesenteroides group [20]. Hence, from this group, the genus Weissella was created and species were named Weissella paramesenteroides, W. confusa, W. kandleri, Weissella minor, W. viridescens, W. halotolerans plus Weissella hellenica, isolated from fermented sausages [19]. Based on 16S and 23S rRNA sequence analyses, three evolutionary lines were distinguished in Leuconostoc spp., nowadays named Leuconostoc sensu strico, W. paramesenteroides and Oenococcus oeni [21]. The genera Oenococcus and Fructobacillus have been proposed from re-classified Leuconostoc spp. [22,23], but interestingly, Weissella was the only genus of the Leuconostocacea family gathering re-classified Leuconostoc and Lactobacillus spp. Currently, the genus Weissella comprises 21 species, isolated from a high variety of sources (Table 1). Based on partial 16S rRNA coding sequence, several branches can be separated within Weissella spp. (Figure 1 and Table 1). Weissella ghanensis, Weissella fabaria, Weissella fabalis, all isolated from fermented cassava, and W. beninensis belong to the same branch. W. ceti, W. halotolerans, W. minor, Weissella uvarum and W. viridescens share another taxonomic branch, which mainly originates from previous Lactobacillus spp. [20]. Species of this branch harbor diverse habitats. W. viridescens and W. halotolerans have been isolated from meat products [20,24], W. minor from milking machine sludge [20], W. ceti from a beaked whale [17] and W. uvarum from wine grapes [25].
Weissella diestrammenae, W. kandleri and Weissella koreensis constitute a third branch with strains from different sources. W. kandleri and W. koreensis were isolated from Koumiss, a fermented drink from mare milk, and from kimchi, respectively. W. cibaria and W. confusa belong to the same branch and share numerous habitats. Both species have been found in fermented foods, animal, clinical samples and human feces. These two species are hardly distinguishable since they share 99.6% of identity in their 16S rRNA coding sequence. Another branch comprises W. bombi, W. hellenica, W. jogaejeotgali, W. paramesenteroides and Weissella thailandensis. All the species from this branch have been isolated from fermented foods except W. bombi isolated from insect gut. Interestingly, W. jogaejeotgali and W. thailandensis are closely related and both have been detected in Jeotgal, a fermented seafood product. W. paramesenteroides and W. hellenica were detected in fermented sausages, fermented milk or cheese.
Finally, two species remained unclassified, W. soli and its closest neighbor species, W. oryzae. W. soli is the only Weissella spp. which has been isolated from soil and W. oryzae has been isolated from fermented rice grains.
All Weissella species except five were isolated from fermented food (Table 1).
3. Occurrence of Weissella in Spontaneous Fermentation
3.1. General Occurrence
Numerous Weissella strains have been isolated from a large panel of spontaneous fermented foods including fruits and vegetables-based foods, fermented meat and fish products and starchy or cereal-based foods (Table 2). A frequent detection of Weissella spp. in a large variety of spontaneous fermented foods strengthens the fact that they are able to adapt to a multitude of environment and play an important role in the fermentation process. Moreover, the identification of Weissella spp., which dominates the microflora of fermented foods, is an important step for the development of new starter cultures for the fermentation industry. These are the reasons why we hypothesized that the high presence and the role of Weissella spp. in fermented food justifies to pay attention to a possible use as starter in the food fermentation industry.
3.2. Fruits and Vegetables-Based Products
W. cibaria, W. confusa and W. paramesenteroides have been isolated from fermented leek, fermented cabbage, fermented cauliflower and sauerkraut [63,65,67,119]. These species but also W. soli, W. thailandensis, W. hellenica, W. koreensis, W. minor, W. halotolerans and W. jogaejeotgali have been isolated from Asian traditional fruits and vegetables-based fermented products, like Kimchi, Yan-dong-gua, Pobuzihi, Jiang-gua, Yan-tsai-shin, Yan-jiang, Yan-taozih, Xi-gua-mian, Dochi, Suan-tsai, Jeotgal and Stinky tofu (Table 2) [29,36,41,42,43,44,46,47,48,49,50,51,52,53,54,56,57]. Kimchi is a Korean traditional fermented vegetable composed of Chinese cabbage and other ingredients like radish, green onion, red pepper power, garlic, ginger and fermented seafood (Jeotgal) [120]. Yan-taozih (pickled peaches), Xi-gua-mian (fermented watermelon) and Pobuzihi (fermented cummingcordia) are popular fermented fruits in China and Taiwan [53,120]. Yan-dong-gua, Jiang-gua, Yan-tsai-shin and Yan-jiang are traditional fermented foods of Taiwan, composed of fermented wax gourd, cucumber, broccoli stems and ginger, respectively.
Interestingly, Weissella spp. are frequently found in vegetables which belong to the cabbage family (cabbage, broccoli, cauliflower and radish) and in fruits from the gourd family (cucumbers, wax gourd, watermelon). Nowadays, cabbages, cucumbers and olives are the main vegetables industrially transformed by lactic acid fermentation [7]. Persistence of Weissella spp. in cabbage and gourd-based fermented products is a cornerstone for its use as starters for these kinds of products.
Actually, the role of Weissella spp. in vegetable fermentation is not clearly understood. They are frequently isolated from fermented vegetables and participate to the final stage of the fermented product. W. cibaria and W. paramesenteroides are responsible for the fermentation of Yan-dong-gua. Surprisingly, W. cibaria was replaced by W. paramesenteroides after 5 days of fermentation. In addition, after 28 days of fermentation, both Weissella species disappeared [47]. These species disappearances remain not understood and need to be investigated. Interestingly, these two species occur together in other fermented foods and similar changes were observed. Both species were detected in the 3-days initial fermentation of Fu-tsai and spontaneous cauliflower and mixed-vegetables fermentation but disappeared during the following steps of the fermentation processes [56,63]. These two species seem to work together in vegetables fermentation and constitute an interesting option for their use as a mixed starter culture. W. cibaria, W. confusa and W. koreensis are frequently involved in Kimchi fermentation. W. confusa (together with Lc. citreum) was present throughout the fermentation process of kimchi, suggesting its importance during the fermentation [46] W. cibaria and W. confusa have been detected in the early stage of kimchi fermentation, but were replaced by Lb. plantarum, Lb. brevis and Lc. pseudomesenteroides. It seemed that W. cibaria and W. confusa started the fermentation and created an environment suitable for the growth of Lactobacillus spp. [11]. It has been shown that W. soli was present during the early stage of fermentation of kimchi but was replaced by W. koreensis [42]. Because kimchi is generally processed by spontaneous fermentation, the LAB microflora of kimchi may vary. However, the LAB species most frequently found in kimchi are Lc. citreum, Lc. mesenteroides, Lc. lactis, W. cibaria, W. confusa, W. koreensis, Lb. plantarum and Lb. brevis. Generally, in vegetable fermentation, Weissella and Leuconostoc spp. start the fermentation and are followed by more acidic tolerant species such as Lactobacillus spp.
3.3. Dairy Fermented Foods
W. confusa, W. cibaria, W. thailandensis, W. paramesenteroides, W. hellenica and W. viridescens have been detected in different types of cheese and fermented milk (Table 2) [68,69,70,71,72,73,74,75,76,77,78,79,80,81].
W. paramesenteroides represented a high proportion of LAB isolated both from the surface and interior of fresh Manura cheese [76]. Besides, it was also one of the predominant LAB species isolated during cheese ripening, suggesting its important role in the production of Manura cheese. W. confusa appeared occasionally in Nunu fermented milk [69]. W. confusa NGB-82 (Lb. confusus) together with L. lactis subsp. lactis MOS-11 dominated in Masai fermented zebu milk. These two strains were used as starter cultures for this kind of milk and were able to rapidly acidify the media and inhibit the growth of S. aureus and E. coli [71]. However, W. confusa was not present in another Masian fermented milk [70]. W. viridescens was occasionally found in Scamorza Altamurana cheese [77] and in Romanian dairy products [80]. W. hellenica 1M30 was able to decrease the pH of skimmed milk for the production of Mozarella cheese and showed a proteolytic activity on milk proteins [78]. This strain may be a suitable starter culture for industrial processing of Mozarella cheese. This was not the case for fermented camel milk since W. hellenica was found in only one sample suggesting a contamination [73].
3.4. Meat and Fish-Based Products
The main Weissella species involved in meat and fish fermentation are W. cibaria, W. confusa, W. paramesenteroides, W. hellenica, W. viridescens and in a lesser part, W. thailandensis (Table 2). W. thailandensis has been first isolated from Pla-ra, a fermented fish product from Thailand [30] and more recently, in Mum, a Thai fermented meat product [86]. Miyashita et al. [86] isolated also W. cibaria, W. confusa, W. paramesenteroides and W. viridescens in several others Thai fermented fish products like, Pla-ra sub, Plaa-som, Pla-jom and Pla-jaw. Presence of W. cibaria and W. confusa in Plaa-som was confirmed by [115,116]. W. cibaria dominated the early stage of fermentation and contributed to the product finishing.
The main LAB involved in dry and fermented sausages are Lb. plantarum, Lb. sakei and Lb. curvatus. However, W. cibaria, W. paramesenteroides, W. hellenica and W. viridescens are frequently detected in fermented sausages [102,105,106,107,108,109,113]. W. hellenica represented a low portion of the LAB isolated from Italian sausages, suggesting that its contribution to the fermentation process was not really important. This observation was confirmed by [107] as W. viridescens has been occasionally detected in Greek-dry fermented sausages. W. minor has been detected in Greek-fermented sausages but should not be used as starter for meat fermentation due to its high production of gas and peroxide [113]. As a whole, generally, Weissella spp. represent a minority part of LAB involved in fermented sausages and their use as starter cultures for fermented meat products is almost not considered.
3.5. Starchy and Cereal-Based Products
As shown in Table 2, W. confusa has been detected in several cereal-based fermented foods like Togwa, Kunu-zaki, Boza, Fura and Borde [39,82,97,98,99]. W. cibaria, W. confusa and W. paramesenteroides have been found in several fermented cassava products, such as Gari, Attieke and Lafun [16,83,84,85,86]. These species have also been isolated from different types of sourdough [10,88,89,90,92].
In Togwa fermentation, W. confusa was present from the beginning to the end of the fermentation, together with Lb. plantarum, Lb. brevis, Lb. fermentum, Lb. cellobiosus and P. pentosaceus. However, Lb. plantarum was the dominant species [82]. W. confusa has been shown to be the dominant species together with Lb. fermentum during the fermentation of Fura. Indeed, these two species were isolated from eight different processing sites and at almost all fermentation stages [98]. W. confusa and W. viridescens were the dominant species during the fermentation of Borde, together with Lb. brevis and P. pentosaceus. These two species are thus adapted to the niche of Borde fermentation and constitute potential starter cultures for the industrial production of Borde [99]. W. confusa together with Lb. plantarum and Lb. fermentum were also the dominant species during the fermentation of Lafun [86].
4. Spontaneous Versus Started Fermentation
4.1. Role of Starters in Food Fermentation
Traditional fermented foods are mainly obtained by a naturally occurred fermentation, resulting from competitive activities of different microorganisms derived from the raw material or the environment, including yeast, fungi and lactic acid bacteria [3,121]. Some traditional fermented foods have been consumed for centuries and remained today an important part of the human diet. Some famous ancient fermented foods are kefir, sauerkraut, Kimchi, kombucha and wine. Kefir, a fermented milk beverage, is a drink originated from the Caucasian mountains and represents today a market of €78.7 million only in North America [122]. Over spontaneous fermentation, microorganisms that are best adapted to the environment and exhibit the highest growth rate, dominate the fermentation process [123]. Generally, a succession of flora is observed [7,124]. Due to multiple microorganisms’ activities, spontaneous fermentation remains difficult to control. Thus, the quality of a spontaneous fermented product is not predictable, undesirable change of the sensory profile can occur and the risk of fermentation failure can be important [7]. Using starter culture is the most efficient way to control the fermentation process, to ensure quality and safety of the fermented product. A starter culture can be defined as a microbial preparation containing a high number of at least one microorganism to produce a fermented food by accelerating its fermentation process and bring desirable sensory properties [8,121]. In 1873, Lister isolated the first pure bacterial culture, Bacterium lactis (nowadays Lactococcus lactis) which became, in 1890, the first starter culture to be used for the cheese and sour-milk production [121]. Nowadays, a number of LAB starters is used in the food industry (Table 3), producing a high diversity of fermented products. The majority of LAB starters belong to the genera Lactobacillus, Lactococcus, Steptococcus, Pediococcus, Leuconostoc and Bifidobacterium. Lactobacillus species are used for the production of dairy, vegetable, cereal and meat products. Lactococcus species are mainly used for dairy products and the main species used as commercial starter is L. lactis. Streptococcus thermophilus is the only species within the genus Streptococcus to be used as commercial starter. It is used for the production of yoghurt and some cheeses, due to its particular rapid acidification in milk fermentation [125]. Among the Leuconostoc spp., Lc. mesenteroides is the main species used as commercial starter. Lc. mesenteroides is used for the production of buttermilk and some cheese [126]. Pediococcus species are mainly involved in the fermentation of meat products and of sourdough. P. pentosaceus and P. acidilactici, together with Lb. sakei, Lb. curvatus, Lb. plantarum, Lb. pentosus and Lb. casei are the species mostly used as commercial starter for meat products [124]. Oenococcus oeni, Lb. plantarum and Lb. hilgardii are available as commercial starter cultures for the malolactic fermentation of wine since the beginning of the 1980s [3].
Before commercialization, starters have to pass many steps of selection and performance tests.
4.2. Expected Characteristics of Starters
General criteria for the selection of starters deal with safety, technological and economical aspects [3]. The starter culture should be able to develop some expected flavor and aroma, to improve safety, nutritional and shelf-life properties of foods. Thus, criteria for selection of starters are mainly divided in four aspects: technological, sensory, nutritional and safety (Table 4). A quick growth and a high acidification rate are the first criteria for the selection of starter. A rapid acidification will create a safe environment at the initial stage of the fermentation and thus reduce food pathogen hazard and spoilage probability. During fermentation, LAB can be exposed to salt stress due to important amounts of added salt, but also in fruits to high sugar levels. Therefore, adaptation to hyperosmotic environments has to be considered, especially for making fruits, vegetables or meat based products.
The synthesis of antimicrobial compounds or bacteriocins is desired during fermentation, as these compounds inhibit the proliferation of pathogens or spoilage microorganisms. Bacteriocins are natural antimicrobial proteins produced by bacteria, including LAB, and their use in food industries in an interesting alternative for food preservation. Nisin, a bacteriocin produced by several L. lactis strains is commercially available in UE as a bio-preservative [127]. Lactic acid, acetic acid, ethanol, hydrogen peroxide (H2O2), diacetyl, reuterin and bacteriocins are produced by LAB and show inhibiting activities against food borne pathogens, including both gram-negative and gram-positive bacteria [128]. In addition, LAB are able to produce antifungal compounds which inhibit mycotoxin producer growth and can bind and reduce the content of mycotoxins [129].
The synthesis of exopolysaccharides (EPS) could be searched, especially for dairy and bakery products, for both sensory and nutritional reasons. According to their composition, EPSs are divided into homopolysaccharides (HoPS) and heteropolysaccharides (HePS). HePSs are composed of repeated units consisting of two or more monosaccharides, mainly galactose, glucose, fructose and rhamnose. HoPS are composed of repeated units of one monosaccharide, mainly glucose or fructose, and are classified in α-d-glucans (dextran, mutan, reuteran and alteran) or in β-d-glucans (levan and inulin-types). EPSs are produced intracellularly or extracellularly, and some bacteria are able to produce capsular EPS (CPS), which are covalently bound to the cell surface. In food industry, EPS are used as viscosifier, water-binding or gelling agents, stabilizers, texturizers and emulsifiers. Besides, EPS may act as antioxidant, antitumor, antiulcer and exert immune-modulating and cholesterol-lowering activities [132].
The ability to increase antioxidant activity is another criterion used to select functional starters for fruits and vegetables fermentation [7]. Several studies demonstrated that lactic acid fermentation increased the polyphenol content associated with an increase of the antioxidant activity [133,134,135,136]. This increase is mainly explained by enzymatic depolymerization of phenolic compounds by LAB [137], leading to more efficient uptake and higher bioavailability of plant polyphenols.
4.3. Non-Expected Characteristics of Starters
Despite of its antimicrobial activity, H2O2 is non-desired for some fermented products, especially in fermented sausages. Indeed, H2O2 can react and degrade organic and antioxidant compounds, increase rancidity and form a discoloration of the final product [7,124]. Papamanoli et al. [107] reported that 100% of Lb. plantarum strains (7), 58% of Lb. curvatus strains (24) and 84% of Lb. sakei strains (49) were able to produce H2O2. It is surprising to note that these species are the main used as starters for fermented sausages.
Others LAB characteristics, which are non-desired for their use as starters, are those associated with health disorders. In particular, biogenic amines, which are organic bases mainly formed by the decarboxylation of amino acids, caused a number of outbreaks, resulting from fish or cheese consumption [138]. Histamine, putrescine, cadaverine, tyramine and β-phenylethylamine are the biogenic amines most frequently found in fermented foods like cheese, wine, sausages, beer, sauerkraut and fish. Bacteria associated with biogenic amines poisoning belong to the Enterobacteriaceae family, such as Morganella morganii, Klebsiella pneumonia and Hafnia alvei. Despite the fact that LAB could produce biogenic amines, no study has reported food poisoning resulting from LAB activities.
4.4. Improvement of Starters
Together with the starter research, several tools have been developed these last decades in order to enhance technological, sensory and nutritional properties of selected starters. Genetic and metabolic engineering facilitates the manufacturing of new starters expected to be superior to that found naturally [139]. It consists in adding or deleting some desired or non-desired criteria, altering a specific trait of a starter or transferring into another more preferable strain. Several tools have been used to develop new interesting and performant starters, such as the use of plasmid vectors, chromosome modification and expression systems, genetic mutants or engineering strains with genes from other LAB or other bacteria [140]. For instance, the gene encoding a bacteriocin from Lb. helveticus was introduced into Lb. acidophilus using a recombinant plasmid [141]. Genes encoding peptidases from a proteolytic Lb. helveticus strain were introduced into L. lactis. New peptidolytic activities were conferred to L. lactis with the aim to accelerate cheese ripening process [142]. Generally, independently of genetic methods used, starter genome sequencing is necessary and constitutes the model for starter engineering. However, authorization for producing these kind of microorganisms is necessary in most countries and consumers are reluctant regarding the use of genetically modified microorganisms in food industry.
Evolutionary engineering is currently the most promising direction for the improvement of starters. It consists of repeated steps of batch cultivation strains in the presence of a selective pressure or under selective conditions. The aim of serial batch cultivations is to increase genetic diversity resulting from mutations. Some mutants will survive and grow better under specific conditions than the initial strains [143,144]. Evolutionary engineering is largely investigated for Saccharomyces cerevisiae [145] and application on LAB deserves a deeper investigation. Experimental evolution has been performed on L. lactis KF147 strain, isolated from mung-bean sprouts [146]. The particularity of this strain is its poor growth in milk compared to other dairy L. lactis strains. L. lactis KF147 was serial cultivated for 1000 generations in skimmed milk and three mutants were selected NZ5521, NZ5522, and NZ5523. The acidification rates of NZ5521, NZ5522 and NZ5523 were increased compared to KF147.
5. Investigation into Using Weissella spp. as a Starter
5.1. Acidification and Growth Performance
Acidification and growth performance are important criteria for the selection of starters. W. koreensis HO20 (and Lc. citreum H012) reached to 6.5 to 9.5 log·cfu·g−1 of dough and decreased the pH from 6.4 to 4.4 after 24 h of sourdough fermentation [147]. As a comparison, Lb. plantarum and Leuconostoc spp. reached to 6.5 to 9.5 log·cfu·g−1 and 6.8 to 9.2 log·cfu·g−1 of dough, respectively after 20 h of sourdough fermentation [148]. W. cibaria MG1 decreased the pH from 6.8 to 4.3 during the fermentation of different sourdough (buckwheat, quinoa, teff and wheat) [149]. W. confusa PE36, Lb. plantarum PE21 and Lb. curvatus PE4 grew from 4.5 to 8.9 log·cfu·g−1 and decreased the pH from 5.0 to 3.7 after 15 h of fermentation of red and yellow peppers [150]. W. thailandensis was not able to acidify milk compared to Lactobacillus spp. [75]. Growth and acidification capacities of Weissella species in food fermentation, more precisely W. cibaria, W. confusa and W. koreensis are similar to those observed for Lactobacillus or Leuconoctoc.
5.2. Production of Exopolysaccharides
Commercial production of EPS has focused mainly on dextran producing Leuconostoc species. However, several studies reported the production of EPS by Weissella spp. (Table 5). EPS are generally produced at levels of 10 mg·L−1 to 1 g·L−1 and have high molecular mass (106 Da). Dextran from Weissella spp. harbor a molecular mass between 104 and 108 Da. Dextran of Weissella may be an alternative for the industry. Dextran production by W. cibaria MG1 during sourdough fermentation was reported by [149,151]. W. cibaria MG1 produced a significant higher amount of EPS (8 g/Kg) than Lb. reuteri Y2 and VIP (0.6 and 3.28 g/Kg, respectively) in sourdough. W. cibaria WC4 and Lb. plantarum PL9 produced the same amount of EPS during sourdough fermentation (2.5 g/Kg). Besides, these two strains increased the viscosity and the resulting bread showed a higher volume and a lower firmness [10]. Production of levan and CPS have been also described for some W. confusa strains [152]. Since Weissella spp. are able to grow in sourdough and produce a high amount of EPS, they present an interest as starters for industrial production of sourdough.
The effect of EPS-producing Weissella was also investigated on vegetable puree. Dextran from W. confusa and Lc. lactis was able to improve the texture and sensory properties of pureed carrots, with pleasant odor and flavor [158].
5.3. Production of Antimicrobial Substances
Ayeni et al. [68] determined lactic acid, acetaldehyde, acetone and ethanol amount production of W. confusa, Lb. brevis and Lb. paracasei strains after 24 h of culture in MRS broth. W. confusa strains produced 13.2 to 15.9 mg·mL−1 lactic acid, 15.8 to 32.8 μg·mL−1 acetone, 35.1 to 64.0 μg·mL−1 acetaldehyde and 2.7 to 5.0 mg·mL−1 ethanol while Lactobacillus strains produced 13.7 to 21.8 mg·mL−1 lactic acid, 4.0 to 38.1 μg·mL−1 acetone, 12.6 to 42.4 μg·mL−1 acetaldehyde and 0.01 to 4.3 mg·mL−1 ethanol in the same conditions. The lowest production of lactic acid was observed for W. confusa. However, the highest productions of acetaldehyde and ethanol were observed for W. confusa strains. W. koreensis HO20 has been shown to produce more ethanol than Lc. citreum HO12 during sourdough fermentation (4.7 and 3.0 mmol·kg−1 dough) [147].
W. cibaria and W. confusa strains showed the same antimicrobial activity against E. coli K12 than Lb. fermentum, Lb. plantarum and P. curvatus strains, however this activity was strain dependent. W. cibaria strains showed zones of inhibition comprised between 14.5 and 23.5 mm, while those of Lb. fermentum strains were comprised between 16.2 to 23.2 mm. Anyway, the highest antagonist activity against E. coli was observed for W. cibaria 142 [60]. In addition, W. confusa has been shown to inhibit the growth of Helicobacter pylori [159].
Bacteriocins isolated from Weissella spp. have been extensively reviewed by [18]. Weissellicin 110, produced by W. cibaria 110 was active against several LAB, including Lactobacillus, Leuconostoc and Weissella species [117]. Several other bacteriocins have been purified from W. hellenica strains (weissellicins D, L, M, Y, 7293A and 7293B) and W. paramesenteroides (weissellicin A) [114,160,161,162,163]. Weissellicin L presented a high homology with leucocin B-TA33a, produced by Lc. mesenteroides TA33a. More recently, Weissellicin MBF has been characterized from W. confusa [164]. Interestingly, weissellicin MBF is encoded by a large transferable plasmid of 17.6 kbp, pWcMBF8-1. To the best of our knowledge, this is the first study which reported plasmid-associated bacteriocin activity in the genus Weissella.
5.4. Probiotic Aspects
Probiotics can be defined as living microorganisms which once ingested exert health beneficial properties. The criteria for the selection of probiotics are: (1) the ability to survive through the passage into the human gastrointestinal tract (GIT), that is to say low pH in the stomach and bile salts in the small intestine; (2) a human origin and isolation from healthy human GIT; (3) a non-pathogenic history; (4) not being associated with diseases; (5) not carrying transmissible antibiotic resistance genes and (6) the Generally Recognized as Safe (GRAS) status. Today, LAB associated with probiotic potential mainly belong to Lactobacillus and Bifidobacterium genera [165]. Recently, investigation of probiotic effect of Weissella spp. have gained [55,60,166].
Patel et al. [60] reported that W. confusa AI10 was the most resistant strain to bile salts (0.3%), with 72% of survival after 24 h at 37 °C. On the contrary, Lb. plantarum AD29 was the less resistant, with 14% of survival. In the same study, W. cibaria 142 showed 131% of survival after 2.5 h at 37 °C in MRS broth adjusted to pH 3, which demonstrated the ability of this strain to grow in acidic conditions. Reference [107] related that 100% of Lb. plantarum strains were resistant to 0.3% of bile salts and none of those strains were able to grow at pH 3. Anandharaj et al. [55] reported that W. koreensis FKI21 was the best resistant strain to pH 1.0, with 29.8% of survival, while Lb. crispatus GI6 showed the lower resistance with 18.3% of survival. In this same study, W. koreensis and Lb. crispatus strains showed approximately the same resistance profile to 0.3% and 0.5% of bile salts. Hence, resistance to bile salts and to acidic conditions are strain dependent and regarding these criteria, probiotic activity should not be assigned to one specific LAB genus or species.
5.5. Role in Bioavailability and Antioxidant Activities
The increase of antioxidant activity occurring over lactic acid fermentation has been related to the activity of several enzymes, such as β-glucosidase (E.C. 3.2.1.21), tannase (E.C. 3.1.1.20), p-coumaric acid decarboxylase (PAD) (E.C. 4.1.1.102) or feruloyl esterase (E.C. 3.1.1.73). These enzymes have been largely described in Lactobacillus and Bifidobacterium spp. [167,168,169,170,171]. Concerning Weissella spp., only a few studies reported these activities.
β-glucosidases catalyze the hydrolysis of diverse compounds with β-D-glucosidic linkages, like oligosaccharides, cell wall components, pigments and cyanogenic glucosides [172]. β-glucosidases have been used in lactic acid fermentation to convert isoflavones glucosides into their corresponding aglycones [173]. Isoflavones constitute a subfamily of the flavonoids. It has been suggested that isoflavones aglycones are absorbed faster and in larger amounts that their corresponding glucosides in humans. Both tannase and β-glucosidase activities were reported for Lb. plantarum, Lb. pentosus, Lc. mesenteroides subsp. mesenteroides and W. cibaria strains, whereas only β-glucosidase activity was detected for W. paramesenteroides BFE 7601 [85]. W. cibaria and W. confusa strains showed up to 50-fold higher β-glucosidase activity than L. lactis subsp. cremoris. W. confusa has been shown to increase aglycone level in fermented soymilk as well as Lb. paraplantarum and Streptococcus salivarius [173]. Soymilk fermented for 12 h with Weissella spp. 4 (not identified) was 100-fold lower in isoflavones glucosides and 3-fold higher in aglycones than the one fermented with Streptococcus infantarius [174].
Phenolic acid decarboxylase is responsible for the decarboxylation of caffeic, ρ-coumaric and ferulic acids into their corresponding vinyl derivatives vinylcatechol, ρ-vinylphenol and vinlguaiacol, respectively. These vinyl derivatives can be reduced into ethylcatechol, ethylphenol and ethylguaiacol, respectively by the phenolic acid reductase. W. cibaria/confusa strains were shown to be able to reduce and decarboxylate caffeic, ρ-coumaric and ferulic acids. Lb. brevis, Lb. fermentum, Lb. curvatus, Lc. mesenteroides and Lb. rossiae were also able to metabolize these phenolic compounds [175]. Kimchi started with W. koreensis OK1-6 has been shown to decrease epidydimal fat pad weight, leptin, triglyceride and cholesterol levels in obese mice [176], suggesting anti-obesity and anti-cholesterol effects of W. koreensis OK1-6.
6. Possible Constraints for the Use of Weissella spp. as Starters
6.1. Biogenic Amine Production of Weissella
Amino acids like histidine, lysine, ornithine and tyrosine can be decarboxylated by LAB in histamine, cadaverine, putrescine and tyramine, respectively. This ability is highly variable and often strain specific [177]. No production of biogenic amine from histidine, lysine, ornithine or tyrosine has been detected for W. confusa strains [118] neither by W. cibaria strains [178] nor by W. viridescens [110]. In these studies, the method used for the detection of biogenic amines was a qualitative method using bromocresol purple as a pH indicator. Using this method, E. faecium and E. faecalis were able to produce biogenic amines [110]. The detection of genes encoding amino acid decarboxylase would help to confirm these observations. On the contrary, some Weissella strains (W. cibaria, W. confusa, W. paramesenteroides) have been shown to produce higher amount of biogenic amines (cadaverine, putrescine, histamine and tyramine) than Leuconostoc strains [179]. W. halotolerans W22 and Lb. hilgardii X1B were able to degrade arginine in putrescine using arginine deiminase or ornithine and arginine decarboxylation pathways [111,180]. Some strains of Lb. brevis, Lb. hilgardii and P. pentosaceus were able to produce tyramine and putrescine [181]. W. hellenica 15.32 has been shown to produce only histamine whereas Lc. mesenteroides and Lb. sakei strains produced histamine plus tyramine in the same conditions [182]. As for other LAB species, biogenic amine production was strain-dependent and some Weissella strains did not produce any of the biogenic amine searched.
6.2. Antibiotic Resistance Profile of Weissella
Antibiotics are frequently used by the health care industries to fight infections. The excessive and inappropriate use of antibiotics lead these last decades to a development of antibiotic resistance in bacteria. Nowadays, it is clearly established that LAB possess several antibiotic resistance genes, and the problem is that they can transfer these genes to pathogenic bacteria [183]. Hence, antibiotic resistance of LAB should be taken carefully regarding their use as starters or probiotics. Several studies reported antibiotic resistance profile of Weissella spp. W. cibaria, W. confusa and W. paramesenteroides strains have been shown to be resistant to streptomycin and gentamicin. Only W. cibaria strains were resistant to penicillin G [179]. Several strains of Lb. fermentum, Lb. plantarum, W. cibaria, W. confusa and P. parvulus were resistant to vancomycin, norfloxacin, gentamicin and kanamycin. The same strains were sensitive to erythromycin, chloramphenicol, ampicillin and tetracycline. Only a few strains of W. cibaria, Lb. fermentum and Lb. plantarum were resistant to polymyxin [60]. D’Aimmo et al. [184] analyzed antibiotic resistance profile of several strains of Bifidobacterium animalis subsp. lactis, B. longum, Lb. acidophilus, Lb. casei and Lb. delbrueckii subsp. bulgaricus and S. thermophilus. All tested strains were resistant to aztreonam, cycloserin, kanamycin, polymyxin B and spectinomycin. Several probiotics LAB have been associated with antibiotic resistance such as Lb. reuteri ATCC 55148, Lb. brevis ATCC 8287 and Lb. plantarum DG507 which have been shown to be resistant to vancomycin and erythromycin [185]. Since, Lactobacillus have been associated with fermented foods consumed for decades, they are considered as safe for humans. Hence, antibiotic resistance profile of these species is often neglected. From our point of view, Weissella genus should not be associated systematically with antibiotic resistance patterns on the grounds that some strains presented resistance to a low number of antibiotics. Besides, Weissella spp. share numerous habitats with Lactobacillus, Leuconostoc, Lactococcus and Pediococcus. All of these species have been associated with antibiotic resistance and each of them may transfer antibiotic resistance gene to each other. As evidence from literature data, antibiotic resistance is strain-dependent, and a careful analysis of each LAB strain should be realized before its application as starters for food fermentation.
6.3. Infections Associated with Weissella
Of the 21 Weissella species, only strains of W. cibaria, W. confusa, W. paramesenteroides and W. viridescens have been isolated from human clinical samples. W. viridescens and W. cibaria have been associated with healthy human vaginal microbiota, together with Lactobacillus, Leuconostoc, Pediococcus, Streptococcus and Enterococcus [186,187,188]. Lee, (2005) [188] suggested W. cibaria PL9023 (W. kimchi) as a probiotic for women, since it has been shown to inhibit the growth of vaginal pathogens like E. coli, Candida albicans and Streptococcus agalactiae. W. confusa, W. cibaria and W. paramesenteroides have been isolated from healthy human feces [166,189,190,191,192]. Interestingly, Weissella spp. mainly isolated from healthy humans (W. cibaria, W. confusa, W. paramesenteroides, W. viridescens) are the same species frequently isolated from fermented foods. Since, they have been isolated from human vagina and feces, Weissella spp. may adhere and colonize the human GIT. These observations support data regarding resistance of Weissella to bile salts and acidic conditions.
Despite the fact that numerous Weissella spp. have been isolated from healthy humans and are suggested as probiotics, they are often classified as opportunistic pathogens since they were occasionally associated with infections. Actually, four Weissella species have been associated with health disorders and infections, both in animals or humans: W. cibaria, W. confusa, W. viridescens and W. ceti. W. ceti is considered to be a pathogen which infects fish and rainbow trout [193,194]. Genome sequence analysis of W. ceti strains revealed the presence of several virulence factors and antibiotic resistance genes [195], which comforts its potential pathogenicity. To the best of our knowledge, no study reported infections associated W. ceti in humans. W. cibaria and W. viridescens have been associated with bacteremia [18] and W. cibaria and W. confusa have been also isolated from dog ear otitis [37]. Concerning W. confusa, several cases reported bacteremia and infections [196,197,198,199,200,201,202,203,204,205,206]. In almost all cases of infections associated W. confusa, patients were immunocompromised or have undergone chemotherapy or organ transplant [207]. These organisms’ weaknesses may facilitate the entry of W. confusa in the body. In almost all cases of W. confusa infections, vancomycin-resistance profile of these species has been point out.
In parallel, [208] reported more than 200 cases of related Lactobacillus associated infections. The Lactobacillus species most frequently associated with bacteremia are Lb. casei and Lb. rhamnosus. Lactobacillus spp. have been shown to be resistant to vancomycin and erythromycin in 27% and 97% of bacteremia associated Lactobacillus, respectively. Surprisingly, Lactobacillus are the main species used as starters or probiotics in food industries. Their potential pathogenicity does not deprive them of their safe status. Because immunodeficiency was the main risk factor of Lactobacillus bacteremia, [209] suggested that Lactobacillus bacteremia should be considered as a negative prognostic marker rather than a pathogen, suggesting comorbidities. Drawing a parallel between the two genera, Weissella spp. should not be considered as pathogens because W. confusa have been associated with bacteremia.
6.4. Bacteriophage Infection
Bacteriophages are ubiquitous, they are considered as the most predominant biological materials present in the world [210]. Phages infecting LAB have been associated with fermented products with inferior quality, especially in dairy products. Adsorption of phage particles to bacterial cells is the initial step of phage infection, then after phages inactivate and lyse microbial cells [211]. Only a few studies reported phages infecting Weissella spp. Bacteriophage Φ22 from W. cibaria N22 was isolated from Nham, a Thai fermented pork sausage [212]. Both Φ22 and ΦYS61 belong to the Podoviridae phage family and harbor size genome of 29 Kb and 33.6 Kb, respectively. Lu et al. [62] isolated 57 phages infecting W. cibaria, W. paramesenteroides, Lb. plantarum and Lb. brevis in commercial cucumber fermentation. Some phages were able to infect species from different genera, for instance Φ3.2.27 was able to infect W. cibaria, Lb. plantarum and Lb. brevis. Interestingly, all Weissella hosts were isolated from day 3 of the fermentation process. The same authors isolated 28 bacteriophages infecting Lc. citreum, Lc. mesenteroides, Lc. pseudomesenteroides, Lc. fallax, Weissella spp., Lb. plantarum, Lb. brevis and Lb. paraplantarum in commercial sauerkraut [66]. Interestingly, all hosts isolated from days 1 and 3 of the fermentation process belong to Leuconostoc and Weissella, while all hosts isolated after 3 days of fermentation belong to Lactobacillus. These observations are correlated to the dominance of Weissella spp. in fermented foods, where they appeared generally at the beginning of the fermentation together with Leuconostoc spp. The authors suggested that bacteriophages may play an important role in the LAB succession occurred in vegetables fermentations. Hence, bacteriophages of Weissella would require a deeper investigation. The disappearance of Weissella observed in vegetables fermentation may be the consequence of dying cells caused by phage infections. The majority of phages infecting Leuconostoc spp. were isolated from dairy products but vegetable fermentation constitutes another source of phages infecting Leuconostoc, Weissella and Lactobacillus spp. [211]. Among LAB, L. lactis, S. thermophilus and Lactobacillus are the main bacteriophages infecting hosts. Indeed, the genome sequence of 58 phages infecting L. lactis have been published [213]. Repeated use of the same starter culture in the same raw material has been suggested to be a risk of phage infection. Thus, development of new starter cultures would be useful both for consumers and food industries.
6.5. Regulations
A substance commonly used in food prior to 1958 is recognized as GRAS (Generally Recognized as Safe) by the Food and Drug Administration (FDA) and the list includes fermentative microorganisms. Since 2007, food microorganisms with a safety assessment are listed as QPS (Qualified Presumption of Safety) by the European Food Safety Authority (EFSA). Probiotics are alive microorganisms which, when ingested in adequate numbers exert a health benefit on the host. Species such as Enterococcus faecium, Lb. plantarum, Lb. acidophilus and Lb. casei subsp. rhamnosus possess the GRAS status [185]. Lactobacillus, Bifidobacterium, S. thermophilus and E. faecium are the probiotics mainly used in the food industry [214].
Bourdichon et al. [215] created a list called “Inventory of Microbial Food Cultures”. The microbial species with a documented presence in fermented foods were included in this list. On the contrary, the species considered as undesirable in food, that do not harbor interested metabolic activity and with a lack of data related interested criteria in food fermentation were excluded of the list. Interestingly, the authors included Weissella species in this list as microorganisms with beneficial use.
However, Weissella spp. are still not recognized as GRAS by the FDA neither as QPS by the EFSA, so not as a probiotic since GRAS status is required as criteria for probiotic status.
Weissella spp. are frequently associated with Lactobacillus, Leuconotocs, Lactococcus and Pediococcus in different habitats, and in food fermentation Weissella spp. participate to the process but Lactobacillus spp. remained generally the dominant species. Since, Lactobacillus are used for centuries in food fermentation, they possess the GRAS status. Industrial people easily prefer using recognized safe LAB rather than those which are not.
7. Concluding Remarks
Starter selection is a complex and long process which requires several steps: (1) isolation and in vitro selection; (2) validation on a lab-scale; (3) validation at factory-scale [216]. Bevilacqua et al. [216] developed a general flow-sheet for the selection of suitable yeast starter cultures, but it can easily be adapted for LAB (Figure 2). Starter cultures may consist of a single culture or a multiple culture containing a mixture of several strains. Self-propagation of the starter culture was initially the preferred method, but with this method several LAB characteristics were lost [8] and it was gradually replaced by ready-to-use frozen, freeze-dried or lyophilized highly concentrated cultures [3,139]. However, the manufacturing of frozen and freeze-dried cultures are long and expensive steps, limiting the production and the development of starter.
Weissella species harbor different sources of habitats, fermented foods, animal, environment and human sources. They share these habitats with LAB commonly used as starters or probiotics, like Lactobacillus, Leuconostoc, Lactococcus and Pediococcus species. Weissella spp. are extensively involved in spontaneous fermented foods, especially fruits and vegetables based products, in which they could dominate the process.
Nowadays, Weissella proposal as starter is still under investigation at the lab-scale, whereas the genus was described 24 years ago. Weissella do not possess the GRAS status so it probably limits investigation for use as starter. From the literature, scientists are divided about using Weissella spp. as starters. Weissella are often classified as opportunistic pathogens, which seems to be a limit for food application. Some LAB species used as commercial starters have been associated with biogenic amine production, antibiotic resistance profile and infections. Surprisingly, these non-expected characteristics do not deprive them of their GRAS status and even less of their commercial starter status. Infections associated Weissella strains do not justify their rejection as commercial starters, especially since infections associated Weissella is strain-dependent. Weissella spp. can be used as commercial starter but safety aspects of the selected Weissella strain need to be investigated.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ross, P.R.; Morgan, S.; Hill, C. Preservation and fermentation: Past, present and future. Int. J. Food Microbiol. 2002, 79, 3–16. [Google Scholar] [CrossRef]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Review: Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef] [PubMed]
- Buckenhüskes, H.J. Selection criteria for lactic acid bacteria to be used as starter cultures for various food commodities. FEMS Microbiol. Rev. 1993, 12, 253–271. [Google Scholar] [CrossRef]
- Caplice, E.; Fitzgerald, G.F. Food fermentations: Role of microorganisms in food production and preservation. Int. J. Food Microbiol. 1999, 50, 131–149. [Google Scholar] [CrossRef]
- Stiles, M.E.; Holzapfel, W.H. Lactic acid bacteria of foods and their current taxonomy. Int. J. Food Microbiol. 1997, 36, 1–29. [Google Scholar] [CrossRef]
- Brandt, M.J. Starter cultures for cereal based foods. Food Microbiol. 2014, 37, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Coda, R.; De Angelis, M.; Gobbetti, M. Exploitation of vegetables and fruits through lactic acid fermentation. Food Microbiol. 2013, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Amari, M.; Arango, L.F.G.; Gabriel, V.; Robert, H.; Morel, S.; Moulis, C.; Gabriel, B.; Remaud-Siméon, M.; Fontagné-Faucher, C. Characterization of a novel dextransucrase from Weissella confusa isolated from sourdough. Appl. Microbiol. Biotechnol. 2013, 97, 5413–5422. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; De Angelis, M.; Limitone, A.; Minervini, F.; Carnevali, P.; Corsetti, A.; Gaenzle, M.; Ciati, R.; Gobbetti, M. Glucan and fructan production by sourdough Weissella cibaria and Lactobacillus plantarum. J. Agric. Food Chem. 2006, 54, 9873–9881. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, S.H.; Jeon, C.O. Kimchi microflora: History, current status, and perspectives for industrial kimchi production. Appl. Microbiol. Biotechnol. 2014, 98, 2385–2393. [Google Scholar] [CrossRef] [PubMed]
- Nuraida, L.; Wacher, M.C.; Owens, J.D. Microbiology of pozol, a Mexican maize dough. World J. Microbiol. Biotechnol. 1995, 11, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Wolkers-Rooijackers, J.C.M.; Thomas, S.M.; Nout, M.J.R. Effects of sodium reduction scenarios on fermentation and quality of sauerkraut. LWT Food Sci. Technol. 2013, 54, 383–388. [Google Scholar] [CrossRef]
- Lynch, K.M.; Lucid, A.; Arendt, E.K.; Sleator, R.D.; Lucey, B.; Coffey, A. Genomics of Weissella cibaria with an examination of its metabolic traits. Microbiology 2015, 161, 914–930. [Google Scholar] [CrossRef] [PubMed]
- Chelo, I.M.; Zé-Zé, L.; Tenreiro, R. Genome diversity in the genera Fructobacillus, Leuconostoc and Weissella determined by physical and genetic mapping. Microbiology 2010, 156, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Padonou, S.W.; Schillinger, U.; Nielsen, D.S.; Franz, C.M.A.P.; Hansen, M.; Hounhouigan, J.D.; Nago, M.C.; Jakobsen, M. Weissella beninensis sp. nov., a motile lactic acid bacterium from submerged cassava fermentations, and emended description of the genus Weissella. Int. J. Syst. Evol. Microbiol. 2010, 60, 2193–2198. [Google Scholar] [CrossRef] [PubMed]
- Vela, A.I.; Fernández, A.; de Quirós, Y.B.; Herráez, P.; Domínguez, L.; Fernández-Garayzábal, J.F. Weissella ceti sp. nov., isolated from beaked whales (Mesoplodon bidens). Int. J. Syst. Evol. Microbiol. 2011, 61, 2758–2762. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Quero, G.M.; Cho, G.-S.; Kabisch, J.; Meske, D.; Neve, H.; Bockelmann, W.; Franz, C.M.A.P. The genus Weissella: Taxonomy, ecology and biotechnological potential. Front. Microbiol. 2015, 6, 155. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.D.; Samelis, J.; Metaxopoulos, J.; Wallbanks, S. Taxonomic studies on some Leuconostoc-like organisms from fermented sausages: Description of a new genus Weissella for the Leuconostoc paramesenteroides group of species. J. Appl. Bacteriol. 1993, 75, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Kandler, O.; Schillinger, U.; Weiss, N. Lactobacillus halotolerans sp.nov., nom.rev. and Lactobacillus minor sp.nov., nom.rev. Syst. Appl. Microbiol. 1983, 4, 280–285. [Google Scholar] [CrossRef]
- Björkroth, J.; Holzapfel, W. Genera Leuconostoc, Oenococcus and Weissella. In The Prokaryotes; Springer: New York, NY, USA, 2006; pp. 267–319. [Google Scholar]
- Dicks, L.M.T.; Dellaglio, F.; Collins, M.D. Proposal To Reclassify Leuconostoc oenos as Oenococcus oeni (corrig.) gen. nov., comb. nov. Int. J. Syst. Bacteriol. 1995, 45, 395–397. [Google Scholar] [CrossRef] [PubMed]
- Endo, A.; Okada, S. Reclassification of the genus Leuconostoc and proposals of Fructobacillus fructosus gen. nov., comb. nov., Fructobacillus durionis comb. nov.; Fructobacillus ficulneus comb. nov. and Fructobacillus pseudoficulneus comb. nov. Int. J. Syst. Evol. Microbiol. 2008, 58, 2195–2205. [Google Scholar] [CrossRef] [PubMed]
- Niven, C.F.; Evans, J.B. Lactobacillus viridescens nov. spec., a heterofermentative species that produces a green discoloration of cured meat pigments. J. Bacteriol. 1957, 73, 758–759. [Google Scholar] [PubMed]
- Nisiotou, A.; Dourou, D.; Filippousi, M.-E.; Banilas, G.; Tassou, C. Weissella uvarum sp. nov.; isolated from wine grapes. Int. J. Syst. Evol. Microbiol. 2014, 64, 3885–3890. [Google Scholar] [CrossRef] [PubMed]
- Snauwaert, I.; Papalexandratou, Z.; De Vuyst, L.; Vandamme, P. Characterization of strains of Weissella fabalis sp. nov. and Fructobacillus tropaeoli from spontaneous cocoa bean fermentations. Int. J. Syst. Evol. Microbiol. 2013, 63, 1709–1716. [Google Scholar] [CrossRef] [PubMed]
- De Bruyne, K.; Camu, N.; De Vuyst, L.; Vandamme, P. Weissella fabaria sp. nov.; from a Ghanaian cocoa fermentation. Int. J. Syst. Evol. Microbiol. 2010, 60, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- De Bruyne, K.; Camu, N.; Lefebvre, K.; De Vuyst, L.; Vandamme, P. Weissella ghanensis sp. nov.; isolated from a Ghanaian cocoa fermentation. Int. J. Syst. Evol. Microbiol. 2008, 58, 2721–2725. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-S.; Lee, S.-H.; Ahn, M.-J.; Ku, H.-J.; Hong, J.-S.; Jeon, C.O.; Lee, J.-H.; Lee, S.H.; Ryu, S.; Shin, H.; et al. Weissella jogaejeotgali sp. nov., isolated from jogae jeotgal, a traditional Korean fermented seafood. Int. J. Syst. Evol. Microbiol. 2015, 65, 4674–4681. [Google Scholar] [CrossRef] [PubMed]
- Tanasupawat, S.; Shida, O.; Okada, S.; Komagata, K. Lactobacillus acidipiscis sp. nov. and Weissella thailandensis sp. nov., isolated from fermented fish in Thailand. Int. J. Syst. Evol. Microbiol. 2000, 50, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Fessard, A.; Kapoor, A.; Patche, J.; Assemat, S.; Hoarau, M.; Bourdon, E.; Bahorun, T.; Remize, F. Lactic Fermentation as an Efficient Tool to Enhance the Antioxidant Activity of Tropical Fruit Juices and Teas. Microorganisms 2017, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Garvie, E.I. The Growth Factor and Amino Acid Requirements of Species of the Genus Leuconostoc, including Leuconostoc paramesenteroides (sp.nov.) and Leuconostoc oenos. J. Gen. Microbiol. 1967, 48, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Praet, J.; Meeus, I.; Cnockaert, M.; Houf, K.; Smagghe, G.; Vandamme, P. Novel lactic acid bacteria isolated from the bumble bee gut: Convivina intestini gen. nov., sp. nov., Lactobacillus bombicola sp. nov., and Weissella bombi sp. nov. Antonie Leeuwenhoek 2015, 107, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Shin, N.R.; Hyun, D.W.; Kim, P.S.; Kim, J.Y.; Kim, M.S.; Yun, J.H.; Bae, J.W. Weissella diestrammenae sp. nov.; isolated from the gut of a camel cricket (Diestrammena coreana). Int. J. Syst. Evol. Microbiol. 2013, 63, 2951–2956. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, W.H.; Van Wyk, E.P. Lactobacillus kandleri sp. nov., a new species of the subgenus betabacterium, with glycine in the peptidoglycan. Zentralblatt Bakteriol. Mikrobiol. Hyg. I. Abt. Orig. C Allg. Angew. Ökologische Mikrobiol. 1982, 3, 495–502. [Google Scholar] [CrossRef]
- Lee, J.; Lee, K.C.; Ahn, J.; Mheen, T.; Pyun, Y.; Park, Y. Weissella koreensis sp. nov.; isolated from kimchi. Int. J. Syst. Evol. Microbiol. 2002, 52, 1257–1261. [Google Scholar] [CrossRef] [PubMed]
- Björkroth, K.J.; Schillinger, U.; Geisen, R.; Weiss, N.; Hoste, B.; Holzapfel, W.H.; Korkeala, H.J.; Vandamme, P. Taxonomic study of Weissella confusa and description of Weissella cibaria sp. nov.; detected in food and clinical samples. Int. J. Syst. Evolut. Microbiol. 2002, 52, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Beleggia, R.; Ferrer, S.; Pardo, I.; Spano, G. A polyphasic approach in order to identify dominant lactic acid bacteria during pasta manufacturing. LWT Food Sci. Technol. 2010, 43, 982–986. [Google Scholar] [CrossRef]
- Osimani, A.; Garofalo, C.; Aquilanti, L.; Milanović, V.; Clementi, F. Unpasteurised commercial boza as a source of microbial diversity. Int. J. Food Microbiol. 2015, 194, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, J.; Jonsson, H.; Schnurer, J.; Roos, S. Weissella soli sp. nov., a lactic acid bacterium isolated from soil. Int. J. Syst. Evol. Microbiol. 2002, 52, 831–834. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Lee, D.; Yang, C.; Jeon, J.; Kim, J.; Han, H. Microbial population dynamics of kimchi, a fermented cabbage product. FEMS Microbiol. Lett. 2006, 257, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Lee, S.H.; Jung, J.Y.; Choi, E.J.; Jeon, C.O. Microbial succession and metabolite changes during long-term storage of Kimchi. J. Food Sci. 2013, 78, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Cho, Y.; Lee, Y.; Han, S.-K.; Kim, C.-G.; Choo, D.-W.; Kim, Y.-R.; Kim, H.-Y. A proteomic approach for rapid identification of Weissella species isolated from Korean fermented foods on MALDI-TOF MS supplemented with an in-house database. Int. J. Food Microbiol. 2017, 243, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Chun, J. Bacterial community structure in kimchi, a Korean fermented vegetable food, as revealed by 16S rRNA gene analysis. Int. J. Food Microbiol. 2005, 103, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Seo, H.N.; Hwang, T.S.; Lee, S.H.; Park, D.H. Characterization of exopolysaccharide (EPS) produced by Weissella cibaria SKkimchi3 isolated from kimchi. J. Microbiol. 2008, 46, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Heo, G.Y.; Jun, W.L.; Oh, Y.J.; Park, J.A.; Park, Y.H.; Pyun, Y.R.; Jong, S.A. Analysis of kimchi microflora using denaturing gradient gel electrophoresis. Int. J. Food Microbiol. 2005, 102, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.-T.T.; Chen, Y.-S.; Yanagida, F. Isolation and characterization of lactic acid bacteria from Yan-dong-gua (fermented wax gourd), a traditional fermented food in Taiwan. J. Biosci. Bioeng. 2009, 108, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-S.; Wu, H.-C.; Wang, C.-M.; Lin, C.-C.; Chen, Y.-T.; Jhong, Y.-J.; Yanagida, F. Isolation and characterization of lactic acid bacteria from pobuzihi (fermented cummingcordia), a traditional fermented food in Taiwan. Folia Microbiol. 2013, 58, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Wu, H.; Lo, H.; Lin, W.; Hsu, W.; Lin, C.; Lin, P.; Yanagida, F. Isolation and characterisation of lactic acid bacteria from jiang-gua (fermented cucumbers), a traditional fermented food in Taiwan. J. Sci. Food Agric. 2012, 92, 2069–2075. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-S.; Liou, M.S.; Ji, S.H.; Yu, C.R.; Pan, S.F.; Yanagida, F. Isolation and characterization of lactic acid bacteria from Yan-tsai-shin (fermented broccoli stems), a traditional fermented food in Taiwan. J. Appl. Microbiol. 2013, 115, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-H.; Chen, Y.-S.; Yanagida, F. Isolation and characterisation of lactic acid bacteria from yan-jiang (fermented ginger), a traditional fermented food in Taiwan. J. Sci. Food Agric. 2011, 91, 1746–1750. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-S.; Wu, H.C.; Pan, S.F.; Lin, B.G.; Lin, Y.H.; Tung, W.C.; Li, Y.L.; Chiang, C.M.; Yanagida, F. Isolation and characterization of lactic acid bacteria from yan-taozih (pickled peaches) in Taiwan. Ann. Microbiol. 2013, 63, 607–614. [Google Scholar] [CrossRef]
- Chen, Y.-S.S.; Wu, H.C.; Yu, C.R.; Chen, Z.Y.; Lu, Y.C.; Yanagida, F. Isolation and characterization of lactic acid bacteria from xi-gua-mian (fermented watermelon), a traditional fermented food in Taiwan. Ital. J. Food Sci. 2016, 28, 9–14. [Google Scholar] [CrossRef]
- Chen, Y.-S.; Yanagida, F.; Hsu, J.S. Isolation and characterization of lactic acid bacteria from dochi (fermented black beans), a traditional fermented food in Taiwan. Lett. Appl. Microbiol. 2006, 43, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Anandharaj, M.; Sivasankari, B.; Santhanakaruppu, R.; Manimaran, M.; Rani, R.P.; Sivakumar, S. Determining the probiotic potential of cholesterol-reducing Lactobacillus and Weissella strains isolated from gherkins (fermented cucumber) and south Indian fermented koozh. Res. Microbiol. 2015, 166, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Chao, S.-H.; Wu, R.-J.; Watanabe, K.; Tsai, Y.-C. Diversity of lactic acid bacteria in suan-tsai and fu-tsai, traditional fermented mustard products of Taiwan. Int. J. Food Microbiol. 2009, 135, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Chao, S.-H.; Tomii, Y.; Watanabe, K.; Tsai, Y.-C. Diversity of lactic acid bacteria in fermented brines used to make stinky tofu. Int. J. Food Microbiol. 2008, 123, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, M.; Malimas, T.; Sugimoto, M.; Yoshino, M. 16S rRNA gene sequence analysis of lactic acid bacteria isolated from fermented foods in Thailand. Microb. Cult. Coll. 2012, 28, 1–9. [Google Scholar]
- Roh, S.W.; Kim, K.; Nam, Y.; Chang, H.; Park, E.; Bae, J.-W. Investigation of archaeal and bacterial diversity in fermented seafood using barcoded pyrosequencing. ISME J. 2009, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Prajapati, J.B.; Holst, O.; Ljungh, A. Probiotic properties of exopolysaccharide producing lactic acid bacteria isolated from vegetables and traditional Indian fermented foods. Food Biosci. 2012, 5, 27–33. [Google Scholar] [CrossRef]
- Yuliana, N.; Dizon, E.I. Phenotypic Identification of Lactic Acid Bacteria Isolated from Tempoyak (Fermented Durian) Made in the Philippines. Int. J. Biol. 2011, 3, 145–152. [Google Scholar] [CrossRef]
- Lu, Z.; Pérez-Díaz, I.M.; Hayes, J.S.; Breidt, F. Bacteriophage ecology in a commercial cucumber fermentation. Appl. Environ. Microbiol. 2012, 78, 8571–8578. [Google Scholar] [CrossRef] [PubMed]
- Wouters, D.; Grosu-Tudor, S.; Zamfir, M.; De Vuyst, L. Bacterial community dynamics, lactic acid bacteria species diversity and metabolite kinetics of traditional Romanian vegetable fermentations. J. Sci. Food Agric. 2013, 93, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ji, Y.; Park, H.; Lee, J.; Park, S.; Yeo, S.; Shin, H.; Holzapfel, W.H. Selection of functional lactic acid bacteria as starter cultures for the fermentation of Korean leek (Allium tuberosum Rottler ex Sprengel.). Int. J. Food Microbiol. 2014, 191, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Wouters, D.; Bernaert, N.; Conjaerts, W.; Van Droogenbroeck, B.; de Loose, M.; De Vuyst, L. Species diversity, community dynamics, and metabolite kinetics of spontaneous leek fermentations. Food Microbiol. 2013, 33, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Breidt, F.; Plengvidhya, V.; Fleming, H.P. Bacteriophage ecology in commercial sauerkraut fermentations. Appl. Environ. Microbiol. 2003, 69, 3192–3202. [Google Scholar] [CrossRef] [PubMed]
- Plengvidhya, V.; Breidt, F.; Lu, Z.; Fleming, H.P. DNA fingerprinting of lactic acid bacteria in sauerkraut fermentations. Appl. Environ. Microbiol. 2007, 73, 7697–7702. [Google Scholar] [CrossRef] [PubMed]
- Ayeni, F.A.; Sánchez, B.; Adeniyi, B.A.; de Los Reyes-Gavilán, C.G.; Margolles, A.; Ruas-Madiedo, P. Evaluation of the functional potential of Weissella and Lactobacillus isolates obtained from Nigerian traditional fermented foods and cow’s intestine. Int. J. Food Microbiol. 2011, 147, 97–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akabanda, F.; Owusu-Kwarteng, J.; Tano-Debrah, K.; Glover, R.L.K.; Nielsen, D.S.; Jespersen, L. Taxonomic and molecular characterization of lactic acid bacteria and yeasts in nunu, a Ghanaian fermented milk product. Food Microbiol. 2013, 34, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Mathara, J.M.; Schillinger, U.; Kutima, P.M.; Mbugua, S.K.; Holzapfel, W.H. Isolation, identification and characterisation of the dominant microorganisms of kule naoto: The Maasai traditional fermented milk in Kenya. Int. J. Food Microbiol. 2004, 94, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Isono, Y.; Shingu, I.; Shimizu, S. Identification and Characteristics of Lactic Acid Bacteria Isolated from Masai Fermented Milk in Northern Tanzania. Biosci. Biotechnol. Biochem. 1994, 58, 660–664. [Google Scholar] [CrossRef]
- Kimura, M.; Danno, K.; Yasui, H. Immunomodulatory Function and Probiotic Properties of Lactic Acid Bacteria Isolated from Mongolian Fermented Milk. Bioscience 2006, 25, 147–155. [Google Scholar] [CrossRef]
- Rahman, N.; Xiaohong, C.; Meiqin, F.; Mingsheng, D. Characterization of the dominant microflora in naturally fermented camel milk Shubat. World J. Microbiol. Biotechnol. 2009, 25, 1941–1946. [Google Scholar] [CrossRef]
- Di Cagno, R.; Buchin, S.; de Candia, S.; De Angelis, M.; Fox, P.F.; Gobbetti, M. Characterization of Italian cheeses ripened under nonconventional conditions. J. Dairy Sci. 2007, 90, 2689–2704. [Google Scholar] [CrossRef] [PubMed]
- Morales, F.; Morales, J.I.; Hernández, C.H.; Hernández-Sánchez, H. Isolation and partial characterization of halotolerant lactic acid bacteria from two mexican cheeses. Appl. Biochem. Biotechnol. 2011, 164, 889–905. [Google Scholar] [CrossRef] [PubMed]
- Gerasi, E.; Litopoulou-Tzanetaki, E.; Tzanetakis, N. Microbiological study of Manura, a hard cheese made from raw ovine milk in the Greek island Sifnos. Int. J. Dairy Technol. 2003, 56, 117–122. [Google Scholar] [CrossRef]
- Baruzzi, F.; Matarante, A.; Morea, M.; Cocconcelli, P.S. Microbial community dynamics during the Scamorza Altamurana cheese natural fermentation. J. Dairy Sci. 2002, 85, 1390–1397. [Google Scholar] [CrossRef]
- Morea, M.; Baruzzi, F.; Cappa, F.; Cocconcelli, P.S. Molecular characterization of the Lactobacillus community in traditional processing of Mozzarella cheese. Int. J. Food Microbiol. 1998, 43, 53–60. [Google Scholar] [CrossRef]
- Williams, A.G.; Banks, J.M. Proteolytic and other hydrolytic enzyme activities in non-starter lactic acid bacteria (NSLAB) isolated from Cheddar cheese manufactured in the United Kingdom. Int. Dairy J. 1997, 7, 763–774. [Google Scholar] [CrossRef]
- Zamfir, M.; Vancanneyt, M.; Makras, L.; Vaningelgem, F.; Lefebvre, K.; Pot, B.; Swings, J.; De Vuyst, L. Biodiversity of lactic acid bacteria in Romanian dairy products. Syst. Appl. Microbiol. 2006, 29, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wang, W.H.; Menghe, B.L.; Jiri, M.T.; Wang, H.M.; Liu, W.J.; Bao, Q.H.; Lu, Q.; Zhang, J.C.; Wang, F.; et al. Diversity of lactic acid bacteria associated with traditional fermented dairy products in Mongolia. J. Dairy Sci. 2011, 94, 3229–3241. [Google Scholar] [CrossRef] [PubMed]
- Mugula, J.; Nnko, S.A.; Narvhus, J.; Sørhaug, T. Microbiological and fermentation characteristics of togwa, a Tanzanian fermented food. Int. J. Food Microbiol. 2003, 80, 187–199. [Google Scholar] [CrossRef]
- Djeni, N.T.; Bouatenin, K.M.J.-P.; Assohoun, N.M.C.; Toka, D.M.; Menan, E.H.; Dousset, X.; Dje, K.M. Biochemical and microbial characterization of cassava inocula from the three main attieke production zones in Côte d’Ivoire. Food Control 2015, 50, 133–140. [Google Scholar] [CrossRef]
- Kostinek, M.; Specht, I.; Edward, V.A.A.; Pinto, C.; Egounlety, M.; Sossa, C.; Mbugua, S.; Dortu, C.; Thonart, P.; Taljaard, L.; et al. Characterisation and biochemical properties of predominant lactic acid bacteria from fermenting cassava for selection as starter cultures. Int. J. Food Microbiol. 2007, 114, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Kostinek, M.; Specht, I.; Edward, V.A.; Schillinger, U.; Hertel, C.; Holzapfel, W.H.; Franz, C.M.A.P. Diversity and technological properties of predominant lactic acid bacteria from fermented cassava used for the preparation of Gari, a traditional African food. Syst. Appl. Microbiol. 2005, 28, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Padonou, W.S.; Nielsen, D.S.; Hounhouigan, J.D.; Thorsen, L.; Nago, M.C.; Jakobsen, M. The microbiota of Lafun, an African traditional cassava food product. Int. J. Food Microbiol. 2009, 133, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Kavitake, D.; Devi, P.B.; Singh, S.P.; Shetty, P.H. Characterization of a novel galactan produced by Weissella confusa KR780676 from an acidic fermented food. Int. J. Biol. Macromol. 2016, 86, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Bounaix, M.-S.; Robert, H.; Gabriel, V.; Morel, S.; Remaud-Siméon, M.; Gabriel, B.; Fontagné-Faucher, C. Characterization of dextran-producing Weissella strains isolated from sourdoughs and evidence of constitutive dextransucrase expression. FEMS Microbiol. Lett. 2010, 311, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Bounaix, M.S.; Gabriel, V.; Morel, S.; Robert, H.; Rabier, P.; Remaud-Siméon, M.; Gabriel, B.; Fontagné-Faucher, C. Biodiversity of exopolysaccharides produced from sucrose by sourdough lactic acid bacteria. J. Agric. Food Chem. 2009, 57, 10889–10897. [Google Scholar] [CrossRef] [PubMed]
- Moroni, A.V.; Arendt, E.K.; Dal Bello, F. Biodiversity of lactic acid bacteria and yeasts in spontaneously-fermented buckwheat and teff sourdoughs. Food Microbiol. 2011, 28, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Pontonio, E.; Nionelli, L.; Curiel, J.A.; Sadeghi, A.; Di Cagno, R.; Gobbetti, M.; Rizzello, C.G. Iranian wheat flours from rural and industrial mills: Exploitation of the chemical and technology features, and selection of autochthonous sourdough starters for making breads. Food Microbiol. 2015, 47, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Robert, H.; Gabriel, V.; Fontagné-Faucher, C. Biodiversity of lactic acid bacteria in French wheat sourdough as determined by molecular characterization using species-specific PCR. Int. J. Food Microbiol. 2009, 135, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Ricciardi, A.; Parente, E. Enzymatic activities of lactic acid bacteria isolated from Cornetto di Matera sourdoughs. Int. J. Food Microbiol. 2007, 115, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Ampe, F.; Ben Omar, N.; Moizan, C.; Wacher, C.; Guyot, J.P. Polyphasic study of the spatial distribution of microorganisms in Mexican pozol, a fermented maize dough, demonstrates the need for cultivation-independent methods to investigate traditional fermentations. Appl. Environ. Microbiol. 1999, 65, 5464–5473. [Google Scholar] [PubMed]
- Tohno, M.; Kitahara, M.; Inoue, H.; Uegaki, R.; Irisawa, T.; Ohkuma, M.; Tajima, K. Weissella oryzae sp. nov., isolated from fermented rice grains. Int. J. Syst. Evol. Microbiol. 2013, 63, 1417–1420. [Google Scholar] [CrossRef] [PubMed]
- Elizaquível, P.; Pérez-Cataluña, A.; Yépez, A.; Aristimuño, C.; Jiménez, E.; Cocconcelli, P.S.; Vignolo, G.; Aznar, R. Pyrosequencing vs. culture-dependent approaches to analyze lactic acid bacteria associated to chicha, a traditional maize-based fermented beverage from Northwestern Argentina. Int. J. Food Microbiol. 2015, 198, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Oguntoyinbo, F.A.; Tourlomousis, P.; Gasson, M.J.; Narbad, A. Analysis of bacterial communities of traditional fermented West African cereal foods using culture independent methods. Int. J. Food Microbiol. 2011, 145, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Kwarteng, J.; Akabanda, F.; Nielsen, D.S.; Tano-Debrah, K.; Glover, R.L.K.; Jespersen, L. Identification of lactic acid bacteria isolated during traditional fura processing in Ghana. Food Microbiol. 2012, 32, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Abegaz, K. Isolation, characterization and identification of lactic acid bacteria involved in traditional fermentation of borde, an Ethiopian cereal beverage. Afr. J. Biotechnol. 2007, 6, 1469–1478. [Google Scholar]
- Camu, N.; De Winter, T.; Verbrugghe, K.; Cleenwerck, I.; Vandamme, P.; Takrama, J.S.; Vancanneyt, M.; De Vuyst, L. Dynamics and biodiversity of populations of lactic acid bacteria and acetic acid bacteria involved in spontaneous heap fermentation of cocoa beans in Ghana. Appl. Environ. Microbiol. 2007, 73, 1809–1824. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.-J.; Nam, Y.-D.; Roh, S.W.; Bae, J.-W. Unexpected convergence of fungal and bacterial communities during fermentation of traditional Korean alcoholic beverages inoculated with various natural starters. Food Microbiol. 2012, 30, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Pringsulaka, O.; Patarasinpaiboon, N.; Suwannasai, N.; Atthakor, W.; Rangsiruji, A. Isolation and characterisation of a novel Podoviridae-phage infecting Weissella cibaria N 22 from Nham, a Thai fermented pork sausage. Food Microbiol. 2011, 28, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Thongsanit, J.; Tanikawa, M.; Yano, S.; Tachiki, T.; Wakayama, M. Identification of glutaminase-producing lactic acid bacteria isolated from Nham, a traditional Thai fermented food and characterisation of glutaminase activity of isolated Weissella cibaria. Ann. Microbiol. 2009, 59, 715–720. [Google Scholar] [CrossRef]
- Wongsuphachat, W.; H-Kittikun, A.; Maneerat, S. Optimization of exopolysaccharides production by Weissella confusa NH 02 isolated from thai fermented sausages. Songklanakarin J. Sci. Technol. 2010, 32, 27–35. [Google Scholar]
- Cocolin, L.; Dolci, P.; Rantsiou, K.; Urso, R.; Cantoni, C.; Comi, G. Lactic acid bacteria ecology of three traditional fermented sausages produced in the North of Italy as determined by molecular methods. Meat Sci. 2009, 82, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Urso, R.; Comi, G.; Cocolin, L. Ecology of lactic acid bacteria in Italian fermented sausages: Isolation, identification and molecular characterization. Syst. Appl. Microbiol. 2006, 29, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Papamanoli, E.; Tzanetakis, N.; Litopoulou-Tzanetaki, E.; Kotzekidou, P. Characterization of lactic acid bacteria isolated from a Greek dry-fermented sausage in respect of their technological and probiotic properties. Meat Sci. 2003, 65, 859–867. [Google Scholar] [CrossRef]
- Rantsiou, K.; Drosinos, E.H.; Gialitaki, M.; Urso, R.; Krommer, J.; Gasparik-Reichardt, J.; Tóth, S.; Metaxopoulos, I.; Comi, G.; Cocolin, L. Molecular characterization of Lactobacillus species isolated from naturally fermented sausages produced in Greece, Hungary and Italy. Food Microbiol. 2005, 22, 19–28. [Google Scholar] [CrossRef]
- Albano, H.; van Reenen, C.A.; Todorov, S.D.; Cruz, D.; Fraga, L.; Hogg, T.; Dicks, L.M.T.; Teixeira, P. Phenotypic and genetic heterogeneity of lactic acid bacteria isolated from “Alheira”, a traditional fermented sausage produced in Portugal. Meat Sci. 2009, 82, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Yüceer, Ö.; Tuncer, B.O. Determination of antibiotic resistance and biogenic amine production of lactic acid bacteria isolated from fermented Turkish sausage (sucuk). J. Food Saf. 2015, 35, 276–285. [Google Scholar] [CrossRef]
- Pereira, C.I.; San Romão, M.V.; Lolkema, J.S.; Crespo, M.T.B. Weissella halotolerans W22 combines arginine deiminase and ornithine decarboxylation pathways and converts arginine to putrescine. J. Appl. Microbiol. 2009, 107, 1894–1902. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Ji, H.; Wang, Q.; Li, B.; Li, K.; Xu, C.; Jiang, C. The effects of starter cultures and plant extracts on the biogenic amine accumulation in traditional Chinese smoked horsemeat sausages. Food Control 2015, 50, 869–875. [Google Scholar] [CrossRef]
- Samelis, J.; Maurogenakis, F.; Metaxopoulos, J. Characterisation of lactic acid bacteria isolated from naturally fermented Greek dry salami. Int. J. Food Microbiol. 1994, 23, 179–196. [Google Scholar] [CrossRef]
- Chen, C.; Chen, X.; Jiang, M.; Rui, X.; Li, W.; Dong, M. A newly discovered bacteriocin from Weissella cibaria D1501 associated with Chinese Dong fermented meat (Nanx Wudl). Food Control 2014, 42, 116–124. [Google Scholar] [CrossRef]
- Kopermsub, P.; Yunchalard, S. Identification of lactic acid bacteria associated with the production of plaa-som, a traditional fermented fish product of Thailand. Int. J. Food Microbiol. 2010, 138, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Paludan-Müller, C.; Madsen, M.; Sophanodora, P.; Gram, L.; Møller, P.L. Fermentation and microflora of plaa-som, a Thai fermented fish product prepared with different salt concentrations. Int. J. Food Microbiol. 2002, 73, 61–70. [Google Scholar] [CrossRef]
- Srionnual, S.; Yanagida, F.; Lin, L.H.; Hsiao, K.N.; Chen, Y.S. Weissellicin 110, a newly discovered bacteriocin from Weissella cibaria 110, isolated from plaa-som, a fermented fish product from Thailand. Appl. Environ. Microbiol. 2007, 73, 2247–2250. [Google Scholar] [CrossRef] [PubMed]
- Thapa, N.; Pal, J.; Prakash, J. Phenotypic identification and technological properties of lactic acid bacteria isolated from traditionally processed fish products of the Eastern Himalayas. Int. J. Food Microbiol. 2006, 107, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Falck, P.; Shah, N.; Immerzeel, P.; Adlercreutz, P.; Stålbrand, H.; Prajapati, J.B.; Holst, O.; Nordberg Karlsson, E. Evidence for xylooligosaccharide utilization in Weissella strains isolated from Indian fermented foods and vegetables. FEMS Microbiol. Lett. 2013, 346, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Swain, M.R.; Anandharaj, M.; Ray, R.C.; Praveen Rani, R. Fermented fruits and vegetables of Asia: A potential source of probiotics. Biotechnol. Res. Int. 2014, 2014, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, W. Use of starter cultures in fermentation on a household scale. Food Control 1997, 8, 241–258. [Google Scholar] [CrossRef]
- Marsh, A.J.; Hill, C.; Ross, R.P.; Cotter, P.D. Fermented beverages with health-promoting potential: Past and future perspectives. Trends Food Sci. Technol. 2014, 38, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Holzapfel, W.H. Appropriate starter culture technologies for small-scale fermentation in developing countries. Int. J. Food Microbiol. 2002, 75, 197–212. [Google Scholar] [CrossRef]
- Ammor, M.S.; Mayo, B. Selection criteria for lactic acid bacteria to be used as functional starter cultures in dry sausage production: An update. Meat Sci. 2007, 76, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Iyer, R.; Tomar, S.K.; Uma Maheswari, T.; Singh, R. Streptococcus thermophilus strains: Multifunctional lactic acid bacteria. Int. Dairy J. 2010, 20, 133–141. [Google Scholar] [CrossRef]
- Ogier, J.C.; Casalta, E.; Farrokh, C.; Saïhi, A. Safety assessment of dairy microorganisms: The Leuconostoc genus. Int. J. Food Microbiol. 2008, 126, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Altuntas, E.G. Bacteriocins: A natural way to combat with pathogens. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Méndez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2013; pp. 1007–1015. [Google Scholar]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Lactic acid bacteria: Their antimicrobial compounds and their uses in food production. Ann. Biol. Res. 2010, 1, 218–228. [Google Scholar]
- Varsha, K.K.; Nampoothiri, K.M. Appraisal of lactic acid bacteria as protective cultures. Food Control 2016, 69, 61–64. [Google Scholar] [CrossRef]
- Hati, S.; Mandal, S.; Prajapati, J. Novel Starters for Value Added Fermented Dairy Products. Curr. Res. Nutr. Food Sci. J. 2013, 1, 83–91. [Google Scholar] [CrossRef]
- Hammes, W.P.; Knauf, H.J. Starters in the Processing of Meat-Products. Meat Sci. 1994, 36, 155–168. [Google Scholar] [CrossRef]
- Zannini, E.; Waters, D.M.; Coffey, A.; Arendt, E.K. Production, properties, and industrial food application of lactic acid bacteria-derived exopolysaccharides. Appl. Microbiol. Biotechnol. 2016, 100, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Surico, R.F.; Siragusa, S.; De Angelis, M.; Paradiso, A.; Minervini, F.; De Gara, L.; Gobbetti, M. Selection and use of autochthonous mixed starter for lactic acid fermentation of carrots, French beans or marrows. Int. J. Food Microbiol. 2008, 127, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, M.; Fernández, D.; Hernández, T.; Estrella, I.; Muñoz, R. Bioactive phenolic compounds of cowpeas (Vigna sinensis L). Modifications by fermentation with natural microflora and with Lactobacillus plantarum ATCC 14917. J. Sci. Food Agric. 2005, 85, 297–304. [Google Scholar] [CrossRef]
- Filannino, P.; Azzi, L.; Cavoski, I.; Vincentini, O.; Rizzello, C.G.; Gobbetti, M.; Di Cagno, R. Exploitation of the health-promoting and sensory properties of organic pomegranate (Punica granatum L.) juice through lactic acid fermentation. Int. J. Food Microbiol. 2013, 163, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Marazza, J.A.; Nazareno, M.A.; Savoy de Giori, G.; Garro, M.S. Enhancement of the antioxidant capacity of soymilk by fermentation with Lactobacillus rhamnosus. J. Funct. Foods 2012, 4, 594–601. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, A.R. Significance of biogenic amines to food safety and human health. Food Res. Int. 1997, 29, 675–690. [Google Scholar] [CrossRef]
- Hansen, E.B. Commercial bacterial starter cultures for fermented foods of the future. Int. J. Food Microbiol. 2002, 78, 119–131. [Google Scholar] [CrossRef]
- Ahmed, F.E. Genetically modified probiotics in foods. Trends Biotechnol. 2003, 21, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Joerger, M.C.; Klaenhammer, T.R. Cloning, expression, and nucleotide sequence of the Lactobacillus helveticus 481 gene encoding the bacteriocin helveticin J. J. Bacteriol. 1990, 172, 6339–6347. [Google Scholar] [CrossRef] [PubMed]
- Joutsjoki, V.; Luoma, S.; Tamminen, M.; Kilpi, M.; Johansen, E.; Palva, A. Recombinant Lactococcus starters as a potential source of additional peptidolytic activity in cheese ripening. J. Appl. Microbiol. 2002, 92, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, H.; Pronk, J.T.; Teusink, B. Evolutionary engineering to enhance starter culture performance in food fermentations. Curr. Opin. Biotechnol. 2015, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cakar, Z.P.; Turanli-Yildiz, B.; Alkim, C.; Yilmaz, U. Evolutionary engineering of Saccharomyces cerevisiae for improved industrially important properties. FEMS Yeast Res. 2012, 12, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Cakar, Z.P.; Seker, U.O.S.; Tamerler, C.; Sonderegger, M.; Sauer, U. Evolutionary engineering of multiple-stress resistant Saccharomyces cerevisiae. FEMS Yeast Res. 2005, 5, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, H.; Starrenburg, M.J.C.; Molenaar, D.; Kleerebezem, M.; Hylckama Vlieg, J.E.T. Microbial domestication signatures of Lactococcus lactis can be reproduced by experimental evolution. Genome Res. 2012, 22, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Kim, Y.-W.; Hwang, I.; Kim, J.; Yoon, S. Evaluation of Leuconostoc citreum HO12 and Weissella koreensis HO20 isolated from kimchi as a starter culture for whole wheat sourdough. Food Chem. 2012, 134, 2208–2216. [Google Scholar] [CrossRef] [PubMed]
- Robert, H.; Gabriel, V.; Lefebvre, D.; Rabier, P.; Vayssier, Y.; Fontagné-Faucher, C. Study of the behaviour of Lactobacillus plantarum and Leuconostoc starters during a complete wheat sourdough breadmaking process. LWT Food Sci. Technol. 2006, 39, 256–265. [Google Scholar] [CrossRef]
- Wolter, A.; Hager, A.-S.; Zannini, E.; Galle, S.; Gänzle, M.G.; Waters, D.M.; Arendt, E.K. Evaluation of exopolysaccharide producing Weissella cibaria MG1 strain for the production of sourdough from various flours. Food Microbiol. 2014, 37, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Surico, R.F.; Minervini, G.; De Angelis, M.; Rizzello, C.G.; Gobbetti, M. Use of autochthonous starters to ferment red and yellow peppers (Capsicum annum L.) to be stored at room temperature. Int. J. Food Microbiol. 2009, 130, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Galle, S.; Schwab, C.; Arendt, E.; Gänzle, M. Exopolysaccharide-forming Weissella strains as starter cultures for sorghum and wheat sourdoughs. J. Agric. Food Chem. 2010, 58, 5834–5841. [Google Scholar] [CrossRef] [PubMed]
- Malang, S.K.; Maina, N.H.; Schwab, C.; Tenkanen, M.; Lacroix, C. Characterization of exopolysaccharide and ropy capsular polysaccharide formation by Weissella. Food Microbiol. 2015, 46, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Zannini, E.; Mauch, A.; Galle, S.; Gänzle, M.; Coffey, A.; Arendt, E.K.; Taylor, J.P.; Waters, D.M. Barley malt wort fermentation by exopolysaccharide-forming Weissella cibaria MG1 for the production of a novel beverage. J. Appl. Microbiol. 2013, 115, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.Z.; Siddiqui, K.; Arman, M.; Ahmed, N. Characterization of high molecular weight dextran produced by Weissella cibaria CMGDEX3. Carbohydr. Polym. 2012, 90, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Rao, T.J.M.; Goyal, A. A novel high dextran yielding Weissella cibaria JAG8 for cereal food application. Int. J. Food Sci. Nutr. 2013, 64, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Tingirikari, J.M.R.; Kothari, D.; Shukla, R.; Goyal, A. Structural and biocompatibility properties of dextran from Weissella cibaria JAG8 as food additive. Int. J. Food Sci. Nutr. 2014, 65, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Maina, N.H.; Tenkanen, M.; Maaheimo, H.; Juvonen, R.; Virkki, L. NMR spectroscopic analysis of exopolysaccharides produced by Leuconostoc citreum and Weissella confusa. Carbohydr. Res. 2008, 343, 1446–1455. [Google Scholar] [CrossRef] [PubMed]
- Juvonen, R.; Honkapää, K.; Maina, N.H.; Shi, Q.; Viljanen, K.; Maaheimo, H.; Virkki, L.; Tenkanen, M.; Lantto, R. The impact of fermentation with exopolysaccharide producing lactic acid bacteria on rheological, chemical and sensory properties of pureed carrots (Daucus carota L.). Int. J. Food Microbiol. 2015, 207, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.; Ha, M.; Bae, O.; Lee, Y. Effect of Weissella confusa strain PL9001 on the adherence and growth of Helicobacter pylori. Appl. Environ. Microbiol. 2002, 68, 4642–4645. [Google Scholar] [CrossRef] [PubMed]
- Leong, K.; Chen, Y.; Lin, Y.; Pan, S.; Yu, B.; Wu, H.; Yanagida, F. Weissellicin L, a novel bacteriocin from sian-sianzih-isolated Weissella hellenica 4–7. J. Appl. Microbiol. 2013, 115, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Zendo, T.; Sawa, N.; Perez, R.H.; Nakayama, J.; Sonomoto, K. Characterization and identification of weissellicin Y and weissellicin M, novel bacteriocins produced by Weissella cibaria QU 13. J. Appl. Microbiol. 2012, 112, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Papagianni, M.; Papamichael, E.M. Purification, amino acid sequence and characterization of the class IIa bacteriocin weissellin A, produced by Weissella paramesenteroides DX. Bioresour. Technol. 2011, 102, 6730–6734. [Google Scholar] [CrossRef] [PubMed]
- Woraprayote, W.; Pumpuang, L.; Tosukhowong, A.; Roytrakul, S.; Perez, R.H.; Zendo, T.; Sonomoto, K.; Benjakul, S.; Visessanguan, W. Two putatively novel bacteriocins active against Gram-negative food borne pathogens produced by Weissella cibaria BCC 7293. Food Control 2015, 55, 176–184. [Google Scholar] [CrossRef]
- Malik, A.; Sumayyah, S.; Yeh, C.W.; Heng, N.C.K. Identification and sequence analysis of pWcMBF8-1, a bacteriocin-encoding plasmid from the lactic acid bacterium Weissella confusa. FEMS Microbiol. Lett. 2016, 363, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.-Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Park, J.Y.; Jeong, H.R.; Heo, H.J.; Han, N.S.; Kim, J.H. Probiotic properties of Weissella strains isolated from human faeces. Anaerobe 2012, 18, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, C.; Jänsch, A.; Ehrmann, M.A.; Toelstede, S.; Vogel, R.F. Characterization of Cinnamoyl Esterases from Different Lactobacilli and Bifidobacteria. Curr. Microbiol. 2017, 74, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, N.; Esteban-Torres, M.; Mancheño, J.M.; de Las Rivas, B.; Muñoz, R. Tannin degradation by a novel tannase enzyme present in some Lactobacillus plantarum strains. Appl. Environ. Microbiol. 2014, 80, 2991–2997. [Google Scholar] [CrossRef] [PubMed]
- Pyo, Y.-H.; Lee, T.-C.; Lee, Y.-C. Enrichment of bioactive isoflavones in soymilk fermented with β-glucosidase-producing lactic acid bacteria. Food Res. Int. 2005, 38, 551–559. [Google Scholar] [CrossRef]
- Rodriguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; Lopez de Felipe, F.; Gomez-Cordoves, C.; Mancheno, J.M.; Munoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, H.; Landete, J.M.; de las Rivas, B.; Muñoz, R. Metabolism of food phenolic acids by Lactobacillus plantarum CECT 748T. Food Chem. 2008, 107, 1393–1398. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.W.; Han, N.S.; Kim, J.H. Purification and characterization of beta-glucosidase from Weissella cibaria 37. J. Microbiol. Biotechnol. 2012, 22, 1705–1713. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Kim, G.M.; Lee, K.W.; Choi, I.D.; Kwon, G.H.; Park, J.Y.; Jeong, S.J.; Kim, J.S.; Kim, J.H. Conversion of isoflavone glucosides to aglycones in soymilk by fermentation with lactic acid bacteria. J. Food Sci. 2007, 72, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Kim, J.S.; Kim, J.H. Enrichment of isoflavone aglycones in soymilk by fermentation with single and mixed cultures of Streptococcus infantarius 12 and Weissella sp. 4. Food Chem. 2008, 109, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Gobbetti, M.; De Angelis, M.; Di Cagno, R. Hydroxycinnamic acids used as external acceptors of electrons: An energetic advantage for strictly heterofermentative lactic acid bacteria. Appl. Environ. Microbiol. 2014, 80, 7574–7582. [Google Scholar] [CrossRef] [PubMed]
- Park, J.A.; Tirupathi Pichiah, P.B.; Yu, J.J.; Oh, S.H.; Daily, J.W.; Cha, Y.S. Anti-obesity effect of kimchi fermented with Weissella koreensis OK1-6 as starter in high-fat diet-induced obese C57BL/6J mice. J. Appl. Microbiol. 2012, 113, 1507–1516. [Google Scholar] [CrossRef] [PubMed]
- Spano, G.; Russo, P.; Lonvaud-Funel, A.; Lucas, P.; Alexandre, H.; Grandvalet, C.; Coton, E.; Coton, M.; Barnavon, L.; Bach, B.; et al. Biogenic amines in fermented foods. Eur. J. Clin. Nutr. 2010, 64, S95–S100. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-atienza, E.; Landeta, G.; De, B.; Gómez-sala, B.; Muñoz, R.; Hernández, P.E.; Cintas, L.M.; Herranz, C. Phenotypic and genetic evaluations of biogenic amine production by lactic acid bacteria isolated from fish and fish products. Int. J. Food Microbiol. 2011, 146, 212–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, D.-W.; Lee, J.-H. Antibiotic resistance, hemolysis and biogenic amine production assessments of Leuconostoc and Weissella isolates for kimchi starter development. LWT Food Sci. Technol. 2015, 64, 1078–1084. [Google Scholar] [CrossRef]
- Arena, M.E.; Manca De Nadra, M.C. Biogenic amine production by Lactobacillus. J. Appl. Microbiol. 2001, 90, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Costantini, A.; Pietroniro, R.; Doria, F.; Pessione, E.; Garcia-Moruno, E. Putrescine production from different amino acid precursors by lactic acid bacteria from wine and cider. Int. J. Food Microbiol. 2013, 165, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Dapkevicius, M.L.N.E.; Nout, M.J.R.; Rombouts, F.M.; Houben, J.H.; Wymenga, W. Biogenic amine formation and degradation by potential fish silage starter microorganisms. Int. J. Food Microbiol. 2000, 57, 107–114. [Google Scholar] [CrossRef]
- Mathur, S.; Singh, R. Antibiotic resistance in food lactic acid bacteria—A review. Int. J. Food Microbiol. 2005, 105, 281–295. [Google Scholar] [CrossRef] [PubMed]
- D’Aimmo, M.R.; Modesto, M.; Biavati, B. Antibiotic resistance of lactic acid bacteria and Bifidobacterium spp. isolated from dairy and pharmaceutical products. Int. J. Food Microbiol. 2007, 115, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Tomar, S.K.; Goswami, P.; Sangwan, V.; Singh, R. Antibiotic resistance among commercially available probiotics. Food Res. Int. 2014, 57, 176–195. [Google Scholar] [CrossRef]
- Jin, L.; Tao, L.; Pavlova, S.I.; So, J.S.; Kiwanuka, N.; Namukwaya, Z.; Saberbein, B.A.; Wawer, M. Species diversity and relative abundance of vaginal lactic acid bacteria from women in Uganda and Korea. J. Appl. Microbiol. 2007, 102, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Silvester, M.E.; Dicks, L.M.T. Identification of lactic acid bacteria isolated from human vaginal secretions. Antonie Leeuwenhoek 2003, 83, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y. Characterization of Weissella kimchii PL9023 as a potential probiotic for women. FEMS Microbiol. Lett. 2005, 250, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Gomathi, S.; Sasikumar, P.; Anbazhagan, K.; Sasikumar, S.; Kavitha, M.; Selvi, M.S.; Selvam, G.S. Screening of indigenous oxalate degrading lactic acid bacteria from human faeces and south Indian fermented foods: Assessment of probiotic potential. Sci. World J. 2014, 2014, 648059. [Google Scholar] [CrossRef] [PubMed]
- Nistal, E.; Caminero, A.; Vivas, S.; Ruiz De Morales, J.M.; Sáenz De Miera, L.E.; Rodríguez-Aparicio, L.B.; Casqueiro, J. Differences in faecal bacteria populations and faecal bacteria metabolism in healthy adults and celiac disease patients. Biochimie 2012, 94, 1724–1729. [Google Scholar] [CrossRef] [PubMed]
- Rubio, R.; Jofré, A.; Martín, B.; Aymerich, T.; Garriga, M. Characterization of lactic acid bacteria isolated from infant faeces as potential probiotic starter cultures for fermented sausages. Food Microbiol. 2014, 38, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.; Hertel, C.; Tannock, G.W.; Lis, C.M.; Munro, K.; Hammes, W.P. Detection of Lactobacillus, Pediococcus, Leuconostoc, and Weissella Species in Human Feces by Using Group-Specific PCR Primers and Denaturing Gradient Gel Electrophoresis. Appl. Environ. Microbiol. 2001, 67, 2578–2585. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.A.A.; Leal, C.A.G.; Schuenker, N.D.; Leite, R.C.; Figueiredo, H.C.P. Characterization of Weissella ceti infections in Brazilian rainbow trout, Oncorhynchus mykiss (Walbaum), farms and development of an oil-adjuvanted vaccine. J. Fish Dis. 2015, 38, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Snyder, A.K.; Hinshaw, J.M.; Welch, T.J. Diagnostic tools for rapid detection and quantification of Weissella ceti NC36 infections in rainbow trout. Lett. Appl. Microbiol. 2015, 60, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Lerma, L.L.; Casado Muñoz, M.D.C.; Montoro, B.P.; Kabisch, J.; Pichner, R.; Cho, G.-S.; Neve, H.; Fusco, V.; Franz, C.M.A.P.; et al. The controversial nature of the Weissella genus: Technological and functional aspects versus whole genome analysis-based pathogenic potential for their application in food and health. Front. Microbiol. 2015, 6, 1197. [Google Scholar] [CrossRef] [PubMed]
- Fairfax, M.R.; Lephart, P.R.; Salimnia, H. Weissella confusa: Problems with identification of an opportunistic pathogen that has been found in fermented foods and proposed as a probiotic. Front. Microbiol. 2014, 5, 254. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, J.D.; Levett, P.N.; Dewhirst, F.E.; Troe, T.E.; Warren, J.R.; Johnson, S. Fatal case of endocarditis due to Weissella confusa. J. Clin. Microbiol. 2003, 41, 2237–2239. [Google Scholar] [CrossRef] [PubMed]
- Harlan, N.P.; Kempker, R.R.; Parekh, S.M.; Burd, E.M.; Kuhar, D.T. Weissella confusa bacteremia in a liver transplant patient with hepatic artery thrombosis. Transpl. Infect. Dis. 2011, 13, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Augustine, D.; Sudhindran, S.; Kurian, A.M.; Dinesh, K.R.; Karim, S.; Philip, R. Weissella confusa: A rare cause of vancomycinresistant Gram-positive bacteraemia. J. Med. Microbiol. 2011, 60, 1539–1541. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Huang, Y.T.; Liao, C.H.; Lai, C.C.; Lee, P.I.; Hsueh, P.R. Bacteraemia caused by Weissella confusa at a university hospital in Taiwan, 1997–2007. Clin. Microbiol. Infect. 2011, 17, 1226–1231. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Cho, S.M.; Kim, M.; Ko, Y.G.; Yong, D.; Lee, K. Weissella confusa bacteremia in an immune-competent patient with underlying intramural hematomas of the aorta. Ann. Lab. Med. 2013, 33, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Medford, R.; Patel, S.N.; Evans, G.A. A confusing case—Weissella confusa prosthetic joint infection: A case report and review of the literature. Can. J. Infect. Dis. Med. Microbiol. 2014, 25, 173–175. [Google Scholar] [CrossRef] [PubMed]
- Olano, A.; Chua, J.; Schroeder, S.; Minari, A.; Salvia, M.L.A.; Hall, G. Weissella confusa (Basonym: Lactobacillus confusus) Bacteremia: A Case Report. J. Clin. Microbiol. 2001, 39, 1604–1607. [Google Scholar] [CrossRef] [PubMed]
- Salimnia, H.; Alangaden, G.J.; Bharadwaj, R.; Painter, T.M.; Chandrasekar, P.H.; Fairfax, M.R. Weissella confusa: An unexpected cause of vancomycin-resistant gram-positive bacteremia in immunocompromised hosts. Transpl. Infect. Dis. 2011, 13, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Kim, D.I.L.; Kim, H.R.; Kim, D.S.; Kook, J.K.; Lee, J.N. Severe infective endocarditis of native valves caused by Weissella confusa detected incidentally on echocardiography. J. Infect. 2007, 54, 149–151. [Google Scholar] [CrossRef] [PubMed]
- Svec, P.; Sevcíková, A.; Sedlácek, I.; Bednárová, J.; Snauwaert, C.; Lefebvre, K.; Vandamme, P.; Vancanneyt, M. Identification of lactic acid bacteria isolated from human blood cultures. FEMS Immunol. Med. Microbiol. 2007, 49, 192–196. [Google Scholar] [CrossRef]
- Kamboj, K.; Vasquez, A.; Balada-Llasat, J.M. Identification and significance of Weissella species infections. Front. Microbiol. 2015, 6, 1204. [Google Scholar] [CrossRef] [PubMed]
- Cannon, J.P.; Lee, T.A.; Bolanos, J.T.; Danziger, L.H. Pathogenic relevance of Lactobacillus: A retrospective review of over 200 cases. Eur. J. Clin. Microbiol. Infect. Dis. 2005, 24, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Franko, B.; Fournier, P.; Jouve, T.; Malvezzi, P.; Pelloux, I.; Brion, J.P.; Pavese, P. Lactobacillus bacteremia: Pathogen or prognostic marker? Med. Mal. Infect. 2017, 47, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Garneau, J.E.; Moineau, S. Bacteriophages of lactic acid bacteria and their impact on milk fermentations. Microb. Cell Factories 2011, 10, S20. [Google Scholar] [CrossRef] [PubMed]
- Kot, W.; Neve, H.; Heller, K.J.; Vogensen, F.K. Bacteriophages of Leuconostoc, Oenococcus, and Weissella. Front. Microbiol. 2014, 5, 186. [Google Scholar] [CrossRef] [PubMed]
- Kleppen, H.P.; Holo, H.; Jeon, S.R.; Nes, I.F.; Yoon, S.S. Novel Podoviridae family bacteriophage infecting Weissella cibaria isolated from kimchi. Appl. Environ. Microbiol. 2012, 78, 7299–7308. [Google Scholar] [CrossRef] [PubMed]
- Mahony, J.; Ainsworth, S.; Stockdale, S.; van Sinderen, D. Phages of lactic acid bacteria: The role of genetics in understanding phage-host interactions and their co-evolutionary processes. Virology 2012, 434, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Klaenhammer, T.R.; Kullen, M.J. Selection and design of probiotics. Int. J. Food Microbiol. 1999, 50, 45–57. [Google Scholar] [CrossRef]
- Bourdichon, F.; Casaregola, S.; Farrokh, C.; Frisvad, J.C.; Gerds, M.L.; Hammes, W.P.; Harnett, J.; Huys, G.; Laulund, S.; Ouwehand, A.; et al. Food fermentations: Microorganisms with technological beneficial use. Int. J. Food Microbiol. 2012, 154, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Selection of Yeasts as Starter Cultures for Table Olives: A Step-by-Step Procedure. Front. Microbiol. 2012, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Neighbour-joining phylogenetic tree based on 16S rRNA gene sequences of Weissella and the related species Leuconostoc mesenteroides obtained from Clustal W2 Multiple Sequence Alignment. The relative genetic distances are indicated by the scale bar and the branch lengths. Weissella species Accession numbers: W. beninensis 2L24P13; EU439435. W. fabaria 257; FM179678. W. ghanensis LMG 24286; AM882997. W. fabalis M75; HE576795. W. halotolerans NRIC 1627; AB022926. W. ceti 1999-1A-09; FN813251. W. viridescens NRIC 1536; AB023236. W. minor NRIC 1625; NR_040809. W. uvarum B18NM42; KF999666. W. diestrammenae ORY33; JQ646523. W. kandleri NRIC 1628; AB022922. W. koreensis S-5623; AY035891. W. cibaria LMG 17699; AJ295989. W. confusa JCM 1093; AB023241. W. thailandensis FS61-T; AB023838. W. jogaejeotgali FOL01; NR_145896. W. paramesenteroides NRIC 1542; AB023238. W. bombi LMG 28290; LK054487. W. hellenica NCFB 2973; X95981. W. soli LMG 20113; AY028260. W. oryzae SG25; AB690345.
Figure 1.
Neighbour-joining phylogenetic tree based on 16S rRNA gene sequences of Weissella and the related species Leuconostoc mesenteroides obtained from Clustal W2 Multiple Sequence Alignment. The relative genetic distances are indicated by the scale bar and the branch lengths. Weissella species Accession numbers: W. beninensis 2L24P13; EU439435. W. fabaria 257; FM179678. W. ghanensis LMG 24286; AM882997. W. fabalis M75; HE576795. W. halotolerans NRIC 1627; AB022926. W. ceti 1999-1A-09; FN813251. W. viridescens NRIC 1536; AB023236. W. minor NRIC 1625; NR_040809. W. uvarum B18NM42; KF999666. W. diestrammenae ORY33; JQ646523. W. kandleri NRIC 1628; AB022922. W. koreensis S-5623; AY035891. W. cibaria LMG 17699; AJ295989. W. confusa JCM 1093; AB023241. W. thailandensis FS61-T; AB023838. W. jogaejeotgali FOL01; NR_145896. W. paramesenteroides NRIC 1542; AB023238. W. bombi LMG 28290; LK054487. W. hellenica NCFB 2973; X95981. W. soli LMG 20113; AY028260. W. oryzae SG25; AB690345.
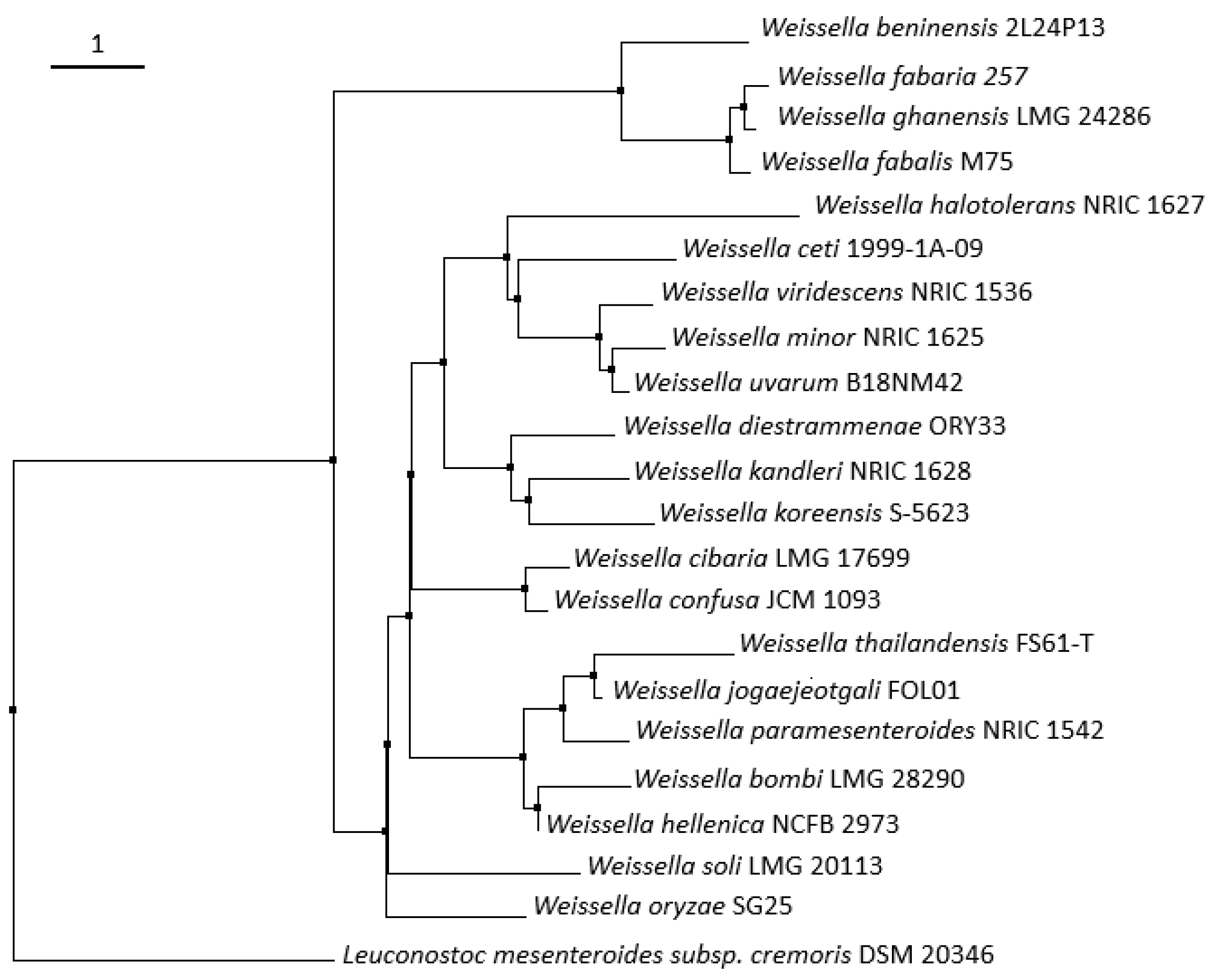
Figure 2.
Flow-sheet for the selection of a starter [216]. Dotted line is for facultative paths.
Figure 2.
Flow-sheet for the selection of a starter [216]. Dotted line is for facultative paths.
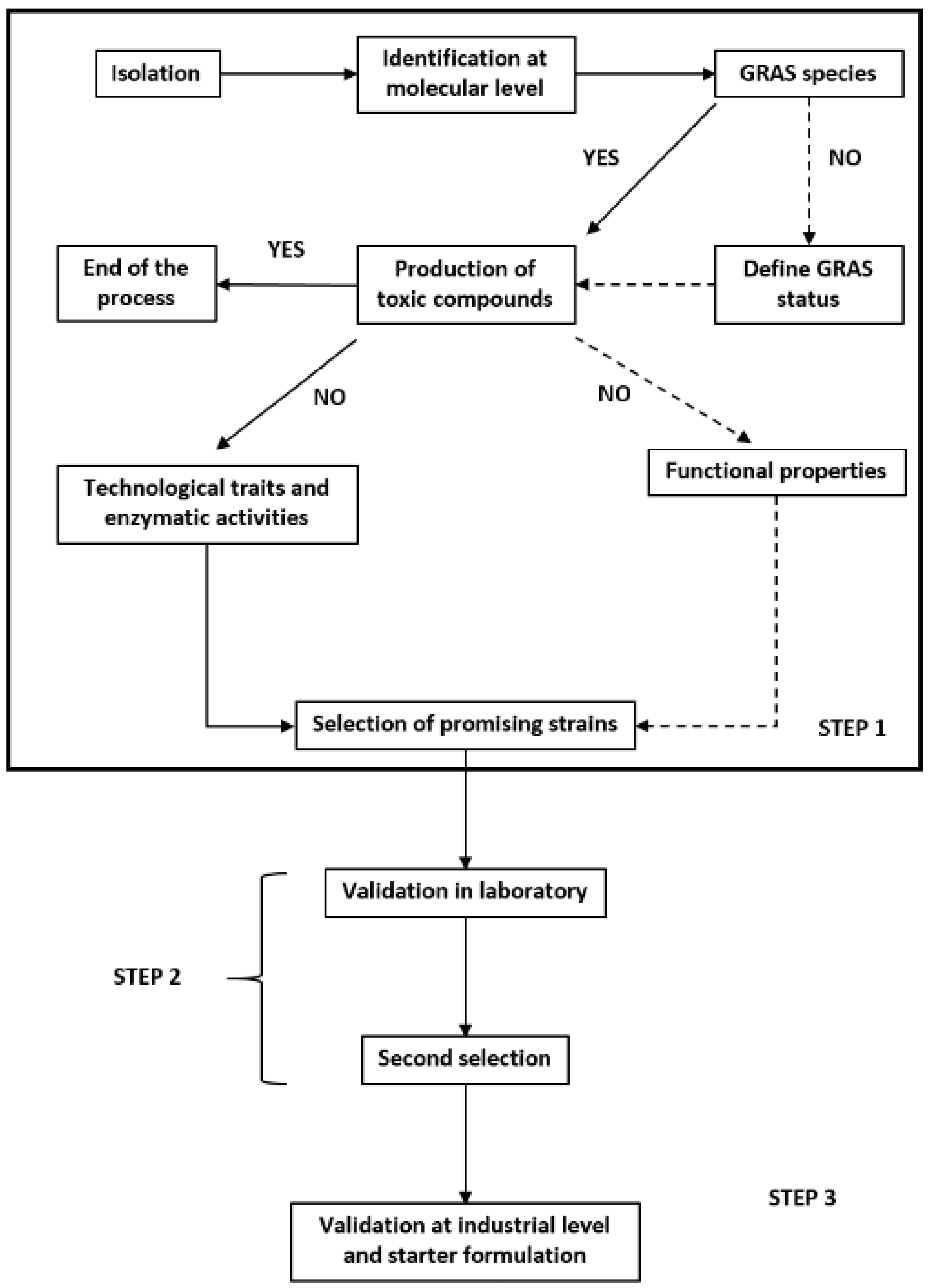

{kind=link}
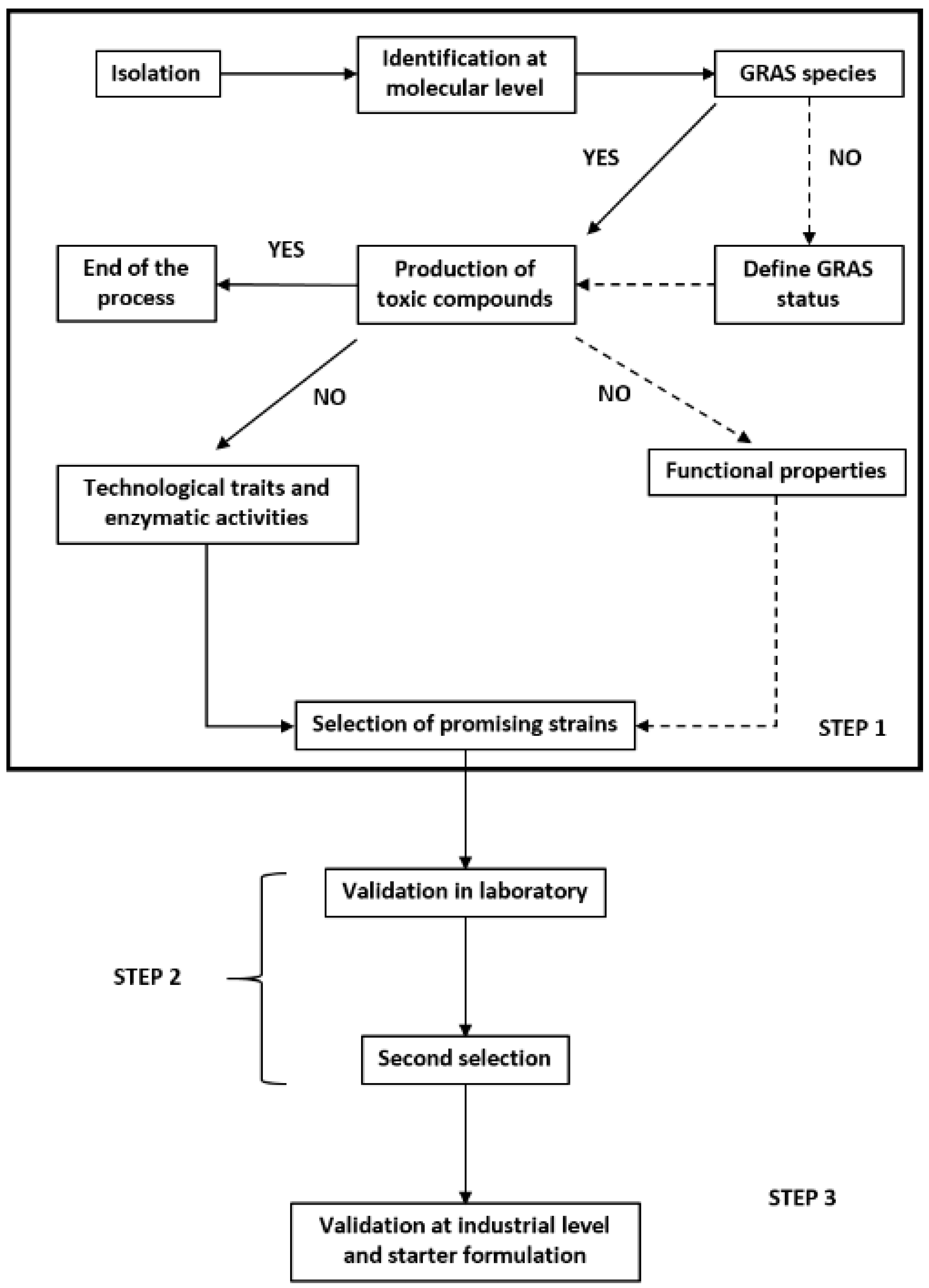{kind=link}
Table 1.
Weissella species, sources, taxonomic branch and genome size.
| Weissella spp. | Sources | Genome Size (Mb) | Taxonomic Branch | References |
|---|---|---|---|---|
| W. fabalis | Cocoa bean fermentation | - | 1 | [26] |
| W. fabaria | Cocoa bean fermentation | - | 1 | [27] |
| W. ghanensis | Cocoa bean fermentation | - | 1 | [28] |
| W. beninensis | Fermented cassava | - | 1 | [16] |
| W. jogaejeotgali | Jeotgal (fermented seafood) | 2.11 (CP014332.1) | 2 | [29] |
| W. thailandensis | Jeotgal, Mexican cheese, Thai fermented meat and fish | 1.97 (HE575133 to HE575182) | 2 | [30]; See Table 2 |
| W. paramesenteroides | Fermented sausages, fermented fruits and vegetables, fermented milk, cheese, fermented cassava, sourdough, fermented cereal, fermented fish | 1.96 (NZ_ACKU00000000.1) | 2 | [31,32]; See Table 2 |
| W. hellenica | Fermented sausages, fermented vegetables, fermented milk, cheese, fermented maize beverage, fermented meat | 1.82–1.92 (NZ_BBIK00000000.1; NZ_FMAW00000000.1) | 2 | [19]; See Table 2 |
| W. bombi | Gut of a bumble bee | - | 2 | [33] |
| W. halotolerans | Meat products, fermented sea food, cheese, fermented sausages | 1.36 (NZ_ATUU00000000.1) | 3 | [20]; See Table 2 |
| W. ceti | Beaked whale | 1.35–1.39 (NZ_ANCA00000000.1; NZ_CP009224.1) | 3 | [17] |
| W. viridescens | Meat products, dry salami, kimchi, fermented milk, cheese, fermented cereal beverage, fermented sausages, fermented fish | 1.53–1.56 (NZ_JQBM00000000.1; NZ_CYXF00000000.1) | 3 | [24]; See Table 2 |
| W. minor | Sludge of milking machine, fermented fruits and vegetables, fermented dry salami | 1.77 (NZ_JQCD00000000.1) | 3 | [20]; See Table 2 |
| W. uvarum | Wine grapes | - | 3 | [25] |
| W. diestrammenae | Gut of a camel cricket | - | 4 | [34] |
| W. kandleri | Desert spring and desert plants | 1.33 (NZ_JQBP00000000.1) | 4 | [35] |
| W. koreensis | Kimchi, fermented vegetables | 1.42–1.73 (NC_015759.1; NZ_AKGG00000000.1) | 4 | [36]; See Table 2 |
| W. cibaria | Malaysian ingredients foods, clinical samples, animals, human feces, fermented fruits and vegetables, fermented dairy products, cheese, fermented cassava, fermented cereal foods and beverages, sourdough, cocoa bean fermentation, fermented sausages, fermented fish | 2.32–2.47 (NZ_AEKT00000000.1; NZ_CP012873.1) | 5 | [37]; See Table 2 |
| W. confusa | Sugar cane, animal, clinical samples, Malaysian ingredients foods, human feces, pasta manufacturing, kimchi, fermented vegetables, fermented milk, cheese, fermented cereal foods, fermented cassava, sourdough, cocoa bean fermentation, pork sausages, fermented fish | 2.18–2.28 (NZ_MNBZ00000000.1; NZ_CAGH00000000.1) | 5 | [37,38]; See Table 2 |
| W. oryzae | Fermented rice grains, cereal based beverage | 2.13 (NZ_BAWR00000000.1) | 0 | [34,39] |
| W. soli | Soil, kimchi, Jeotgal, stinky tofu, leek fermentation, sliced cabbage | - | 0 | [40]; See Table 2; [31] |
Table 2.
Detection of Weissella spp. in fermented foods.
| Food Category | Fermented Product | Weissella Species | Reference |
|---|---|---|---|
| Fruits and Vegetables Fermented Food | Kimchi (mix vegetables) | W. koreensis, W. cibaria, W. confusa, W. soli, W. hellenica | [36,41,42,43,44,45,46] |
| Yan-dong-gua (wax gourd) | W. cibaria, W. paramesenteroides | [47] | |
| Pobuzihi (cummingcordia) | W. cibaria, W. paramesenteroides | [48] | |
| Jiang-gua (cucumbers) | W. cibaria, W. hellenica | [49] | |
| Yan-tsai-shin (brocoli stems) | W. cibaria, W. paramesenteroides, W. minor | [50] | |
| Yan-Jiang (ginger) | W. cibaria | [51] | |
| Yan-taozih (pickled peaches) | W. cibaria, W. paramesenteroides, W. minor | [52] | |
| Xi-gua-mian (watermelon) | W. paramesenteroides | [53] | |
| Dochi (black beans) | W. paramesenteroides | [54] | |
| Koozh (cucumber) | W. koreensis | [55] | |
| Suan-tsai, fu-tsai (mustard product) | W. cibaria, W. paramesenteroides | [56] | |
| Jeotgal (sea food) | W. thailandensis, W. halotolerans, W. soli, W. cibaria, W. jogaejeotgali | [29,43] | |
| Stinky tofu | W. cibaria, W. confusa, W. paramesenteroides, W. soli | [57] | |
| Tuaw jaew (soybeans) | W. confusa | [58] | |
| Fermented sea food | W. paramesenteroides | [59] | |
| Fermented cabbage | W. cibaria | [60] | |
| Tempoyak (durian) | W. paramesenteroides | [61] | |
| Commercial cucumber fermentation | W. cibaria, W. paramesenteroides | [62] | |
| Cauliflower and mixed vegetables | W. cibaria, W. paramesenteroides | [63] | |
| Well-fermented leek kimchi | W. viridescens, W. confusa, W. cibaria | [64] | |
| Leek fermentation | W. soli, Weissella spp. | [65] | |
| Commercial sauerkraut | Weissella spp. | [66] | |
| Sauerkraut | Weissella confusa/cibaria | [67] | |
| Dairy Fermented Food | Nono (Fermented skimmed milk), cheese | W. confusa | [68] |
| Nunu (Ghanaian fermented milk) | W. confusa | [69] | |
| Kule naoto (Masain fermented milk in Kenya) | W. paramesenteroides/Lc mesenteroides | [70] | |
| Maisian fermented zebu milk | W. confusa (Lb confusus), W. viridescens (Lb viridescens) | [71] | |
| Mongolian fermented milk | W. viridescens | [72] | |
| Shubat (fermented camel milk) | W. hellenica | [73] | |
| Dahi (Indian yogurt like product) | W. cibaria | [60] | |
| Italian cheese | W. confusa, W. cibaria | [74] | |
| Mexican cheese (Cotija cheese) | W. thailandensis | [75] | |
| Manura (hard cheese) | W. paramesenteroides, W. viridescens | [76] | |
| Scamorza Altamurana Cheese | W. viridescens | [77] | |
| Mozarella cheese | W. hellenica | [78] | |
| Cheddar cheese | W. halotolerans, W. viridescens | [79] | |
| Romanian cheese | W. viridescens | [80] | |
| Mongolian dairy products | W. cibaria | [81] | |
| Starchy or Cereal-Based Fermented Food and Diverse | Togwa (sorghum based food) | W. confusa | [82] |
| Gari, Attieke, Lafun (cassava) | W. paramesenteroides, W. cibaria, W. confusa, W. beninensis | [16,83,84,85,86] | |
| Idli batter (Indian fermented rice and black gram based food) | W. confusa, W. cibaria | [60,87] | |
| French wheat sourdough, buckwheat and teff sourdough, Italian sourdough, Turkish sourdough, spontaneous sourdough | W. confusa, W. cibaria, W. paramesenteroides | [10,88,89,90,91,92,93] | |
| Mexican Pozol | W. paramesenteroides | [94] | |
| Fermented rice grains | W. oryzae | [95] | |
| Chicha (maize based beverage) | W. confusa, W. hellenica, W. paramesenteroides | [96] | |
| Kunu-zaki (Nigerian cereal based food) | W. confusa | [97] | |
| Boza (Bulgarian cereal-based beverage) | W. confusa, W. oryzae | [39] | |
| Fura (African millet based food) | W. confusa | [98] | |
| Borde (Ethiopian cereal beverage) | W. confusa, W. viridescens | [99] | |
| Cocoa bean fermentation | W. cibaria, W. ghanensis, W. confusa, W. paramesenteroides, W. fabaria, W. fabalis | [26,27,28,100] | |
| Makgeolli (Korean starchy alcoholic beverage) | W. confusa, W. cibaria, W. paramesenteroides | [101] | |
| Fermented Meat and Fish Product | Nham (Thai fermented pork sausage) | W. cibaria, W. confusa | [102,103,104] |
| Italian fermented sausages | W. hellinica, W. paramesenteroides | [105,106] | |
| Greek dry-fermented sausages | W. viridescens | [107] | |
| Fermented sausages in Hungary | W. paramesenteroides/W. hellenica, W. viridescens | [108] | |
| Alheira (Fermented sausage in Portugal) | W. cibaria, W. viridescens | [109] | |
| Sucuk (fermented turkish sausages) | W. viridescens | [110] | |
| Portuguese fermented sausage | W. halotolerans | [111] | |
| Smoked horsemeat sausage | W. hellenica, Weissella spp. | [112] | |
| Fermented sausages | W. hellenica | [19] | |
| Fermented Greek dry salami | W. hellenica, W. viridescens, W. paramesenteroides, W. minor, W. halotolerans | [113] | |
| Chinese dong fermented meat | W. hellenica | [114] | |
| Mum (Thai fermented meat) | W. thailandensis | [58] | |
| Pla-ra, Pla-ra sub, plaa-som, pla-jom, pla-jaw (Thai fermented fish) | W. thailandensis, W. cibaria, W. confusa, W. paramesenteroides, W. viridescens | [30,58,115,116,117] | |
| Sidra (Fish products) | W. confusa | [118] |
Table 3.
Lactic acid bacteria used as commercial starters in the food industry.
| Type of Fermented Product | Product | LAB Species Used as Commercial Starters | References |
|---|---|---|---|
| Dairy Fermented Foods | Yoghurt | S. thermophilus, Lb. delbruecki subsp. bulgaricus | [130] |
| Fermented/Probiotic milk | Lb. casei, Lb. acidophilus, Lb. rhamnosus, Lb. johnsonii, B. lactis, B. bifidum, B. brevis | ||
| Kefir | Lb. kefir, Lb. kefiranofacies, Lb. brevis | ||
| Butter and buttermilk | L. lactis susbp. lactis, L. lactis subsp. diacetylactis, L. lactis subsp. cremoris, Lc. mesenteroides subsp. cremoris | ||
| Swiss and Italian type cheeses | Lb. delbrueckii subsp. lactis, Lb. helveticus, Lb. casei, Lb. delbrueckii subsp. bulgaricus, S. thermophilus | ||
| Cheeses (with or without eyes) | L. lactis subsp. Lactis, L. lactis subsp. cremoris, L. lactis subsp. diacetylactis, Lc. mesenteroides subsp. cremoris | ||
| Fermented Cereals | Sourdough | Lb. brevis, Lb. plantarum, Lb. sanfranciscensis, Lb. casei, Lb. delbrueckii, Lb. fermentum, P. pentosaceus, P. acidilactic, Lb. pontis, Lb. crispatus, Lb. paracasei, Lb. helveticus, Lb. paralimentarius, Lc. lastis | [6] |
| Fruits and Vegetables | Sauerkraut | Lc. mesenteroides, Lb. plantarum, P. acidilactici | [8] |
| Pickles | Lc. mesenteroides, P. cerevisiae, Lb. brevis, Lb. plantarum | ||
| Fermented olives | Lb. paracasei, Lb. pentosus, Lb. plantarum | ||
| Fermented vegetables | P. acidilactici, P. pentosaceus, Lb. plantarum, Lb. fermentum | ||
| Vegetable juices | Lb. acidophilus, Lb. bavaricus, Lb. bifidus, Lb. brevis, Lb. casei, Lb. delbrueckii, Lb. helveticus, Lb. plantarum, Lb. salivarius, Lb. xylosus, L. lactis, Lc. mesenteroides | [3] | |
| Meat Products | Sausages | P. acidilactici, P. pentosaceus, Lb. sakei, Lb. curvatus, Lb. plantarum, Lb. pentosus, Lb, casei, L. lactis | [124,131] |
| Fermented Beverages | Wine | Oenococcus oeni, Lb. plantarum, Lb. hilgardii | [3] |
Table 4.
Criteria for the selection of starters for fruits and vegetables, cereal, dairy and meat fermentation.
Table 4.
Criteria for the selection of starters for fruits and vegetables, cereal, dairy and meat fermentation.
| Criteria Category | Fruits and Vegetables | Dairy Products | Meat Products | Cereal Based Foods |
|---|---|---|---|---|
| Technological | - Growth and acidification rate - Salt tolerance - Tolerance to low values of pH - Growth at low temperature - Completeness of fermentation - Malolatic fermentation - Tolerance to phenols - Synthesis of antimicrobial substances - No formation of hydrogen peroxide - Pectinolytic activity | - Growth and acidification rate - Production of nutraceuticals - Accelerate ripening of cheese - Resistance to bacteriophage - Proteinase and peptidase activity | - Fast production of lactic acid - Growth rate at different temperatures - Salt and pH tolerance - Persistence over the whole fermentation and ripening process - Nitrate and nitrite reduction - Catalase positive - Lactose negative - Proteolytic and lipolytic enzyme activities - No formation of hydrogen peroxide - Antagonism against pathogens - Improve the nutritional value of the sausages | - Growth and acidification rate - Salt tolerance - Growth at low temperatures - Synthesis of antimicrobial compounds |
| Sensory | - Hetero-fermentative metabolism - Synthesis of aroma compounds and their precursors | - Production of aroma and flavor - Synthesis of exopolysaccharides | - Formation of flavor - Hetero-fermentative metabolism | - Hetero-fermentative metabolism - Synthesis of aroma compounds and their precursors |
| Nutritional | - Synthesis of exopolysaccharides - Increase of the antioxidant activity - Synthesis of biogenic compounds - Bacteriocin production | - Synthesis of exo-polysaccharides - Bacteriocins production - Reduction of toxic or antinutritional factors - Low-calorie sugar production - Vitamin production - Bioactive peptide production - Production of conjugated linolein acid (CLA) | - Tolerance or even synergy to other microbial components or starters - No formation of ropy slime - Bacteriocin production - Probiotic features | - Release of free amino acids - Synthesis of biogenic compounds - Degradation of antinutritionnal factor (phytic acid) - Increase the antioxidant activity - Synthesis of exopolysaccharides |
| Safety | - No formation of biogenic amines - No antibiotic resistance profile | |||
Table 5.
Characterization of EPS from Weissella species.
| Species | Source | Type of EPS | Molecular Mass (Da) | Amount Produced | Media | Reference |
|---|---|---|---|---|---|---|
| W. cibaria (WC4) | Sourdough | Glucan | 1.1–1.3 × 104 | 7.9 g·L−1 (6 d) | MRS + Sucrose | [10] |
| 2.5 g·kg−1 | Sourdough | |||||
| W. cibaria (WC3) | Sourdough | Glucan | 1.1–1.3 × 104 | 6.7 g·L−1 (6 d) | MRS + Sucrose | [10] |
| W. cibaria (WC9) | Sourdough | Glucan | 1.1–1.3 × 104 | 5.5 g·L−1 (6 d) | MRS + Sucrose | [10] |
| W. cibaria (MG1) | Sourdough | Dextran | 106 to 107 | 0.9 g·kg−1 | Sourdough | [149] |
| 3.2 g·kg−1 | ||||||
| 4.2 g·kg−1 | ||||||
| 7.2 × 108 | 8 g·kg−1 | [151] | ||||
| 36.4 g·L−1 | MRS + Sucrose | [153] | ||||
| W. cibaria (11GM-2) | Sour milk | Dextran | >2 × 107 | ND | MRS + Sucrose | [152] |
| W. cibaria (CMGDEX3) | Cabbage | Dextran | >2 × 106 | 0.24 g·L−1 | MRS + Sucrose | [154] |
| W. cibaria (JAG8) | Apple | Dexran | 8 × 105 | 38 g·L−1 (12 h) | Tsuchiya medium | [155,156] |
| W. confusa (E392) | Soured carrot mash | Dextran | ND | ND | MRS + Sucrose | [157] |
| W. confusa (KR780676) | Idli batter | Galactan | ND | 17.2 g·kg−1 (dry weight) | MRS + Sucrose | [87] |
| W. confusa (F3/2-2) | Cassava fermentation | Dextran | >2 × 107 | ND | MRS + Sucrose | [152] |
| Levan | 2 × 105 | ND | MRS + Raffinose | |||
| W. confusa (8CS-2) | Sour milk | HePS | ND | ND | MRS + Glucose | [152] |
| Dextran | >2 × 107 | ND | MRS + Sucrose | |||
| W. confusa (NH 02) | Nham | ND | 1.13 × 106 | 18.08 g·L−1 | MRS + Sucrose | [104] |
| W. hellenica (SKkimchi3) | Kimchi | Glucan | 2.03 × 105 | 5.12 g·L−1 | MRS + Sucrose | [45] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fessard, A.; Remize, F. Why Are Weissella spp. Not Used as Commercial Starter Cultures for Food Fermentation? Fermentation 2017, 3, 38. https://doi.org/10.3390/fermentation3030038
AMA Style
Fessard A, Remize F. Why Are Weissella spp. Not Used as Commercial Starter Cultures for Food Fermentation? Fermentation. 2017; 3(3):38. https://doi.org/10.3390/fermentation3030038
Chicago/Turabian StyleFessard, Amandine, and Fabienne Remize. 2017. "Why Are Weissella spp. Not Used as Commercial Starter Cultures for Food Fermentation?" Fermentation 3, no. 3: 38. https://doi.org/10.3390/fermentation3030038
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.