Sero-Surveillance of Lyssavirus Specific Antibodies in Nigerian Fruit Bats (Eidolon helvum)

 and
and 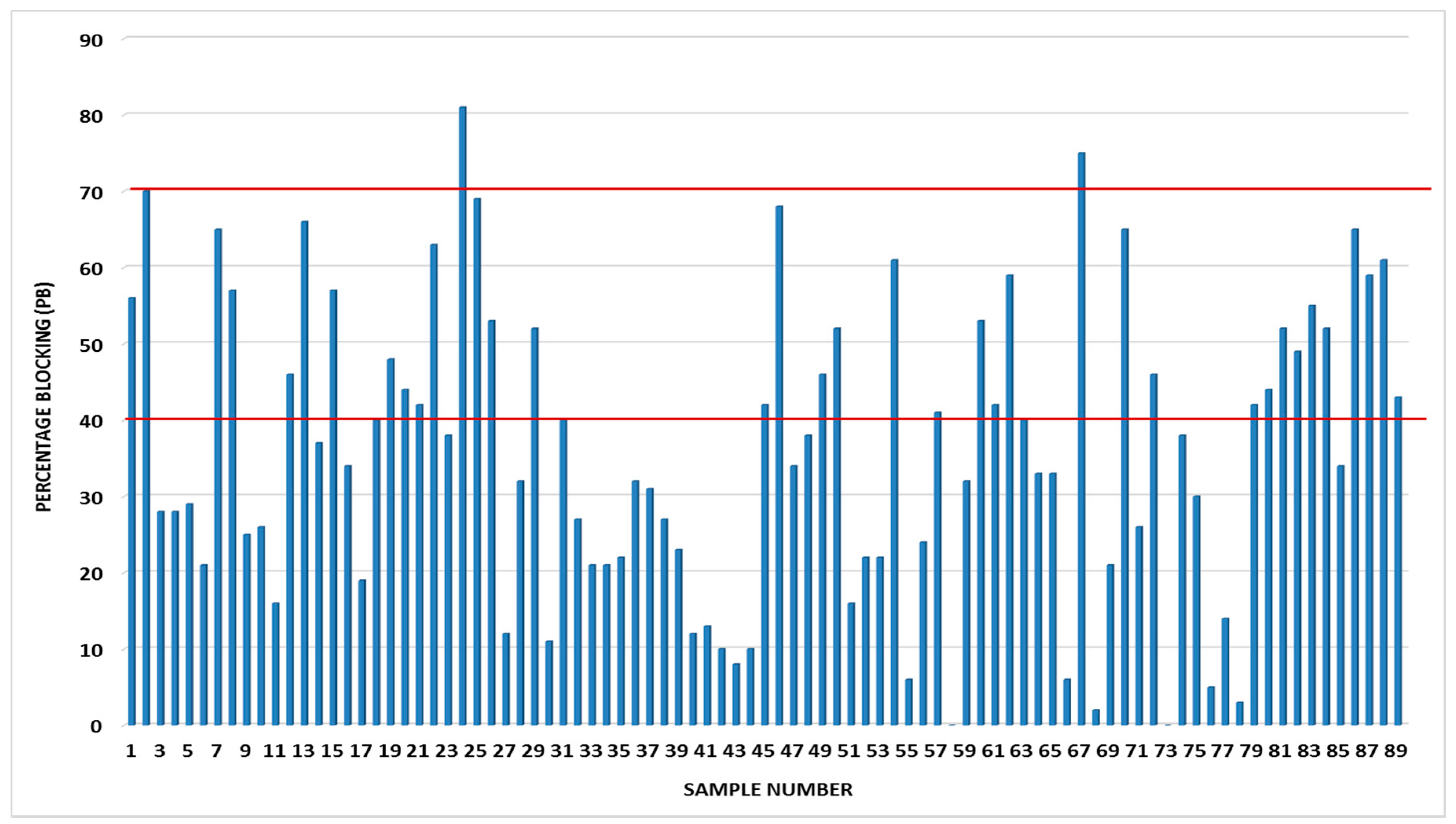{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
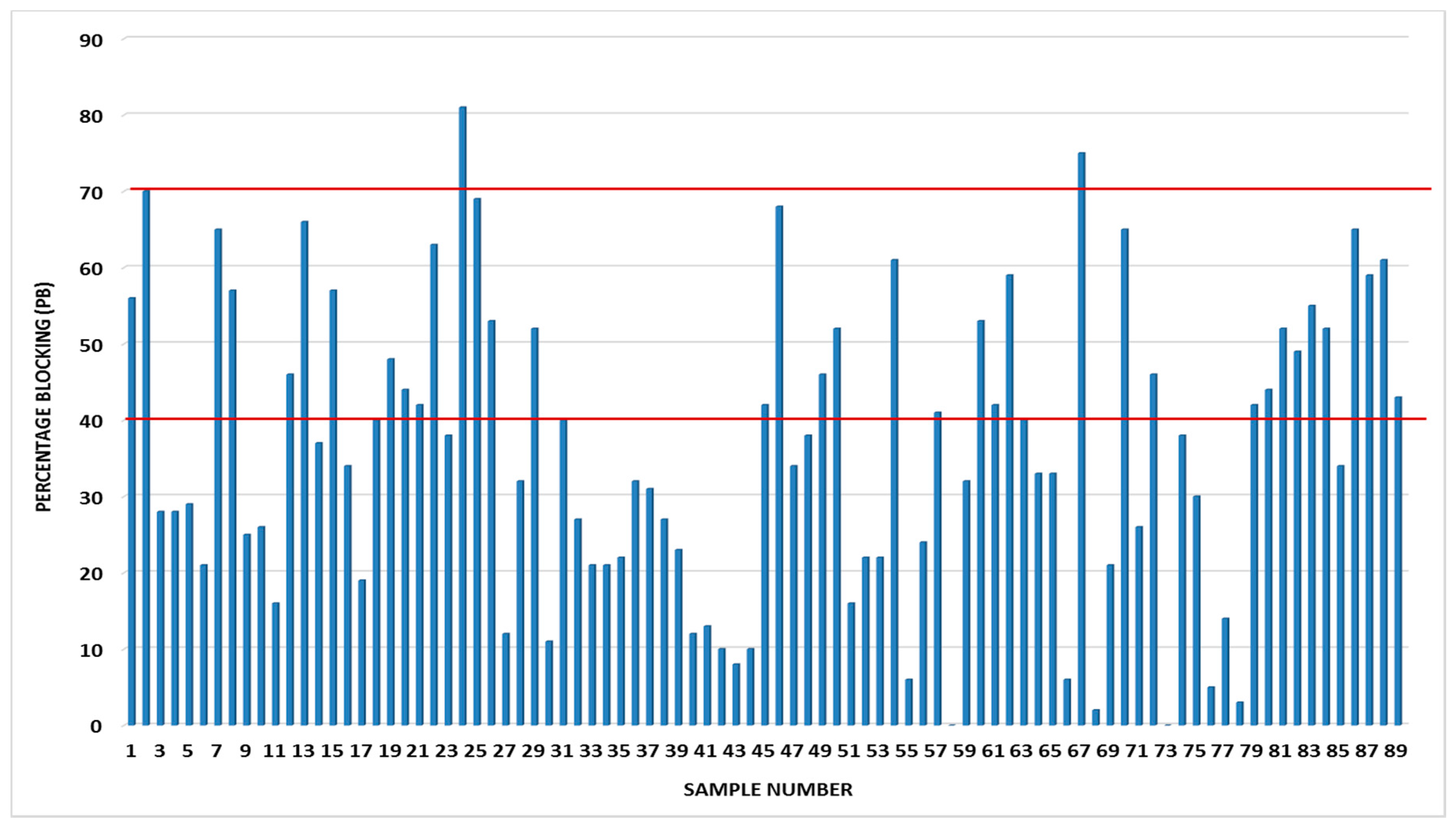
3. Results and Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- ICTV (International Committee on Taxonomy of Viruses). Available online: http://www.ictvonline.org/virusTaxonomy.asp?bhcp=1 (accessed on 30 May 2017).
- Marston, D.A.; Ellis, R.J.; Wise, E.L.; Aréchiga-Ceballos, N.; Freuling, C.M.; Banyard, A.C.; Mcelhinney, L.M.; De Lamballerie, X.; Müller, T.; Fooks, A.R.; et al. Complete genome sequence of Lleida bat lyssavirus. Genome Announc. 2017, 5, e01427–16. [Google Scholar] [CrossRef] [PubMed]
- Aréchiga, C.N.; Vázquez, M.S.; Berciano, J.M.; Nicolás, O.; Aznar, L.C.; Juste, J.; Rodríguez, N.C.; Aguilar, S.A.; Echevarría, J.E. Novel lyssavirus in bat, Spain. Emerg. Infect. Dis. 2013, 19, 793–795. [Google Scholar]
- Gunawardena, P.S.; Marston, D.A.; Ellis, R.J.; Wise, E.L.; Karawita, A.C.; Breed, A.C.; Mcelhinney, L.M.; Johnson, N.; Banyard, A.C.; Fooks, A.R. Lyssavirus in Indian flying foxes, Sri Lanka. Emerg. Infect. Dis. 2016, 22, 1456–1459. [Google Scholar] [CrossRef] [PubMed]
- Badrane, H.; Bahloul, C.; Perrin, P.; Noel, T. Evidence of two Lyssavirus phylogroups with distinct pathogenicity and immunogenicity. J. Virol. 2001, 75, 3268–3276. [Google Scholar] [CrossRef] [PubMed]
- Hanlon, C.A.; Kuzmin, I.V.; Blanton, J.D.; Weldon, W.C.; Manangan, J.S.; Rupprecht, C.E. Efficacy of rabies biologics against new lyssaviruses from Eurasia. Virus Res. 2005, 111, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Mayer, A.E.; Niezgoda, M.; Markotter, W.; Agwanda, B.; Breiman, R.F.; Rupprecht, C.E. Shimoni bat virus, a new representative of the Lyssavirus genus. Virus Res. 2010, 149, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Hayman, D.T.S.; Fooks, A.R.; Marston, D.A.; Garcia-R, J.C. The Global Phylogeography of Lyssaviruses—Challenging the ‘Out of Africa’ hypothesis. PLoS Neglect. Trop. Dis. 2016, 10, e0005266. [Google Scholar] [CrossRef] [PubMed]
- Fooks, A.R.; Banyard, A.C.; Horton, D.L.; Johnson, N.; McElhinney, L.M.; Jackson, A.C. Current status of rabies and prospects for elimination. Lancet 2014, 384, 1389–1399. [Google Scholar] [CrossRef]
- Hampson, K.; Coudeville, L.; Lembo, T.; Sambo, M.; Kieffer, A.; Attlan, M.; Barrat, J.; Blanton, J.D.; Briggs, D.J.; Cleaveland, S.; et al. Estimating the global burden of endemic canine rabies. PLoS Neglect. Trop. Dis. 2015, 9, e0003786. [Google Scholar] [CrossRef] [PubMed]
- Boulger, I.R.; Porterfield, J.S. Isolation of a virus from Nigerian fruit bats. Trans. Roy. Soc. Trop. Med. Hyg. 1958, 52, 421–424. [Google Scholar] [CrossRef]
- Dzikwi, A.A.; Kuzmin, I.; Umoh, U.J.; Kwage, J.K.P.; Ahmad, A.A.; Rupprecht, C.E. Evidence of Lagos bat virus circulation among Nigerian fruit bats. J. Wildl. Dis. 2010, 46, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Hayman, D.T.S.; Fooks, A.R.; Rowcliffe, J.M.; McCrea, R.; Restif, O.; Baker, K.S.; Horton, D.L.; Suu-Ire, R.; Cunningham, A.A.; Wood, J.L.N. Endemic Lagos bat virus infection in Eidolon helvum. Epidemiol. Infect. 2012, 140, 2163–2171. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Niezgoda, M.; Franka, R.; Agwanda, B.; Markotter, W.; Beagley, J.C.; Urazova, O.Y.; Breiman, R.F.; Rupprecht, C.E. Lagos bat virus in Kenya. J. Clin. Microbiol. 2008, 46, 1451–1461. [Google Scholar] [CrossRef] [PubMed]
- Fernandes de Almeida, M.; Alves Martorelli, L.F.; Sodré, M.M.; Arruda Geraldes Kataoka, A.P.; da Rosa, A.R.; de Oliveira, M.L.; Amatuzzi, E. Rabies diagnosis and serology in bats from the State of São Paulo, Brazil. Rev. Soc. Bras. Med. Trop. 2011, 44, 140–145. [Google Scholar] [CrossRef]
- Wasniewski, M.; Almeida, I.; Baur, A.; Bedekovic, T.; Boncea, D.; Chaves, L.B.; David, D.; de Benedictis, P.; Dobrostana, M.; Giraud, P.; et al. First international collaborative study to evaluate rabies antibody detection method for use in monitoring the effectiveness of oral vaccination programmes in fox and raccoon dog in Europe. J. Virol. Methods 2016, 238, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Cliquet, F.; Aubert, M.; Sagne, L. Development of a fluorescent antibody virus neutralisation test (FAVN test) for the quantitation of rabies-neutralising antibody. J. Immunol. Methods 1998, 212, 79–87. [Google Scholar] [CrossRef]
- Aghomo, H.O.; Ako-Nai, A.K.; Oduye, O.O.; Tomori, O.; Rupprecht, C.E. Detection of rabies virus antibodies in fruit bats (Eidolon helvum) from Nigeria. J. Wildl. Dis. 1990, 26, 258–261. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyem, D.A.; Dogonyaro, B.B.; Woma, T.A.; Ngoepe, E.C.; Sabeta, C.T. Sero-Surveillance of Lyssavirus Specific Antibodies in Nigerian Fruit Bats (Eidolon helvum). Trop. Med. Infect. Dis. 2017, 2, 26. https://doi.org/10.3390/tropicalmed2030026
Tyem DA, Dogonyaro BB, Woma TA, Ngoepe EC, Sabeta CT. Sero-Surveillance of Lyssavirus Specific Antibodies in Nigerian Fruit Bats (Eidolon helvum). Tropical Medicine and Infectious Disease. 2017; 2(3):26. https://doi.org/10.3390/tropicalmed2030026
Chicago/Turabian StyleTyem, Dinchi A., Banenat B. Dogonyaro, Timothy A. Woma, Ernest Chuene Ngoepe, and Claude Taurai Sabeta. 2017. "Sero-Surveillance of Lyssavirus Specific Antibodies in Nigerian Fruit Bats (Eidolon helvum)" Tropical Medicine and Infectious Disease 2, no. 3: 26. https://doi.org/10.3390/tropicalmed2030026