The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor

Department of Biology, University of North Florida, Jacksonville, FL 32224, USA
Fermentation 2018, 4(1), 20; https://doi.org/10.3390/fermentation4010020
Submission received: 2 February 2018
/
Revised: 12 March 2018
/
Accepted: 13 March 2018
/
Published: 19 March 2018
(This article belongs to the Special Issue Brewing & Distilling)
Abstract
:Beer is a complex beverage containing a myriad of flavor- and aroma-active compounds. Brewers strive to achieve an appropriate balance of desired characters, while avoiding off-aromas and flavors. Phenolic compounds are always present in finished beer, as they are extracted from grains and hops during the mashing and brewing process. Some of these compounds have little impact on finished beer, while others may contribute either desirable or undesirable aromas, flavors, and mouthfeel characteristics. They may also contribute to beer stability. The role of simple phenolic compounds on the attributes of wort and beer are discussed.

1. Introduction
There are approximately 1000–2000 chemical compounds in beer [1]. Many of these are present at levels well below their flavor, aroma, and other perception thresholds, and therefore, do not contribute to beer perceptions. The wide array of flavors, aromas, and mouthfeel attributes across world beer styles result from the presence, to varying degrees, of many organic and inorganic compounds. For many compounds, we have little understanding of the potential contribution to beer aroma and flavor, while others have been investigated extensively.
A wide range of phenolic compounds are present in beer, extracted from raw materials during the malting, mashing, and brewing process. These include complex polyphenols that contribute to mouthfeel, beer haze, antioxidant properties and stability, as well as foam retention [2,3]. Simple phenolic compounds can have a major impact on beer aroma and flavor, both positive and negative. In this paper, the role of these simple phenolic compounds in raw ingredients, their transformation during brewing and fermentation, and their impact on beer organoleptic properties is reviewed.
2. The Role of Phenolic Compounds in Plants
Simple and volatile phenols in beer all originate from plant material, primarily malted grains and hops. The vast majority are extracted from raw materials during the brewing process (see Section 3 below). Other sources are less direct, either through extraction of phenols from wood during barrel aging, or from burning of wood in smoked malt beers. These indirect sources are still plant-derived.
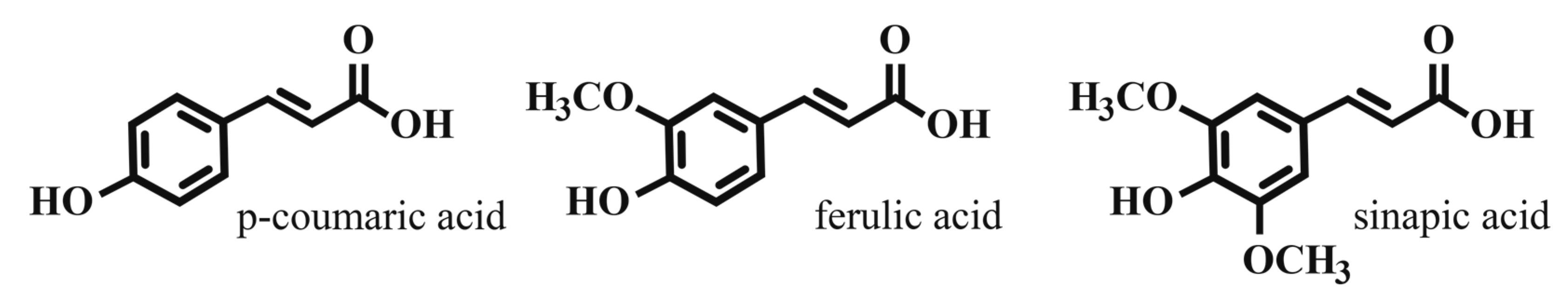
Phenolic compounds are an integral and abundant component of the plant cell wall. Ferulic acid, p-coumaric acid, and sinapic acid are hydroxycinnamic acids (HCAs) that are converted to their hydroxyl-cinnamyl alcohol derivatives. These compounds are exported to the plant cell wall in a poorly understood process. In the cell wall, these “monolignols” react to form the complex polymer lignin, a major cell wall component that protects cell wall fibers and contributes to cell rigidity [4].
These same hydroxycinnamic acids also provide crucial cross-links between lignin and non-lignin fiber components of the cell wall, most commonly hemicellulose. Ester linkages between HCAs and cell wall polysaccharides, particularly arabinoxylans are well-documented. Dehydrodiferulic acid also participates in polysaccharide-polysaccharide and polysaccharide-lignin bridges [5,6].
In addition to their structural role, HCAs contribute to plant defense against pathogenic microbes. They reduce the biodegradability of cell wall polymers, serving as a structural defense to cell wall breakdown [6]. Free HCAs as well as other phenolic cell wall components are directly toxic to microbial cell membranes, leading to increased ion permeability, thereby reducing cell viability [7]. Common representative HCA structures are shown in Figure 1.
3. Extraction of Simple Phenolics into Beer Wort
3.1. Raw Materials
3.1.1. Grains and Malt
Cereal grains and malt account for the majority of the total phenolic acid content of beer wort [8]. In barley malt, ferulic acid and p-coumaric acid predominate. HCA levels vary among barley varieties, and may be influenced by malting process, growing environment and barley handling post-harvest. Published values vary widely, ranging from 10 to 15 µg/g dry weight combined [9,10] to highs of ~100–300 µg/g dry weight for p-coumaric acid, and ~400–600 µg/g dry weight for ferulic acid [11,12]. These wide variances may be due to extraction method, analytical methods, and whether free, bound, or all forms of HCAs are analyzed. In all varieties investigated, ferulic acid predominates by a factor of 2–5 [3]. HCAs in malt are available for extraction into wort in two forms, free and esterified. Esterified HCAs are primarily bound to arabinoxylans. Greater than 90% of the HCA in malt is ester-bound, and its availability for malt extraction and later conversion to free HCA is dependent on arabinoxylan-degrading enzymes in the barley, mainly active during the mashing process [11,12,13,14,15].
Most analyses of barley and barley malt have been carried out on pilsner or other base malt varieties. Vanbeneden et al. [13] have shown that some specialty malts with higher degrees of kilning have altered potential extraction of HCAs. Worts produced with lighter kilned specialty malts such as Munich and Vienna did not differ in HCA extraction from base malt worts. Higher kilned malts (caramunich and carafa) yielded lower ferulic acid to the wort, with a corresponding increase in 4-vinylguaiacol, the decarboxylation product of ferulic acid [13,16]. It is proposed that higher kilning temperatures lead to some degree of thermal decarboxylation during the kilning process. These analyses were carried out on laboratory worts prepared from 50% pilsner malt and 50% specialty malt. It is expected that at the lower percent of specialty malts often used in brewing that these malts will not significantly impact the HCA composition of the wort [13].
Release of ester-bound HCAs into wort occurs primarily during mashing. Barley enzymes include arabinofuranosidases and β-xylan endohydrolases, which degrade and solubilize the arabinoxylan chains and allow extraction into wort [15]. Perhaps more important is cinnamoyl esterase, which releases HCAs from xylan subunits, making them available for further metabolism [11,12,13,15,17]. These enzymes are active under typical mashing conditions, but have temperature optima lower than many mash regimens. If a high level of volatile phenol precursors (ferulic acid) is desired, a 40 °C mash rest is recommended [13].
Not surprisingly, there is notable variability in extraction of free and ester-bound HCAs into wort using a standard congress mash. In one study, 6–13 µg/g representing 0.7–1.5 ppm p-coumaric acid was released into wort. This represented only 2.3–5.5% of the available p-coumaric acid in the malt. Extraction of ferulic acid was slightly more efficient, yielding 7.1–12.5% of that available in the malt. This achieved 42–70 µg/g, or 4.9–8.3 ppm ferulic acid in the wort [11]. These numbers are in general agreement with similar studies that used different sets of barley malts [9,14,18,19,20], however wide ranges can be found within the published literature [21]. This observed variability is most commonly linked to alternative extraction methods between researchers. Interestingly, the proportion of free versus ester-bound p-coumaric acid released to wort was significantly and consistently higher than for ferulic acid: 60–80% for p-coumaric acid and 20–43% for ferulic acid. A similar proportion of the two free HCAs was determined to have been enzymatically released during mashing [11]. These wort values also correlate well with analysis of HCAs present in finished beer [22]. Overall, these data reflect the consistent observation that most barley varieties are significantly higher in ferulic acid compared to p-coumaric acid.
Malted and unmalted wheat makes up a significant and important proportion of the grist of several specialty beer styles, including Bavarian weizen (weissbier), Berlinerweiss, Belgian witbier, lambic, and geueze. It is a common ingredient in lower proportions in many other styles, and is often added at 2–5% of the grist of beers where additional head retention and mouthfeel are desired, due to the higher protein content of wheat versus barley [23]. Analysis of wheat varieties for phenolic acid content reveals a much higher overall yield than barley, with ferulic acid constituting 70–90% of the available phenolic acids [24,25,26,27,28]. Weissbier style is characterized by a clove-like aroma and flavor [13,29]. This is a result of the volatile phenol 4-vinylguaiacol at levels above the aroma/flavor threshold. This compound is generated primarily by metabolism of ferulic acid extracted from malted wheat (and barley) grist by “phenolic off-flavor positive” (POF+) yeast strains (see Section 4.2 below). When high 4-vinylguaiacol is desired in this (or other) styles, a ferulic acid rest can be added to the mash regimen, allowing feruoyl esterase to release additional free ferulic acid into the wort [13].
3.1.2. Hops
There is less published work on HCAs and other phenolic acids for hops compared to cereal grains. Published experimental values are wide ranging and difficult to interpret, at least partly due to variations in extraction procedures [30,31,32,33]. A general observation is that hops appear to be somewhat higher (two to five fold) in HCA content compared to barley, when analyzed simultaneously [9,10]. Hopping rates vary widely among different beer styles, and the use and form of hops will also vary. Hop cone plant material will contribute the bulk of the simple phenolics. Modern breweries have many choices of hop form, including whole cone, plugs, and pellets, as well as newer hop extracts. Pellets are available as type-90 or type-45, which contain 90 percent and 45 percent, respectively, of raw hop cone plant material [30]. This may impact the potential extraction of phenolic acids since the lupulin gland contents will be concentrated relative to other hop cone components; however limited experimental data has been inconclusive [30]. A general conclusion is that while the phenolic acid constituent of hops may be slightly higher than for barley, the very high dry mass ratio of malt to hops in most beer recipes makes the hop contribution to beer volatile phenols low relative to grain.
3.2. Wood Aging
Historically, beer has been fermented, aged, and served from wooden vessels [34]. Long-term wood-aging has been carried through for certain specialty beer styles, such as lambic and Flanders red ales [35,36,37,38,39,40,41]. There has been a resurgence of barrel and wood-aging in recent decades, particularly among the craft brewing industry. Often the goal of the brewer is to impart the character of distilled or fermented beverages that were previously held in the barrel, but characteristics from the wood itself will be transferred to the beer during aging, and may contribute significantly to the organoleptic properties of the finished beer. A wide range of volatile compounds are transferred to beer and other beverages aged in barrels or on wood chips [42,43]. Of the phenolic compounds, vanillin and vanillic acid are of high interest, imparting sweet or vanilla-like flavors to the beer [10]. Guaiacol, contributing to smoky or burnt character, will also be significant if the barrel is charred or heavily toasted [44]. The content and extraction of compounds from wood during aging has been shown to vary dramatically based on a number of variables. Extractable compounds vary by wood species, as well as geographical region of origin [45,46,47]. The degree of wood seasoning as well as toast level also influences phenolic content, with higher toast levels generally showing higher phenolic content [42,45,47]. Breweries often reuse barrels, and the history of barrel use will greatly influence the contribution of the wood to the finished beer.
4. Transformation of Hydroxycinnamic Acids to Volatile Compounds
Hydroxycinnamic acids are always present in wort and beer, extracted from grains and hops. These compounds have high flavor/aroma thresholds, generally greater than 500 ppm [14,48], and are present in wort or beer and levels far below this threshold (see Section 3.1.1). HCAs can be converted into vinyl derivatives and possibly ethyl derivatives by several mechanisms. These new compounds have very low organoleptic thresholds [14,48] and may make significant contributions to the aroma and flavor of the finished beer.
4.1. Thermal Decarboxylation to Vinyl Derivatives
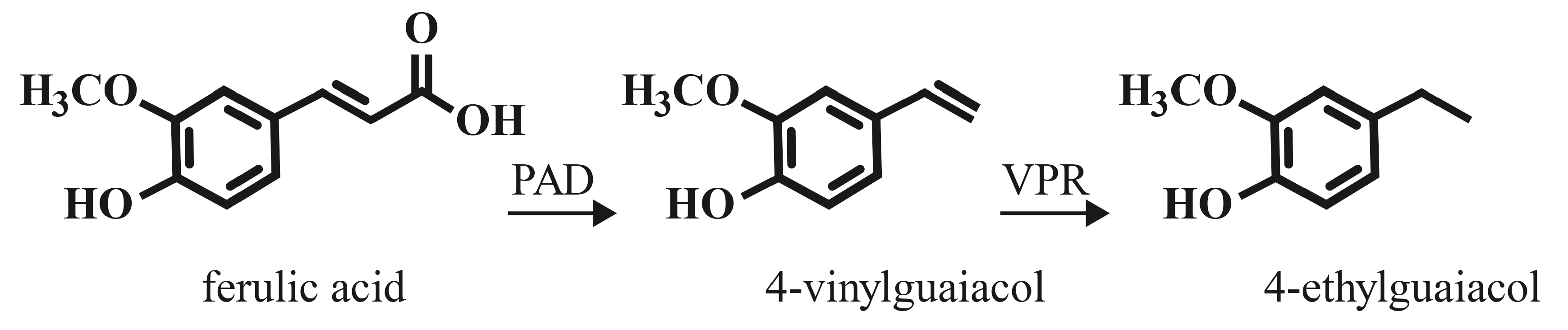
4-vinylguaiacol (4-VG) and 4-vinylphenol (4-VP) are the decarboxylation products of ferulic acid and p-coumaric acid, respectively, the two most abundant phenolic acids in cereal grains and beer wort. There do not appear to be active phenolic acid decarboxylase enzymes in barley malt, as mashing regimens do not produce 4-VG in unboiled wort [18]. This experimental data also demonstrates that mashing temperatures are not adequate for thermal decarboxylation of ferulic acid [18]. Over the course of a three hour boil, levels of 4-VG increased, with highest levels generated in congress wort that utilized a portion of wheat malt, with levels approached threshold values (0.3 mg/L) [18,49]. This value represents about ten percent of the free ferulic acid present in the pre-boil wort. Related HCAs would be expected to undergo similar thermal decarboxylation during wort boiling.
4.2. Microbial Enzymatic Decarboxylation to Vinyl Derivatives
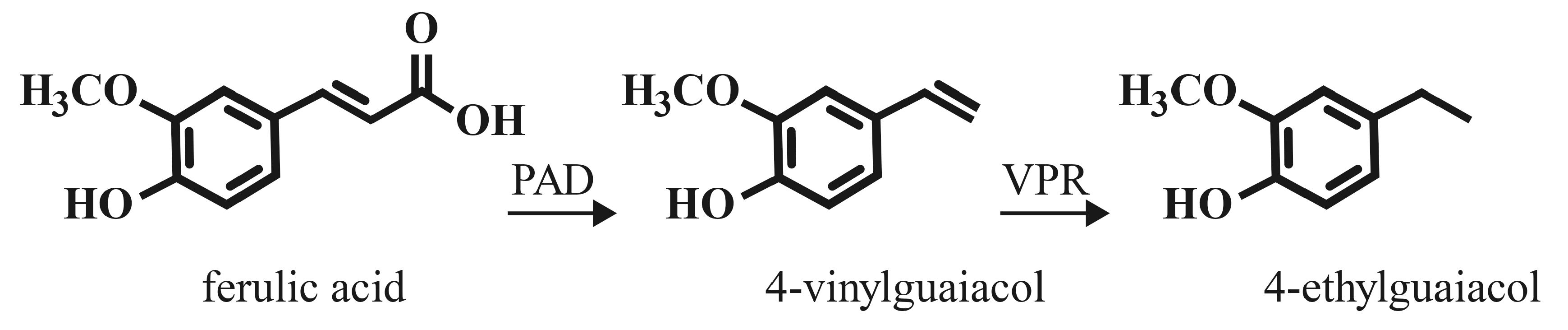
A wide variety of bacteria and fungi encode a phenolic acid decarboxylase enzyme (PAD; also known as ferulic acid decarboxylase, courmaric acid decarboxylase, cinnamate decarboxylase). These include species which represent contaminants in a brewing environment, but also species and strains utilized in brewing. Many strains of brewer’s yeast (Saccharomyces cerevisiae), as well as Brettanomyces sp., Lactobacillus, and Pediococcus are PAD+ [50,51,52,53,54]. These non-Saccharomyces genera of microbes are utilized in traditional sour beers of Belgium, and increasingly in other “wild” and sour beer styles popular in the craft brewing industry [36,37,39,40,41,50]. PAD catalyzes the enzymatic decarboxylation of HCAs to their vinyl derivatives (see Figure 2). This enzyme activity may have evolved in microbes that occupy an environment rich in plant matter (fruit skins, soil, etc.) as a stress-response countermeasure to the antimicrobial activity of the HCA compounds [51].
In Saccharomyces yeast strains, a POF+ phenotype results from active decarboxylation of cinnamic acid derivatives. This “phenolic off-flavor” character requires two enzymes, a phenylacrylic acid decarboxylase (PAD1) and a ferulic acid decarboxylase (FDC1). Somewhat confusingly, the PAD1 enzyme is not a decarboxylase, but catalyzes the synthesis of an FMN-related co-factor required for the function of FDC1 [52,55]. FDC1 enzyme catalyzes the decarboxylation of cinnamic acid and derivatives. Among the well-characterized microbial decarboxylase enzymes, the Saccharomyces FDC1 is of the few that can use cinnamic acid as a substrate yielding styrene as a product; others are only functional on hydroxycinnamic acids [52].
S. cerevisiae Pad1 and Fdc1 genes are analogous to the UbiX and UbiD genes of E. coli, respectively, involved in the ubiquinone biosynthetic pathway [53,55]. The recently determined 3D structure of the full-length 503 amino acid FDC1 protein reveals a cinnamic acid substrate binding site that can be modeled to accept the much larger substrate predicted for a role in the ubiquinone pathway [52]. Pad1/Fdc1 double knockout yeast are not deficient in ubiquinone biosynthesis however, suggesting the possible presence of additional genes for this biosynthetic pathway [55]. A survey of industrial brewing yeasts reveals a wide range of PAD activity [49]. Further evidence for a primary role for PAD1/FDC1 in cinnamic acid metabolism can readily be seen in a large-scale genomic analysis of industrial yeast strains [56]. Of 157 sequenced strains of S. cerevisiae, the majority of strains selected for use in alcoholic beverages have lost cinnamic acid decarboxylation function. A variety of loss-of-function mutations are found in either Pad1, Fdc1, or both among beer, wine, and sake strains (POF−), however all strains sequenced that fall into “wild”, industrial, or bread baking groups retain POF activity. Among the strains sequenced are three Bavarian wheat beer strains, where POF+ activity is essential for the clove/spice character attributed to 4-VG. These strains broadly align with other beer strains, but show significant mosaicism. The authors suggest a more recent hybridization for these strains, in which previously selected traits desirable for beer brewing combined with functional Pad1 and Fdc1 alleles likely derived from wine strains [56].
PAD enzymes have been characterized from several sour ale brewing-relevant bacteria, including Lactobacillus brevis and L. plantarum. The sequence of these enzymes reveals significant conservation across genera, and a structure and mechanism that does not require a known co-factor, unlike the S. cerevisiae PAD1/FDC1 system [54,57]. This group constitutes a PAD enzyme family that shares a flattened, compact beta-barrel structure, with the substrate binding site within the core of the barrel [57]. In limited testing of substrate specificity across strains, there appear to be two subgroups of enzymes. One group, including L. brevis, Bacillus subtilis, and B. pumilus, shows similar substrate specificity for the three main substrates p-coumaric acid, ferulic acid, and caffeic acid. The group including L. plantarum and P. pentosaceus strongly prefers p-coumaric acid over ferulic acid [58,59].
Yeast of the Brettanomyces genus are well-known as spoilage organisms in the wine and beer industries. Interestingly, they also contribute key organoleptic qualities to specialty beer styles including lambic, gueuze, Flanders red and brown ales, Berlinerweiss, and others [36,37,39,40,41,50,60,61,62,63]. They are increasingly utilized in other new “wild” ale styles in the craft brewing industry. The contribution to beer aroma and flavor resulting from Brettanomyces fermentation is complex and generally poorly characterized, however volatile phenol formation is a well-known component of Brettanomyces metabolism in beer and wine. Commercial interest in these yeasts has resulted in a growing body of data related to their contribution to wine and beer flavor. Genetic and physiological analysis of B. bruxellensis strains that vary in geographical and source environment suggests niche adaptation, as strains cluster more strongly by the isolation environment than by geography [64,65,66,67,68]. There are notable differences between beer and wine strains at the whole genome level and for SNPs, including genes involved in volatile phenol production [66,67]. The Brettanomyces PAD enzyme has been characterized and cloned from several strains, including beer and wine isolates. The production of vinyl phenols is strongly strain and growth medium dependent [67,69,70].
4.3. Vinylphenol Reductase Enzyme Activity
The presence of phenolic acid decarboxylase activity is fairly widespread among bacteria and fungi that utilize plant matter habitats. These enzymes decarboxylate hydroxycinnamic acids to less toxic vinyl derivatives as described above (Section 4.2). A few microbes encode a vinylphenol reductase (VPR) enzyme, which uses the vinyl intermediates as substrates to generate ethyl derivatives (see Figure 2). Among brewing relevant organisms, a few Lactobacillus species (L. brevis, L. collinoides, L. plantarum), and P. damnosus, as well as members of the Brettanomyces yeast genus have active VPR enzymes [71,72]. VPR activity appears to be rare among yeast species, but has been demonstrated for Pichia guilliermondii isolated from winery environments [73], although this activity is strain-dependent [74]. The most common products are 4-ethylphenol and 4-ethylguaiacol, derived from p-coumaric acid and ferulic acid, respectively. These compounds have similar organoleptic properties to the vinyl substrates (see Section 5), but have somewhat lower aroma and flavor thresholds [3,49,71].
5. Organoleptic Properties of Volatile Phenols in Beer
The most prevalent HCAs in wort and beer are p-coumaric acid and ferulic acid (see Section 3.1), and most microbial enzymes that metabolize HCAs utilize these two substrates preferentially (see Section 4.2). Therefore, the main volatile phenols found in finished beer are 4-vinylguaiacol and 4-ethylguaiacol (derived from ferulic acid) and 4-vinylphenol and 4-ethylphenol (derived from p-coumaric acid). The HCA precursors have flavor/aroma thresholds well above the amounts found in most beers; 20–50 mg/L threshold [3] versus 1–5 mg/L present in beer [14,22,49,75,76,77,78]. The metabolic products have far lower thresholds, ranging from 0.08 to 0.5 mg/L, values readily achievable from the available substrates [3,49,71]. The aroma and flavor descriptors used for volatile phenols are wide-ranging, and include “stable, barnyard, horsey, leathery, smoky, spicy, clove, medicinal, band-aid®”, and others [63,71,79,80]. The specific aroma and flavor profile contributed by volatile phenols in a particular sample is likely to be influenced by relative concentrations of the compounds present, the total summative concentration of compounds, interactions with other compounds present, and variation among individuals for sensitivity to these compounds. Individual compounds contribute the properties shown in Table 1.
5.1. Desirable Characteristics
In beers brewed with POF+ strains of S. cerevisiae, 4-vinylguaiacol is the most prevalent volatile phenol, as ferulic acid dominates the wort HCAs. This is especially true of beers with a high proportion of wheat, such as Bavarian wheat beers and Belgian white beers [8,85,86,87,88,89,90,91]. The most common descriptors for the phenolic contribution to aroma and flavor in wheat-based beers is “clove” and “spice”, in agreement with Table 1. The high proportion of wheat and corresponding high proportion of ferulic acid makes the spicy phenolic character of these styles a defining feature, although 4-vinylguaiacol will be present in varying concentrations in individual varieties [49,86].
In many Belgian-style specialty beers, POF+ S. cerevisiae strains are required to impart spice notes in the finished beer [92]. The phenolic character is generally more subtle than the wheat-based styles described above. This is likely due at least in part to a lower level of ferulic acid precursors in the wort in the absence of a high proportion of wheat, but variation in POF enzyme activity may also play a role. Several studies have investigated variation in POF activity among a range of brewing yeast strains demonstrating significant variability, however specific yeast varieties were not identified [49,93]. In addition to the clove-like spice commonly encountered in Belgian styles, a “black pepper” character is commonly encountered. This character is considered phenolic, however a specific metabolic compound has not been ascribed to this unique feature. “Peppery” may be a result of a particular interaction between yeast metabolic products, ingredients, and brewing conditions, but is reproducible and attributed consistently to specific examples of Belgian style beers, particularly saisons [94,95,96]. Only a few commercially available brewing yeast strains specifically list “peppery” as an expected descriptor for the finished beer. These include White Labs WLP565, Wyeast 3711, Wyeast 3726, BSI S-11, BSI S-26, and BSI 565. All of these strains are identified by the supplier as a most suitable for saison-style beers [97,98,99].
A desirable smoky character may be present in certain beer styles, particularly smoked beers and to a lesser extent beers with a high proportion of dark roasted barley and malts such as porters and stouts. Smoke character in non-smoked beers is attributed primarily to 4-vinylguaiacol through thermal decarboxylation of ferulic acid during kilning of the dark roasted malts [13,16]. This character may also be derived from enzymatic decarboxylation by POF+ yeast or bacteria. In beers where smoke aroma and flavor are a major component (classic rauchbier, licthenhainer, piwo grodziskie, and other smoked specialty styles), the smoke character comes from the use of malts that are wood-smoked. The smoke character is therefore a result of lignin pyrolysis during wood burning. There is a wide range of pyrolysis products from this process, combinations of which will provide unique properties and variation between wood types. A major contributor to “smoke” character regardless of wood source is the volatile phenol guaiacol [83].
5.2. Undesirable/Spoilage Characteristics
In most beers styles, phenolic character is undesirable and considered a flaw. This can range from inappropriate levels of otherwise pleasant aromas and flavors (spice, clove), to strong and offensive medicinal, plastic strip, goaty, burnt, or creosote character, among others [62,63,64]. The former situation may be due to inappropriate yeast strain choice (POF+ versus POF−), or fermentation at a temperature above the recommended range [91].
For beers that are dominated by offensive phenolic character, the most likely source of volatile phenols is metabolic activity of contaminating spoilage organisms. Most beer spoilage organisms fall into one of several categories: gram positive lactic acid bacteria, gram negative acetic acid bacteria, wild Saccharomyces yeast, and non-Saccharomyces yeast [100,101]. Lactic acid bacteria include a number of species within both the Lactobacillus and Pediococcus genera. While a number of species within these genera possess a PAD enzyme gene and can be considered POF+ (see Section 4.2), they are primarily known for spoilage by acidification of wort or beer through lactic acid fermentation, rather than for generating threshold levels of volatile phenols, although diacetyl formation is also a potential spoilage problem with Pediococcus [100]. Gluconobacter and Acetobacter are the most common brewery contaminants spoiling through metabolism of ethanol to acetic acid, but are also not known for detectable volatile phenol formation [100,102].
Most beer spoilage yeasts that have been characterized exhibit a POF+ phenotype, and volatile phenol production is a major mechanism of spoilage by yeasts. This includes wild strains of S. cerevisiae, as well as members of the Pichia, Candida, Toulaspora, Kloeckera, Brettanomyces, and Schizosaccharomyces genera most commonly associated with brewery contamination, although the degree of volatile phenol production varies widely among isolates [49,93,100,103,104]. A designation of POF+ is usually associated with an analysis of yeasts ability to metabolize HCA precursor compounds. Most POF+ strains are active for phenolic acid decarboxylase activity, leading to generation of vinyl derivatives (see Section 4.2). Among the strains tested, only Brettanomyces isolates have demonstrated vinylphenol reductase enzyme, completing HCA metabolism to ethyl derivatives, primarily 4-ethylphenol and 4-ethylguaiacol (Section 4.3). The ethyl compounds have comparable aroma and flavor thresholds, and are generally associated with less desirable characteristics of barnyard and creosote [3,71].
Phenolic compounds have recently been identified as potential contributors to staling of beer through aging or improper storage. Aging beer has been shown to acquire concentrations of 4-vinyl syringol that are perceived as a strong “old beer” character. This is likely due to degradation of sinapic acid glycosides during aging [3,8]. Long-term storage may also lead to altered flavor profile due to degradation of phenolic compounds. Beers with a desired “spice” aroma and flavor from 4-vinylguaiacol may over time lose this character in favor of a sweeter taste as 4-VG degrades to vanillin. This process is proposed to occur via a pH dependent reaction [84].
6. Conclusions
Simple phenolic compounds are always present in beer, as they are released into wort from grains and hops during the mashing and brewing process. Nevertheless, a brewer has considerable control over aroma and flavor impacts of phenolic compounds on the finished beer. Desirable character can be increased by choice of mashing and wort boiling regimens, as well as ingredient and yeast choice. Reducing the influence of unwanted phenolic character can likewise be achieved by choices in the brewing process, as well as by avoiding unwanted microbes through proper sanitation and proper storage and aging.
Acknowledgments
Thanks to Michael Boyles of the University of North Florida Center for Instruction and Research and Technology for figure preparation, and to Jamie Lynch for critical reading of the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- The Chemistry of Beer. Available online: https://www.newfoodmagazine.com/article/9164/the-chemistry-of-beer/ (accessed on 17 January 2018).
- Bamforth, C. Beer: A Quality Perspective; Elsevier: London, UK, 2009; pp. 12–27. [Google Scholar]
- Callemien, D.; Collin, S. Structure, organoleptic properties, quantification methods, and stability of phenolic compounds in beer—A review. Food Rev. Int. 2009, 26, 1–84. [Google Scholar] [CrossRef]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Iiyama, K.; Lam, T.B.T.; Stone, B.A. Covalent cross-links in the cell wall. Plant Physiol. 1994, 104, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Faulds, C.B.; Williamson, G. The role of hydroxycinnamates in the plant cell wall. J. Sci. Food Agric. 1999, 79, 393–395. [Google Scholar] [CrossRef]
- Campos, F.M.; Couto, J.A.; Figueiredo, A.R.; Tóth, I.V.; Rangel, A.O.S.S.; Hogg, T.A. Cell membrane damage induced by phenolic acids on wine lactic acid bacteria. Int. J. Food Microbiol. 2009, 135, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Collin, S.; Jerkovic, V.; Bröhan, M.; Callemien, D. Polyphenols and Beer Quality. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2333–2359. [Google Scholar]
- McMurrough, I.; Roche, G.P.; Cleary, K.G. Phenolic acids in beers and worts. J. Inst. Brew. 1984, 90, 181–187. [Google Scholar] [CrossRef]
- Wackerbauer, K.; Kramer, P.; Siepert, J. Phenolic aromatic substances in beer. Phenolic carboxylic acid and phenols in the raw material and beer preparation. Brauwelt 1982, 122, 618–620. [Google Scholar]
- Vanbeneden, N.; Gils, F.; Delvaux, F.; Delvaux, F.R. Variability in the release of free and bound hydroxycinnamic acids from diverse malted barley (Hordeum vulgare L.) cultivars during wort production. J. Agric. Food Chem. 2007, 55, 11002–11010. [Google Scholar] [CrossRef] [PubMed]
- Hernanz, D.; Nuñez, V.; Sancho, A.I.; Faulds, C.B.; Williamson, G.; Bartolomé, B.; Gómez-Cordovés, C. Hydroxycinnamic acids and ferulic acid dehydrodimers in barley and processed barley. J. Agric. Food Chem. 2001, 49, 4884–4888. [Google Scholar] [CrossRef] [PubMed]
- Vanbeneden, N.; Van Roey, T.; Willems, F.; Delvaux, F.; Delvaux, F.R. Release of phenolic flavour precursors during wort production: Influence of process parameters and grist composition on ferulic acid release during brewing. Food Chem. 2008, 111, 83–91. [Google Scholar] [CrossRef]
- McMurrough, I.; Madigan, D.; Donnelly, D.; Hurley, J.; Doyle, A.-H.; Hennigan, G.; McNulty, N.; Smyth, M.R. Control of ferulic acid and 4-vinylguaiacol in brewing. J. Inst. Brew. 1996, 102, 327–332. [Google Scholar] [CrossRef]
- Jamar, C.; du Jardin, P.; Fauconnier, M.-L. Cell wall polysaccharides hydrolysis of malting barley (Hordeum vulgare L.): A review. Biotechnol. Agron. Soc. Environ. 2011, 15, 301–313. [Google Scholar]
- Samaras, T.S.; Camburn, P.A.; Chandra, S.X.; Gordon, M.H.; Ames, J.M. Antioxidant properties of kilned and roasted malts. J. Agric. Food Chem. 2005, 53, 8068–8074. [Google Scholar] [CrossRef] [PubMed]
- Sancho, A.I.; Faulds, C.B.; Bartolomé, B.; Williamson, G. Characterisation of feruloyl esterase activity in barley. J. Sci. Food Agric. 1999, 79, 447–449. [Google Scholar] [CrossRef]
- Coghe, S.; Benoot, K.; Delvaux, F.; Vanderhaegen, B.; Delvaux, F.R. Ferulic acid release and 4-vinylguaiacol formation during brewing and fermentation: Indications for feruloyl esterase activity in Saccharomyces cerevisiae. J. Agric. Food Chem. 2004, 52, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Han, Z.; Huang, Y.; Chen, Z.-H.; Zhang, G.; Dai, F. Genetic diversity of individual phenolic acids in barley and their correlation with barley malt quality. J. Agric. Food Chem. 2015, 63, 7051–7057. [Google Scholar] [CrossRef] [PubMed]
- Dvořáková, M.; Douanier, M.; Jurková, M.; Kellner, V.; Dostálek, P. Comparison of antioxidant activity of barley (Hordeum vulgare L.) and malt extracts with the content of free phenolic compounds measured by high performance liquid chromatography coupled with coularray detector. J. Inst. Brew. 2008, 114, 150–159. [Google Scholar] [CrossRef]
- Carvalho, D.O.; Gonçalves, L.M.; Guido, L.F. Overall antioxidant properties of malt and how they are influenced by the individual constituents of barley and the malting process. Compr. Rev. Food Sci. Food Saf. 2016, 15, 927–943. [Google Scholar] [CrossRef]
- Nardini, M.; Ghiselli, A. Determination of free and bound phenolic acids in beer. Food Chem. 2004, 84, 137–143. [Google Scholar] [CrossRef]
- Shewry, P.R. Improving the protein content and composition of cereal grain. J. Cereal Sci. 2007, 46, 239–250. [Google Scholar] [CrossRef]
- Mpofu, A.; Sapirstein, H.D.; Beta, T. Genotype and environmental variation in phenolic content, phenolic acid composition, and antioxidant activity of hard spring wheat. J. Agric. Food Chem. 2006, 54, 1265–1270. [Google Scholar] [CrossRef] [PubMed]
- Dinelli, G.; Segura-Carretero, A.; Di Silvestro, R.; Marotti, I.; Arráez-Román, D.; Benedettelli, S.; Ghiselli, L.; Fernadez-Gutierrez, A. Profiles of phenolic compounds in modern and old common wheat varieties determined by liquid chromatography coupled with time-of-flight mass spectrometry. J. Chromatogr. A 2011, 1218, 7670–7681. [Google Scholar] [CrossRef] [PubMed]
- Zuchowski, J.; Jonczyk, K.; Pecio, L.; Oleszek, W. Phenolic acid concentrations in organically and conventionally cultivated spring and winter wheat. J. Sci. Food Agric. 2011, 91, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemicals and antioxidant activity of milled fractions of different wheat varieties. J. Agric. Food Chem. 2005, 53, 2297–2306. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Tsao, R.; Yang, R.; Cui, S.W. Phenolic acid profiles and antioxidant activities of wheat bran extracts and the effect of hydrolysis conditions. Food Chem. 2006, 95, 466–473. [Google Scholar] [CrossRef]
- Back, W.; Diener, C.; Sacher, B. Hefeweizenbier—Taste spectrum and technology. Brauwelt Int. 2000, 18, 112–119. [Google Scholar]
- Kowalczyk, D.; Świeca, M.; Cichocka, J.; Gawlik-Dziki, U. The phenolic content and antioxidant activity of the aqueous and hydroalcoholic extracts of hops and their pellets. J. Inst. Brew. 2013, 119, 103–110. [Google Scholar] [CrossRef]
- Kellner, V.; Jurková, M.; Čulík, J.; Horák, T.; Čejka, P. Some phenolic compounds in Czech hops and beer of Pilsner type. Brew. Sci. 2007, 60, 32–37. [Google Scholar]
- Pascoe, H.M.; Ames, J.M.; Chandra, S. Critical stages of the brewing process for changes in antioxidant activity and levels of phenolic compounds in ale. J. Am. Soc. Brew. Chem. 2003, 61, 203–209. [Google Scholar]
- Proestos, C.; Komaitis, M. Antioxidant capacity of hops. Beer Health Dis. Prev. 2009, 467–475. [Google Scholar]
- Pasteur, L.; Robb, D.C.; Faulkner, F. Studies on Fermentation; the Diseases of Beer, Their Causes, and the Means of Preventing Them; A translation, Made with the Author’s Sanction, of Études sur la bière; with Notes, Index, and Original Illustrations; MacMillan: Basingstoke, UK, 1879. [Google Scholar]
- Flanders Red Ales. Available online: http://www.milkthefunk.com/wiki/Flanders_Red_Ale (accessed on 18 January 2018).
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The microbial diversity of an industrially produced lambic beer shares members of a traditionally produced one and reveals a core microbiota for lambic beer fermentation. Food Microbiol. 2015, 49, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Snauwaert, I.; Roels, S.P.; Van Nieuwerburg, F.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. Microbial diversity and metabolite composition of Belgian red-brown acidic ales. Int. J. Food Microbiol. 2016, 221, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Martens, H.; Iserentant, D.; Verachtert, H. Microbiological aspects of a mixed yeast-bacterial fermentation in the production of a special Belgian acidic ale. J. Inst. Brew. 1997, 103, 85–91. [Google Scholar] [CrossRef]
- Van Oevelen, D.; Spaepen, M.; Timmermans, P.; Verachtert, H. Microbial aspects of spontaneous fermentation in production of lambic and gueuze. J. Inst. Brew. 1977, 83, 356–360. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Bamforth, C.W.; Mills, D.A. Brewhouse-resident microbiota are responsible for multi-stage fermentation of American coolship ale. PLoS ONE 2012, 7, e35507. [Google Scholar] [CrossRef] [PubMed]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Daniel, H.-M.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The microbial diversity of traditional spontaneously fermented lambic beer. PLoS ONE 2014, 9, e95384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, W.; Xu, Y.; Yu, A. Influence of oak chips geographical origin, toast level, dosage and aging time on volatile compounds of apple cider. J. Inst. Brew. 2006, 112, 255–263. [Google Scholar] [CrossRef]
- Wyler, P.; Angeloni, L.H.P.; Alcarde, A.R.; da Cruz, S.H. Effect of oak wood on the quality of beer. J. Inst. Brew. 2015, 121, 62–69. [Google Scholar] [CrossRef]
- Cadahía, E.; Fernández de Simón, B.; Jalocha, J. Volatile compounds in Spanish, French, and American oak woods after natural seasoning and toasting. J. Agric. Food Chem. 2003, 51, 5923–5932. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.P.; Howell, G.S.; Michaelis, C.S.; Dickmann, D.I. The content of phenolic acid and aldehyde flavor components of white oak as affected by site and species. Am. J. Enol. Vitic. 1992, 43, 333–338. [Google Scholar]
- Sara, C.; Concei, L.M.; Isabel, S.M.; Pedro, B.A. Influence of botanical species and geographical origin on the content of low molecular weight phenolic compounds of woods used in Portuguese cooperage. Holzforschung 2005, 54, 255–261. [Google Scholar]
- Garcia, R.; Soares, B.; Dias, C.B.; Freitas, A.M.C.; Cabrita, M.J. Phenolic and furanic compounds of Portuguese chestnut and French, American and Portuguese oak wood chips. Eur. Food Res. Technol. 2012, 235, 457–467. [Google Scholar] [CrossRef]
- Meilgaard, M.C. Flavor chemistry of beer: Part II: Flavour and threshold of 239 aroma volatiles. MBAA Tech. Q. 1975, 3, 151–168. [Google Scholar]
- Vanbeneden, N.; Gils, F.; Delvaux, F.; Delvaux, F.R. Formation of 4-vinyl and 4-ethyl derivatives from hydroxycinnamic acids: Occurrence of volatile phenolic flavour compounds in beer and distribution of Pad1-activity among brewing yeasts. Food Chem. 2008, 107, 221–230. [Google Scholar] [CrossRef]
- Spitaels, F.; Kerrebroeck, S.V.; Wieme, A.D.; Snauwaert, I.; Aerts, M.; Landschoot, A.V.; De Vuyst, L.; Vandamme, P. Microbiota and metabolites of aged bottled gueuze beers converge to the same composition. Food Microbiol. 2015, 47, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Barthelmebs, L.; Divies, C.; Cavin, J.M. Knockout of the p-coumarate decarboxylase gene from Lactobacillus plantarum reveals the existence of two other inducible enzymatic activities involved in phenolic acid metabolism. Appl. Environ. Microbiol. 2000, 66, 3368–3375. [Google Scholar] [CrossRef] [PubMed]
- Bhuiya, M.W.; Lee, S.G.; Jez, J.M.; Yu, O. Structure and mechanism of ferulic acid decarboxylase (FDC1) from Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2015, 81, 4216–4223. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Ferguson, K.L.; Boyer, D.R.; Lin, X.N.; Marsh, E.N.G. Isofunctional enzymes PAD1 and UbiX catalyze formation of a novel cofactor required by ferulic acid decarboxylase and 4-hydroxy-3-polyprenylbenzoic acid decarboxylase. ACS Chem. Biol. 2015, 10, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Lind, M.E.S.; Himo, F. Theoretical study of the reaction mechanism of phenolic acid decarboxylase. FEBS J. 2015, 282, 4703–4713. [Google Scholar] [CrossRef] [PubMed]
- Mukai, N.; Masaki, K.; Fujii, T.; Kawamukai, M.; Iefuji, H. PAD1 and FDC1 are essential for the decarboxylation of phenylacrylic acids in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2010, 109, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and divergence of Saccharomyces cerevisiae beer yeasts. Cell 2016, 166, 1397–1410. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.; Angulo, I.; de las Rivas, B.; Campillo, N.; Páez, J.A.; Muñoz, R.; Mancheño, J.M. p-Coumaric acid decarboxylase from Lactobacillus plantarum: Structural insights into the active site and decarboxylation catalytic mechanism. Proteins: Struct. Funct. Bioinform. 2010, 78, 1662–1676. [Google Scholar] [CrossRef] [Green Version]
- Barthelmebs, L.; Diviès, C.; Cavin, J.F. Expression in Escherichia coli of native and chimeric phenolic acid decarboxylases with modified enzymatic activities and method for screening recombinant E. coli strains expressing these enzymes. Appl. Environ. Microbiol. 2001, 67, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Rodríguez, H.; Curiel, J.A.; de las Rivas, B.; Mancheño, J.M.; Muñoz, R. Gene cloning, expression, and characterization of phenolic acid decarboxylase from Lactobacillus brevis RM84. J. Ind. Microbiol. Biotechnol. 2010, 37, 617–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annemüller, G. Die Berliner Weiße, 1st ed.; VLB: Berlin, Germany, 2008. [Google Scholar]
- Van Oevelen, D.; Delescaille, F.; Verachtert, H. Synthesis of aroma compounds during spontaneous fermentation of lambic and gueuze. J. Inst. Brew. 1976, 82, 322–326. [Google Scholar] [CrossRef]
- Spaepen, M.; Van Oevelen, D.; Verachtert, H. Fatty acids and esters produced during spontaneous fermentation of lambic and gueuze. J. Inst. Brew. 1978, 84, 278–282. [Google Scholar] [CrossRef]
- Crauwels, S.; Steensels, J.; Aerts, G.; Willems, K.; Verstrepen, K.; Lievens, B. Brettanomyces bruxellensis, essential contributor in spontaneous beer fermentations providing novel opportunities for the brewing industry. Brew. Sci. 2015, 68, 110–121. [Google Scholar]
- Conterno, L.; Joseph, C.M.L.; Arvik, T.J.; Henick-Kling, T.; Bisson, L.F. Genetic and physiological characterization of Brettanomyces bruxellensis strains isolated from wines. Am. J. Enol. Vitic. 2006, 57, 139–147. [Google Scholar]
- Lentz, M.; Harris, C. Analysis of growth inhibition and metabolism of hydroxycinnamic acids by brewing and spoilage strains of Brettanomyces yeast. Foods 2015, 4, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Crauwels, S.; Zhu, B.; Steensels, J.; Busschaert, P.; De Samblanx, G.; Marchal, K.; Willems, K.A.; Verstrepen, K.J.; Lievens, B. Assessing genetic diversity among Brettanomyces yeasts by DNA fingerprinting and whole-genome sequencing. Appl. Environ. Biol. 2014, 80, 4398–4413. [Google Scholar] [CrossRef] [PubMed]
- Crauwels, S.; Van Opstaele, F.; Jaskula-Goiris, B.; Steensels, J.; Verreth, C.; Bosmans, L.; Paulussen, C.; Herrera-Malaver, B.; de Jonge, R.; De Clippeleer, J.; et al. Fermentation assays reveal differences in sugar and (off-)flavor metabolism across different Brettanomyces bruxellensis strains. FEMS Yeast Res. 2017, 17, fow105. [Google Scholar] [CrossRef] [PubMed]
- Crauwels, S.; Van Assche, A.; de Jonge, R.; Borneman, A.R.; Verreth, C.; Troels, P.; De Samblanx, G.; Marchal, K.; Van, d.P.Y.; Willems, K.A.; et al. Comparative phenomics and targeted use of genomics reveals variation in carbon and nitrogen assimilation among different Brettanomyces bruxellensis strains. Appl. Microbiol. Biotechnol. 2015, 99, 9123–9134. [Google Scholar] [CrossRef] [PubMed]
- Godoy, L.; Garrido, D.; Martinez, C.; Saaverda, J.; Combina, M.; Ganga, M.A. Study of the coumarate decarboxylase and vinylphenol reductase activities of Dekkera bruxellensis (anamorph Brettanomyces bruxellensis) isolates. Lett. Appl. Microbiol. 2009, 48, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Vigentini, I.; Romano, A.; Compagno, C.; Merico, A.; Molinari, F.; Tirelli, A.; Foschino, R.; Volonterio, G. Physiological and oenological traits of different Dekkera/Brettanomyces bruxellensis strains under wine-model conditions. FEMS Yeast Res. 2008, 8, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Chatonnet, P.; Dubourdieu, D.; Boidron, J.N.; Pons, M. The origin of ethylphenols in wines. J. Sci. Food Agric. 1992, 60, 165–178. [Google Scholar] [CrossRef]
- Romano, D.; Valdetara, F.; Zambelli, P.; Galafassi, S.; De Vitis, V.; Molinari, F.; Compagno, C.; Foschino, R.; Vigentini, I. Cloning the putative gene of vinyl phenol reductase of Dekkera bruxellensis in Saccharomyces cerevisiae. Food Microbiol. 2017, 63, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.; Dias, S.; Sancho, T.; Stender, H.; Querol, A.; Malfeito-Ferreira, M.; Loureiro, V. Identification of yeasts isolated from wine-related environments and capable of producing 4-ethylphenol. Food Microbiol. 2003, 20, 567–574. [Google Scholar] [CrossRef]
- Sangorrín, M.P.; García, V.; Lopes, C.A.; Sáez, J.S.; Martínez, C.; Ganga, M.A. Molecular and physiological comparison of spoilage wine yeasts. J. Appl. Microbiol. 2013, 114, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Piazzon, A.; Forte, M.; Nardini, M. Characterization of phenolics content and antioxidant activity of different beer types. J. Agric. Food Chem. 2010, 58, 10677–10683. [Google Scholar] [CrossRef] [PubMed]
- Achilli, G.; Piero Cellerino, G.; Gamache, P.H.; Vico Melzi d’Eril, G. Identification and determination of phenolic constituents in natural beverages and plant extracts by means of a coulometric electrode array system. J. Chromatogr. A 1993, 632, 111–117. [Google Scholar] [CrossRef]
- Floridi, S.; Montanari, L.; Marconi, O.; Fantozzi, P. Determination of free phenolic acids in wort and beer by coulometric array detection. J. Agric. Food Chem. 2003, 51, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Montanari, L.; Perretti, G.; Natella, F.; Guidi, A.; Fantozzi, P. Organic and phenolic acids in beer. LWT—Food Sci. Technol. 1999, 32, 535–539. [Google Scholar] [CrossRef]
- Heresztyn, T. Metabolism of volatile phenolic compounds from hydroxycinnamic acids by Brettanomyces yeast. Arch. Microbiol. 1986, 146, 96–98. [Google Scholar] [CrossRef]
- Oelofse, A.; Lonvaud-Funel, A.; du Toit, M. Molecular identification of Brettanomyces bruxellensis strains isolated from red wines and volatile phenol production. Food Microbiol. 2009, 26, 377–385. [Google Scholar] [CrossRef] [PubMed]
- The Good Scents Company. Available online: http://www.thegoodscentscompany.com/ (accessed on 5 December 2017).
- Flavor and Extract Manufacturers Association of the United States. Available online: https://www.femaflavor.org/about (accessed on 5 December 2017).
- Wasserman, A.E. Organoleptic evaluation of three phenols present in wood smoke. J. Food Sci. 1966, 31, 1005–1010. [Google Scholar] [CrossRef]
- Vanbeneden, N.; Saison, D.; Delvaux, F.; Delvaux, F.R. Decrease of 4-vinylguaiacol during beer aging and formation of apocynol and vanillin in beer. J. Agric. Food Chem. 2008, 56, 11983–11988. [Google Scholar] [CrossRef] [PubMed]
- Wackerbauer, K.; Kramer, P. Bavarian wheat beer—An alternative. Production and composition. Brauwelt 1982, 122, 758–760. [Google Scholar]
- Langos, D.; Granvogl, M.; Schieberle, P. Characterization of the key aroma compounds in two Bavarian wheat beers by means of the sensomics approach. J. Agric. Food Chem. 2013, 61, 11303–11311. [Google Scholar] [CrossRef] [PubMed]
- Langos, D.; Granvogl, M. Studies on the simultaneous formation of aroma-active and toxicologically relevant vinyl aromatics from free phenolic acids during wheat beer brewing. J. Agric. Food Chem. 2016, 64, 2325–2332. [Google Scholar] [CrossRef] [PubMed]
- Narziss, L.; Miedaner, H.; Neizsche, F. The formation of 4-vinylguaiacol during the manufacture of Bavarian wheat beers (weizenbier). Monatsschrift Fuer Brauwissenschaft 1990, 43, 96–100. [Google Scholar]
- Kieninger, H.; Narziss, L.; Miedaner, H.; Hecht, S. Concerning the changes of quality determining molecular groups in the preparation of Bavarian wheat beers. Brauwissenschaft 1984, 37, 9–16. [Google Scholar]
- Schwarz, K.J.; Boitz, L.I.; Methner, F.-J. Release of phenolic acids and amino acids during mashing dependent on temperature, pH, time, and raw materials. J. Am. Soc. Brew. Chem. 2012, 70, 290–295. [Google Scholar] [CrossRef]
- Schwarz, K.J.; Stübner, R.; Methner, F.-J. Formation of styrene dependent on fermentation management during wheat beer production. Food Chem. 2012, 134, 2121–2125. [Google Scholar] [CrossRef] [PubMed]
- White, C.; Zainasheff, J. Yeast: The Practical Guide to Beer Fermentation; Brewers: Boulder, CO, USA, 2010. [Google Scholar]
- Mertens, S.; Steensels, J.; Gallone, B.; Souffriau, B.; Malcorps, P.; Verstrepen, K.J. Rapid screening method for phenolic off-flavor (POF) production in yeast. Am. Soc. Brew. Chem. 2017, 75, 318–323. [Google Scholar] [CrossRef]
- 2015 Beer Style Guidelines. Available online: https://www.bjcp.org/docs/2015_Guidelines_Beer.pdf (accessed on 22 January 2018).
- Competition Style List, Descriptions, and Specifications. 2017. Available online: https://www.greatamericanbeerfestival.com/wp-content/uploads/2017/05/17_GABF_Beer_Style_Guidelines_Final.pdf (accessed on 22 January 2018).
- Competition Style List, Descriptions, and Specifications. 2018. Available online: https://www.worldbeercup.org/wp-content/uploads/2013/09/18_WBC_Style_Guidelines_Final.pdf (accessed on 22 January 2018).
- Yeast Bank. Available online: https://www.whitelabs.com/yeast-bank (accessed on 22 January 2018).
- Yeast Strains for Beer. Available online: http://www.wyeastlab.com/beer-strains (accessed on 22 January 2018).
- Yeast Descriptions Guide. Available online: http://www.brewingscience.com/PDF/prodlist/BSI_Yeast_Descriptions_Guide.pdf (accessed on 18 January 2018).
- Jespersen, L.; Jakobsen, M. Specific spoilage organisms in breweries and laboratory media for their detection. Int. J. Food Microbiol. 1996, 33, 139–155. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Hiralal, L.; Mokoena, M.P.; Pillay, B. Flavour-active volatile compounds in beer: Production, regulation and control. J. Inst. Brew. 2017, 123, 13–23. [Google Scholar] [CrossRef]
- Lawrence, D.R. Spoilage Organisms in Beer. In Developments in Food Microbiology; Robinson, R.K., Ed.; Elsevier: London, UK, 1988; pp. 1–48. [Google Scholar]
- Shinohara, T.; Kubodera, S.; Yanagida, F. Distribution of phenolic yeasts and production of phenolic off-flavors in wine fermentation. J. Biosci. Bioeng. 2000, 90, 90–97. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411–430. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Hydroxycinnamic acid structures.
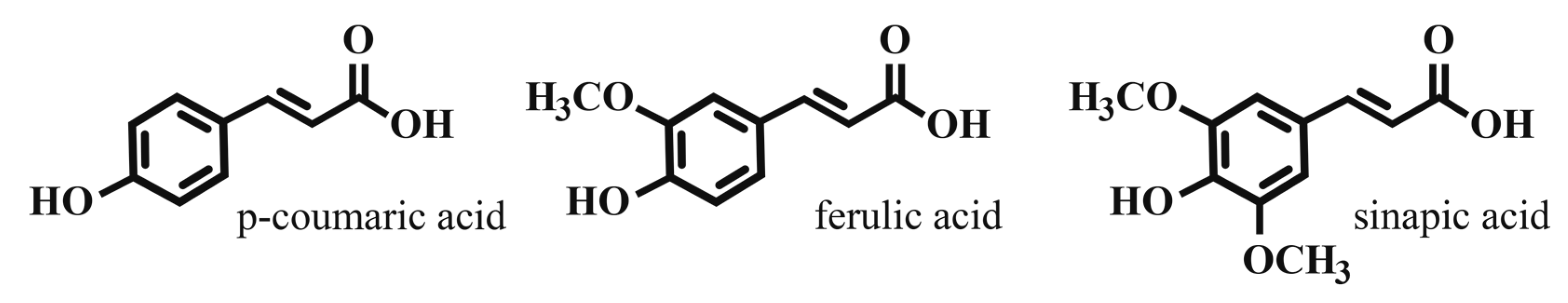
Figure 2.
Enzymatic conversion of hydroxycinnamic acid (ferulic acid). PAD, phenolic acid decarboxylase; VPR, vinylphenol reductase.
Figure 2.
Enzymatic conversion of hydroxycinnamic acid (ferulic acid). PAD, phenolic acid decarboxylase; VPR, vinylphenol reductase.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Flavor and aroma descriptors for common volatile phenols.
| Compound | Descriptors | Sources | References |
|---|---|---|---|
| 4-vinylguaiacol | Clove, curry, spice, smoky, bacon | Thermal or enzymatic decarboxylation of ferulic acid | [3,18,81] |
| 4-vinylphenol | Phenolic, medicinal, spicy | Thermal or enzymatic decarboxylation of p-coumaric acid | [18,81] |
| 4-ethylguaiacol | Clove, phenol, spice, woody, smoky, vanilla | Enzymatic reduction of 4-vinyl guaiacol | [3,71,81,82] |
| 4-ethylphenol | Leather, phenol, spice, stable, smoke, creosote | Enzymatic reduction of 4-vinyl phenol | [3,71,81,82] |
| Guaiacol | Smoke, bacon | Lignin pyrolysis | [3,83] |
| Vanillin | Sweet, vanilla | Lignin pyrolysis, wood aging, degradation of 4-vinyl guaiacol | [3,84] |
| 4-vinyl syringol | Stale, “old beer” | Degradation of synapic acid glycosides | [3,8] |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lentz, M. The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor. Fermentation 2018, 4, 20. https://doi.org/10.3390/fermentation4010020
AMA Style
Lentz M. The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor. Fermentation. 2018; 4(1):20. https://doi.org/10.3390/fermentation4010020
Chicago/Turabian StyleLentz, Michael. 2018. "The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor" Fermentation 4, no. 1: 20. https://doi.org/10.3390/fermentation4010020
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.