A Microtiter Plate Assay as a Reliable Method to Assure the Identification and Classification of the Veil-Forming Yeasts during Sherry Wines Ageing

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Mitochondrial DNA (mtDNA) Restriction Analysis
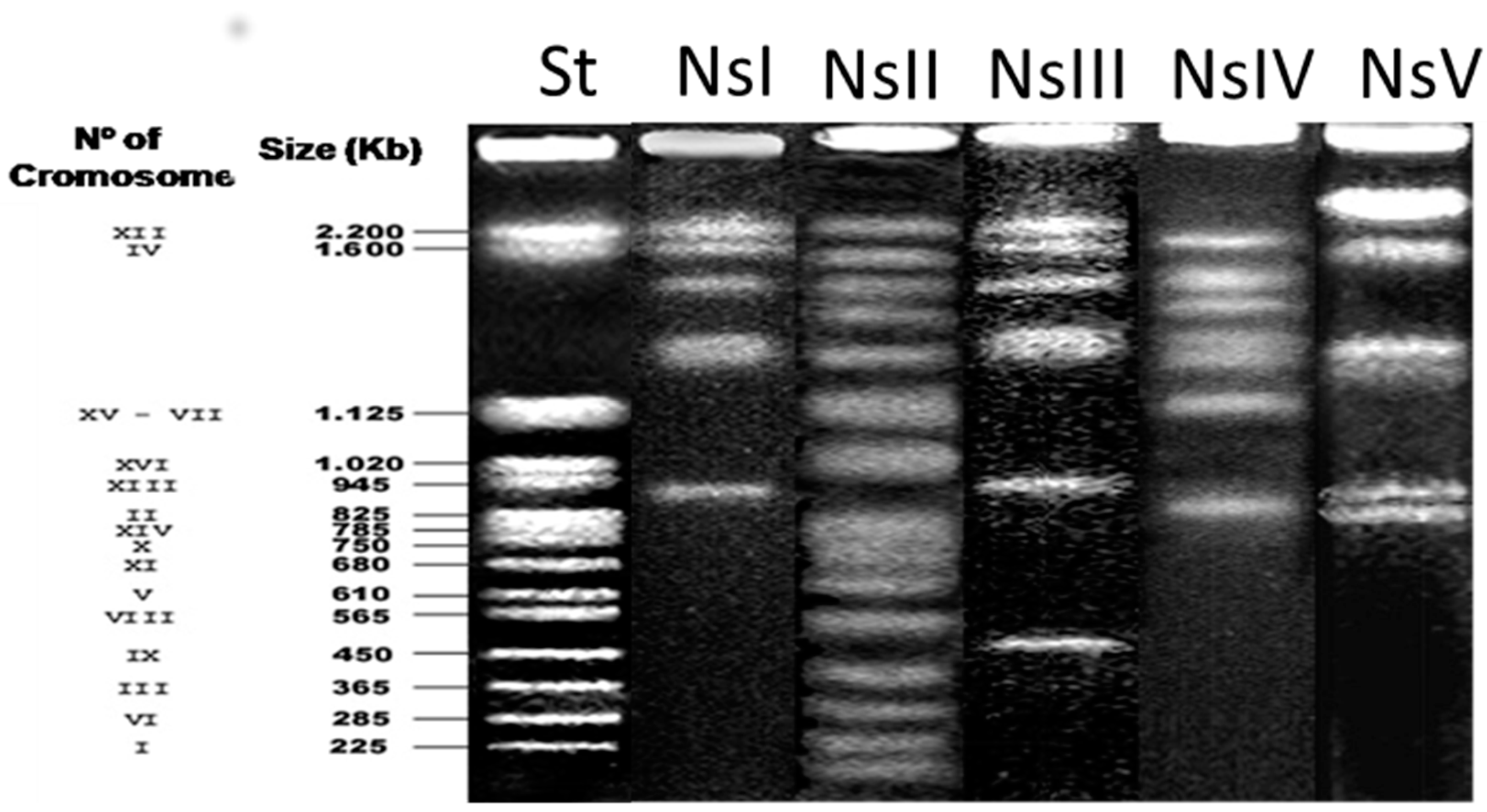
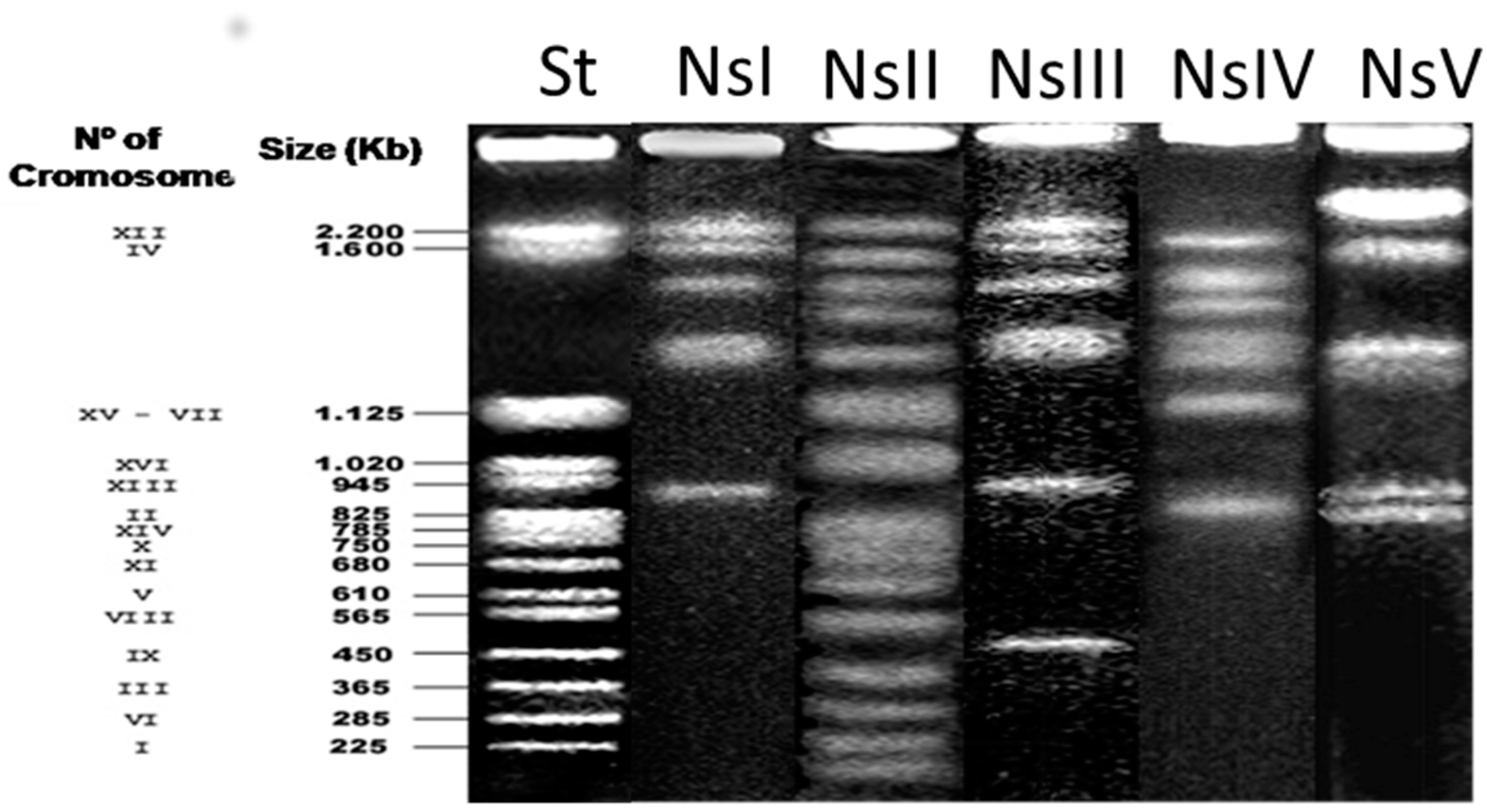
2.3. Electrophoretic Karyotype
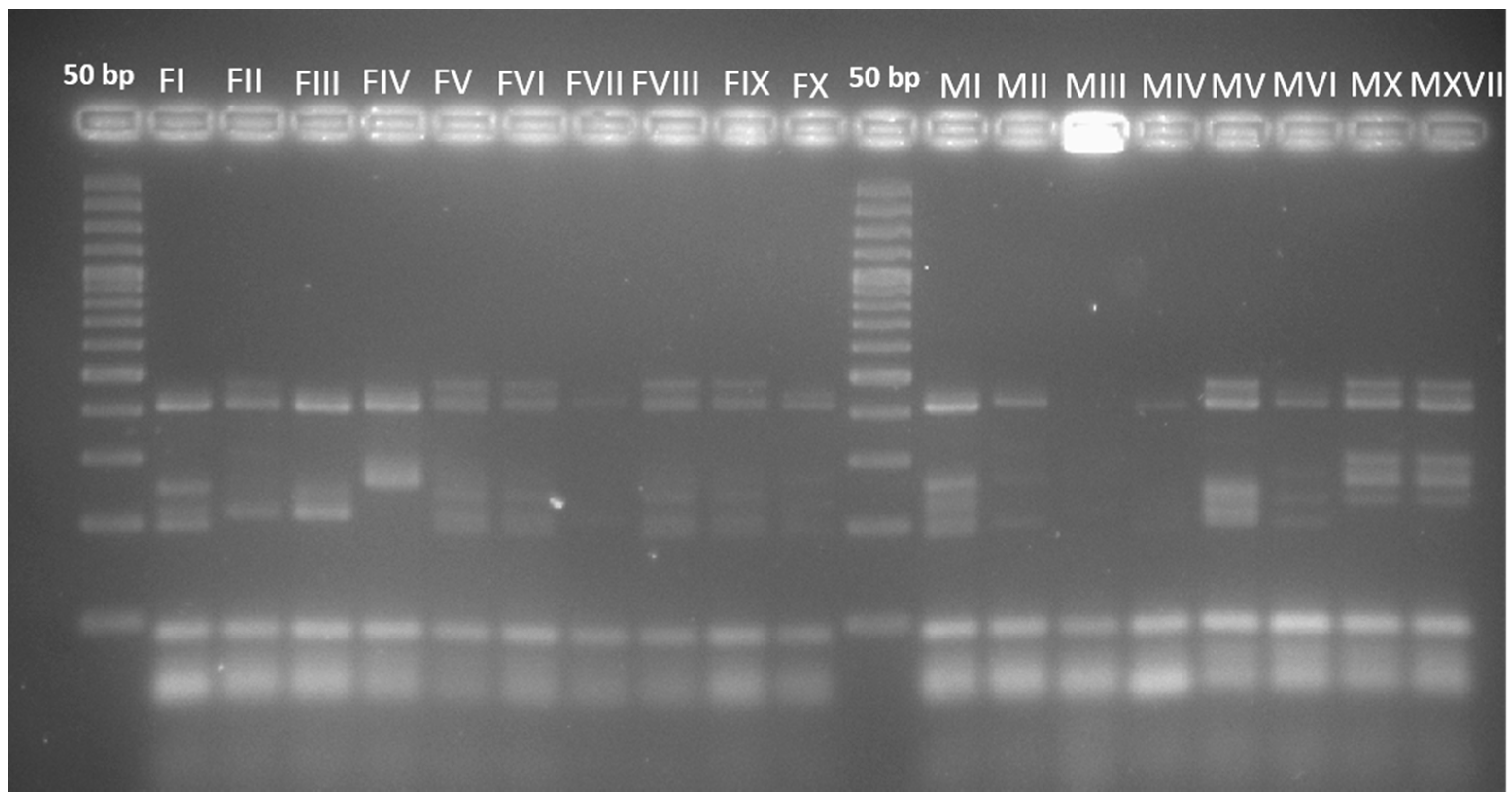
2.4. Multiplex PCR-Microsatellite (SSR) Analysis
2.5. Physiological and Biochemical Tests
2.6. Validation and Reproducibility of the Tests in Microtiter Plates
3. Results
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rodríguez, M.E.; Infante, J.J.; Mesa, J.J.; Rebordinos, L.; Cantoral, J.M. Enological behavior of biofilms formed by genetically-characterized strains of sherry flor yeast. Open Biotechnol. J. 2013, 7, 23–29. [Google Scholar] [CrossRef]
- Mesa, J.J.; Infante, J.J.; Rebordinos, L.; Cantoral, J.M. Characterisation of yeasts involved in the biological ageing of sherry wines. LWT-Food Sci. Technol. 1999, 32, 114–120. [Google Scholar] [CrossRef]
- Esteve-Zarzoso, B.; Peris-Torán, M.J.; García-Maiquez, E.; Uruburu, F.; Querol, A. Yeast population dynamics during the fermentation and biological aging of sherry wines. Appl. Environ. Microbiol. 2001, 67, 2056–2061. [Google Scholar] [CrossRef] [PubMed]
- Martínez, P.; Codon, A.C.; Pérez, L.; Benítez, T. Physiological and molecular characterization of flor yeasts: Polymorphism of flor yeast population. Yeast 1995, 11, 1399–1411. [Google Scholar] [CrossRef] [PubMed]
- Boekhout, T.; Kurtzman, C.P. Principles and methods used in yeast classification, and an overview of currently accepted yeast genera. In Nonconventional Yeasts in Biotechnology; Springer: Berlin/Heidelberg, Germany, 1996; pp. 1–81. ISBN 978-3-642-79856-6. [Google Scholar]
- Vancanneyt, B.P.; Hennebert, G.; Kersters, K. Differentiation of yeast species based on electrophoretic whole-cell protein patters. Syst. Appl. Microbiol. 1991, 14, 23–32. [Google Scholar] [CrossRef]
- Silva, M.M.D.; Malfeito-Ferreira, M.; Loureiro, V.; Aubyn, A.S. Long-chain fatty acid composition as a criterion for yeast distinction in the brewing industry. J. Inst. Brew. 1994, 100, 17–22. [Google Scholar] [CrossRef]
- Esteve-Zarzoso, B.; Fernández-Espinar, M.T.; Querol, A. Authentication and identification of Saccharomyces cerevisiae “flor” yeast races involved in sherry ageing. Antonie van Leewenhoek 2004, 85, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Marin-Menguiano, M.; Romero-Sánchez, S.; Barrales, R.R.; Ibeas, J.I. Population analysis of biofilm yeasts during fino sherry wine aging in the Montilla Moriles D.O. region. Int. J. Food Microbiol. 2017, 244, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.E.; Infante, J.J.; Molina, M.; Rebordinos, L.; Cantoral, J.M. Using RFLP-mtDNA for the rapid monitoring of the dominant inoculated yeast strains in industrial wine fermentation. Int. J. Food Microbiol. 2011, 145, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Querol, A.; Barrio, E.; Huerta, T.; Ramón, D. Molecular monitoring of wine fermentatios conducted by active dry yeast strains. Appl. Environ. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [PubMed]
- Vaudano, E.; García-Moruno, E. Discrimination of Saccharomyces cerevisiae wine strains using microsatellite multiplex PCR and band pattern analysis. Food Microbiol. 2008, 25, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Legras, J.C.; Moreno-García, J.; Zara, S.; Zara, G.; García-Martínez, T.; Mauricio, J.C.; Mannazzu, I.; Coi, A.L.; Zeidan, M.B.; Dequin, S.; et al. Flor yeast: New perspectives beyond wine aging. Front. Microbiol. 2016, 7, 503. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study; Elvesier: London, UK, 2011; p. 2363. [Google Scholar]
- Fernández-Espinar, M.T.; Esteve-Zarzoso, B.; Querol, A.; Barrio, E. RFLP analysis of the ribosomal internal transcribed spacers and the 5.8 S rRNA gene region of the genus Saccharomyces: A fast method for species identification and the differentiation of flor yeasts. Antonie van Leeuwenhoek 2000, 78, 87–97. [Google Scholar] [CrossRef]
- Lopandic, K.; Pfliegler, W.P.; Tiefenbrunner, W.; Gangl, H.; Sipiczki, M.; Sterlinger, K. Genotypic and phenotypic evolution of yeast interspecies hybrids during high-sugar fermentation. Appl. Microbiol. Biotechnol. 2016, 100, 6331–6343. [Google Scholar] [CrossRef] [PubMed]
- Espinazo-Romeu, M.; Cantoral, J.M.; Matallana, E.; Aranda, A. Btn2p is involved in ethanol and biofilm formation in flor yeast. FEMS Yeast Res. 2008, 8, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Bueso, G.; Rodríguez, M.E.; Garrido, C.; Cantoral, J.M. Rapid and not culture-dependent assay based on multiplex PCR-SSR analysis for monitoring inoculated yeast strains in industrial wine fermentations. Arch. Microbiol. 2017, 199, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.E.; Infante, J.J.; Molina, M.; Domínguez, M.; Rebordinos, L.; Cantoral, J.M. Genomic characterization and selection of wine yeast to conduct industrial fermentations of a white wine produced in a SW Spain winery. J. Appl. Microbiol. 2010, 108, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Van Der Walt, J.P.; Yarrow, D. Methods for the isolation, maintenance, classification and identification of yeasts. In The Yeasts: A Taxonomic Study, 3rd ed.; Kreger-van Rij, N.J.W., Ed.; Elvesier Science Publishers: Amsterdan, The Netherlands, 1984; pp. 45–104. [Google Scholar]
- Sipiczki, M. Overwintering of vineyard yeasts: Survival of interacting yeast communities in grapes mummified on vines. Front. Microbiol. 2016, 7, 212. [Google Scholar] [CrossRef] [PubMed]
- Ibeas, J.I.; Jiménez, J. Mitochondrial DNA loss caused by etanol in Saccharomyces flor yeasts. Appl. Environ. Microbiol. 1997, 63, 7–12. [Google Scholar] [PubMed]
- Kvitek, D.J.; Will, J.L.; Gasch, A.P. Variations in Stress Sensitivity and Genomic Expression in Diverse S. cerevisiae Isolates. PLoS Genet. 2008, 4, e1000223. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, H. Flor yeasts of Saccharomyces cerevisiae—Their ecology, genetics and metabolism. Int. J. Food Microbiol. 2013, 167, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J.A.; Payne, R.W.; Yarrow, D. Yeasts: Characterisation and Identification, 3rd ed.; Cambridge University Press: Cambridge, UK, 2000; p. 1139. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| McFarland Standard | 0.5 | 1 | 2 | 3 | 4 |
|---|---|---|---|---|---|
| 1% Barium chloride (mL) | 0.05 | 0.1 | 0.2 | 0.3 | 0.4 |
| 1% Sulfuric acid (mL) | 9.95 | 9.9 | 9.8 | 9.7 | 9.6 |
| Cell density ± 108 cells/mL | 1.5 | 3 | 6 | 9 | 12 |
| Absorbance (610 nn) | 0.1 | 0.25 | 0.45 | 0.60 | 0.67 |
| Strain Designation | Sugar Fermentation | |||||
|---|---|---|---|---|---|---|
| Fructose | Glucose | Lactose | Maltose | Rafinose | Sucrose | |
| FI | − | + | − | − | − | − |
| FII | + | + | − | − | + | + |
| FIII | + | + | − | − | − | − |
| FIV | + | + | − | − | − | + |
| FV | + | + | − | − | − | + |
| FVI | + | + | − | − | + | + |
| FIX | − | + | − | − | + | + |
| FX | − | + | + | − | − | − |
| MI | + | − | − | − | − | + |
| MII | + | + | + | − | + | + |
| MIII | − | − | + | − | − | − |
| MIV | + | + | + | − | + | − |
| MV | − | + | − | + | − | − |
| MVI | + | + | − | − | + | + |
| MX | + | + | − | − | + | + |
| Strain Designation | Carbon/Nitrogen Compounds | ||||||
|---|---|---|---|---|---|---|---|
| Ammonium Citrate | Glucose | Inulin | Melibiose | Nitrite | Starch | Urea | |
| FI | + | − | − | − | − | − | − |
| FII | + | − | − | − | − | . | − |
| FIII | + | − | − | − | − | − | − |
| FIV | + | − | − | − | − | − | − |
| FV | + | − | − | − | − | − | − |
| FVI | − | − | − | − | − | − | − |
| FIX | − | + | + | − | − | − | − |
| FX | − | − | − | − | − | − | − |
| MI | + | − | − | − | − | − | − |
| MII | + | − | + | − | − | − | + |
| MIII | + | − | + | − | − | − | + |
| MIV | + | − | − | − | − | − | − |
| MV | − | + | − | + | + | − | − |
| MVI | + | − | − | − | − | + | − |
| MX | − | − | + | − | − | + | − |
| Strain Designation | Carbon/Nitrogen Compounds | |||||||
|---|---|---|---|---|---|---|---|---|
| Raffinose | Galactose | Inulin | Melibiose | Maltose | Lactose | l-Sorbose | Ethanol | |
| NsI | − | + | − | − | − | − | − | + |
| NsII | + | + | + | − | + | − | − | + |
| NsIII | − | + | − | − | + | − | − | + |
| NsIV | + | + | − | − | + | − | − | + |
| NsV | − | − | − | − | − | + | + | + |
| Strain Designation | Osmotolerance: Growth on Media Supplemented with 1:1 (Glucose:Fructose) and Cycloheximide Resistance | ||||||
|---|---|---|---|---|---|---|---|
| 2% | 20% | 30% | 40% | 50% | Cycloheximide 0.01% | Cycloheximide 0.1% | |
| FI | + | + | + | − | − | − | − |
| FII | + | + | − | − | − | − | − |
| FIII | + | + | + | − | − | − | − |
| FIV | + | + | + | − | − | − | − |
| FV | + | + | + | − | − | − | − |
| FVI | + | + | + | − | − | − | − |
| FVII | + | + | − | − | − | − | − |
| FVIII | + | + | − | − | − | − | − |
| FIX | + | + | + | − | − | − | − |
| FX | + | + | + | − | − | − | − |
| MI | + | + | + | − | − | − | − |
| MII | + | + | + | − | − | − | − |
| MIII | + | + | + | − | − | − | − |
| MIV | + | + | + | − | − | − | − |
| MV | + | + | + | − | − | − | − |
| MVI | + | + | + | − | − | + | − |
| MX | + | + | + | − | − | + | − |
| MXVIII | + | + | + | − | − | − | − |
| NsI | + | + | − | − | − | − | − |
| NsII | + | + | + | + | + | − | − |
| NsIII | + | + | + | + | + | − | − |
| NsIV | + | + | − | − | − | + | − |
| NsV | + | + | − | − | − | + | − |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruíz-Muñoz, M.; Bernal-Grande, M.D.C.; Cordero-Bueso, G.; González, M.; Hughes-Herrera, D.; Cantoral, J.M. A Microtiter Plate Assay as a Reliable Method to Assure the Identification and Classification of the Veil-Forming Yeasts during Sherry Wines Ageing. Fermentation 2017, 3, 58. https://doi.org/10.3390/fermentation3040058
Ruíz-Muñoz M, Bernal-Grande MDC, Cordero-Bueso G, González M, Hughes-Herrera D, Cantoral JM. A Microtiter Plate Assay as a Reliable Method to Assure the Identification and Classification of the Veil-Forming Yeasts during Sherry Wines Ageing. Fermentation. 2017; 3(4):58. https://doi.org/10.3390/fermentation3040058
Chicago/Turabian StyleRuíz-Muñoz, Marina, Maria Del Carmen Bernal-Grande, Gustavo Cordero-Bueso, Mónica González, David Hughes-Herrera, and Jesús Manuel Cantoral. 2017. "A Microtiter Plate Assay as a Reliable Method to Assure the Identification and Classification of the Veil-Forming Yeasts during Sherry Wines Ageing" Fermentation 3, no. 4: 58. https://doi.org/10.3390/fermentation3040058