Enterococci Isolated from Cypriot Green Table Olives as a New Source of Technological and Probiotic Properties



Department of Agricultural Sciences, Biotechnology and Food Science, Cyprus University of Technology, Limassol 3036, Cyprus
*
Author to whom correspondence should be addressed.
Fermentation 2018, 4(2), 48; https://doi.org/10.3390/fermentation4020048
Submission received: 9 May 2018
/
Revised: 2 June 2018
/
Accepted: 8 June 2018
/
Published: 20 June 2018
(This article belongs to the Special Issue Safety and Microbiological Quality)
Abstract
:Τable olive is one of the main fermented vegetable worldwide and can be processed as treated or natural product. Lactic Acid Bacteria (LAB) are responsible for the fermentation of treated olives. The aim of this work was to study the technological characteristics and the potential probiotic properties of LAB isolated from Cypriot green table olives. This is the first comprehensive report on the isolation and characterization of LAB isolates retrieved from Cypriot green table olives. From a collection of 92 isolates from spontaneously fermenting green olives, 64 g positive isolates were firstly identified to genus level using biochemical tests, and secondly to species level using multiplex species specific polymerase chain reaction (PCR) amplifications of the sodA gene. Moreover, each of our isolates were tested for their technological and probiotics properties, as well as for their safety characteristics, using biochemical and molecular methods, in order to be used as starter cultures. Finally, to discriminate the most promising isolates on the base of their technological and probiotics properties, Principal component analysis was used. All the isolates were identified as Enteroccocus faecium, having interesting technological properties, while pathogenicity determinants were absent. Principal component analysis showed that some isolates had a combination of the tested parameters. These findings demonstrate that enteroccoci from Cypriot table olives should be considered as a new source of potential starter cultures for fermented products, having possibly promising technological and probiotic attributes.

1. Introduction
Table olive is considered the most well-known among fermented foods in the Mediterranean area, constituting an important sector for the agro-industrial economy. Olives are the fruits produced by the olive tree (Olea europea). World production reach an average of 2.3 million tons per year, while the Mediterranean region is the main producer [1]. Most table olives (60% of the total production) are prepared following the Spanish-style (alkali treated green olives). However, nowadays, producers have displayed renewed and increasing interest in Greek-style fermentation, prepared with directly brined fruits [2]. Among the microorganisms involved in the fermentation process, Lactic Acid Bacteria (LAB) is the most essential and abundantly found group [3]. The development of LAB in the fermentation process, leads to a rapid acidification of the brine and a pH drop under 4.5 units [4].
LABs are gram-positive, non-spore forming bacteria. Due to the fact that they are facultative anaerobes they use their fermentative metabolism to provide energy. In general, LAB are a genetically diverse group of bacteria, encompassing rod-shaped bacteria such as lactobacilli, and cocci like streptococci, lactococci, enterococci, pediococci, or leuconostocs [5].
Enterococci are Gram-positive, non-spore forming, catalase negative, oxidase negative cocci and they occur singly, in pairs, or in chains. From a taxonomic point of view, the genus Enterococcus has been reviewed several times [6]. The genus includes more than 20 species, with Enterococcus faecium and Enterococcus faecalis being the most prevalent species in fermented foods [7]. It has been shown that those species produce bile salt hydrolases, presenting potential probiotic properties related to a reduction of serum cholesterol levels by promoting a higher excretion of deconjugated bile salts [8]. Due to their medium and/or high acid and salt tolerance, enterococci may be used in food fermentations as starter cultures, being responsible for the formation of unique flavors, as well as the production of enterocins [9]. Moreover, enterococci seem to be involved in table olive fermentation; In previous studies, it has been reported the presence of those LAB in Spanish-style green olives [9,10]. More specifically, Randazzo et al. [11] isolated four strains of enterococci belonging to the species E. faecium, Enterococcus casseliflavus, and Enterococcus hirae from naturally fermented green olives collected from different areas of Sicily region. Furthermore, it has been proposed the use of enterococci as starter culture for olive fermentation, because they play a crucial role at the initial stage of the process, due to their tolerance to high pH and salinity [11,12].
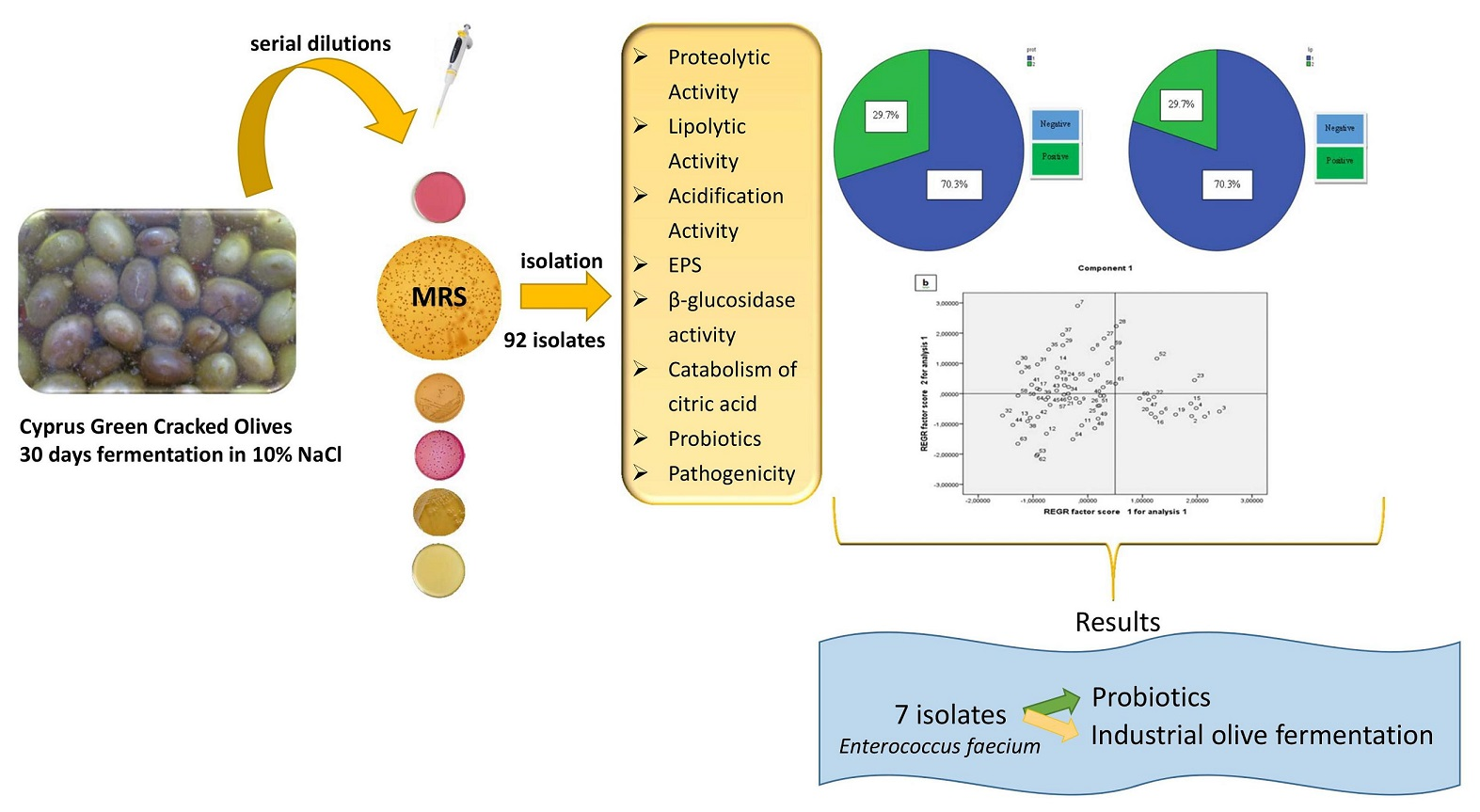
The aim of this work was to study the technological characteristics and the potential probiotic properties of LAB isolated from Cypriot green table olives, in order to discover new potential starter cultures for food fermentation industry.
2. Materials and Methods
2.1. Samples Collection
Nine samples of Cyprus variety of green cracked olives in round containers of 100 L brine (10%) fermenting for a period of one month (average temperature 25 °C), were collected from the company named “King of olives” in Agglisides, Cyprus, which is the largest producer of table olives in the country and processes olives from most production regions. This one month period was decided because in Cyprus the general practice is to consider the green cracked olives ready for consumption in 30 days of fermentation/debittering process. This is due to local market preferences of consuming green cracked olives while keeping their “freshness” or otherwise bitter taste. The brined samples were placed in sterile plastic containers and transported to the laboratory where they were stored in darkness at room temperature for analysis within 24 h. pH was measured in all nine samples by a portable pH meter (Hanna, UK).
2.2. Enumeration of Microorganisms
The samples were evaluated for their Total Viable Count (TVC), Enterobacteriaceae, LAB, yeasts, coliforms, Micrococcaceae, Gram-negative cocci and salt tolerant using the standard spread plate method after serial dilutions in saline water (0.85% (w/v)) (Table 1). More specifically, 10 g of table olives (flesh tissue) were transferred aseptically to stomacher bags with 90 mL saline solution (0.85% (w/v) NaCl) and homogenized for 2 min using a Stomacher at 220 rpm for 2 min (Bug Mixer, Interscience, Saint Nom, France). Volumes of 0.1 mL or 1 mL (spread and pour plate, respectively) of serial dilutions in saline solution, were placed in petri dishes for enumeration of the microorganisms described. All samples were analyzed in triplicates.
2.3. Isolation of LAB
Ten to 12 representative colonies according to their different morphological characteristics (size, shape, color etc.) were retrieved from De Man-Rogosa-Sharpe (MRS) agar Petri dishes with colony counts at the range of 30–300. Purity of the isolates was checked by streaking twice on MRS agar, followed by microscopic examination. Finally, stock cultures of purified isolates were stored at −80 °C in 20% (v/v) glycerol/MRS broth.
2.4. Physiological and Molecular Characterization of LAB Strains
The purified isolates were examined for Gram staining and catalase [13]. Gram-positive, catalase negative cocci were presumptively identified as LAB. Further classification was applied according to the biochemical criteria, such as growth at various temperatures (10 °C, 15 °C, and 45 °C), salt tolerance (2, 4, 6.5, 8% (w/v) NaCl), production of CO2 from glucose as unique source of carbon and pH (9.6) [14,15].
The identification of the isolates at species level was achieved by applying molecular techniques by multiplex PCR amplifications of the sodA gene [16]. Three PCR master mixes consisting of different primer sets for each species were prepared (Table 2); Group 1 was Enterococcus durans, E. faecalis, E. faecium; group 2 was E. casseliflavus, Enterococcus gallinarum; group 3 was E. hirae. PCR reactions contained 2.5 μL template DNA (10 ng), 1× PCR reaction buffer, 0.2 mM of each deoxynucleotide triphosphate (dNTP), 3 mМ MgCl2, 16 μM of each primer, 2.5 U of Kappa Hot Start DNA polymerase (KAPPA Biosystems), and distilled water was added to a final volume of 22.5 μL. Following an initial denaturation at 95 °C for 4 min, products were amplified by 35 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for Group 1, 57 °C for Group 2 and 60 °C for Group 3 for 1 min, and elongation at 72 °C for 1 min. Finally, amplification was followed by a final extension at 72 °C for 7 min. Eight μL of PCR product was electrophorized on a 1.2% agarose gel, 1× TAE (Tris-acetate-EDTA), stained with SYBR™ Safe DNA Gel Stain (Invitrogen, Carlsbad, CA, USA) and visualized under UV light. A 100 bp DNA ladder (Nippon Genetics) was used as marker.
2.5. Technological Characteristics
2.5.1. Acidification Activity
Acidification activity of the Enterococcus isolates was tested according to Fuka et al. [17]. In particular, tubes containing 10 mL of sterile skimmed milk (D 10% (w/v); Oxoid) were inoculated with fresh overnight cultures (1% (v/v)) and incubated at 37 °C. The pH value was determined after incubation for 0, 6 and 24 h using a pH meter (Hanna instruments). The analysis was carried out in triplicate. A non-inoculated skim milk was used as negative control. The acidification rate was calculated as ΔpH = pH (0h) − pH (6 h or 24 h respectively).
2.5.2. Proteolytic Activity
For the determination of proteolytic activity, 2 μL of fresh overnight cultures (108 cfu/mL) were spotted on the surface of a skim milk agar (10% (w/v) skim milk, 2% (w/v) agar, Oxoid), and were incubated at 37 °C from 2 to 4 days [18]. The results were considered as positive when a clear zone was formed around the colonies. The analysis was performed in triplicate.
2.5.3. Lipolytic Activity
For the determination of lipolytic activity, a loopful of fresh culture (24 h) was placed on tributyrin agar (Oxoid) and was incubated at 37 °C for 4 days [19]. The results were considered as positive when a clear zone appeared around the colonies. The analysis was repeated three times.
2.5.4. Exopolysaccharide Production (EPS)
EPS production was evaluated with ruthenium red staining method in ruthenium milk agar (0.5% yeast extract, 10% skim milk powder, 1% sucrose, 1.6% agar, 0.08 g/L ruthenium red) [20]. The coagulated cultures were considered as EPS-positive when a white loop was formed after incubation at 37 °C for 24 h. The analysis was conducted in triplicate.
2.5.5. β-Glucosidase Activity
β-glucosidase activity was determined using X-Gluc-5-bromo-4-chloro-3-indolyl-β-d-glucopyranoside (Sigma, Saint Louis, MO, USA) as substrate. The test was conducted by spreading 0.2 mL of a N,N-dimethyl formamide solution to a final concentration of 0.3% (w/v) X-Gluc on MRS agar. The plates were left to dry for 3 h in darkness, and were incubated at 37 °C for 48 h after the inoculation (2 μL) [21]. Isolates producing β-glucosidase were recognized when the colony was colored blue. The analysis was repeated three times.
2.5.6. Catabolism of Citric Acid
The determination of catabolism of citric acid was performed by growth in Simmons Citrate Agar (Oxoid), which contains citric acid as the only carbon source. Freshly prepared cultures were inoculated by spread to the medium and the plates were incubated at 37 °C for 7 days. Results were considered positive when colonies turned the color of the substrate from green to blue, otherwise the test was considered as negative. The analysis was performed in triplicates.
2.6. Pathogenicity
2.6.1. Hemolytic Activity
Freshly prepared cultures were inoculated by spread on Columbia Agar Base (Oxoid), supplemented with 5% (v/v) horse blood and incubated at 37 °C for 24 h. Hemolytic activity was characterized “α” when a green zone was formed round the colonies, “β” when clear zones were created around colonies, and “γ” when no zones were appeared around colonies [17]. The analysis was repeated in triplicates.
2.6.2. DNAse Production
Screening for DNAse production was held according to Ribeiro et al. [22]. The tested strains were grown at 37 °C in MRS broth overnight. A loopful of freshly prepared cultures was spotted on DNAse agar (Oxoid) and a few drops of 1 N HCl were added onto the colony, after incubation for 24 h at 37 °C. Production of DNAse was indicated when a clear zone appeared around the colonies. The analysis was repeated three times.
2.6.3. Virulence Activity Using Genotypic Tests
All the selected strains were checked for the presence of genes encoding virulence, antibiotic resistance and amino acid decarboxylase activity [23,24]. DNA was extracted using Bacterial DNA kit (Macherey-Nagel, Duren, Germany) according to the manufacturer’s instructions, following by amplifications of genes described in Table 3 using specific primers. PCR reactions contained 1 μL template DNA (10 ng), 1× PCR reaction buffer, 0.2 mM of each dNTP, 1.5 mM MgCl2, 16 μМ of each primer, 1 U of Kappa Taq DNA polymerase (KAPPA Biosystems), and distilled water was added to a final volume of 20 μL. Following an initial denaturation at 95 °C for 4 min, products were amplified by 30 cycles of denaturation at 95 °C for 30 s, annealing of each primer for 30 s, and elongation at 72 °C for 1 min. Finally, amplification was followed by a final extension at 72 °C for 7 min. Eight μL of PCR product was electrophorized on a 1.2% agarose gel, 1× TAE (Tris-acetate-EDTA), stained with SYBR™ Safe DNA Gel Stain (Invitrogen, Carlsbad, CA, USA) and visualized under UV light. A 100 bp DNA ladder (Nippon Genetics) was used as marker. The DNA of the reference strain E. faecium ATCC 29212 was used as positive control in the corresponding PCR reactions.
2.7. Screening for Probiotics Characteristics
2.7.1. Resistance to Low pH
The survival of cultures on MRS broth buffered at four different pH values (6, 4, 3, and 2) was studied. The isolates were inoculated (1% (v/v)) in MRS broth with 4 different pH values described above and incubated at 37 °C for 48 h. Furthermore, 1ml of sample was taken at 0, 1 and 3 h of incubation and following appropriate dilutions, was inoculated into MRS agar. Finally, after the incubation at 37 °C for 48 h the forming colonies were counted. The survival ratio of the 1st and the 3rd hour was calculated using the log of the surviving cells (cfu/mL) in 1 and 3 h, divided by the log of the starting cells (cfu/mL) at 0 h [28]. The analysis was repeated three individual times.
2.7.2. Resistance to Bile Salts
Resistance to MRS broth containing 0.3% of bile acids (Sigma-Aldrich, Saint Louis, MO, USA) was tested as follows. Firstly, the samples (1% v/v)) of 0 and 3 h were received from a water bath (37 °C), following by appropriate dilutions and finally inoculated into MRS agar. Colony counting took place after 48 h of incubation at 37 °C. The survival ratio was calculated using the log of the surviving cells (cfu/mL) in 3 h divided by the log of the original cells (cfu/mL) at 0 h [29]. The analysis was repeated three individual times.
2.8. Statistical Analysis
The results were expressed as the mean and standard deviation with triplicate determinations, followed by frequencies distribution. Significant differences for microbiological and technological analyses between samples were determined using the statistical software MINITAB 12.0 by the method of one way analysis of variance (ANOVA).
Principal component analysis (PCA) was used to discriminate all isolates on the base of their technological and probiotics properties, while the discriminating variables were proteolytic activity, lipolytic activity, acidification activity, survival to pH 2 at 1 h, survival to pH 3 at 1 h, survival to pH 3 at 2 h, survival to bile salts (0.3%). Factors with eigenvalues higher than 1.00 were retained according to the Kaiser criterion [29]. In the case of not numeric values (proteolytic and lypolitic activity), before the use of PCA, it was used the method of Optimal Scaling, so our factorial points of the objects obtain new, complex, quantitative variables, which are linearly independent. PCA was performed using the statistical software SPSS v.20.
3. Results and Discussion
3.1. Microbial Enumeration
The results of total aerobic bacteria, Enterobacteriaceae, LAB on MRS and M17 agar, yeasts and molds, coliforms, Micrococcaceae, and Gram negative cocci of the fermented green cracked Cypriot table olives are summarized in Table 4. As shown in Table 4, LAB were the predominant microorganisms, having an average value of 8.04 ± 0.04 log10cfu/g. However, yeasts and molds counts were counted at lower levels (3.48 ± 0.06 log10cfu/g). In accordance with our results, in other study it was found that LAB from Spanish style fermented “Bella di Cerignola” table olives were the predominant microorganisms, having a population of 6.74 ± 0.16 log10cfu/g, at the end of fermentation process, while yeasts population was limited [30]. Another study reports that in natural processing of olives the predominant organisms are yeasts contrary to Lactic Acid Bacteria dominating the fermentation of Spanish style treated olives [31]. It is worthwhile to mention that in the present study, the population of Enterobacteriaceae and coliforms was found at quite high levels (4.66 ± 0.02 log10cfu/g and 3.83 ± 0.01 log10cfu/g, respectively), despite the low pH values (3.89 ± 0.01). Enterobacteriaceae can be found at the beginning of the fermentation and are quickly inhibited by pH decrease due to LAB activity which produce lactic acid as well as bacteriocins responsible of the safety of fermented products [32]. A possible justification for these high numbers of Enterobacteriaceae could be the short period of time (30 days) of fermentation not affecting their counts. Reports from other studies show that Enterobacteriaceae disappear in 30–40 days [32].
3.2. Isolation and Identification of LAB
Ninety two colonies were randomly picked from MRS plates containing 30–300 colonies. MRS was chosen as the source of isolates because it showed the highest populations. Sixty-four Gram positive isolates were considered as LAB due to the fact that, they were cocci and catalase negative, and thus it seems that they belong to the genus of Enterococcus, according to the phenotypic identification (growth to all NaCl concentrations, pH and temperatures, negative to heterofermentation). Molecular identification via multiplex species specific PCR showed that all 64 isolates were E. faecium.
Enterococci were isolated from Spanish-style green olive fermentations [33]. Isolates have been identified, characterized, and utilized with Lactobacillus pentosus as starter cultures for Spanish-style green olive fermentation. In another study [10], three strains of Enterococci were isolated from olives, belonged to the species E. faecium. Furthermore, a total number of 52 LAB have been isolated from naturally fermented green olives collected from different areas of Sicily [11]. Even though the majority of these strains belonged to the genus of Lactobacillus, some of them were identified as Enterococcus spp., as well.
3.3. Technological Properties
3.3.1. Acidification Activity
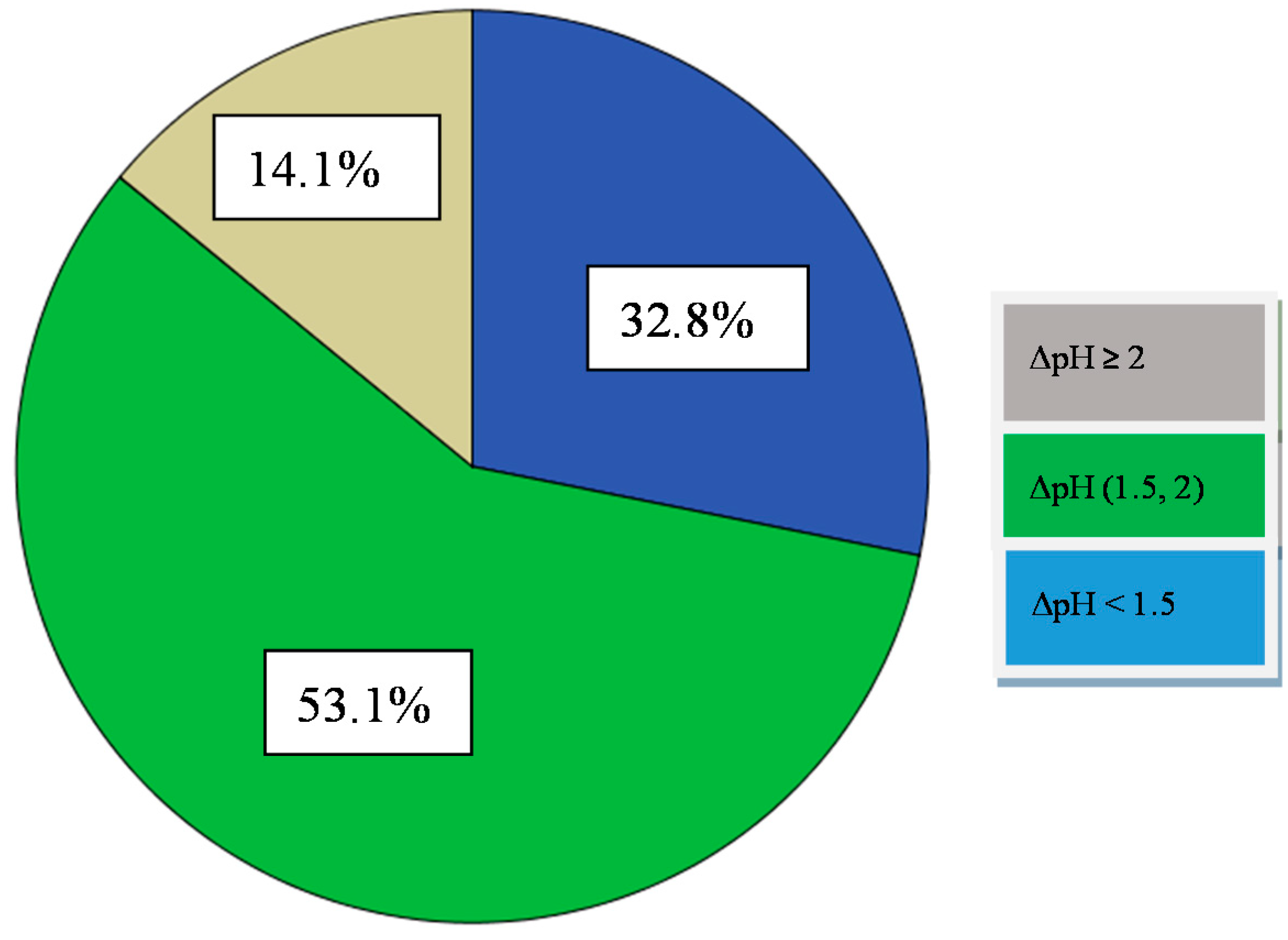
A good mesophilic fast acid producing starter culture will reduce the pH of the milk at 5.3 after 6 h of incubation at 37 °C [24]. Of the Enterococci of the present study, none of them reduced the milk pH to <6.0 after 6 h of incubation (Figure 1, Table 5). However, after 24 h of incubation, nine out of 64 isolates reduced milk pH levels to >4.60, 34 of 64 from 5.10 to 4.60, while 21 of them kept the pH value to <5.10. Thus, those isolates could be considered as slow or medium acidifiers [34], and could be classified in three main groups: (i) those showing a high acidifying capacity, with a pH decrease of 2 pH units (14.1%) and more, (ii) those with an intermediate acidifying activity showing a pH decrease ranging from 1.5 to 2.0 pH units (53.1% of the isolates), and (iii) those with low acidifying capacity (32.8% of the isolates) showing a decrease in pH value less than 1.5 pH units (Figure 1, Table 5). It is known that the rate of acid production is a crucial factor when it comes to select a proper starter culture. Besides, pH prevents the growth of undesirable microorganisms such as spoilage and pathogenic bacteria, is also responsible for the organoleptic properties of the final product. Τhe high capacity acidifiers (14.1%) identified, could be used as starter cultures for the production of fermented table olives because they could reduce the pH value and metabolize lactose to lactic acid. Slow or medium acidifiers (85.9%), could also be used as starter cultures in combination with the high capacity acidifiers because they have other interesting technological characteristics.
According to literature, considerable work has been done on acid production of Enterococcus species. In general, enterococci show low or medium acidifying ability, a fact that agrees with the present study [22]. Particularly, in another work, it has been reported that the pH value of milk did not fall below 5.5 pH units after 24 h inoculated with E. faecium [35]. Some other studies on enterococci confirmed the poor acidifying activity of these microorganisms in milk, giving a small percentage of strains showing a pH below 5.0–5.2 after 16–24 h of incubation at 37 °C [28,30,31]. It has been also shown that E. faecalis has higher acidification activity than E. faecium in general [36].
3.3.2. Proteolytic Activity
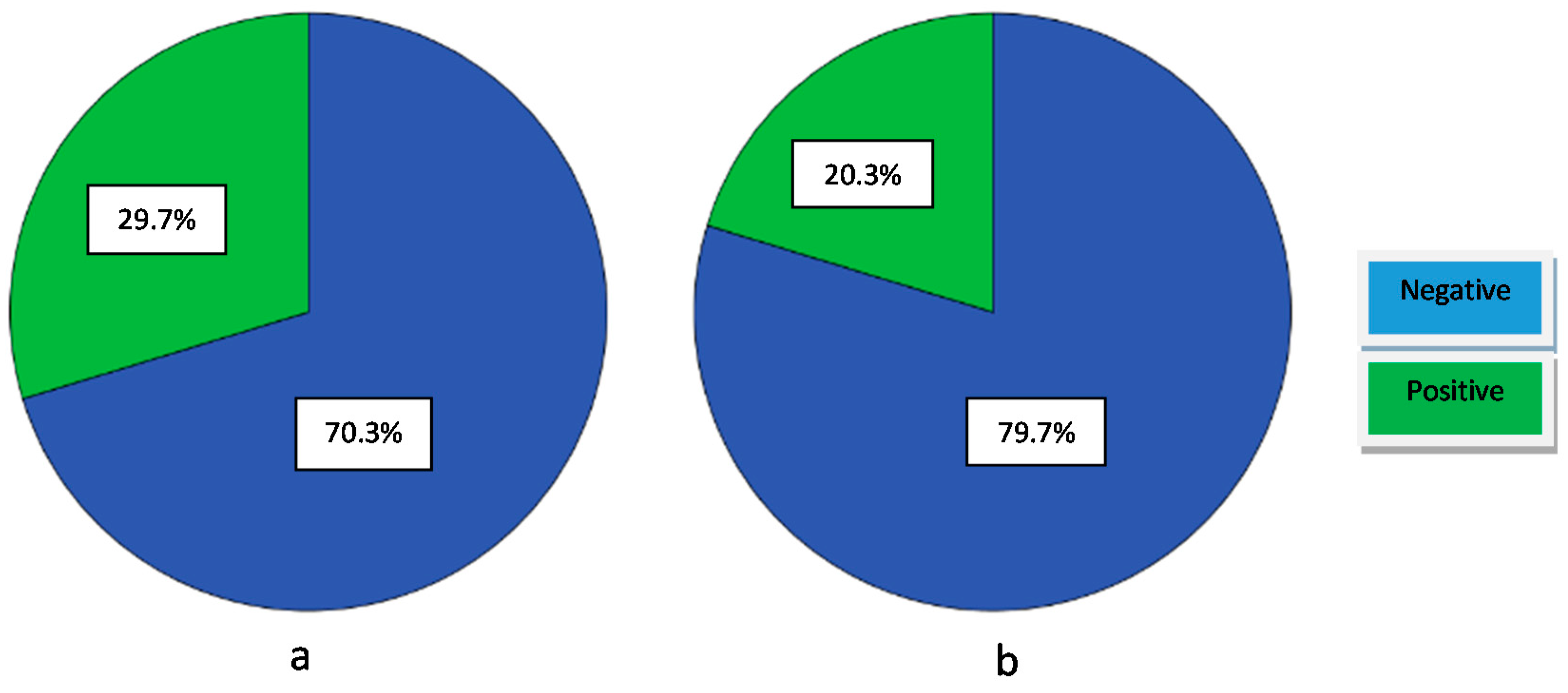
Proteolysis is one of the main industrial phenomena that contribute to the development of the organoleptic characteristics of a fermented product. The proteolytic activity of the isolates was recorded by the presence of a clear halo (positive result) on 10% Skim Milk agar. Nineteen out of 64 strains (29.7%) showed proteolytic activity. According to the literature, it has been reported that between seven strains of Enterococcus, only one was able to degrade casein, being positive for proteolytic activity [22]. Our isolates have less proteolytic activity (Figure 2a, Table 5) in comparison with other Enterococcus strains that have been characterized in other studies. In particular, according to other studies, enterococci were characterized having weak proteinase activity [33,36]. The same conclusion was drawn in another systematic study [34], in which 129 E. faecium, E. faecalis, and E. durans strains were screened for their technological characteristics. It was found that all strains showed low extracellular proteolytic activity, with the E. faecalis strains being more active. Generally, there are few data regarding the proteolytic system of enterococci in comparison with other LAB species. The proteolytic activity is low in Enterococcus strains, except of E. faecalis strains [30,32]. It is worthwhile to mention, that in the present study there is no correlation between proteolytic and acidification activities, because the fact that only two strains were positive to proteolytic activity, were able to reduce pH value to more than two units, as well. This finding agrees with those of a study performed by other researchers [37], suggesting that no clear relationship was observed between proteolytic and acidification activities. However, according to other report, it has been observed that the majority of the acidifying strains also had proteolytic activity [36]. Nevertheless, more and deeper physiological studies are needed in order to investigate whether or not the proteolytic ability of enterococci is one of its characteristics and how is regulated.
3.3.3. Lipolytic Activity
The lipolytic activity is a desirable characteristic of a strain to be used as a starter culture in order to highlight certain organoleptic characteristics of the final product. Limited reports exist on the lipolytic activity of enterococci, especially those isolated from table olives. Regarding the results of this study, it seems that enterococci have generally low lipolytic activity when using tributyrin agar. Namely, only 20.31% (13 out of 64) of strains were positive to lipolytic activity (Figure 2b, Table 5). Additionally, there are some studies showing low or no lipolytic activity of Enterococcus species [27,31,32]. Moreover, in another work, it has been showed that the existence of lipolytic activity was only confirmed for two out of seven Enterococcus isolates [22]. However, there is a study reported that among 129 enterococci, the majority of them (90%) hydrolyzed all tributyrin substrates [34]. Furthermore, in the same work, it has been concluded that E. faecalis strains were the most lipolytic, followed by the E. faecium and E. durans strains. All studies lead to the conclusion that lipolytic activity of enterococci, is strain depended as well as related to the type of the examined food.
3.3.4. Exopolysaccharide Production (EPS)
According to our results about EPS production, all isolates were not able to produce EPS since they were showing pink to red colonies. EPS is a protection barrier against lethal influence of the environment (desiccation, phagocytosis, phage attack, osmotic stress, antibiotics or toxic compounds) and the major component of bacterial biofilm, enhancing the colonization of probiotic bacteria in cell-host interactions in the gastrointestinal tract [38]. This is a beneficial trait for probiotics in their endeavor to colonize the gut. However, according to two other studies 26 out of 72 tested strains were able to produce EPS and seven out of 25 strains, respectively [28,35]. However, more studies are required in order to learn more about the EPS production from Enterococcus spp.
3.3.5. β-Glucosidase Activity
None of the isolates gave positive result (blue colony), so they are not able to produce the β-glucosidase enzyme. The enzyme of β-glucosidase is closely related to the oleuropein hydrolysis and the debittering process in table olives so it is a desirable characteristic for a potential starter culture [39].
3.3.6. Catabolism of Citric Acid
Citrate metabolism by LAB is essential in a wide range of fermented foods and beverages, since it serves as a precursor for the formation of plenty other compounds contributing to the final organoleptic characteristics. None of the isolates could catabolize the citric acid as unique source of carbon. The metabolism of citric acid has been extensively studied and it is well documented in several Lactococcus, Lactobacillus, and Leuconostoc species [40]. In contrast, only a few data deals with citrate metabolism by Enterococcus strains are available. According to them, strains of E. faecium have less ability to utilize citrate as unique source of carbon [34].
3.4. Pathogenicity
Pathogenicity tests (DNase production, hemolytic activity) were negative for all strains, suggesting their safety as starter cultures but the study of virulence factors by molecular methods in Enterococcus spp. coming from foods, is necessary due to the risk of genetic transfer since these genes are usually located in conjugative plasmids [23,41]. The presence of virulence factors in enterococci can greatly contribute to enhance the severity of hospital infections. The isolates were tested for the presence of genes encoding potential virulence factors and biogenic amines. Five (7.8%) of our isolates were positive for gelE gene. However, in previous studies, it has been found that strains having this gene did not produce gelatinase [24,38], because the fsr operon could be damaged, lost, or suffer deletions due to physiological stresses from laboratory storage.. Hemolysin production increases the severity of enterococcal infections, and the presence of hemolysin/cytolysin genes is considered a risk pathogenicity factor. Regarding the cylA gene, two (3.1%) isolates gave a positive result, but according to phenotypical tests, only γ-hemolysis was observed. cylA has been considered as a “silent gene” and its gene expression can be influenced by the environmental factors and conditions used for phenotypic tests [24]. The genes esp, efaA, and ace have been involved in the colonization and adhesion at biotic and non-biotic surfaces, and the host immune system evasion. The genes esp, efaA, and ace were not detected in none of the isolates. However other authors found high incidence of esp, asa1, and efaA genes in E. faecalis [39,42]. Moreover, none of the isolates gave a positive result for hyl, which is related to the production of hyaluronidase facilitating the spread of toxins and bacteria throughout the host tissue by causing tissue damage. Similarly, aggregation substance (asa1) was not detected in our isolates. This gene is a sex pheromone plasmid-encoded surface protein, which promotes the conjugative transfer of sex pheromone plasmids by formation of mating aggregates between donor and recipient cells. Finally, none of the isolates was positive for vanA and vanB genes. This is in agreement with the results from previous studies [24,43], reporting that vanA and vanB genes have not been found frequently in enterococci isolates from food sources. However, in other work, it has been reported that three strains (50%) of Enterococcus faecalis were positive to vanA gene [22]. Regarding the presence of several-amino decarboxylase genes, histidine (hdc1 and hdc2), ornithine (odc), and tyrosine (tdc) decarboxylase, no amplification occurred for either hdc1 and hdc2, or odc. However, the tdc gene was present in 11 (17.1%) isolates, which is in accordance with the literature reported that despite the fact that tyramine production is a common characteristic of enterococcal isolates it is considered to be a negative trait for their possible use in foodstuffs [21,28,44].
The safety profile of enterococci isolated from Cypriot green table olive revealed that all of them were negative to the most clinically relevant antibiotics, such as vanA and vanB genes. The van gene is transferable, making this antibiotic resistance the most important safety factor to be evaluated in food-grade enterococci [45]. It is crucial to mention that the lack of the determinants of infectivity and antibiotic resistance in our enterococcus strains raises optimism about their further application in fermented foods. It must be mentioned that the presence of some virulence genes such like gelE, which was found in some of our isolates, cannot be considered as a negative trend, since enterococci are a part of the spontaneous microbiota of table olives and these genes have been also found in commercial enterococci starter cultures with a long history of safe use [44].
3.5. Screening for Probiotic Potential
Probiotic potential is one of the main factors in the case of choosing a starter culture. Tolerance to bile salts is a prerequisite for colonization and metabolic activity of probiotic bacteria in the small intestine of the host. Results indicated that all tested isolates were resistant to bile salts since the majority of them were grown successfully in MRS broth supplemented with 0.3% bile salts representing the physiological concentration of human bile (Table 6). Moreover, the isolates exhibited high tolerance to acidic conditions, surviving in pH 4.0, 5.0, and 6.0 (100% survival), in pH 2 (after 1 h of incubation), as well as in pH 3.0 (after 1 and 3 h of incubation), having high survival rates. In addition, after 3 h of incubation at pH 2.0, none of the isolates survived. However, all of isolates could be characterized as potential probiotics. According to other study among seven selected strains, three strains of E. faecium survived at pH 3.0 after 3 h (over 85%), while the other four strains showed lower survival rates [46]. The results indicated that all tested strains (Table 7) could possibly survive through the human stomach and might possess the ability to reach the intestinal environment in which they may effectively work. In the same work all tested strains survived in the presence of 1% bile salts over the rate 85%. Based on these results, E. faecium strains have the prerequisites to survive in the gastrointestinal tract. Finally, other researchers revealed that Enterococcus spp. strains from different sources could have the ability to reach the intestinal lumen and stay alive in that environment [28,47] However, further in vitro studies are needed with these strains in order to establish their probiotic potential.
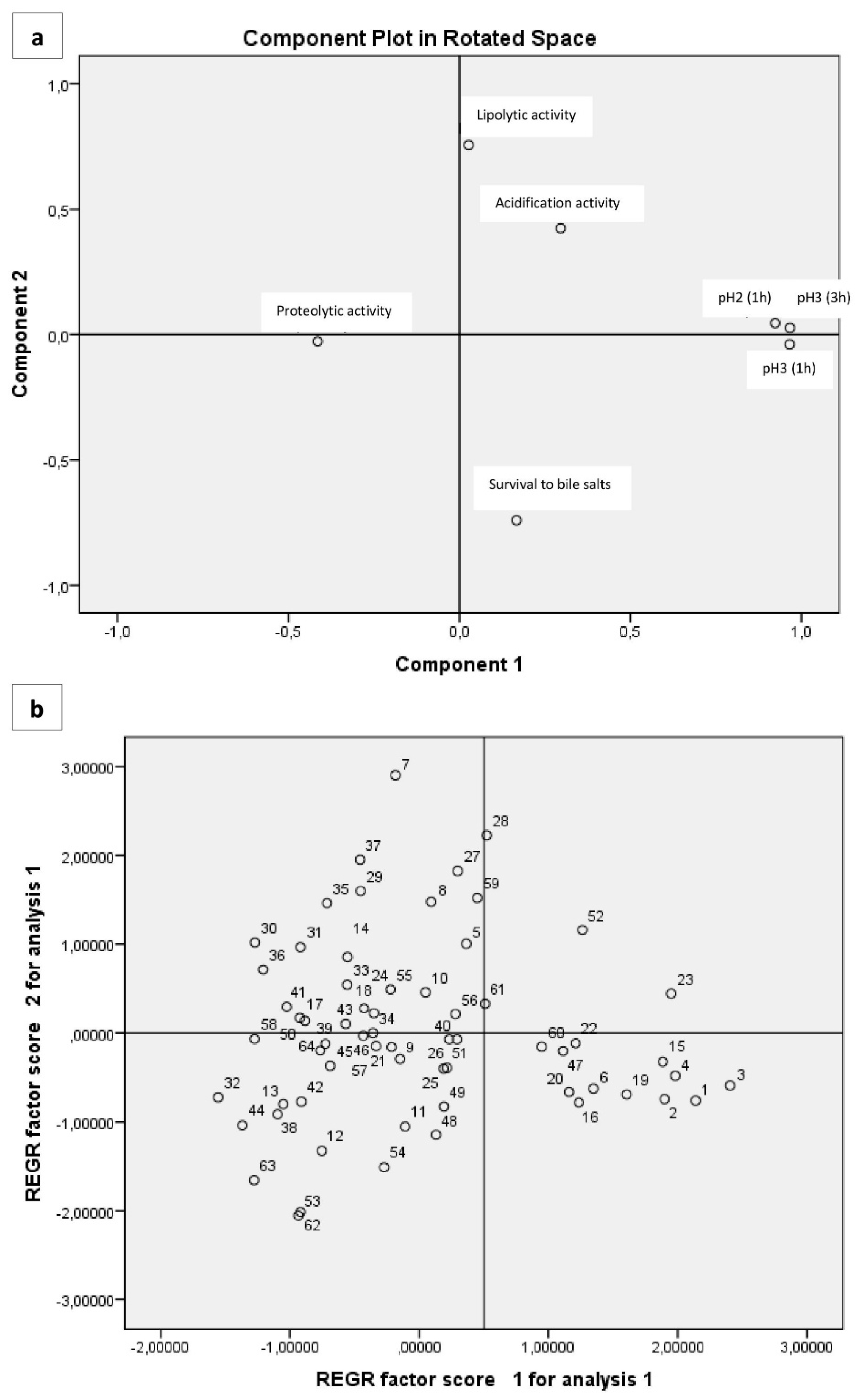
3.6. Multivariate Analysis of Phenotypic Characteristics Related to Probiotic Potential
In our study, three eigenvalues had value higher than 1. The study of the contribution of variables to factors (Table 8) showed that Factor 1 (43.19% of variance) was related to five variables (acidification activity, survival to bile salts, survival to pH 21h, survival to pH 31h, survival to pH 33h), Factor 2 (18.515% of variance) was related to one variable (lipolytic activity), and Factor 3 (13.093% of variance) to one variable (proteolytic activity) (Figure 3a). A projection of the variables on the plane formed by the first two factors (Figure 3b) shows a clear relationship between the variables described above. However, they are quite scattered among them, but they can be distinguished mainly by three groups in which are close to each other. The first group is placed on the negative part of the two factors and involves isolates that do not combine proteolysis and lipolysis (or absence of both), having low resistance to pH but quite good resistance to bile salts (up to 75%), as well. Also, the majority of them are characterized as medium acidifiers. In the second group (negative of first factor and positive of second one), they are isolates with a greater resistance to pH and bile salts than the first group, and proteolytic and lypolitic activity are combined (both presence and absence). However, in this group microorganisms are characterized as slow and/or medium acidifiers. In the third group, which is placed on the positive side of factor 1 and negative of factor 2, there are isolates with high survival to pH (more than 75% in pH 3 after 3 h) and bile salts (up to 90%). Furthermore the majority of them are high acidifiers, but they do not have proteolytic and lypolytic activity. Apart from three main groups described above, there are also a few isolates separated from the rest. For example isolates 11 and 12 have medium resistance to pH but high resistance to bile salts (93% and 100% respectively). They are also a few low acidifiers but they have both proteolytic and lipolytic activity. Furthermore, another group of four isolates (1, 2, 3, 4) was characterized by high values of acidification, while they have high resistance to pH (average 74% to pH 2 and 84% to pH 3 after 3 h) and bile salts (100%, 97% 100%, and 94%, respectively). None of these isolates have proteolytic and lipolytic activity. Isolate 16 which is depicted a bit away from the others (on positive side of factor 1 and negative of factor 2) presents intermediate values for all the parameters. According to PCA analysis, we conclude to seven isolates (1, 2, 3, 4, 11, 12, 16) having possibly promising possibly promising technological and probiotic attributes, as described above.
4. Conclusions
This is the first comprehensive report on the characterization of LAB isolates from fermented Cypriot table olives. Enterococci were the predominant group of LAB, and it seems that those strains could find possible applications in food industry, due to their interesting technological and probiotic properties, such as their resistance to low pH and to bile salts. However, more research is required in those essential characteristics, and scientists should give more attention in the future to the genus as it becomes apparent that is a dominant group of microorganisms in fermented food products.
Author Contributions
Conceptualization, D.A.A. and D.T.; Methodology, D.A.A., D.B., D.T.; Formal Analysis, D.A.A.; Investigation, D.A.A.; Resources, D.T.; Data Curation, D.A.A., D.T.; Writing—Original Draft Preparation, D.A.A. and D.B.; Writing—Review & Editing, D.T.; Project Administration, D.T.; Funding Acquisition, D.T.
Funding
This study was funded by internal funds from the Cyprus University of Technology to Dimitris Tsaltas.
Acknowledgments
The authors gratefully acknowledge the material provided by A. Agathokleous (King of Olives, Agglissides, Larnaca, Cyprus) and the financial support from the Cyprus University of Technology to Dimitris Tsaltas.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Olives, T. No 66—November 2012; International Olive Council: Madrid, Spain, 2012. [Google Scholar]
- López, F.N.A.; Romero, C.; Quintana, M.D.C.D.; López, A.L.; García, P.G.; Fernández, A.G. Kinetic study of the physicochemical and microbiological changes in ‘seasoned’ olives during the shelf-life period. J. Agric. Food Chem. 2005, 53, 5285–5292. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Gallego, J.; Arroyo-López, F.N.; Rantsiou, K.; Jiménez-Díaz, R.; Garrido-Fernández, A.; Cocolin, L. Screening of lactic acid bacteria isolated from fermented table olives with probiotic potential. Food Res. Int. 2013, 50, 135–142. [Google Scholar] [CrossRef]
- Montet, D.; Ray, R.C.; Zakhia-Rozis, N. Lactic Acid Fermentation of Vegetables and Fruits. In Microorganisms and Fermentation of Traditional Foods; CRC Press: Boca Raton, FL, USA, 2014; pp. 108–140. [Google Scholar]
- Stiles, M.E.; Holzapfel, W.H. Lactic acid bacteria of foods and their current taxonomy. Int. J. Food Microbiol. 1997, 36, 1–29. [Google Scholar] [CrossRef]
- Fisher, K.; Phillips, C. The ecology, epidemiology and virulence of Enterococcus. Microbiology 2009, 155, 1749–1757. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, F.; Willems, R.J.L.; Gilmore, M.S. Enterococcus Diversity, Origins in Nature, and Gut Colonization. In Enterococci: From Commensals to Lead. Causes Drug Resistant Infection; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014; pp. 1–56. [Google Scholar]
- Franz, C.M.A.P.; Huch, M.; Abriouel, H.; Holzapfel, W.; Gálvez, A. Enterococci as probiotics and their implications in food safety. Int. J. Food Microbiol. 2011, 151, 125–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, B.C.; Dora, B.; de Melo, G.; De Martinis, E.C.P.; Paulo, S. Dualistic aspects of Enterococcus spp. in foods. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 1119–1125. [Google Scholar]
- Omar, N.B.; Castro, A.; Lucas, R.; Abriouel, H.; Yousif, N.M.K.; Franz, C.M.A.P.; Holzapfel, W.H.; Ruben, P.P.; Martínez-Canãmero, M.; Gálvez, A. Functional and Safety Aspects of Enterococci Isolated from Different Spanish Foods. Syst. Appl. Microbiol. 2004, 27, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Randazzo, C.L.; Restuccia, C.; Romano, A.D.; Caggia, C. Lactobacillus casei, dominant species in naturally fermented Sicilian green olives. Int. J. Food Microbiol. 2004, 90, 9–14. [Google Scholar] [CrossRef]
- Gonza, J.M.; Lucena-padro, H.; Ruiz-barba, L.; Maldonado-Barraga, A. Enterococcus olivae sp. nov., isolated from Spanish-style green-olive fermentations. Int. J. Syst. Evol. Microbiol. 2014, 64, 2534–2539. [Google Scholar]
- Shinozaki-Kuwahara, N.; Saito, M.; Hirasawa, M.; Hirasawa, M.; Takada, K. Streptococcus dentiloxodontae sp. nov., isolated from the oral cavity of elephants. Int. J. Syst. Evol. Microbiol. 2016, 66, 3878–3883. [Google Scholar] [PubMed]
- Cogan, T.M. Characterization of the lactic acid bacteria in artisanal dairy products. J. Dairy Res. 1997, 64, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Estifanos, H. Isolation and identification of probiotic lactic acid bacteria from curd and in vitro evaluation of its growth inhibition activities against pathogenic bacteria. Afr. J. Microbiol. Res. 2014, 8, 1419–1425. [Google Scholar] [CrossRef]
- Jackson, C.R.; Fedorka-Cray, P.J.; Barrett, J.B. Use of a Genus- and Species-Specific Multiplex PCR for Identification of Enterococci Use of a Genus- and Species-Specific Multiplex PCR for Identification of Enterococci. J. Clin. Microbiol. 2004, 42, 3558. [Google Scholar] [CrossRef] [PubMed]
- Fuka, M.M.; Maksimovic, A.Z.; Tanuwidjaja, I.; Hulak, N.; Schloter, M. Characterization of enterococcal community isolated from an Artisan Istrian raw milk cheese: Biotechnological and safety aspects. Food Technol. Biotechnol. 2017, 55, 368–380. [Google Scholar]
- Franciosi, E.; Settanni, L.; Cavazza, A.; Poznanski, E. Biodiversity and technological potential of wild lactic acid bacteria from raw cows’ milk. Int. Dairy J. 2009, 19, 3–11. [Google Scholar] [CrossRef]
- Dinçer, E.; Kıvanç, M. Lipolytic Activity of Lactic Acid Bacteria Isolated from Turkish Pastırma. Anadolu Univ. J. Sci. Technol. C Life Sci. Biotechnol. 2018, 7, 12–19. [Google Scholar] [CrossRef]
- Imène, K.; Halima, Z.-K.; Nour-Eddine, K. Screening of exopolysaccharide-producing coccal lactic acid bacteria isolated from camel milk and red meat of Algeria. Afr. J. Biotechnol. 2017, 16, 1078–1084. [Google Scholar] [CrossRef]
- Ghabbour, N.; Lamzira, Z.; Thonart, P.; Cidalia, P.; Markaoui, M.; Asehraou, A. Selection of oleuropein-degrading lactic acid bacteria strains isolated from fermenting Moroccan green olives. Grasas Aceites 2011, 62, 84–89. [Google Scholar] [Green Version]
- Ribeiro, S.C.; Coelho, M.C.; Todorov, S.D.; Franco, B.D.G.M.; Dapkevicius, M.L.E.; Silva, C.C.G. Technological properties of bacteriocin-producing lactic acid bacteria isolated from Pico cheese an artisanal cow’s milk cheese. J. Appl. Microbiol. 2014, 116, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Łaniewska-Trokenheim, Ł. Virulence factors of Enterococcus spp. presented in food. LWT Food Sci. Technol. 2017, 75, 670–676. [Google Scholar] [CrossRef]
- Aspri, M.; Bozoudi, D.; Tsaltas, D.; Hill, C.; Papademas, P. Raw donkey milk as a source of Enterococcus diversity: Assessment of their technological properties and safety characteristics. Food Control 2017, 73, 81–90. [Google Scholar] [CrossRef]
- Vankerckhoven, V.; van Autgaerden, T.; Vael, C.; Lammens, C.; Chapelle, S.; Rossi, R.; Jabes, D.; Goossens, H. Development of a multiplex PCR for the detection of asaI, gelE, cylA, esp, and hyl genes in enterococci and survey for virulence determinants among european hospital isolates of Enterococcus faecium. J. Clin. Microbiol. 2004, 42, 4473–4479. [Google Scholar] [CrossRef] [PubMed]
- Martín-Platero, A.M.; Valdivia, E.; Maqueda, M.; Martínez-Bueno, M. Characterization and safety evaluation of enterococci isolated from Spanish goats’ milk cheeses. Int. J. Food Microbiol. 2009, 132, 24–32. [Google Scholar] [CrossRef] [PubMed]
- De las Rivas, B.; Marcobal, Á.; Muñoz, R. Improved multiplex-PCR method for the simultaneous detection of food bacteria producing biogenic amines. FEMS Microbiol. Lett. 2005, 244, 367–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilavenil, S.; Park, H.S.; Vijayakumar, M.; Arasu, M.V.; Kim, D.H.; Ravikumar, S.; Choi, K.C. Probiotic Potential of Lactobacillus Strains with Antifungal Activity Isolated from Animal Manure. Sci. World J. 2015, 2015, 802570. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Yun, B.; Moon, J.-H.; Park, D.-J.; Lim, K.; Oh, S. Characterization of Selected Lactobacillus Strains for Use as Probiotics. Korean J. Food Sci. Anim. Resour. 2015, 35, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Campaniello, D.; Bevilacqua, A.; D’Amato, D.; Corbo, M.R.; Altieri, C.; Sinigaglia, M. Microbial characterization of table olives processed according to Spanish and natural styles. Food Technol. Biotechnol. 2005, 43, 289–294. [Google Scholar]
- Aponte, M.; Ventorino, V.; Blaiotta, G.; Volpe, G.; Farina, V.; Avellone, G.; Lanza, C.M.; Moschetti, G. Study of green Sicilian table olive fermentations through microbiological, chemical and sensory analyses. Food Microbiol. 2010, 27, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Panagou, E.Z.; Tassou, C.C.; Katsaboxakis, C.Z. Induced lactic acid fermentation of untreated green olives of the Conservolea cultivar by Lactobacillus pentosus. J. Sci. Food Agric. 2003, 83, 667–674. [Google Scholar] [CrossRef]
- De Castro, A.; Montano, A.; Casado, F.-J.; Sanchez, A.-H.; Rejano, L. Utilization of Enterococcus casseliflavus and Lactobacillus pentosus as starter cultures for Spanish-style green olive fermentation. Food Microbiol. 2002, 19, 637–644. [Google Scholar] [CrossRef]
- Sarantinopoulos, P.; Andrighetto, C.; Georgalaki, M.D.; Rea, M.C.; Lombardi, A.; Cogan, T.M.; Kalantzopoulos, G.; Tsakalidou, E. Biochemical properties of enterococci relevant to their technological performance. Int. Dairy J. 2001, 11, 621–647. [Google Scholar] [CrossRef]
- Morea, M.; Baruzzi, F.; Cocconcelli, P.S. Molecular and physiological characterization of dominant bacterial populations in traditional Mozzarella cheese processing. J. Appl. Microbiol. 1999, 87, 574–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzzi, G.; Caruso, M.; Gardini, F.; Lombardi, A.; Vannini, L.; Guerzoni, M.E.; Andrighetto, C.; Lanorte, M.T. A survey of the enterococci isolated from an artisanal Italian goat’s cheese (semicotto caprino). J. Appl. Microbiol. 2000, 89, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Durlu özkaya, F.; Xanthopoulos, V.; Tunail, N.; Litopoulou-Tzanetaki, E. Technologically important properties of lactic acid bacteria isolates from Beyaz cheese made from raw ewes’ milk. J. Appl. Microbiol. 2001, 91, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Kanmani, P.; Suganya, K.; Kumar, R.S.; Yuvaraj, N.; Pattukumar, V.; Paari, K.A.; Arul, V. Synthesis and functional characterization of antibiofilm exopolysaccharide produced by Enterococcus faecium mc13 isolated from the gut of fish. Appl. Biochem. Biotechnol. 2013, 169, 1001–1015. [Google Scholar] [CrossRef] [PubMed]
- Charoenprasert, S.; Mitchell, A. Factors influencing phenolic compounds in table olives (Olea europaea). J. Agric. Food Chem. 2012, 60, 7081–7095. [Google Scholar] [CrossRef] [PubMed]
- Laëtitia, G.; Pascal, D.; Yann, D. The Citrate Metabolism in Homo- and Heterofermentative LAB: A Selective Means of Becoming Dominant over Other Microorganisms in Complex Ecosystems. Food Nutr. Sci. 2014, 5, 953–969. [Google Scholar] [CrossRef]
- González, L.; Sacristán, N.; Arenas, R.; Fresno, J.M.; Tornadijo, M.E. Enzymatic activity of lactic acid bacteria (with antimicrobial properties) isolated from a traditional Spanish cheese. Food Microbiol. 2010, 27, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Creti, R.; Imperi, M.; Bertuccini, L.; Fabretti, F.; Orefici, G.; Rosa, R.D.; Baldassarri, L. Survey for virulence determinants among Enterococcus faecalis isolated from different sources. J. Med. Microbiol. 2004, 53, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Franz, C.; Muscholl-Silberhorn, A.; Yousif, N.; Vancanneyt, M.; Swings, J.; Holzapfel, W. Incidence of virulence factors and antibiotic resistance among enterococci isolated from food. Appl. Environ. Microbiol. 2001, 67, 4385–4389. [Google Scholar] [CrossRef] [PubMed]
- Rosado, D.; Brito, J.C.; Harris, D.J. Molecular screening of Hepatozoon (Apicomplexa: Adeleorina) infections in Python sebae from West Africa using 18S rRNA gene sequences. Herpetol. Notes 2015, 8, 461–463. [Google Scholar]
- Klein, G. Taxonomy, ecology and antibiotic resistance of enterococci from food and the gastro-intestinal tract. Int. J. Food Microbiol. 2003, 88, 123–131. [Google Scholar] [CrossRef]
- Strompfová, V.; Lauková, A.; Ouwehand, A.C. Selection of enterococci for potential canine probiotic additives. Vet. Microbiol. 2004, 100, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.; Garriga, M.; Hugas, M.; Aymerich, T. Genetic diversity and safety aspects of enterococci from slightly fermented sausages. J. Appl. Microbiol. 2005, 98, 1177–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Percentage of number of isolates categorized in three acidification activity groups (low ΔpH < 1.5, medium ΔpH (1.5, 2) and high ΔpH ≥ 2) after 24 h incubation at 37 °C in 10% skim milk.
Figure 1.
Percentage of number of isolates categorized in three acidification activity groups (low ΔpH < 1.5, medium ΔpH (1.5, 2) and high ΔpH ≥ 2) after 24 h incubation at 37 °C in 10% skim milk.

Figure 2.
Percentage of number of: (a) proteolytic positive isolates and (b) lipolytic positive isolates.
Figure 2.
Percentage of number of: (a) proteolytic positive isolates and (b) lipolytic positive isolates.

Figure 3.
Projection of the variables (a) cases (b) isolates onto the plane formed by the first two factors.
Figure 3.
Projection of the variables (a) cases (b) isolates onto the plane formed by the first two factors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Microbiological media used for microflora enumeration.
| Growth Media | Microorganisms | Method | Incubation Conditions |
|---|---|---|---|
| Plate Count Agar (PCA) (Merck, Darmstadt, Germany) | Total viable count | Spread plate | 30 °C/72 h |
| De Man-Rogosa-Sharpe Agar (MRS) (Oxoid, Basingstoke, UK) + natamycin 4% | Lactic acid bacteria | Pour plate/Overlay | 30 °C/72 h |
| M17 (Oxoid, Basingstoke, UK) | Lactic acid bacteria | Pour plate | 37 °C/72 h |
| Sabouraud Agar (Oxoid, Basingstoke, UK) | Yeast and molds | Spread plate | 25 °C/3 day |
| Violet Red Bile Glycose Agar (VRBGA) (BD, Sparks, MD, USA) | Enterobacteriaceae | Pour plate/Overlay | 37 °C/24 h |
| Violet Red Bile Lactose Agar (VRBL) (Oxoid, Basingstoke, UK) | Coliforms | Pour plate/Overlay | 30 °C/24 h |
| Baird Parker egg yolk tellurite (BPM) (Oxoid, Basingstoke, UK) | Micrococcaceae | Spread plate | 37 °C/48 h |
| Nutrient Agar Crystal Violet (NACV) (Oxoid, Basingstoke, UK) | Gram- cocci | Spread plate | 21 °C/48 h |
| Mannitol Salt Agar (MSA) (Oxoid, Basingstoke, UK) | Salt resistant | Spread plate | 30 °C/48 h |
Table 2.
Polymerase chain reaction (PCR) primers, products, and reference strains.
| Strain | Primer | Sequence (5′–3′) | Product Size (bp) |
|---|---|---|---|
| E. durans ATCC19432 | DU1 | CCTACTGATATTAAGACAGCG | 295 |
| DU2 | TAATCCTAAGATAGGTGTTTG | ||
| E. faecalis ATCC19433 | FL1 | ACTTATGTGACTAACTTAACC | 360 |
| FL2 | TAATGGTGAATCTTGGTTTGG | ||
| E. faecium ATCC19434 | FM1 | GAAAAAACAATAGAAGAATTAT | 215 |
| FM2 | TGCTTTTTTGAATTCTTCTTTA | ||
| E. casseliflavus ATCC25788 | CA1 | TCCTGAATTAGGTGAAAAAAC | 288 |
| CA2 | GCTAGTTTACCGTCTTTAACG | ||
| E. gallinarum ATCC49673 | GA1 | TTACTTGCTGATTTTGATTCG | 173 |
| GA2 | TGAATTCTTCTTTGAAATCAG | ||
| E. hirae ATCC 8043 | HI1 | CTTTCTGATATGGATGCTGTC | 187 |
| HI2 | TAAATTCTTCCTTAAATGTTG |
Table 3.
List of primers used for the amplification of pathogenicity related genes.
| Target Gene | Primer Sequence | Annealing Temperature (°C) | Fragment Size (bp) | Reference |
|---|---|---|---|---|
| Aggregationsubstance (asa1) | GCACGCTATTACGAACTATGA TAAGAAAGAACATCACCACGA | 50 | 375 | [25] |
| Adhesion of collagen protein (ace) | GAATTGAGCAAAAGTTCAATCG GTCTGTCTTTTCACTTGTTTC | 48 | 1008 | [26] |
| Cytolysin (cylA) | ACTCGGGGATTGATAGGC GCTGCTAAAGCTGCGCTT | 52 | 688 | [25] |
| Endocartidis antigen (efaA) | GCCAATTGGGACAGACCCTC CGCCTTCTGTTCCTTCTTTGGC | 57 | 688 | [26] |
| Enterococcal surface protein (esp) | AGATTTCATCTTTGATTCTTG AATTGATTCTTTAGCATCTGG | 50 | 510 | [25] |
| Gelatinase (gelE) | TATGACAATGCTTTTTGGGAT AGATGCACCCGAAATAATATA | 47 | 213 | [25] |
| Hyluronidase (hyl) | ACAGAAGAGCTGCAGGAAATG GACTGACGTCCAAGTTTCCAA | 53 | 276 | [25] |
| Vancomycin Resistance (vanA) | TCTGCAATAGAGATAGCCGC GGAGTAGCTATCCCAGCATT | 52 | 377 | [26] |
| Vancomycin Resistance (vanB) | GCTCCGCAGCCTGCATGGACA ACGATGCCGCCATCCTCCTGC | 60 | 529 | [26] |
| Histidine decarboxylase (hdc1) | AGATGGTATTGTTTCTTATG AGACCATACACCATAACCTT | 46 | 367 | [27] |
| Histidine decarboxylase (hdc2) | AAYTCNTTYGAYTTYGARAARGARG ATNGGNGANCCDATCATYTTRTGNCC | 50 | 534 | [27] |
| Tyrosine decarboxylase (tdc) | GAYATNATNGGNATNGGNYTNGAYCARG CCRTARTCNGGNATAGCRAARTCNGTRTG | 55 | 924 | [27] |
| Ornithinedecarboxylase (odc) | GTNTTYAAYGCNGAYAARCANTAYTTYGT ATNGARTTNAGTTCRCAYTTYTCNGG | 54 | 1446 | [27] |
Table 4.
Mean values (log10cfu/g; x ± SD) of the counts of different microbial groups, as well as pH values of Cypriot green cracked table olives. Mean values of three individual plate counts.
Table 4.
Mean values (log10cfu/g; x ± SD) of the counts of different microbial groups, as well as pH values of Cypriot green cracked table olives. Mean values of three individual plate counts.
| Medium | Log cfu mL−1 |
|---|---|
| TAC a | 9.18 ± 0.07 |
| Yeasts and Molds | 3.48 ± 0.06 |
| LAB on MRS agar | 8.04 ± 0.04 |
| Coliforms | 4.66 ± 0.02 |
| Enterobacteriaceae | 3.83 ± 0.01 |
| Micrococacceae | nd * |
| LAB on M17 agar | 4.86 ± 0.06 |
| Salt Resistant Bacteria | 7.81 ± 0.08 |
| Gram -ve | 5.37 ± 0.03 |
| pH | 3.89 ± 0.01 |
* nd: not detected; a TAC: Total Aerobic Counts.
Table 5.
Isolates of Enterococcus spp. showing positive or negative proteolytic activity, positive or negative lipolytic activity, and acidification activity of high medium or low level (low ΔpH < 1.5, medium ΔpH (1.5,2) and high ΔpH ≥ 2 after 24 h incubation at 37 °C in 10% skim milk).
Table 5.
Isolates of Enterococcus spp. showing positive or negative proteolytic activity, positive or negative lipolytic activity, and acidification activity of high medium or low level (low ΔpH < 1.5, medium ΔpH (1.5,2) and high ΔpH ≥ 2 after 24 h incubation at 37 °C in 10% skim milk).
| Proteolytic Activity (Positives) | Proteolytic Activity (Negatives) | Lipolytic Activity (Positives) | Lipolytic Activity (Negatives) | Acidification Activity (High) | Acidification Activity (Medium) | Acidification Activity (Low) | |
|---|---|---|---|---|---|---|---|
| Isolates (Enterococcus) | 7,11–14,16–18,27–37 | 1–6,8–10,15,19–26,38–64 | 11–13,16–18,32,36,42,53,54,62,63 | 1–10,14,15,19–31,33,34,35,37–41,43–52,55–61,64 | 3,23,26,27,28,42,47,52,59 | 1,2,4–10,13,14,18,21,22,24,25,29–33,35,37,39,40,41,43,50,51,55,56,57,60–64 | 11,12,15,16,17,19,20,34,36,38,44,45,46,48,49,53,54,58 |
Table 6.
Percentage of isolates for survival at various rates to low pH and bile salts.
| Survival Rate | pH 2 (1 h) | pH 3 (1 h) | pH 3 (3 h) | Bile Salts 0.3% |
|---|---|---|---|---|
| >70% | 89% | 57.8% | 82.7% | 10.9% |
| (70%, 80%) | 11% | 20.3% | 9.5% | 20.3% |
| (80%, 90%) | 0% | 12.5% | 7.8% | 34.4% |
| <90% | 0% | 9.4% | 0% | 34.4% |
Table 7.
Isolates of Enterococcus spp. surviving at different rates for various pH and exposure times or bile salts.
Table 7.
Isolates of Enterococcus spp. surviving at different rates for various pH and exposure times or bile salts.
| Survival Rate | pH 2 (1 h) | pH 3 (1 h) | pH 3 (3 h) | Bile Salts 0.3% |
|---|---|---|---|---|
| >70% | 2–14,17,18,20,21,22,24–64 | 7,8,9,10,12,13,14,17,18,21,24,25,26,29–39,41–46,50,53,57,58,62,63,64 | 5,7,8,9,10,11,12,13,14,17,18,20,21,22,24–51,53,54,55,56,57,58,59,61,62,63,64 | 5,7,8,17,36,37,59 |
| (70%, 80%) | 1,3,4,15,16,19,23 | 5,11,27,28,40,48,49,51,54,55,56,59,61 | 1,2,3,4,6,16,19,23,52,60 | 10,18,23,24,28,29,35,43,45,46,52,55,58 |
| (80%, 90%) | - | 6,16,19,20,22,47,52,60 | 1,2,3,4,15, | 9,14,15,16,20,21,22,27,30,31,39,40,41,42,50,51,53,54,56,57,60,61 |
| ≤90% | - | 1,2,3,4,15,23 | - | 1,2,3,4,6,11,12,13,19,25,26,32,33,34,38,44,47,48,49,62,63,64 |
Table 8.
Contribution of variables (proteolytic activity, lipolytic activity, acidification activity, survival to pH 2 for 1 h, survival to pH 3 for 1 h, survival to pH 3 for 3 h and survival to bile salts) to the factors in the PCA based on correlations.
Table 8.
Contribution of variables (proteolytic activity, lipolytic activity, acidification activity, survival to pH 2 for 1 h, survival to pH 3 for 1 h, survival to pH 3 for 3 h and survival to bile salts) to the factors in the PCA based on correlations.
| Variable | Factor 1 | Factor 2 | Factor 3 |
|---|---|---|---|
| Proteolytic activity | −0.416 | <0.01 | 0.683 |
| Lipolytic activity | 0.055 | 0.753 | 0.250 |
| Acidification activity | 0.311 | 0.413 | −0.545 |
| Survival to pH2 1h | 0.925 | 0.011 | 0.240 |
| Survival to pH3 1h | 0.967 | <0.01 | 0.117 |
| Survival to pH3 3h | 0.964 | <0.01 | 0.119 |
| Survival to bile salts | 0.139 | −0.747 | <0.01 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Anagnostopoulos, D.A.; Bozoudi, D.; Tsaltas, D. Enterococci Isolated from Cypriot Green Table Olives as a New Source of Technological and Probiotic Properties. Fermentation 2018, 4, 48. https://doi.org/10.3390/fermentation4020048
AMA Style
Anagnostopoulos DA, Bozoudi D, Tsaltas D. Enterococci Isolated from Cypriot Green Table Olives as a New Source of Technological and Probiotic Properties. Fermentation. 2018; 4(2):48. https://doi.org/10.3390/fermentation4020048
Chicago/Turabian StyleAnagnostopoulos, Dimitrios A., Despina Bozoudi, and Dimitrios Tsaltas. 2018. "Enterococci Isolated from Cypriot Green Table Olives as a New Source of Technological and Probiotic Properties" Fermentation 4, no. 2: 48. https://doi.org/10.3390/fermentation4020048
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.