Soil-Transmitted Helminths in Tropical Australia and Asia
by
 ,
,
Catherine A. Gordon
1,* ,
,
Johanna Kurscheid
2,
Malcolm K. Jones
3,
Darren J. Gray
2 and
Donald P. McManus
1 1
QIMR Berghofer Medical Research Institute, Molecular Parasitology Laboratory, Queensland 4006, Australia
2
Australian National University, Department of Global Health, Research School of Population Health, Australian Capital Territory 2601, Australia
3
School of Veterinary Science, University of Queensland, Brisbane, QLD 4067, Australia
*
Author to whom correspondence should be addressed.
Trop. Med. Infect. Dis. 2017, 2(4), 56; https://doi.org/10.3390/tropicalmed2040056
Submission received: 29 August 2017
/
Revised: 16 October 2017
/
Accepted: 17 October 2017
/
Published: 23 October 2017
(This article belongs to the Special Issue Neglected and Emerging Tropical Diseases in South and Southeast Asia and Northern Australia)
Abstract
:Soil-transmitted helminths (STH) infect 2 billion people worldwide including significant numbers in South-East Asia (SEA). In Australia, STH are of less concern; however, indigenous communities are endemic for STH, including Strongyloides stercoralis, as well as for serious clinical infections due to other helminths such as Toxocara spp. The zoonotic hookworm Ancylostoma ceylanicum is also present in Australia and SEA, and may contribute to human infections particularly among pet owners. High human immigration rates to Australia from SEA, which is highly endemic for STH Strongyloides and Toxocara, has resulted in a high prevalence of these helminthic infections in immigrant communities, particularly since such individuals are not screened for worm infections upon entry. In this review, we consider the current state of STH infections in Australia and SEA.
1. Introduction
Soil-transmitted helminths (STH) are estimated to infect 2 billion people worldwide. Many of these infections occur in South-East Asia (SEA) [1,2]. Species included in the term STH are the human hookworm species Ancylostoma duodenale and Necator americanus, the human roundworm Ascaris lumbricoides, and the human whipworm Trichuris trichiura [3]. Hookworm and Trichuris have zoonotic counterparts (A. caninum, A. ceylanicum, T. suis, and T. vulpis) [4,5,6,7,8,9,10,11,12,13,14]. A. lumbricoides itself is a zoonosis as the previously-identified pig roundworm, A. suum, has been found through molecular characterisation to be nearly identical to A. lumbricoides, and instead represents a haplotype of A. lumbricoides [6,15]. Toxocara canis and Strongyloides stercoralis are additional important nematode species of dogs that can also infect humans and are included in this review. Strongyloides is estimated to infect 30–100 million people [16,17], while the seroprevalence (2%–5% in urban areas, 14.2%–37% in rural areas) of Toxocara in developed countries indicates that the number of people at risk of infection may be in the millions [18]. While Toxocara and Strongyloides have low prevalences in Australia, our closest neighbours in SEA, as well as many countries from which refugees and immigrants originate, are highly endemic for these parasites. These helminths are also considered neglected tropical diseases (NTD) due to their occurrence in low socio-economic regions and have thus not received as much attention as other diseases occurring in developed countries. Primarily, these helminthic infections are endemic in tropical and subtropical areas due to the requirements for warm moist soil for egg or larval development.
Another potential reason for their status as NTDs is the chronic rather than acute nature of the infections they cause. Symptoms are similar between the causative species and are generally non-specific; namely nausea and/or vomiting, diarrhoea, abdominal pain, and fever. In adults the impact can be seen as lower ability to work. As such, the impact of these parasites should be examined by considering the disability adjusted life years (DALYs) to measure disease burden. According to the WHO definition, one DALY is one year of life quality lost compared to a healthy individual, and the sum of these DALYs across a population is a measurement of the gap between current health status and an ideal health situation [19].
As of 2010 there were an estimated 438.9 million people infected globally with hookworm, 819.0 million with A. lumbricoides, and 464.4 million with T. trichiura [20]. It was calculated that STH contributed to 4.98 million years lived with disability (YLDs), with 65% attributed to hookworm, 22% to A. lumbricoides, and 13% to T. trichiura [20]. The DALYs for intestinal helminths (including only A. lumbricoides, T. trichiura, and hookworm) have been reduced from 170 per 100,000 (94–290) in 1990 to 75 per 100,000 (43–128) in 2010 and 69.4 per 100,000 (43.3–106.4) in 2013 [21,22]. DALYs are not considered a good measurement of the burden of disease for S. stercoralis since the majority of infections cause limited clinical symptoms; the most common complaint and symptom is stomachache. Poor diagnostics for S. stercoralis also result in the true prevalence being underestimated [23].
Infection with hookworm or hyper-infection with S. stercoralis can result in anaemia, and hookworm can also present with cutaneous rash from larval migration. Ascaris, Strongyloides and hookworm larvae migrate to the lungs to be coughed up and swallowed, thus entering the gut where they mature. Lung-stage infection by Ascaris can cause pneumonia, called Loeffler’s pneumonia, while disseminated Strongyloides can also cause pneumonia and pulmonary haemorrhage; hookworm-associated pneumonia, ‘eosinophilic pneumonia’, is a rare manifestation [24,25,26]. Toxocara infections can result in a range of symptoms depending on where the larvae migrate. The migrating larvae themselves, much like in Strongyloides and hookworm infections, can result in a rash, or larval tracks, due to inflammation. In the eye, Toxocara can cause partial or total retinal detachment leading to blindness and may result in neurological symptoms if the larvae are present in the brain [18,27].
Strongyloides stercoralis, which is endemic in aboriginal communities of Australia, can be a serious roundworm infection with severe health implications due to autoinfection and dissemination. Infection in immunocompromised individuals is particularly serious, and can be fatal. Autoinfection with S. stercoralis occurs when the larvae produced by the adult worms cause reinfection without ever having to leave the body. In such instances there is no immune response against the migrating larvae and this can lead to hyperinfection and dissemination. Continuous reinfection with S. stercoralis through autoinfection can also lead to persistent infections lasting many years (Figure 1). Infections in immigrants in Australia have been found more than 20 years after moving away from an endemic area (Table 1). Disseminated strongyloidiasis occurs when the parasite is distributed throughout the body and is more commonly seen in people with impaired immune systems [28]. It can lead to abdominal pain and swelling, pulmonary and neurological complications and meningitis, depending on where the parasite is located, as well as septicaemia, a leading cause of death in S. stercoralis infection [28,29]. Septicaemia occurs due to migration of larvae through the gastrointestinal wall. While generally considered as a human parasite, S. stercoralis has also been found in non-human primates and dogs [30,31,32]. There are also haplotypes identified for this species, and grouping of haplotypes from humans with haplotypes from canids and indicates potential zoonotic transmission [33,34]. Sequencing of isolates found in dogs and humans in Cambodia found two haplotypes of S. stercoralis in dogs, one of which was indistinguishable from that found in humans, again indicating the potential for zoonotic transmission [30]. Dogs are an important reservoir for zoonotic infections in terms of transmission as they live in close contact with humans, increasing the likelihood of transmission when compared with other potential zoonotic hosts such as non-human primates, which have a far more limited association with humans. Australia has a high pet ownership, particularly dogs and cats, with many also living inside homes, thus providing clear potential for transmission [35].
Recent molecular analysis of hookworm species has been important for speciation and shows a different epidemiological pattern than previously thought, including a much higher prevalence of A. ceylanicum, which had been considered to be only a rare infection in humans [4,11,50,51,52,53]. This is due in part to the morphological similarity of eggs and larval stages between hookworm species leading to misdiagnosis, and partly due to initial erroneous assumptions of their epidemiology [53]. While dogs are thought to be the primary source of zoonotic A. ceylanicum, there is evidence that human–human transmission can occur [11]. Two haplotypes have been identified, with the zoonotic haplotype also identified in cats [11,54]. This has public health implications, since dogs and cats will act as reservoir hosts and will thus need to be taken into consideration for control. Dogs are also the main host for A. caninum, which can cause gastric enteritis in humans, and Toxocara canis which can cause serious eye disease often resulting in blindness, as well as neurological symptoms depending on where the parasite migrates to in the body. T. cati, found in cats, can also cause similar pathology.
We review the STH, Strongyloides and Toxocara in SEA and Australia, considering their lifecycles; prevalence in SE Asia and Australia; diagnosis; and treatment and control. In addition, their zoonotic potential will be further explored.
2. Lifecycles
The lifecycles of the STH are shown in Figure 1, illustrating key differences in infection strategy and migration pathways. Adults of all STH species and S. stercoralis live in the gastrointestinal tract (GIT) and produce eggs that are excreted into the environment via the stool. STH and S. stercoralis require moist, warm soil to develop, largely restricting these parasites to tropical areas. Hookworm eggs hatch in the faecal mass and moult from L1 to infective L3 larvae. The L3 larvae then migrate onto vegetation, penetrate the skin, are carried via the blood to the lungs where they undergo tracheal migration, pass through to the small intestine, mature into adults and attach to the gut wall [55]. For both T. trichiura and A. lumbricoides, eggs are passed unembryonated and mature to an infectious stage after 15 days. The now infectious eggs are ingested, often due to poor hygiene and contaminated food, and hatch in the small intestine. Larvae of T. trichiura then mature in the small intestine into adults [56]. A. lumbricoides larvae penetrate the gut and undergo tracheal migration similar to hookworm larvae, and develop into mature adults once in the small intestine (Figure 1) [57].
The lifecycle of S. stercoralis is more complicated than other STH since it has a free-living stage in addition to the parasitic lifecycle [58]. Adults in the gut produce eggs that hatch into first-stage larvae, which have a distinctive oesophageal appearance that gives rise to the descriptive term ‘rhabditiform larva’. This first stage larva will moult to become either free-living, a dioecious adult or an infectious larva, the filariform larva. Free-living adults produce eggs that hatch as rhabditiform larvae that moult twice to become 3rd stage filariform larvae. Filariform larvae then penetrate the skin, much like hookworm L3 larvae, and migrate to the gut – this can be via the lungs and they are coughed up as occurs in the hookworm lifecycle, or direct travel to the gut. Autoinfection occurs when eggs from adult parasites hatch into rhabditiform larvae that become filariform larvae while still in the gut. The filariform larvae can complete the lifecycle in the gut, or disseminate, migrating to other organs and tissues (Figure 1). The free-living cycle for S. stercoralis only persists for one generation, not indefinitely as in Parastrongyloides sp. [59,60].
Humans are accidental hosts of Toxocara spp. and become infected by ingesting eggs in contaminated soil [61]. The eggs hatch in the gut and the larvae penetrate the gut and are carried by the blood to different organs where they can promote a local reaction, which is the cause of toxocariasis. Visceral and ocular migrans are the most common presentations. There is also a suggested link between seropositivity for toxocariasis and epilepsy [62].
3. STH and Strongyloides in SEA
STH infections are common in SEA, where approximately one-third of global STH cases occur, with active, stable transmission occurring in all countries of the region [63,64,65] (Figure 2). Risk factors for infection include poverty, lack of access to clean water and toilets, as well as unhygienic practices such as not washing hands [64].
T. trichiura, A. lumbricoides and hookworm are the most prevalent STH and infected individuals are often found with co-infections. Polyparasitism with STH is very common in SEA, mainly due to the shared geographical locations in tropical areas, where these worms are endemic. Additionally, the infection pathways of the STH are similar (Figure 1). Polyparasitism with STH is more likely than mono-infection in endemic areas, where co-infection with other helminths and protozoan parasites is also very high [66,67,68,69,70,71]. Because STH endemicity is quite low in Australia, data for polyparasitism is limited, although it may occur in remote Aboriginal communities [42].
Co-infections with parasites have been identified with increased disease status, and synergism between parasite infections. Infection intensity of helminths in co-infections also seems to differ, with hookworm infection intensity significantly increased with multiple infections [72]. Maternal infection with STH may also increase susceptibility of the unborn child to infection with STH, but it is unclear if this is due to shared environmental factors [73,74]. Co-infections may necessitate combination chemotherapy depending on drug efficacy for the infecting species.
The precise global prevalence of Strongyloides is unknown although it is estimated to infect 30–100 million people worldwide [75]. There are also issues with diagnosis because serology can return negative results, particularly in early infections, and may not pick up disseminated cases [16]. Eggs are rarely seen in the stool; rather the rhabditiform larvae are present instead, although often in low numbers. Both copro-culture, which can be laborious, and direct smears, can be used to identify larvae [76]. Other methods include the formalin ethyl acetate sedimentation method, which has low sensitivity, immunodiagnostics, and molecular methods [77]. Molecular methods, specifically PCR, have been used to sensitively diagnose Strongyloides [78,79]. Strongyloides is endemic in SEA and there are a number of reports from countries in the region using an array of different diagnostic techniques [80,81,82,83]. A comprehensive review is available of its global distribution based on community, hospital, and refugee and immigrant surveys [84] (Figure 3). Depending on the diagnostic method used, the prevalence of S. stercoralis given in Figure 3 may be an underestimate due to the low sensitivity of many of the procedures used, and the low number of larvae that are excreted, even in heavy infections [77,79,84].
4. STH and Strongyloides in Australia
Australia has a low number of STH cases, largely because these worm infections can be readily controlled by good hygiene, access to safe, clean water, and the use of toilets. The overall prevalence of hookworm and T. trichiura in the Australian Northern Territory is quite low at 0.17% and 0.65%, respectively (Table 1), based on hospital data collected between 2002 and 2012 [42,43]. These cases were primarily detected in people admitted to hospital for reasons other than hookworm or Trichuris infection; thus the true prevalence may be much higher, and may follow a more focal intensity that would not be accounted for by recording state-wide or country-wide prevalence. For both species the prevalence has been reduced; with T. trichiura, there was a drop from 123.1 cases per 100,000 in 2002 to 35.8 cases per 100,000 in 2012 [42]. The zoonotic STHs, particularly A. caninum and A. ceylanicum, are of importance in Australia since they are found in domestic and wild canids (Table 2) [85,86]. Intestinal parasites of pigs in Australia also include T. suis and A. suum, which can infect humans, although the extent of human disease is unclear [4,87].
In 1918, hookworm was considered such a serious problem in North Queensland that a five-year campaign to eradicate the disease was instigated and although considered successful, hookworm infection continued to be a problem in Aboriginal communities [53,88]. The majority of published papers on STH in Australia are relatively old, with very few published in the last 10 years. Prociv and Luke [88] provide a solid review of the early history of hookworm infections in Australia. Ascaris spp. infection has never been very prevalent despite the requisite tropical climate and moist soil existing in Australia [89].
S. stercoralis appears to be more prevalent in Australia than the other STH, or at least, there are more published data available (Table 1) (Figure 2). The parasite is endemic in tropical regions of Australia including Queensland, the Northern Territory, Western Australia, as well as Northern NSW. It is primarily found among Aboriginal people living in remote communities, with a prevalence of >60% recorded (Table 1) [90,91,92,93,94]. This worm has persisted due to lack of attention to the disease it causes, despite the potential for high morbidity and mortality in immunosuppressed individuals [94]. Its true prevalence is unknown in Australia, and probably globally. As reported by Speare et al – “if you don’t look, you won’t find” [94]. As a human-only parasite, it can be readily treated with ivermectin and eliminated from the community. A retrospective study examined indigenous Australians in Central Australia who were positive for S. stercoralis infection and who may also have been positive for human T cell lymphotropic virus type I (HTLV-I). This virus invades adult T cells, thereby reducing the effectiveness of the immune system [29]. Of these subjects, eleven (n = 18) were tested for HTLV-I, of which seven were positive. Of those who tested positive for HTLV-I, four were never treated for Strongyloides, and of those who were treated, many were not treated at initial diagnosis and infection status was not checked on subsequent visits [29]. Of these eighteen patients, fifteen died from sepsis.
A complication of disseminated strongyloidiasis is secondary bacterial infection that can become systemic, and has likely contributed to mortality in at least four cases. Further studies have found high prevalence of HTLV-I in Aboriginal communities (33.6% n = 889), which may lead to more cases of hyperinfection with S. stercoralis in the future [118,119]. Along with immune suppression due to infection with HIV or HTLV-I, and immunosuppression due to organ transplantation, treatment with steroid drugs suppresses the inflammatory response and can also result in hyperinfection [29,119,120,121,122,123,124]. There is a clear need to increase knowledge of physicians in endemic areas, perhaps as part of a database for NTDs. Speare et al [93] advocated that S. stercoralis be added to the national notifiable diseases surveillance system in Australia to help combat this sadly neglected disease. Notification would bring with it greater oversight and available information to physicians, which would help lead to better management of cases, including the provision of effective treatment. To date this has not occurred and, indeed, there are no helminth diseases on the list; malaria is the sole parasite infection listed.
5. Immigration Screening in Australia
Outside remote communities, it is primarily in immigrants from developing nations and returning travellers that STH cases occur in Australia (Table 1) (Figure 2). Current health screening for immigrants does not include testing for parasites and focuses on notifiable diseases such as tuberculosis and HIV/AIDS. Since STH infections are not notifiable, it is possible that there are autochthonous and returned traveller cases occurring in Australia that are not identified or reported.
Cross-sectional surveys have been performed on recent (1997–2000) and long-term immigrants to Australia in the East African and Cambodian communities in 2000 and 2002, and long-term immigrants from Cambodia [36] (Table 1). S. stercoralis and T. trichiura were identified in the East African cohort, with only S. stercoralis present in the Cambodian cohort. Despite having been in Australia for some years and subject to immigration screening, the high prevalence recorded, particularly in the Cambodian cohort (42%, Table 1), indicates a need to include NTDs such as STH in pre-immigration screening. Entamoeba histolytica, Hymenolepis nana, Schistosoma spp. (East African cohort), and Dientamoeba fragilis were also identified [36]. Far from being an isolated occurrence, there is a history of STH found in resettled immigrants (Table 1). An earlier study on immigrants from Laos, who had been resettled in Australia for at least 12 years prior to the survey, found S. stercoralis in 24% of participants (23/95) [37]. Strongyloides is the most commonly reported helminth infection in immigrants (Table 1). Generally S. stercoralis infections are asymptomatic. More severe complications from infection include eosinophilic pneumonia, malnutrition, and disseminated strongyloidiasis.
While screening for helminth and protozoan parasites does not occur upon entry to Australia, the government does provide reimbursement for GPs who perform heath assessments within 12 months of arrival [125]. There are also state-funded refugee services in most states and territories [126,127,128,129,130]. In theory, these services could include parasitological identification. As of June 2016, 28.5% of Australians were born overseas, with five of the top 10 countries of birth in SEA (China, India, the Philippines, Vietnam, Malaysia) [131], countries with high STH endemicity. A global distribution map for STH [64] shows stable transmission occurring in SEA and Africa, origins of many immigrants coming to Australia (Figure 2 and Figure 3).
6. Returned Service Personnel
Another cohort for Australian STH infections are Australian army veterans, including older veterans who served in STH-endemic areas from World War II onwards (Table 2) [39,40,41,132]. Strongyloides is the most commonly identified helminth infection in this cohort, possibly the only STH actually considered. In Vietnam veterans, 11.6% had positive serology in 2013 for Strongyloides, despite serving between 1962 and 1975 [40]. This shows that the adult worms can persist for many years, as highlighted earlier for immigrants who had been long-term residents in Australia still testing positive for this disease.
7. Diagnostics
Microscopy
Stool-based microscopy remains the most common diagnostic method for STH, including in Australia, with the formol ethyl acetate sedimentation technique most commonly employed [133,134]. For S. stercoralis, serology is recommended, due to low and irregular numbers of larvae excreted in the stool even in heavy infections, with stool microscopy used to rule out other infections [133,135]. Diagnosis of STH in Australia is largely done on a case-by-case basis rather than by case detection. Case detection, involving diagnostic sweeps of a community, is more likely to occur as part of a research program in endemic countries. The diagnostic method used will vary. Case-by-case studies are likely to use more sensitive, albeit more laborious, diagnostics than prevalence surveys, which examine a large number or individuals and usually necessitating faster, cheaper diagnostics such as the Kato-Katz (KK) method.
In Asia, a number of different diagnostics have been employed, often as part of specific research projects or for assessing government control programs. The main diagnostic employed is the KK method, a tool also used for diagnosis of schistosomiasis and STH in the Philippines and China. The KK procedure is cheap and easy to perform, particularly under field conditions, which are the reasons it is generally used in large-scale studies. However, the KK is known to lack sensitivity, particularly in low prevalence/intensity infections, and particularly for Strongyloides [136,137,138]. Additionally, hookworm eggs hatch rapidly after stool deposition, with the result that KK slides need to be prepared and examined quickly before eggs lyse [139,140]. FLOTAC is another microscopic, albeit more recent, technique that has been used in STH diagnostics, and has a higher sensitivity than the KK procedure [136,137,141,142,143,144]. The main disadvantage of FLOTAC is the length of time it takes to implement, with a single FLOTAC taking around 30 minutes to produce a result [145]. Other methods include the Baermann technique, which is based on the movement of larvae out of stool, the formalin-ether concentration technique (FECT), and coproculture [136]. Strongyloides has low sensitivity on stool examination so either multiple stool samples need to be examined or serology undertaken, which is the recommended diagnostic approach [77]. Other methods such as PCR, agar plate culture, and Baermann sedimentation can also be performed on stool samples, and have a higher sensitivity than microscopy [77]. Both agar plate and the Baermann technique can be time consuming. While PCR is more expensive than microscopy, agar plate, or Baermann, it achieves higher sensitivity than all of these techniques [78].
8. Immunodiagnostics
As indicated, serology is often used in Australia for diagnosis of S. stercoralis [133], with microscopy employed for the other STH. In general, this holds true for Asia as well. For hookworms, which as discussed earlier, have fragile eggs, immunodiagnostics can be a more sensitive detection method than faecal microscopy. However, lack of specificity, cross-reactivity, and the inability to distinguish between past and current infections are limitations of many immunodiagnostic tests. The most common immunodiagnostic methods used for STH detection are enzyme-linked immunosorbent assays (ELISAs), western blots, and ELISPOT [14,146,147]. Dipstick assays are rapid diagnostics that usually detect antibodies in blood to a target parasite. A dipstick developed for S. stercoralis, which is no longer available, had a similar sensitivity (91%) to the ELISA assays it was compared to and a specificity of 97.7% [148]. While serology is recommended for strongyloidiasis due to the poor sensitivity of stool examination, serology can also miss heavy infections as demonstrated by a recent fatal case from Israel of hyperinfection with S. stercoralis that was ELISA-negative [23]. An assortment of immunodiagnostics are available for Strongyloides, which have a range of sensitivities and specificities [149].
Dried blood spot (DBS) testing occurs by blotting blood samples onto filter paper. Samples can be collected and stored for later analysis, allowing for large numbers of samples to be collected but without requiring large amounts of storage space, and allows relative ease of collection because only a drop from a fingerprick is required. Dried blood spots have been used for molecular diagnostic tests, but can also be used for immunodiagnostics [46]. Most recently, the application of DBS was utilised in serology to diagnose S. stercoralis in a remote community in Northern Australia [46].
9. Molecular Diagnostics
There is a range of molecular diagnostic tests available that offer higher sensitivity than microscopy-based diagnostics, albeit with a higher price tag, and these have been reviewed elsewhere [150]. The main benefit of molecular methods is the ability to multiplex assays, that is, identify multiple species using a single assay. However, as well as being more costly than microscopy techniques, they also require specialised equipment. An exception is loop-mediated isothermal amplification (LAMP), which can be performed in the field due to much reduced equipment requirements [151,152]. However LAMP does not allow for multiplexing and to date, of the STH, has only been used to detect hookworm infections [152].
Molecular methods with multiplexing capabilities include conventional polymerase chain reaction (cPCR), real-time PCR (qPCR), multiparallel and tandem qPCR, and digital droplet PCR (ddPCR). Both cPCR and ddPCR are endpoint PCRs, which rely on designing primers that produce amplicons of different lengths to distinguish individual parasite species. ddPCR also uses fluorescent dyes, while real-time PCR reactions utilise fluorescent probes to distinguish between amplicons; the use of taqman probes can increase the sensitivity and specificity of an assay. Of these methods only ddPCR has yet to be used to diagnose STH infections, although it has been utilised for other parasites such as Schistosoma spp., detecting cell-free DNA (cfDNA) in a range of body fluids (stool, serum, urine, saliva) [153,154]. The assay provides absolute quantification and can detect very low levels of target DNA; it is also more sensitive than qPCR. While currently only providing for two channels, it is possible to multiplex the reactions for four targets by utilising different size amplicons, much as for a conventional PCR multiplex, because targets can be separated based on size. While schistosomes are blood parasites, and are thus in contact with host blood and tissues, the STH live in the gut and the potential for detection of parasite cfDNA in body fluids such as blood, urine, and saliva would likely be reduced. However, since hookworm is a blood feeder it does gain access to the host blood stream and it is possible that hookworm cfDNA would be detectable in sera. Likewise, the larvae of hookworm and Ascaris penetrate the alveolae of the lungs to be coughed up and swallowed, thereby reaching the gut. It is therefore possible that a saliva or sputum sample would yield cfDNA or the larvae themselves. Regardless, the use of ddPCR on stool samples will readily amplify target parasite genes with very high sensitivity. Because it confers absolute quantification and by partitioning the PCR mix (containing master mix, primers, and DNA) into ~20,000 droplets pre-amplification, it effectively means that each sample has ~20,000 technical replicates. Therefore there is no need to run samples in duplicate or triplicate for ddPCR as there is for qPCR. This can save on costs, although in practice the cost of ddPCR and qPCR is similar.
10. Costs of Diagnostics
The cost of a single KK slide, excluding personnel costs and stool collection costs, is US$0.30 [155]. The total cost for single and duplicate KK slides have been estimated to be $US1.73 and US$2.06, respectively, while the FLOTAC costs $2.35 [145]. In comparison, a multiplex qPCR costs $7.68. For qPCR the major costs result from DNA extraction as the qPCR assay itself costs $1.68 per sample in triplicate. A multiparallel qPCR has been costed at $1 per sample, excluding DNA extraction costs [156].
11. Treatment and Mass Drug Administration (MDA) of STH Infections
Mass drug administration (MDA) is a hallmark of many control programs aimed at controlling STH infections. The benzimidazoles (albendazole and mebendazole) are the most commonly used drugs for STH infection in humans, and are recommended for MDA. In the Philippines there is an annual deworming program among school-aged children. In 2003 the prevalence of STH in pre-school children was 66%, after which the Philippines Department of Health introduced the national school deworming program, The Integrated Helminth Control Program (IHCP) [157], with albendazole or mebendazole being recommended for use. The program has been successful in reducing overall prevalence at least in some areas, although MDA coverage can vary. Chemotherapy does not prevent re-infection, and once out of the school program there is no mandated MDA for STH treatment in adults. Fear of birth defects has also been recorded as a reason for refusing STH treatment by pregnant women in the Philippines [158]. School-based MDA has a compliance of >75% while community wide MDAs tend toward low compliance (25–65%) [158,159]. In some cases this is due to poor community involvement, but also because of concerns with possible side effects of the drugs. In the IHCP school deworming program, there was an increase in STH in at least one city (46.05% in 2007 to 56.60% in 2011), and the overall prevalence remained high at 45% as of 2011 [160]. Assuming 100% coverage, the problem of STH will still exist, since re-infection can occur very quickly after treatment, and STH eggs/larvae can live in the environment for several weeks to months, remaining viable for infection. Another issue is drug efficacy. While most available drugs for STH are highly efficacious for A. lumbricoides, there are varying efficacies for hookworm, Strongyloides spp. and Trichuris spp. Efficacy varies depending on the drug given, whether the drug is given as a single or multiple dose, and the amount given. Most programs of MDA rely on a single dose, being easier and not relying on individuals returning for treatment on multiple days. Drug efficacy for hookworm species can be difficult to untangle, as many studies do not speciate the infecting worms. There are documented differences in drug efficacy between A. duodenale and N. americanus, with mebendazole less effective against N. americanus, and pyrantel pamoate less effective against A. duodenale [161]. For all STH, drug efficacy likely varies on a regional level depending on parasite populations and treatment programs, particularly those involving MDA.
A World Health Organization (WHO) report [162] showed a range of efficacies against STH for mebendazole, albendazole, pyrantel, levamisole, and ivermectin. For A. lumbricoides, efficacy was up to 100% for all drugs except ivermectin. For hookworm the highest efficacy, in terms of cure rate (CR), was achieved with levamisole (66%–100%), and for T. trichiura with mebendazole (45%–100%). In the same report comparing differing single doses of albendazole and mebendazole (recommended by the WHO for STH control and treatment) with multiple doses of mebendazole, multiple doses resulted in a higher CR for both hookworm and Trichuris spp., albeit the median CR was still around 80% [162]. However, the WHO also recommends periodic worming with albendazole or mebendazole where prevalence is >20% [163]. The aim of the WHO with regards to STH control is to carry out preventative worming in endemic countries with a prevalence of >20%. However if drug efficacy is low, particularly for hookworm and Trichuris spp., this approach is unlikely to decrease prevalence in the long term.
Of relevance to treatment of STH cases in Australia and SEA, a study on immigrant populations (in Canada) found an overall reduction in intestinal parasites (STH and Giardia) 6 years after resettlement and treatment with thiabendazole, from 63.7% down to 21.9%; however the prevalence of S. stercoralis remained relatively high with a reduction from 15% to 11% [37,164]. However, since that report, ivermectin has been designated the drug of choice for Strongyloides spp. infection because this has a high efficacy given singly as a 200 mcg/kg dose (96% CR) or as a split 400 mcg/kg dose (98% CR) [165]. This highlights the need for combined chemotherapy when treating individuals infected with more than one species of STH.
Cure rates may be lower than reported due to insensitive diagnostics used in many drug efficacy trials, primarily stool microscopy, and variation in CR given by the same drug regimens in different trials may be due to the methods used to assess treatment success or failure. The sensitivity of diagnosis varies considerably with the more traditional microscopic diagnostics, such as the Kato-Katz procedure, lacking the sensitivity of more recently developed techniques such as real-time PCR-based diagnostics. Serological diagnosis measuring antibodies should not be used to assess CR since they will detect antibodies from previous infections for several months post-treatment, assuming 100% efficacy. Antigen-based tests are more specific but many detected antigens tend to break down quickly in the body.
In an interesting development, the bacterium Bacillus subtilis has been engineered to express the anthelmintic protein Cry5B, which proved lethal to Caenorhabditis elegans and experimental A. ceylanicum hookworm infections in hamsters [166]. This approach could be used by modifying ‘good’ bacteria that are safe for human ingestion. There are numerous probiotics on the market, of which the most common bacterial species are Lactobacillus acidophilus, L. casei, Bifidobacterium lactis, B. bifidum, and Bacillus subtilis.
Resistance to anthelmintics continues to be a concern. Currently, resistance has been reported for the human helminth, Onchocerca volvulus against ivermectin [167]. N. americanus eggs heterozygous for a β-tubulin mutation associated with resistance to benzimidazoles (albendazole, mebendazole) in A. caninum were recovered from a small number of individuals (n = 28) in Haiti [168]; homozygous eggs were not identified, but may be present. This raises the possibility of emerging benzimidazole resistance in human hookworm infections, as it has in veterinary hookworm. Drug resistance is common in helminths of veterinary importance where resistance has been reported against not only benzimidazoles but also levamisole, avermectins, and milbemycins (which include ivermectin) [169,170]. Resistance has not been reported for any of the zoonotic helminths. With the increase of MDA programs in STH-endemic areas, there is increasing evolutionary pressure on parasite populations, which may lead to resistance. To rigorously assess resistance, however, sensitive diagnostics will be required, and research needs to be undertaken in the search for alleles conferring resistance. Wolstenholme et al [170] have provided an excellent overview of drug resistance in veterinary helminths, while Vercruysse et al [171] consider the potential of resistance to currently-available drugs developing in human helminths.
12. Control Programs
STH reinfections can largely be controlled with appropriate hygiene, including washing hands after defecation, and using a toilet, as augmentation to treatment. There are several programs operating in Asia that seek to combat STH infection by using educational interventions. These include the ‘Magic Glasses’ program in China [172,173], which has now been extended to the Philippines, and the WASH (water, sanitation, and hygiene) program, which aims to provide clean water, toilets, and promoting good hygiene practices, implemented, for example, in Timor-Leste [174,175]. Helminth infection is of lesser concern in developed countries, where toilets and clean running water are available. While STH chemotherapy is effective, it does not prevent re-infections that can happen very quickly after treatment [176]. The aim of currently applied interventions is to prevent re-infection, and thus reduce the overall prevalence and eventually eliminate STH from a community.
The Magic Glasses program targets school aged children in China to reduce prevalence of STH and to increase knowledge of STH parasites to reduce re-infections occurring [172]. The focal point of the intervention is a cartoon, produced along with other teaching aids, to teach children about STH and what they can do personally and in their homes to prevent infection, including washing hands after defecation, only using a toilet, covering food, and wearing shoes. In some schools, water tanks were provided outside toilets to facilitate handwashing. Parasite prevalence and intensity levels were assessed pre-intervention and a year later post-intervention; knowledge was also tested at these times using a questionnaire and quiz. The project was highly successful in reducing re-infection and increasing knowledge in Hunan province [172], and has been further trialled in Yunnan province. It is ongoing in the Philippines, where a new video was created to match local popular cartoons, culture and language. The program has the potential to be modified for many different cultures and areas, including Australia.
WASH for WORMS has been implemented in Timor-Leste as part of a trial where intervention villages had WASH implemented alongside mass drug administration (MDA) using albendazole, while control villages only received MDA [174,175]. Intervention villages were provided with access to clean water, the building of latrines and improving hygiene, particularly handwashing. Results of this intervention and the Magic Glasses trial in Yunnan and the Philippines are currently being formulated.
In both these types of interventions community involvement was crucial. With the Magic Glasses, one of the key components was to increase STH knowledge, focusing on school children whereas the WASH program was community-wide. Targeting only children, or any one group, may not significantly impact transmission in a community, but educational interventions in schools can be delivered by teachers, with the children and teachers taking the lessons learnt back to their families, thereby increasing community knowledge and practice [177]. Paradoxically, children often have the highest helminth infection prevalence in a community [178].
13. Zoonotic Roundworms
Zoonotic Hookworms
Ancylostoma duodenale and N. americanus are responsible for the majority of human hookworm infections. However, there are two prominent zoonotic hookworm species, A. ceylanicum and A. caninum, which can also infect humans. A. ceylanicum, particularly, is gaining prominence since many cases originally identified as A. duodenale may actually be due to A. ceylanicum [51]. Both A. caninum and A. ceylanicum are found in dogs, and A. ceylanicum infects cats (Table 2). A. caninum has been identified in cats, although this is uncommon (Table 2). While human infections with A. ceylanicum do occur, there is some discussion around whether infection with this species produces hookworm disease and morbidity (Table 1) [50,179]. Table 1 shows recent cases of A. ceylanicum infection in humans and animals since 2000. Conlan et al [179] provide a review of the historical perspective of A. ceylanicum. Infection with A. ceylanicum was confirmed by molecular methods using PCR, PCR-RFLP, sequencing, or microscopy. In Australia there have been only two recent studies, including one case study of a returned peacekeeper, identifying human infection with A. ceylanicum. Studies in dogs have shown that this species is present in Australia, but to a much lesser degree than A. caninum. In the Asia-Pacific, the main hookworm species identified is N. americanus (81.8%), while 18.18% harboured A. ceylanicum, including one individual who harboured both species [132]. An equal prevalence of A. ceylanicum and N. americanus was found in Cambodia (51.6%) [96,132]. Prevalence of hookworm in animals in Asia is also high, with dual infections of A. ceylanicum and A. caninum also occurring (Table 2). Certainly, therefore, the presence of A. ceylanicum in animals in Asia and Australia poses a risk to human health.
Pets or companion animals are increasingly popular and a potential source of zoonotic hookworms. Observing good hygiene practices, wearing footwear, and regular worming of pets will help prevent transmission of zoonotic hookworms to humans. However, studies on pets and their owners in Europe, which presents a similar socio-economic situation as Australia where handwashing practices, access to clean water and toilets, are similar, have shown that pet owners do not always wash their hands after handling their pets [180]. Only 15% of dog owners and 8% of cat owners stated that they always washed their hands. The same studies have also found Toxocara eggs on the fur of the study animals (both cats and dogs) [180,181,182,183,184]. Cats pose a particular risk for egg contamination because cat litter trays reside inside the house and are cleaned by the owner, while dog owners may also be required to pick up after their animals as well.
The rate of pet ownership in Australia is very high. As of 2016, an estimated 62% of households had at least one pet, with dogs the most popular (39%) followed by cats (29%) [35]. Handwashing among Australia children was examined, with only 41% reported to always or mostly wash their hands after playing with animals, indicating that this is an issue in Australia as with comparable countries in Europe [185]. Pet ownership data are also available for China (25% dogs, 10% cats), South Korea (20% dogs, 6% cats), Japan (7% dogs, 14% cats) and Hong Kong (14% dogs, 10% cats) [35].
There have been few human infections in Australia and Asia identified as A. caninum, with the two most recent reports of human infection with this species both coming from Asia (Laos and India) (Table 2). In Australia, A. caninum has historically been associated with eosinophilic enteritis [186,187,188,189,190,191,192], although there are limited reports on this condition since the mid-1990s.
14. Toxocara
Toxocariasis is caused by the migration of Toxocara larvae to various tissues, causing visceral larva migrans. Traditionally the species involved are T. canis and T. cati nematodes of dogs and cats respectively, with the latter most likely to be involved in human disease [193]. Other Toxocara species may also cause infection such as T. malaysiensis, which is found in Malaysia, Vietnam, and China, although its potential to infect humans at this point is unknown [193,194,195]. Toxocara malaysiensis also infects cats. Stray dogs and cats are a primary source of infection in developing countries, while high pet ownership in developed countries means that pets are the main source of infection there. Soil samples taken from playgrounds in Malaysia found that 95.7% of samples tested had Toxocara eggs, and 88.3% had hookworm, showing very high contamination of the local soil with parasite eggs infectious to humans [196].
The most serious result of infection with Toxocara is ocular toxocariasis, which can lead to blindness. Humans are dead-end hosts; the parasite larvae migrate to many tissues, and while they do not develop further they can cause granulomas and inflammation in the tissues they reside in [197].
In Australia T. canis has long been known to exist with high prevalence found in dogs and in environmental samples [198,199]. There are very few reports of recent surveys of animals, environmental samples, or humans in Australia. In the 1990s the T. canis prevalence in the general population of Australia was 5.7% while in Aboriginal communities it ranged from 11.1%–43% [47]. Historical infections reported included serological examination of patients with ocular symptoms in Victoria (3.86% n = 621) [200]; 7% (n = 660) seroprevalence was recorded in healthy blood donors from the Australia Capital Territory [201]. More recently 21% (n = 29) prevalence was reported in a remote Aboriginal community in the Northern Territory [47].
15. Ascaris suum
It was originally thought that A. suum infected humans only rarely, but molecular tools have shown that the two species (A. lumbricoides and A. suum), which are morphologically identical, may in fact be one species and A. suum represents a haplotype of A. lumbricoides. It has been recognised that the haplotype, A. suum, can cause human infections; and has also been found in non-human primates [15,202]. A study in Japan sequenced the ITS1 region of Ascaris derived from humans and pigs, finding that 3 of 9 isolates derived from humans were identical to those derived from pigs [6]. A phylogeny study performed in China utilising the mitochondrial genes cox1 and nad1, and found a high level of gene flow between human- and pig-derived Ascaris, as well as indicating the presence of 20 haplotypes based on the cox1 gene and 26 based on the nad1 [203]. There is also evidence from China of hybrid forms of Ascaris [204]. In India PCR-RFLP was performed on dog stools identifying the presence of A. lumbricoides eggs [205].
A. suum has been identified in pigs in Australia; however, limited molecular work has been done to characterise the genotypes. Due to control measures in pigs it is estimated that only 3% of pigs in Australia harbour Ascaris [206]. There is also limited information on A. suum human infections in Australia. Two cases of Ascaris infection in Tasmania were thought to be A. suum based on morphology of mouthparts, and based on history of contact with pigs; no molecular tools were utilised [207].
Worldwide, 800 million people are estimated to be infected with Ascaris. A number of prevalence studies have been done in developing countries using both microscopy, primarily KK, and molecular techniques [66,208,209,210]. Speciation, or the presence of haplotypes, is rarely looked for in prevalence studies; it is therefore difficult to calculate the number of Ascaris infections due to pig-derived worms.
16. Trichuris suis and T. vulpis
Pigs and canines are the primary hosts of T. suis and T. vulpis, respectively. Human infections with both species have been recorded, as have hybrids of T. suis and the human species T. trichiura [5,9,10,13]. In Thailand, both dogs and humans harbour T. vulpis and T. trichiura, indicating the likelihood of zoonotic transmission occurring for both species [5]. However, dogs in India appear to only carry T. trichiura, while further human infections with T. vulpis have occurred in North America and Africa [13,205,211].
Genetic analysis of Trichuris species in humans, pigs, and non-human primates indicates that there may be several genotypes of Trichuris circulating in humans and animals [212,213,214]. Sequencing of pig and human derived Trichuris indicates that they are separate species [215], unlike A. suum discussed above, which is considered to be the same species as A. lumbricoides. Experimental infections of humans with T. suis documented the previously undocumented symptoms of infection which included flatulence, diarrhoea, and upper abdominal pain, although these symptoms regressed over time, after repeated exposure to eggs, to subclinical symptoms [216].
17. Helminth Therapy
While not all STH have as severe consequences as Strongyloides, it is clear that treatment of STH is needed in infected individuals. However, it has also been suggested that intestinal worms may actually be beneficial to human health, particularly to bowel health and diseases of hypersensitivity. This is usually described as part of the hygiene hypothesis, linking ‘cleaner’ living conditions with higher levels of allergic disease, and now extended to inflammatory disorders such as inflammatory bowel disease (IBD) [217]. Helminth parasites are known to modulate the host immune system and it is thought that by doing so for self-benefit, this may cause a bystander effect on other immune-related diseases [218,219].
There have been clinical trials with hookworm and Trichuris for treatment of a range of syndromes, including coeliac disease, asthma, allergies, and psoriasis [220]. Hookworm and Trichuris eggs are available online through companies such as Wormswell [221], which provides N. americanus eggs, and Tanawisa, which provides T. suis (porcine Trichuris species) (TSO) [222] eggs. The link between asthma regulation and helminths has been studied for some time, although concrete significance in trials has yet to eventuate, and exact mechanisms of action are unclear [223]. Recent trials for asthma, where individuals were experimentally infected with hookworm, indicated some improvement between groups with hookworm and those without, but the results were not significant [220]. A study in Uganda showed maternal hookworm infections decreased the risk of childhood eczema [224]. Australian studies of hookworm therapy largely focus on coeliac disease, which has had mixed results with null or positive associations; any causal link between allergies and parasite infection has not been shown [225,226]. Gut microbiota may also play a role, although it is still unclear how parasites and the microbiota interact with each other [227]. While T. suis is primarily a parasite of pigs, it can establish patent infections in humans [9,10,228] and treatment with TSO has been linked to improvement of psoriasis [229].
So, whereas STH and Strongyloides are generally thought to be detrimental to health, the presence of a few worms might be beneficial, although more research needs to be performed to confirm this. Because of the risk of self-infection with helminth eggs, most current work aimed at clinical trials is focused on the isolation of particular components in worm secretions that provide beneficial protection or immune modulation of IBD diseases [230,231].
18. Discussion and Conclusion
While STH infections are generally low in Australia, they are still endemic and of significant health importance in remote Aboriginal communities, particularly S. stercoralis, which can be fatal in immunocompromised people. Limited data are available for hyperinfection and mortality due to strongyloidiasis in the Asia-Pacific area, but in Australia, infection has led to fatalities, which, in such a resource-rich country, is alarming and unacceptable. Part of the problem may be due to a general lack of awareness of worm infections, which have a low prevalence in the country as a whole. A physician in an urban area may never see, diagnose or treat a helminthiasis case [93,94]. In Aboriginal communities, where the prevalence of strongyloidiasis can be quite high (0.25%–59.5%) [90,91,92,94,232,233], the very high cost of healthcare delivery limits available services, and physicians may only be resident for a short time so that knowledge about STH and S. stercoralis, in particular, may not be being passed on or recorded in the hospital system. Adding STH, particularly especially S. stercoralis, to the national notifiable disease system would help increase knowledge and provide access to more information about the appropriate handling of these parasitic worm infections.
High prevalence of STH in refugees and immigrants from endemic areas, particularly the Asia-Pacific, where the majority of immigrants to Australia now originate, highlights a need for better and more comprehensive health screenings of these groups that include parasite diagnosis and treatment. This holds true for army veterans and current members who have served, or are serving, in STH-endemic areas, including long-term follow-ups in case of poor drug efficacy or ongoing autoinfection in the case of S. stercoralis. The Asia-Pacific area has particularly high endemicity for STH and, with so much movement between Asia-Pacific and Australia, better understanding and treatment of STH infections are key from a public health perspective.
Author Contributions
CAG wrote the first draft of the manuscript, CAG, DPM, MJ, JK, and DJG contributed to editing and rewrites. All authors have read and approved the final paper.
Conflicts of Interest
The authors declare no conflict of interest.
List of Abbreviations
| STH | soil transmitted helminths |
| SEA | South-East Asia |
| DNA | deoxyribonucleic acid |
| qPCR | quantitative real-time polymerase chain reaction |
| cPCR | conventional polymerase chain reaction |
| ddPCR | digital droplet polymerase chain reaction |
| RFLP-PCR | restriction fragment length polymorphism polymerase chain reaction |
| Cox 1 | cytochrome oxidase 1 |
| LAMP | Loop-mediated isothermal amplification |
| GIT | gastrointestinal tract |
| WASH | water, sanitation, and hygiene |
| MDA | mass drug administration |
| NTDs | neglected tropical diseases |
| DALYs | disability adjusted life years |
| YLDs | years lived with disability |
| HTLV-I | human T cell lymphotropic virus type I |
| CR | cure rate |
| IHCP | the integrated helminth control program |
| WHO | World Health Organization |
| IBD | inflammatory bowel disease |
| KK | Kato-Katz |
| ELISA | enzyme linked immunosorbent assay |
| DBS | dried blood spot |
| TOS | Trichuris suis eggs |
References
- Bethony, J.; Brooker, S.; Alboico, M.; Geirger, S.M.; Loukas, A.; Diemart, D.; Hotez, P.J. Soil-transmitted helminth infections: Ascariasis, trichuriasis, and hookworm. Lancet 2006, 367, 1521–1532. [Google Scholar] [CrossRef]
- Utzinger, J.; Keiser, J. Schistosomiasis and soil-transmitted helminthiasis: Common drugs for treatment and control. Expert Opin. Pharmacother. 2004, 5, 263–285. [Google Scholar] [CrossRef] [PubMed]
- WHO; UNICEF. Prevention and Control of Schistosomiasis and Soil-Transmitted Helminthiasis; WHO: Geneva, Switzerland, 2004. [Google Scholar]
- Gordon, C.A.; McManus, D.P.; Jones, M.K.; Gray, D.J.; Gobert, G.N. The increase of exotic zoonotic helminth infections: The impact of urbanization, climate change and globalization. Adv. Parasitol. 2016, 91, 311–397. [Google Scholar] [PubMed]
- Areekul, P.; Putaporntip, C.; Pattanawong, U.; Sitthicharoenchai, P.; Jongwutiwes, S. Trichuris vulpis and T. trichiura infections among schoolchildren of a rural community in northwestern Thailand: The possible role of dogs in disease transmission. Asian Biomed. 2010, 4, 49–60. [Google Scholar]
- Arizono, N.; Yoshimura, Y.; Tohzaka, N.; Yamada, M.; Tegoshi, T.; Onishi, K.; Uchikawa, R. Ascariasis in Japan: Is pig-derived Ascaris infecting humans? Jpn. J. Infect. Dis. 2010, 63, 447–448. [Google Scholar] [PubMed]
- Betson, M.; Nejsum, P.; Bendall, R.P.; Deb, R.M.; Stothard, J.R. Molecular epidemiology of ascariasis: A global perspective on the transmission dynamics of Ascaris in people and pigs. J. Infect. Dis. 2014, 210, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Dutto, M.; Petrosillo, N. Hybrid ascaris suum/lumbricoides (ascarididae) infestation in a pig farmer: A rare case of zoonotic ascariasis. Cent. Eur. J. Public Health 2013, 21, 224–226. [Google Scholar] [PubMed]
- Cutillas, C.; Callejon, R.; de Rojas, M.; Tewes, B.; Ubeda, J.M.; Ariza, C.; Guevara, D.C. Trichuris suis and Trichuris trichiura are different nematode species. Acta Trop. 2009, 111, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Nissen, S.; Al-Jubury, A.; Hansen, T.V.; Olsen, A.; Christensen, H.; Thamsborg, S.M.; Nejsum, P. Genetic analysis of Trichuris suis and Trichuris trichiura recovered from humans and pigs in a sympatric setting in Uganda. Vet. Parasitol. 2012, 188, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Ngui, R.; Lim, Y.A.; Traub, R.; Mahmud, R.; Mistam, M.S. Epidemiological and genetic data supporting the transmission of Ancylostoma ceylanicum among human and domestic animals. PLoS Negl. Trop. Dis. 2012, 6, e1522. [Google Scholar] [CrossRef] [PubMed]
- Phosuk, I.; Intapan, P.M.; Thanchomnang, T.; Sanpool, O.; Janwan, P.; Laummaunwai, P.; Aamnart, W.; Morakote, N.; Maleewong, W. Molecular detection of Ancylostoma duodenale, Ancylostoma ceylanicum, and Necator americanus in humans in northeastern and southern Thailand. Korean J. Parasitol. 2013, 51, 747–749. [Google Scholar] [CrossRef] [PubMed]
- George, S.; Geldhof, P.; Albonico, M.; Ame, S.M.; Bethony, J.M.; Engels, D.; Mekonnen, Z.; Montresor, A.; Hem, S.; Tchuem-Tchuente, L.A.; et al. The molecular speciation of soil-transmitted helminth eggs collected from school children across six endemic countries. Trans. R. Soc. Trop. Med. Hyg. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bahgat, M.A.; El Gindy, A.E.; Mahmoud, L.A.; Hegab, M.H.; Shahin, A.M. Evaluation of the role of Ancylostoma caninum in humans as a cause of acute and recurrent abdominal pain. J. Egypt. Soc. Parasitol. 1999, 29, 873–882. [Google Scholar] [PubMed]
- Leles, D.; Gardner, S.L.; Reinhard, K.; Iniguez, A.; Araujo, A. Are Ascaris lumbricoides and Ascaris suum a single species? Parasit. Vectors 2012, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, A.A.; Berk, S.L. Diagnosis of Strongyloides stercoralis infection. Clin. Infect. Dis. 2001, 33, 1040–1047. [Google Scholar] [CrossRef] [PubMed]
- Genta, R.M. Global prevalence of strongyloidiasis: Critical review with epidemiologic insights into the prevention of disseminated disease. Rev. Infect. Dis. 1989, 11, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Magnaval, J.-F.; Glickman, L.T.; Dorchies, P.; Morassin, B. Highlights of human toxocariasis. Korean J. Parasitol. 2001, 39, 1–11. [Google Scholar] [CrossRef] [PubMed]
- WHO. Metrics: Disability-Adjusted Life Year (DALY). Available online: http://www.who.int/healthinfo/global_burden_disease/metrics_daly/en/ (accessed on 6 July 2017).
- Pullan, R.L.; Smith, J.L.; Jasrasaria, R.; Brooker, S.J. Global numbers of infection and disease burden of soil transmitted helminth infections in 2010. Parasit. Vectors 2014, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Vos, T.; Lozano, R.; Naghavi, M.; Flaxman, A.D.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S.; et al. Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2197–2223. [Google Scholar] [CrossRef]
- Murray, C.J.; Barber, R.M.; Foreman, K.J.; Ozgoren, A.A.; Abd-Allah, F.; Abera, S.F.; Aboyans, V.; Abraham, J.P.; Abubakar, I.; Abu-Raddad, L.J.; et al. Global, regional, and national disability-adjusted life years (DALYs) for 306 diseases and injuries and healthy life expectancy (HALE) for 188 countries, 1990–2013: Quantifying the epidemiological transition. Lancet 2015. [Google Scholar] [CrossRef]
- Osiro, S.; Hamula, C.; Glaser, A.; Rana, M.; Dunn, D. A case of Strongyloides hyperinfection syndrome in the setting of persistent eosinophilia but negative serology. Diagn. Microbiol. Infect. Dis. 2017, 88, 168–170. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, K.G. Anaesthesia and Ascaris pneumonia (Loeffler’s syndrome). Indian J. Anaesth. 2015, 59, 125–126. [Google Scholar] [CrossRef] [PubMed]
- Cataño, J.C.; Pinzón, M.A. Strongyloides Pneumonia. Am. J. Trop. Med. Hyg. 2012, 87, 195. [Google Scholar] [CrossRef] [PubMed]
- Cheepsattayakorn, A.; Cheepsattayakorn, R. Parasitic pneumonia and lung involvement. BioMed Res. Int. 2014, 2014, 18. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.K.; Holland, C.V.; Loxton, K.; Barghouth, U. Cerebral toxocariasis: Silent progression to neurodegenerative disorders? Clin. Microbiol. Rev. 2015, 28, 663–686. [Google Scholar] [CrossRef] [PubMed]
- Keiser, P.B.; Nutman, T.B. Strongyloides stercoralis in the immunocompromised population. Clin. Microbiol. Rev. 2004, 17, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Einsiedel, L.; Fernandes, L. Strongyloides stercoralis: A cause of morbidity and mortality for indigenous people in Central Australia. Intern. Med. J. 2008, 38, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Jaleta, T.G.; Zhou, S.; Bemm, F.M.; Schar, F.; Khieu, V.; Muth, S.; Odermatt, P.; Lok, J.B.; Streit, A. Different but overlapping populations of Strongyloides stercoralis in dogs and humans - dogs as a possible source for zoonotic strongyloidiasis. PLoS Negl. Trop. Dis. 2017, 11, e0005752. [Google Scholar] [CrossRef] [PubMed]
- Kouassi, R.Y.; McGraw, S.W.; Yao, P.K.; Abou-Bacar, A.; Brunet, J.; Pesson, B.; Bonfoh, B.; N’Goran, E.K.; Candolfi, E. Diversity and prevalence of gastrointestinal parasites in seven non-human primates of the Tai National Park, Cote d’Ivoire. Parasite 2015, 22, 1. [Google Scholar] [CrossRef] [PubMed]
- Thanchomnang, T.; Intapan, P.M.; Sanpool, O.; Rodpai, R.; Tourtip, S.; Yahom, S.; Kullawat, J.; Radomyos, P.; Thammasiri, C.; Maleewong, W. First molecular identification and genetic diversity of Strongyloides stercoralis and Strongyloides fuelleborni in human communities having contact with long-tailed macaques in Thailand. Parasitol. Res. 2017, 116, 1917–1923. [Google Scholar] [CrossRef] [PubMed]
- Bergquist, R.; Yang, G.J.; Knopp, S.; Utzinger, J.; Tanner, M. Surveillance and response: Tools and approaches for the elimination stage of neglected tropical diseases. Acta Trop. 2015, 141, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Nagayasu, E.; Aung, M.; Hortiwakul, T.; Hino, A.; Tanaka, T.; Higashiarakawa, M.; Olia, A.; Taniguchi, T.; Win, S.M.T.; Ohashi, I.; et al. A possible origin population of pathogenic intestinal nematodes, Strongyloides stercoralis, unveiled by molecular phylogeny. Sci. Rep. 2017, 7, 4844. [Google Scholar] [CrossRef] [PubMed]
- Pet Ownership Statistics; Animal Medicines Australia: Barton ACT, Australia, 2016.
- Caruana, S.R.; Kelly, H.A.; Ngeow, J.Y.; Ryan, N.J.; Bennett, C.M.; Chea, L.; Nuon, S.; Bak, N.; Skull, S.A.; Biggs, B.A. Undiagnosed and potentially lethal parasite infections among immigrants and refugees in Australia. J. Travel Med. 2006, 13, 233–239. [Google Scholar] [CrossRef] [PubMed]
- De Silva, S.; Saykao, P.; Kelly, H.; MacIntyre, C.R.; Ryan, N.; Leydon, J.; Biggs, B.A. Chronic Strongyloides stercoralis infection in Laotian immigrants and refugees 7–20 years after resettlement in Australia. Epidemiol. Infect. 2002, 128, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Einsiedel, L.; Spelman, D. Strongyloides stercoralis: Risks posed to immigrant patients in an Australian tertiary referral centre. Intern. Med. J. 2006, 36, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Speare, R.; Bradbury, R.S.; Croese, J. A case of Ancylostoma ceylanicum infection occurring in an Australian soldier returned from Solomon Islands. Korean J. Parasitol. 2016, 54, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Rahmanian, H.; MacFarlane, A.C.; Rowland, K.E.; Einsiedel, L.J.; Neuhaus, S.J. Seroprevalence of Strongyloides stercoralis in a South Australian Vietnam veteran cohort. Aust. N. Z. J. Public Health 2015, 39, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Pattison, D.A.; Speare, R. Strongyloidiasis in personnel of the Regional Assistance Mission to Solomon Islands (RAMSI). Med. J. Aust. 2008, 189, 203–206. [Google Scholar] [PubMed]
- Crowe, A.L.; Smith, P.; Ward, L.; Currie, B.J.; Baird, R. Decreasing prevalence of Trichuris trichiura (whipworm) in the Northern Territory from 2002 to 2012. Med. J. Aust. 2014, 200, 286–289. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Majumdar, S.S.; Forbes, R.T.; Smith, P.; Currie, B.J.; Baird, R.W. Hookworm in the Northern Territory: Down but not out. Med. J. Aust. 2013, 198, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Chaves, N.J.; Gibney, K.B.; Leder, K.; O’Brien, D.P.; Marshall, C.; Biggs, B.-A. Screening practices for infectious diseases among Burmese refugees in Australia. Emerg. Infect. Dis. 2009, 15, 1769–1772. [Google Scholar] [CrossRef] [PubMed]
- Biggs, B.A.; Caruana, S.; Mihrshahi, S.; Jolley, D.; Leydon, J.; Chea, L.; Nuon, S. Management of chronic strongyloidiasis in immigrants and refugees: Is serologic testing useful? Am. J. Trop. Med. Hyg. 2009, 80, 788–791. [Google Scholar] [PubMed]
- Mounsey, K.; Kearns, T.; Rampton, M.; Llewellyn, S.; King, M.; Holt, D.; Currie, B.J.; Andrews, R.; Nutman, T.; McCarthy, J. Use of dried blood spots to define antibody response to the Strongyloides stercoralis recombinant antigen NIE. Acta Trop. 2014, 138, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Shield, J.; Aland, K.; Kearns, T.; Gongdjalk, G.; Holt, D.; Currie, B.; Prociv, P. Intestinal parasites of children and adults in a remote Aboriginal community of the Northern Territory, Australia, 1994. West. Pac. Surveill. Response J. 2015, 6, 44–51. [Google Scholar] [CrossRef]
- Lim, L.; Biggs, B.A. Fatal disseminated strongyloidiasis in a previously-treated patient. Med. J. Aust. 2001, 174, 355–356. [Google Scholar] [PubMed]
- Koehler, A.V.; Bradbury, R.S.; Stevens, M.A.; Haydon, S.R.; Jex, A.R.; Gasser, R.B. Genetic characterization of selected parasites from people with histories of gastrointestinal disorders using a mutation scanning-coupled approach. Electrophoresis 2013, 34, 1720–1728. [Google Scholar] [CrossRef] [PubMed]
- Traub, R.J. Ancylostoma ceylanicum, a re-emerging but neglected parasitic zoonosis. Int. J. Parasitol. 2013, 43, 1009–1015. [Google Scholar] [CrossRef] [PubMed]
- Traub, R.J.; Inpankaew, T.; Sutthikornchai, C.; Sukthana, Y.; Thompson, R.C.A. PCR-based coprodiagnostic tools reveal dogs as reservoirs of zoonotic ancylostomiasis caused by Ancylostoma ceylanicum in temple communities in Bangkok. Vet. Parasitol. 2008, 155, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Schär, F.; Inpankaew, T.; Traub, R.J.; Khieu, V.; Dalsgaard, A.; Chimnoi, W.; Chhoun, C.; Sok, D.; Marti, H.; Muth, S.; et al. The prevalence and diversity of intestinal parasitic infections in humans and domestic animals in a rural Cambodian village. Parasitol. Int. 2014, 63, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, R.; Traub, R.J. Hookworm Infection in Oceania. In Neglected Tropical Diseases—Oceania; Loukas, A., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 33–68. [Google Scholar]
- Ngui, R.; Mahdy, M.A.; Chua, K.H.; Traub, R.; Lim, Y.A. Genetic characterization of the partial mitochondrial cytochrome oxidase c subunit I (cox 1) gene of the zoonotic parasitic nematode, Ancylostoma ceylanicum from humans, dogs and cats. Acta Trop. 2013, 128, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Hookworm biology. Available online: https://www.cdc.gov/parasites/hookworm/biology.html (accessed on 18 August 2017).
- Whipworm biology. Available online: https://www.cdc.gov/parasites/whipworm/biology.html (accessed on 18 August 2017).
- Ascariasis biology. Available online: https://www.cdc.gov/parasites/ascariasis/biology.html (accessed on 18 August 2017).
- Strongyloidiasis biology. Available online: https://www.cdc.gov/dpdx/strongyloidiasis/index.html (accessed on 18 August 2017).
- Yamada, M.; Matsuda, S.; Nakazawa, M.; Arizono, N. Species-specific differences in heterogonic development of serially transferred free-living generations of Strongyloides planiceps and Strongyloides stercoralis. J. Parasitol. 1991, 77, 592–594. [Google Scholar] [CrossRef] [PubMed]
- Viney, M.E.; Lok, J.B. The biology of Strongyloides spp. WormBook Online Rev. C Elegans Biol. 2015, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Toxocariasis biology. Available online: https://www.cdc.gov/parasites/toxocariasis/biology.html (accessed on 18 August 2017).
- Quattrocchi, G.; Nicoletti, A.; Marin, B.; Bruno, E.; Druet-Cabanac, M.; Preux, P.M. Toxocariasis and epilepsy: Systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2012, 6, e1775. [Google Scholar] [CrossRef] [PubMed]
- Jex, A.R.; Lim, Y.A.; Bethony, J.M.; Hotez, P.J.; Young, N.D.; Gasser, R.B. Soil-transmitted helminths of humans in Southeast Asia - towards integrated control. Adv. Parasitol. 2011, 74, 231–265. [Google Scholar] [PubMed]
- Brooker, S.J.; Bundy, D.A.P. 55—Soil-transmitted Helminths (Geohelminths). In Manson’s Tropical Infectious Diseases (Twenty-Third Edition); Farrar, J., Hotez, P.J., Junghanss, T., Kang, G., Lalloo, D., White, N.J., Eds.; W.B. Saunders: London, UK, 2014. [Google Scholar]
- De Silva, N.R.; Brooker, S.; Hotez, P.J.; Montresor, A.; Engels, D.; Savioli, L. Soil-transmitted helminth infections: Updating the global picture. Trends Parasitol. 2003, 19, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.A.; McManus, D.P.; Acosta, L.P.; Olveda, R.; Williams, M.; Ross, A.G.; Gray, D.J.; Gobert, G.N. Multiplex real-time PCR monitoring of intestinal helminths in humans reveals widespread polyparasitism in Northern Samar, the Philippines. Int. J. Parasitol. 2015, 45, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Laymanivong, S.; Hangvanthong, B.; Keokhamphavanh, B.; Phommasansak, M.; Phinmaland, B.; Sanpool, O.; Maleewong, W.; Intapan, P.M. Current status of human hookworm infections, ascariasis, trichuriasis, schistosomiasis mekongi and other trematodiases in Lao People’s Democratic Republic. Am. J. Trop. Med. Hyg. 2014, 90, 667–669. [Google Scholar] [CrossRef] [PubMed]
- Rim, H.-J.; Chai, J.-Y.; Min, D.-Y.; Cho, S.; Eom, K.S.; Hong, S.; Sohn, W.; Yong, T.; Deodato, G.; Standgaard, H.; et al. Prevalence of intestinal parasite infections on a national scale among primary schoolchildren in Laos. Parasitol. Res. 2003, 91, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Conlan, J.V.; Khamlome, B.; Vongxay, K.; Elliot, A.; Pallant, L.; Sripa, B.; Blacksell, S.D.; Fenwick, S.; Thompson, R.C. Soil-transmitted helminthiasis in Laos: A community-wide cross-sectional study of humans and dogs in a mass drug administration environment. Am. J. Trop. Med. Hyg. 2012, 86, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Meurs, L.; Polderman, A.M.; Vinkeles Melchers, N.V.; Brienen, E.A.; Verweij, J.J.; Groosjohan, B.; Mendes, F.; Mechendura, M.; Hepp, D.H.; Langenberg, M.C.; et al. Diagnosing polyparasitism in a high-prevalence setting in Beira, Mozambique: Detection of intestinal parasites in fecal samples by microscopy and real-time PCR. PLoS Negl. Trop. Dis. 2017, 11, e0005310. [Google Scholar] [CrossRef] [PubMed]
- Elyana, F.N.; Al-Mekhlafi, H.M.; Ithoi, I.; Abdulsalam, A.M.; Dawaki, S.; Nasr, N.A.; Atroosh, W.M.; Abd-Basher, M.H.; Al-Areeqi, M.A.; Sady, H.; et al. A tale of two communities: Intestinal polyparasitism among Orang Asli and Malay communities in rural Terengganu, Malaysia. Parasit. Vectors 2016, 9, 398. [Google Scholar] [CrossRef] [PubMed]
- Fleming, F.M.; Brooker, S.; Geiger, S.M.; Caldas, I.R.; Correa-Oliveira, R.; Hotez, P.J.; Bethony, J.M. Synergistic associations between hookworm and other helminth species in a rural community in Brazil. Trop. Med. Int. Health 2006, 11, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.S.; Rodriguez, A.; Chico, M.; Guadalupe, I.; Broncano, N.; Sandoval, C.; Tupiza, F.; Mitre, E.; Cooper, P.J. Maternal geohelminth infections are associated with an increased susceptibility to geohelminth infection in children: A case-control study. PLoS Negl. Trop. Dis. 2012, 6, e1753. [Google Scholar] [CrossRef] [PubMed]
- Menzies, S.K.; Rodriguez, A.; Chico, M.; Sandoval, C.; Broncano, N.; Guadalupe, I.; Cooper, P.J. Risk Factors for soil-transmitted helminth infections during the first 3 years of life in the tropics; findings from a birth cohort. PLoS Negl. Trop. Dis. 2014, 8, e2718. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.; van Lieshout, L.; Marti, H.; Polderman, T.; Polman, K.; Steinmann, P.; Stothard, R.; Thybo, S.; Verweij, J.J.; Magnussen, P. Strongyloidiasis--the most neglected of the neglected tropical diseases? Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Ngui, R.; Halim, N.A.; Rajoo, Y.; Lim, Y.A.; Ambu, S.; Rajoo, K.; Chang, T.S.; Woon, L.C.; Mahmud, R. Epidemiological characteristics of strongyloidiasis in inhabitants of indigenous communities in Borneo island, Malaysia. Korean J. Parasitol. 2016, 54, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Requena-Mendez, A.; Chiodini, P.; Bisoffi, Z.; Buonfrate, D.; Gotuzzo, E.; Munoz, J. The laboratory diagnosis and follow up of strongyloidiasis: A systematic review. PLoS Negl. Trop. Dis. 2013, 7, e2002. [Google Scholar] [CrossRef] [PubMed]
- Verweij, J.J.; Canales, M.; Polman, K.; Ziem, J.; Brienen, E.A.T.; Polderman, A.M.; van Lieshout, L. Molecular diagnosis of Strongyloides stercoralis in faecal samples using real-time PCR. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Intapan, P.M.; Maleewong, W.; Wongsaroj, T.; Singthong, S.; Morakote, N. Comparison of the quantitative formalin ethyl acetate concentration technique and agar plate culture for diagnosis of human strongyloidiasis. J. Clin. Microbiol. 2005, 43, 1932–1933. [Google Scholar] [CrossRef] [PubMed]
- Boonjaraspinyo, S.; Boonmars, T.; Kaewsamut, B.; Ekobol, N.; Laummaunwai, P.; Aukkanimart, R.; Wonkchalee, N.; Juasook, A.; Sriraj, P. A cross-sectional study on intestinal parasitic infections in rural communities, northeast Thailand. Korean J. Parasitol. 2013, 51, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Kling, K.; Kuenzli, E.; Blum, J.; Neumayr, A. Acute strongyloidiasis in a traveller returning from South East Asia. Travel Med. Infect. Dis. 2016, 14, 535–536. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Cheong, F.W.; Liew, J.W.; Lau, Y.L. Seroprevalence of fascioliasis, toxocariasis, strongyloidiasis and cysticercosis in blood samples diagnosed in Medic Medical Center Laboratory, Ho Chi Minh City, Vietnam in 2012. Parasit. Vectors 2016, 9, 486. [Google Scholar] [CrossRef] [PubMed]
- Nontasut, P.; Muennoo, C.; Sa-nguankiat, S.; Fongsri, S.; Vichit, A. Prevalence of Strongyloides in Northern Thailand and treatment with ivermectin vs albendazole. Southeast Asian J. Trop. Med. Public Health 2005, 36, 442–444. [Google Scholar] [PubMed]
- Schär, F.; Trostdorf, U.; Giardina, F.; Khieu, V.; Muth, S.; Marti, H.; Vounatsou, P.; Odermatt, P. Strongyloides stercoralis: Global distribution and risk factors. PLoS Negl. Trop. Dis. 2013, 7, e2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smout, F.A.; Skerratt, L.F.; Butler, J.R.A.; Johnson, C.N.; Congdon, B.C.; Thompson, R.C.A. The hookworm Ancylostoma ceylanicum: An emerging public health risk in Australian tropical rainforests and Indigenous communities. One Health (Amsterdam, Netherlands) 2017, 3, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Smout, F.A.; Thompson, R.C.A.; Skerratt, L.F. First report of Ancylostoma ceylanicum in wild canids. Int. J. Parasitol. Parasites Wildl. 2013, 2, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Lee, A. Internal parasites of pigs. Available online: https://www.dpi.nsw.gov.au/__data/assets/pdf_file/0019/433018/internal-parasites-of-pigs.pdf (accessed on 4 July 2017).
- Prociv, P.; Luke, R.A. The changing epidemiology of human hookworm infection in Australia. Med. J. Aust. 1995, 162, 150–154. [Google Scholar] [PubMed]
- Health, V. Ascariasis (roundworm infection). Available online: https://www2.health.vic.gov.au/public-health/infectious-diseases/disease-information-advice/ascariasis-roundworm-infection (accessed on 27 July 2017).
- Johnston, F.H.; Morris, P.S.; Speare, R.; McCarthy, J.; Currie, B.; Ewald, D.; Page, W.; Dempsey, K. Strongyloidiasis: A review of the evidence for Australian practitioners. Aust. J. Rural Health 2005, 13, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.I. Intestinal parasite infections in Western Australian Aborigines. Med. J. Aust. 1980, 2, 375–380. [Google Scholar] [PubMed]
- Prociv, P.; Luke, R. Observations on strongyloidiasis in Queensland aboriginal communities. Med. J. Aust. 1993, 158, 160–163. [Google Scholar] [PubMed]
- Speare, R.; Miller, A.; Page, W.A. Strongyloidiasis: A case for notification in Australia? Med. J. Aust. 2015, 202, 523–524. [Google Scholar] [CrossRef] [PubMed]
- Page, W.; Speare, R. Chronic strongyloidiasis—Don’t look and you won’t find. Aust. Fam. Physician 2016, 45, 40–44. [Google Scholar] [PubMed]
- Hsu, Y.; Lin, J. Intestinal Infestation with Ancylostoma ceylanicum. N. Engl. J. Med. 2012, 366. [Google Scholar] [CrossRef] [PubMed]
- Inpankaew, T.; Schar, F.; Dalsgaard, A.; Khieu, V.; Chimnoi, W.; Chhoun, C.; Sok, D.; Marti, H.; Muth, S.; Odermatt, P.; et al. High prevalence of Ancylostoma ceylanicum hookworm infections in humans, Cambodia, 2012. Emerg. Infect. Dis. 2014, 20, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zheng, G.; Alsarakibi, M. The zoonotic risk of Ancylostoma ceylanicum isolated from stray dogs and cats in Guangzhou, South China. BioMed Res. Int. 2014, 2014, 208759. [Google Scholar] [CrossRef] [PubMed]
- Mahdy, M.A.; Lim, Y.A.; Ngui, R.; Siti Fatimah, M.R.; Choy, S.H.; Yap, N.J.; Al-Mekhlafi, H.M.; Ibrahim, J.; Surin, J. Prevalence and zoonotic potential of canine hookworms in Malaysia. Parasit. Vectors 2012, 5, 88. [Google Scholar] [CrossRef] [PubMed]
- Mohd Zain, S.N.; Sahimin, N.; Pal, P.; Lewis, J.W. Macroparasite communities in stray cat populations from urban cities in Peninsular Malaysia. Vet. Parasitol. 2013, 196, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Ngui, R.; Lim, Y.A.; Chua, K.H. Rapid detection and identification of human hookworm infections through high resolution melting (HRM) analysis. PLoS ONE 2012, 7, e41996. [Google Scholar] [CrossRef] [PubMed]
- Ngui, R.; Lim, Y.A.; Ismail, W.H.; Lim, K.N.; Mahmud, R. Zoonotic Ancylostoma ceylanicum infection detected by endoscopy. Am. J. Trop. Med. Hyg. 2014, 91, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Pa Pa Aung, W.; Htoon, T.T.; Tin, H.H.; Sanpool, O.; Jongthawin, J.; Sadaow, L.; Phosuk, I.; Ropai, R.; Intapan, P.M.; Maleewong, W. First molecular identifications of Necator americanus and Ancylostoma ceylanicum infecting rural communities in lower Myanmar. Am. J. Trop. Med. Hyg. 2017, 96, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.S.; Traub, R.J.; Robertson, I.D.; Hobbs, R.P.; Elliot, A.; While, L.; Rees, R.; Thompson, R.C. The veterinary and public health significance of hookworm in dogs and cats in Australia and the status of A. ceylanicum. Vet. Parasitol. 2007, 145, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Traub, R.J.; Pednekar, R.P.; Cuttell, L.; Porter, R.B.; Abd Megat Rani, P.A.; Gatne, M.L. The prevalence and distribution of gastrointestinal parasites of stray and refuge dogs in four locations in India. Vet. Parasitol. 2014, 205, 233–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traub, R.J.; Robertson, I.D.; Irwin, P.; Mencke, N.; Thompson, R.C. Application of a species-specific PCR-RFLP to identify Ancylostoma eggs directly from canine faeces. Vet. Parasitol. 2004, 123, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Tun, S.; Ithoi, I.; Mahmud, R.; Samsudin, N.I.; Kek Heng, C.; Ling, L.Y. Detection of helminth eggs and identification of hookworm species in stray cats, dogs and soil from Klang Valley, Malaysia. PLoS ONE 2015, 10, e0142231. [Google Scholar] [CrossRef] [PubMed]
- Kaya, D.; Yoshikawa, M.; Nakatani, T.; Tomo-Oka, F.; Fujimoto, Y.; Ishida, K.; Fujinaga, Y.; Aihara, Y.; Nagamatsu, S.; Matsuo, E.; et al. Ancylostoma ceylanicum hookworm infection in Japanese traveler who presented chronic diarrhea after return from Lao People’s Democratic Republic. Parasitol. Int. 2016, 65, 737–740. [Google Scholar] [CrossRef] [PubMed]
- George, S.; Levecke, B.; Kattula, D.; Velusamy, V.; Roy, S.; Geldhof, P.; Sarkar, R.; Kang, G. Molecular identification of hookworm isolates in humans, dogs and soil in a tribal area in Tamil Nadu, India. PLoS Negl. Trop. Dis. 2016, 10, e0004891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pumidonming, W.; Salman, D.; Gronsang, D.; Abdelbaset, A.E.; Sangkaeo, K.; Kawazu, S.I.; Igarashi, M. Prevalence of gastrointestinal helminth parasites of zoonotic significance in dogs and cats in lower Northern Thailand. J. Vet. Med. Sci. 2017, 78, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Brunet, J.; Lemoine, J.P.; Lefebvre, N.; Denis, J.; Pfaff, A.W.; Abou-Bacar, A.; Traub, R.J.; Pesson, B.; Candolfi, E. Bloody diarrhea associated with hookworm infection in traveler returning to France from Myanmar. Emerg. Infect. Dis. 2015, 21, 1878–1879. [Google Scholar] [CrossRef] [PubMed]
- Ng-Nguyen, D.; Hii, S.F.; Nguyen, V.A.; Van Nguyen, T.; Van Nguyen, D.; Traub, R.J. Re-evaluation of the species of hookworms infecting dogs in Central Vietnam. Parasit. Vectors 2015, 8, 401. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Yu, X.G.; Wu, S.; Tan, L.P.; Song, M.R.; Abdulahi, A.Y.; Wang, Z.; Jiang, B.; Li, G.Q. Levels of Ancylostoma infections and phylogenetic analysis of cox 1 gene of A. ceylanicum in stray cat faecal samples from Guangzhou, China. J. Helminthol. 2016, 90, 392–397. [Google Scholar] [CrossRef] [PubMed]
- George, S.; Kaliappan, S.P.; Kattula, D.; Roy, S.; Geldhof, P.; Kang, G.; Vercruysse, J.; Levecke, B. Identification of Ancylostoma ceylanicum in children from a tribal community in Tamil Nadu, India using a semi-nested PCR-RFLP tool. Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 283–285. [Google Scholar] [CrossRef] [PubMed]
- Jiraanankul, V.; Aphijirawat, W.; Mungthin, M.; Khositnithikul, R.; Rangsin, R.; Traub, R.J.; Piyaraj, P.; Naaglor, T.; Taamasri, P.; Leelayoova, S. Incidence and risk factors of hookworm infection in a rural community of central Thailand. Am. J. Trop. Med. Hyg. 2011, 84, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Sanguankiat, S.; Yoonuan, T.; Pongvongsa, T.; Keomoungkhoun, M.; Phimmayoi, I.; Boupa, B.; Moji, K.; Waikagul, J. Copro-molecular identification of infections with hookworm eggs in rural Lao PDR. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Scholz, T.; Uhlirova, M.; Ditrich, O. Helminth parasites of cats from the Vientiane province, Laos, as indicators of the occurrence of causative agents of human parasitoses. Parasite 2003, 10, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.S.; Lin, C.K.; Su, K.E.; Liu, C.Y.; Lin, C.C.; Liang, C.C.; Lee, T.H. Diagnosis of Ancylostoma ceylanicum infestation by single-balloon enteroscopy (with video). Gastrointest. Endosc. 2012, 76, 671–672. [Google Scholar] [CrossRef] [PubMed]
- Einsiedel, L.J.; Pham, H.; Woodman, R.J.; Pepperill, C.; Taylor, K.A. The prevalence and clinical associations of HTLV-1 infection in a remote Indigenous community. Med. J. Aust. 2016, 205, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Einsiedel, L.; Woodman, R.J.; Flynn, M.; Wilson, K.; Cassar, O.; Gessain, A. Human T-Lymphotropic Virus type 1 infection in an Indigenous Australian population: Epidemiological insights from a hospital-based cohort study. BMC Public Health 2016, 16, 787. [Google Scholar] [CrossRef] [PubMed]
- Al Maslamani, M.A.; Al Soub, H.A.; Al Khal, A.L.M.; Al Bozom, I.A.; Abu Khattab, M.J.; Chacko, K.C. Strongyloides stercoralis hyperinfection after corticosteroid therapy: A report of two cases. Ann. Saudi Med. 2009, 29, 397–401. [Google Scholar] [PubMed]
- Koticha, A.; Kuyare, S.; Nair, J.; Athvale, A.; Mehta, P. Strongyloides stercoralis hyperinfection syndrome in patients on prolonged steroid treatment: Two case reports. J. Indian Med. Assoc. 2013, 111, 272–274. [Google Scholar] [PubMed]
- Aru, R.G.; Chilcutt, B.M.; Butt, S.; deShazo, R.D. Novel findings in HIV, immune reconstitution disease and Strongyloides stercoralis Infection. Am. J. Med. Sci. 2017, 353, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Llenas-Garcia, J.; Fiorante, S.; Salto, E.; Maseda, D.; Rodriguez, V.; Matarranz, M.; Hernando, A.; Rubio, R.; Pulido, F. Should we look for Strongyloides stercoralis in foreign-born HIV-infected persons? J. Immigr. Minor. Health 2013, 15, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Mobley, C.M.; Dhala, A.; Ghobrial, R.M. Strongyloides stercoralis in solid organ transplantation: Early diagnosis gets the worm. Curr. Opin. Organ Transplant. 2017, 22, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Medicinewise, N. Albendazole (Zentel) listing extended to treat hookworm and strongyloidiasis. Available online: https://www.nps.org.au/radar/articles/albendazole-zentel-listing-extended-to-treat-hookworm-and-strongyloidiasis (accessed on 4 July 2017).
- Refugee health services. Available online: https://www.health.qld.gov.au/public-health/groups/multicultural/refugee-services (accessed on 27 September 2017).
- Refugee and asylum seeker health and wellbeing. Available online: https://www2.health.vic.gov.au/about/populations/refugee-asylum-seeker-health (accessed on 27 September 2017).
- NSW refugee health service. Available online: https://www.swslhd.health.nsw.gov.au/refugee/ (accessed on 27 September 2017).
- Humanitarian entrant health service. Available online: http://ww2.health.wa.gov.au/Articles/F_I/Humanitarian-Entrant-Health-Service (accessed on 27 September 2017).
- Refugee Health Program. Available online: https://www.ntphn.org.au/refugee-health (accessed on 27 September 2017).
- Statistics, A.B.O. Migration, Australia, 2015–2016. Available online: http://www.abs.gov.au/ausstats/[email protected]/mf/3412.0 (accessed on 4 July 2017).
- Bradbury, R.S.; Hii, S.F.; Harrington, H.; Speare, R.; Traub, R. Ancylostoma ceylanicum hookworm in the Solomon Islands. Emerg. Infect. Dis. 2017, 23, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Hanieh, S.; Ryan, N.; Biggs, B. Assessing enteric helminths in refugees, asylum seekers and new migrants. Microbiol. Aust. 2016, 15–19. [Google Scholar] [CrossRef]
- Mayer-Coverdale, J.K.; Crowe, A.; Smith, P.; Baird, R.W. Trends in Strongyloides stercoralis faecal larvae detections in the Northern Territory, Australia: 2002 to 2012. Trop. Med. Infect. Dis. 2017, 2. [Google Scholar] [CrossRef]
- Montes, M.; Sawhney, C.; Barros, N. Strongyloides stercoralis: There but not seen. Curr. Opin. Infect. Dis. 2010, 23, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Knopp, S.; Salim, N.; Schindler, T.; Voules, D.A.K.; Rothen, J.; Lweno, O.; Mohammed, A.S.; Singo, R.; Benninghoff, M.; Nsojo, A.A.; et al. Diagnostic accuracy of Kato-Katz, FLOTAC, Baermann, and PCR methods for the detection of light-intensity hookworm and Strongyloides stercoralis infections in Tanzania. Am. J. Trop. Med. Hyg. 2014, 90, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Knopp, S.; Speich, B.; Hattendorf, J.; Rinaldi, L.; Mohammed, K.A.; Khamis, I.S.; Mohammed, A.S.; Albonico, M.; Rollinson, D.; Marti, H.; et al. Diagnostic accuracy of Kato-Katz and FLOTAC for assessing anthelmintic drug efficacy. PLoS Negl. Trop. Dis. 2011, 5, e1036. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.-D.; Liu, J.-X.; Liu, Y.-M.; Hu, F.; Zhang, Y.-Y.; Xu, J.-M.; Li, J.-Y.; Ji, M.-J.; Bergquist, R.; Wu, G.-L.; et al. Routine Kato-Katz technique underestimates the prevalence of Schistosoma japonicum: A case study in an endemic area of the People’s Republic of China. Parasitol. Int. 2008, 57, 281–286. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.S.; Lustigman, S.; Yang, G.-J.; Barakat, R.M.; García, H.H.; Sripa, B.; Willingham, A.L.; Prichard, R.K.; Basáñez, M.-G. A researcha genda for helminth diseases of humans: Diagnostics for control and elimination programmes. PLoS Negl. Trop. Dis. 2012, 6, e1601. [Google Scholar] [CrossRef] [PubMed]
- Dacombe, R.J.; Crampin, A.C.; Floyd, S.; Randall, A.; Ndhlovu, R.; Bickle, Q.; Fine, P.E.M. Time delays between patient and laboratory selectively affect accuracy of helminth diagnosis. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Cringoli, G. FLOTAC, a novel apparatus for a multivalent faecal egg count technique. Parassitologica 2006, 48, 381–384. [Google Scholar]
- Glinz, D.; Silué, K.D.; Knopp, S.; Lohouringnon, L.K.; Yao, K.P.; Steinmann, P.; Rinaldi, L.; Cringoli, G.; N’Goran, E.K.; Utzinger, J. Comparing diagnostic accuraccy of Kato-Katz, koga agar plate, ether-concentration, and FLOTAC for Schistosoma mansoni and soli-transmitted helminths. PLoS Negl. Trop. Dis. 2010, 4, e754. [Google Scholar] [CrossRef] [PubMed]
- Habtamu, K.; Degarege, A.; Ye-Ebiyo, Y.; Erko, B. Comparison of the Kato-Katz and FLOTAC techniques for the diagnosis of soil-transmitted helminth infections. Parasitol. Int. 2011, 60, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Knopp, S.; Rinaldi, R.; Khamis, I.S.; Stothard, J.R.; Rollinson, D.; Maurelli, M.P.; Steinmann, P.; Marti, H.; Cringoli, G.; Utzinger, J. A single FLOTAC is more sensitive than triplicate Kato-Katz for the diagnosis of low intensity soil-transmitted helminth infections. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Speich, B.; Knopp, S.; Mohammed, A.K.; Khamis, I.S.; Rinaldi, L.; Cringoli, G.; Rollinson, D.; Utzinger, J. Comparative cost assessment of the Kato-Katz and FLOTAC techniques for soil-transmitted helminth diagnosis in epidemiological surveys. Parasit. Vectors 2010, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buonfrate, D.; Formenti, F.; Perandin, F.; Bisoffi, Z. Novel approaches to the diagnosis of Strongyloides stercoralis infection. Clin. Microbiol. Infect. 2015, 21, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Buonfrate, D.; Perandin, F.; Formenti, F.; Bisoffi, Z. A retrospective study comparing agar plate culture, indirect immunofluorescence and real-time PCR for the diagnosis of Strongyloides stercoralis infection. Parasitology 2017, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, H.R.; Koelewijn, R.; Hofwegen, H.; Gilis, H.; Wetsteyn, J.C.; Wismans, P.J.; Sarfati, C.; Vervoort, T.; van Gool, T. Use of enzyme-linked immunosorbent assay and dipstick assay for detection of Strongyloides stercoralis infection in humans. J. Clin. Microbiol. 2007, 45, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.W.; Klein, D.M.; Dornink, S.M.; Jespersen, D.J.; Kubofcik, J.; Nutman, T.B.; Merrigan, S.D.; Couturier, M.R.; Theel, E.S. Comparison of three immunoassays for detection of antibodies to Strongyloides stercoralis. Clin. Vaccine Immunol. CVI 2014, 21, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.A.; Gray, D.J.; Gobert, G.N.; McManus, D.P. DNA amplification approaches for the diagnosis of key parasitic helminth infections of humans. Mol. Cell. Probes 2011, 25, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, T.; Furushima-Shimogawara, R.; Ohmae, H.; Wang, T.P.; Lu, S.; Chen, R.; Wen, L.; Ohta, N. Detection of early and single infections of Schistosoma japonicum in the intermediate host snail, Oncomelania hupensis, by PCR and loop-mediated isothermal amplification (LAMP) assay. Am. J. Trop. Med. Hyg. 2010, 83, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Mugambi, R.M.; Agola, E.L.; Mwangi, I.N.; Kinyua, J.; Shiraho, E.A.; Mkoji, G.M. Development and evaluation of a loop mediated isothermal amplification (LAMP) technique for the detection of hookworm (Necator americanus) infection in fecal samples. Parasit. Vectors 2015, 8, 574. [Google Scholar] [CrossRef] [PubMed]
- Weerakoon, K.G.; Gordon, C.A.; Cai, P.; Gobert, G.N.; Duke, M.; Williams, G.M.; McManus, D.P. A novel duplex ddPCR assay for the diagnosis of schistosomiasis japonica: Proof of concept in an experimental mouse model. Parasitology 2017, 144, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Weerakoon, K.G.; Gordon, C.A.; Gobert, G.N.; Cai, P.; McManus, D.P. Optimisation of a droplet digital PCR assay for the diagnosis of Schistosoma japonicum infection: A duplex approach with DNA binding dye chemistry. J. Microbiol. Methods 2016, 125, 19–27. [Google Scholar] [CrossRef] [PubMed]
- WHO. KITS; WHO: Geneva, Switzerland, 1998. [Google Scholar]
- Pilotte, N.; Papaiakovou, M.; Grant, J.R.; Bierwert, L.A.; Llewellyn, S.; McCarthy, J.S.; Williams, S.A. Improved PCR-based detection of soil transmitted helminth infections using a next-generation sequencing approach to assay design. PLoS Negl. Trop. Dis. 2016, 10, e0004578. [Google Scholar] [CrossRef] [PubMed]
- Belizario, V.Y.; Totanes, F.I.; de Leon, W.U.; Matias, K.M. School-based control of soil-transmitted helminthiasis in western Visayas, Philippines. Southeast Asian J. Trop. Med. Public Health 2014, 45, 556–567. [Google Scholar] [PubMed]
- Insetta, E.R.; Soriano, A.J.; Totanes, F.I.; Macatangay, B.J.; Belizario, V.Y., Jr. Fear of birth defects is a major barrier to soil-transmitted helminth treatment (STH) for pregnant women in the Philippines. PLoS ONE 2014, 9, e85992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, A.; Olveda, R.; Olveda, D.; Harn, D.; Gray, D.J.; McManus, D.P.; Tallo, V.; Chau, T.; Williams, G. Can mass drug administration lead to the sustainable control of schistosomiasis in the Philippines? J. Infect. Dis. 2015, 211, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Sanza, M.; Totanes, F.I.; Chua, P.L.; Belizario, V.Y., Jr. Monitoring the impact of a mebendazole mass drug administration initiative for soil-transmitted helminthiasis (STH) control in the Western Visayas Region of the Philippines from 2007 through 2011. Acta Trop. 2013, 127, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Geerts, S.; Gryseels, B. Drug resistance in human helminths: Current situation and lessons from livestock. Clin. Microbiol. Rev. 2000, 13, 207–222. [Google Scholar] [CrossRef] [PubMed]
- WHO. Monitoring Anthelmintic Efficacy for Soil Transmitted Helminths (STH); WHO: Geneva, Switzerland, 2008. [Google Scholar]
- WHO. Soil-transmitted helminthiases: Number of children treated in 2014. Wkly. Epidemiol. Rec. 2015, 90, 701–712. [Google Scholar]
- Gyorkos, T.W.; MacLean, J.D.; Viens, P.; Chheang, C.; Kokoskin-Nelson, E. Intestinal parasite infection in the Kampuchean refugee population 6 years after resettlement in Canada. J. Infect. Dis. 1992, 166, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Zaha, O.; Hirata, T.; Kinjo, F.; Saito, A.; Fukuhara, H. Efficacy of ivermectin for chronic strongyloidiasis: Two single doses given 2 weeks apart. J. Infect. Chemother. 2002, 8, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Miller, M.M.; Derman, A.I.; Ellis, B.L.; Monnerat, R.G.; Pogliano, J.; Aroian, R.V. Bacillus subtilis strain engineered for treatment of soil-transmitted helminth diseases. Appl. Environ. Microbiol. 2013, 79, 5527–5532. [Google Scholar] [CrossRef] [PubMed]
- Osei-Atweneboana, M.Y.; Awadzi, K.; Attah, S.K.; Boakye, D.A.; Gyapong, J.O.; Prichard, R.K. Phenotypic evidence of emerging ivermectin resistance in Onchocerca volvulus. PLoS Negl. Trop. Dis. 2011, 5, e998. [Google Scholar] [CrossRef] [PubMed]
- Diawara, A.; Schwenkenbecher, J.M.; Kaplan, R.M.; Prichard, R.K. Molecular and biological diagnostic tests for monitoring benzimidazole resistance in human soil-transmitted helminths. Am. J. Trop. Med. Hyg. 2013, 88, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Brockwell, Y.M.; Elliott, T.P.; Anderson, G.R.; Stanton, R.; Spithill, T.W.; Sangster, N.C. Confirmation of Fasciola hepatica resistant to triclabendazole in naturally infected Australian beef and dairy cattle. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Wolstenholme, A.J.; Fairweather, I.; Prichard, R.; von Samson-Himmelstjerna, G.; Sangster, N.C. Drug resistance in veterinary helminths. Trends Parasitol. 2004, 20, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Vercruysse, J.; Albonico, M.; Behnke, J.M.; Kotze, A.C.; Prichard, R.K.; McCarthy, J.S.; Montresor, A.; Levecke, B. Is anthelmintic resistance a concern for the control of human soil-transmitted helminths? Int. J. Parasitol. Drugs Drug Resist. 2011, 1, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Bieri, F.A.M.; Gray, D.J.P.; Williams, G.M.P.; Raso, G.P.; Li, Y.-S.P.; Yuan, L.P.; He, Y.M.P.H.; Li, R.S.B.; Guo, F.-Y.B.A.; Li, S.-M.B.A.; et al. Health-education package to prevent worm infections in Chinese schoolchildren. N. Engl. J. Med. 2013, 368, 1603–1612. [Google Scholar] [CrossRef] [PubMed]
- McManus, D.P.; Bieri, F.A.; Li, Y.S.; Williams, G.M.; Yuan, L.P.; Henglin, Y.; Du, Z.W.; Clements, A.C.; Steinmann, P.; Raso, G.; et al. Health education and the control of intestinal worm infections in China: A new vision. Parasit. Vectors 2014, 7, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, N.E.; Clements, A.C.; Bryan, S.; McGown, J.; Gray, D.; Nery, S.V. Investigating the differential impact of school and community-based integrated control programmes for soil-transmitted helminths in Timor-Leste: The (S)WASH-D for Worms pilot study protocol. Pilot Feasibility Stud. 2016, 2, 69. [Google Scholar] [CrossRef] [PubMed]
- Nery, S.V.; McCarthy, J.S.; Traub, R.; Andrews, R.M.; Black, J.A.; Gray, D.J.; Weking, E.; Atkinson, J.A.; Campbell, S.; Francis, N.; et al. A cluster-randomised controlled trial integrating a community-based water, sanitation and hygiene programme, with mass distribution of albendazole to reduce intestinal parasites in Timor-Leste: The WASH for WORMS research protocol. Br. Med. J. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Yap, P.; Du, Z.W.; Wu, F.W.; Jiang, J.Y.; Chen, R.; Zhou, X.N.; Hattendorf, J.; Utzinger, J.; Steinmann, P. Rapid re-infection with soil-transmitted helminths after triple-dose albendazole treatment of school-aged children in Yunnan, People’s Republic of China. Am. J. Trop. Med. Hyg. 2013, 89, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Mascarini-Serra, L. Prevention of soil-transmitted helminth infection. J. Glob. Infect. Dis. 2011, 3, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J.; Bundy, D.A.P.; Beegle, K.; Brooker, S.; Drake, L.; de Silva, N.; Montresor, A.; Engels, D.; Jukes, M.; Chitsulo, L.; et al. Helminth infections: soil-transmitted helminth infections and schistosomiasis. In Disease Control Priorities in Developing Countries; Jamison, D.T., Breman, J.G., Measham, A.R., Alleyne, G., Claeson, M., Evans, D.B., Jha, P., Mills, A., Musgrove, P., Eds.; World Bank, The International Bank for Reconstruction and Development/The World Bank Group: Washington, DC, USA, 2006. [Google Scholar]
- Conlan, J.V.; Sripa, B.; Attwood, S.; Newton, P.N. A review of parasitic zoonoses in a changing Southeast Asia. Vet. Parasitol. 2011, 182, 22–40. [Google Scholar] [CrossRef] [PubMed]
- Overgaauw, P.A.M.; van Zutphen, L.; Hoek, D.; Yaya, F.O.; Roelfsema, J.; Pinelli, E.; van Knapen, F.; Kortbeek, L.M. Zoonotic parasites in fecal samples and fur from dogs and cats in The Netherlands. Vet. Parasitol. 2009, 163, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Overgaauw, P.A.M.; van Knapen, F. Veterinary and public health aspects of Toxocara spp. Vet. Parasitol. 2013, 193, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Simonato, G.; Frangipane di Regalbono, A.; Cassini, R.; Traversa, D.; Beraldo, P.; Tessarin, C.; Pietrobelli, M. Copromicroscopic and molecular investigations on intestinal parasites in kenneled dogs. Parasitol. Res. 2015, 114, 1963–1970. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, B.; Traversa, D.; Iorio, R.; De Berardinis, A.; Bartolini, R.; Salini, R.; Di Cesare, A. Zoonotic parasites in feces and fur of stray and private dogs from Italy. Parasitol. Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Roddie, G.; Stafford, P.; Holland, C.; Wolfe, A. Contamination of dog hair with eggs of Toxocara canis. Vet. Parasitol. 2008, 152, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Heyworth, J.S.; Cutt, H.; Glonek, G. Does dog or cat ownership lead to increased gastroenteritis in young children in South Australia? Epidemiol. Infect. 2006, 134, 926–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, N.I.; Croese, J.; Clouston, A.D.; Parry, M.; Loukas, A.; Prociv, P. Eosinophilic enteritis in northeastern Australia. Pathology, association with Ancylostoma caninum, and implications. Am. J. Surg. Pathol. 1995, 19, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Croese, J.; Loukas, A.; Opdebeeck, J.; Prociv, P. Occult enteric infection by Ancylostoma caninum: A previously unrecognized zoonosis. Gastroenterology 1994, 106, 3–12. [Google Scholar] [CrossRef]
- Loukas, A.; Croese, J.; Opdebeeck, J.; Prociv, P. Detection of antibodies to secretions of Ancylostoma caninum in human eosinophilic enteritis. Trans. R. Soc. Trop. Med. Hyg. 1992, 86, 650–653. [Google Scholar] [CrossRef]
- Loukas, A.; Opdebeeck, J.; Croese, J.; Prociv, P. Immunologic incrimination of Ancylostoma caninum as a human enteric pathogen. Am. J. Trop. Med. Hyg. 1994, 50, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Prociv, P.; Croese, J. Human enteric infection with Ancylostoma caninum: Hookworms reappraised in the light of a ‘new’ zoonosis. Acta Trop. 1996, 62, 23–44. [Google Scholar] [CrossRef]
- Prociv, P.; Croese, J. Human eosinophilic enteritis caused by dog hookworm Ancylostoma caninum. Lancet 1990, 335, 1299–1302. [Google Scholar] [CrossRef]
- Croese, J.; Fairley, S.; Loukas, A.; Hack, J.; Stronach, P. A distinctive aphthous ileitis linked to Ancylostoma caninum. J. Gastroenterol. Hepatol. 1996, 11, 524–531. [Google Scholar] [CrossRef] [PubMed]
- McGuinness, S.L.; Leder, K. Global burden of toxocariasis: A common neglected infection of poverty. Curr. Trop. Med. Rep. 2014, 1, 52–61. [Google Scholar] [CrossRef]
- Le, T.H.; Anh, N.T.; Nguyen, K.T.; Nguyen, N.T.; Thuy do, T.T.; Gasser, R.B. Toxocara malaysiensis infection in domestic cats in Vietnam—An emerging zoonotic issue? Infect. Genet. Evol. 2016, 37, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Li, M.W.; Lin, R.Q.; Song, H.Q.; Wu, X.Y.; Zhu, X.Q. The complete mitochondrial genomes for three Toxocara species of human and animal health significance. BMC Genom. 2008, 9, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohd Zain, S.N.; Rahman, R.; Lewis, J.W. Stray animal and human defecation as sources of soil-transmitted helminth eggs in playgrounds of Peninsular Malaysia. J. Helminthol. 2015, 89, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Arevalo, J.F.; Espinoza, J.V.; Arevalo, F.A. Ocular toxocariasis. J. Pediatr. Ophthalmol. Strabismus 2013, 50, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Blake, R.T.; Overend, D.J. The prevalence of Dirofilaria immitis and other parasites in urban pound dogs in north-eastern Victoria. Aust. Vet. J. 1982, 58, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Dunsmore, J.D.; Thompson, R.C.A.; Bates, I.A. Prevalence and survival of Toxocara canis eggs in the urban environment of Perth, Australia. Vet. Parasitol. 1984, 16, 303–311. [Google Scholar] [CrossRef]
- Carden, S.M.; Meusemann, R.; Walker, J.; Stawell, R.J.; MacKinnon, J.R.; Smith, D.; Stawell, A.M.; Hall, A.J. Toxocara canis: Egg presence in Melbourne parks and disease incidence in Victoria. Clin. Exp. Ophthalmol. 2003, 31, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, W.L.; Stewart, A.C.; Walker, J.C. Toxocariasis: A serological survey of blood donors in the Australian Capital Territory together with observations on the risks of infection. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 217–221. [Google Scholar] [CrossRef]
- Nejsum, P.; Bertelsen, M.F.; Betson, M.; Stothard, J.R.; Murrell, K.D. Molecular evidence for sustained transmission of zoonotic Ascaris suum among zoo chimpanzees (Pan Troglodytes). Vet. Parasitol. 2010, 171, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Li, M.; Yuan, K.; Hu, N.; Peng, W. Phylogeography of Ascaris lumbricoides and A. suum from China. Parasitol. Res. 2011, 109, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Li, M.; Yuan, K.; Deng, S.; Peng, W. Pig Ascaris: An important source of human ascariasis in China. Infect. Genet. Evol. 2012, 12, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Traub, R.J.; Robertson, I.D.; Irwin, P.J.; Mencke, N.; Thompson, R.C.A.A. Canine gastrointestinal parasitic zoonoses in India. Trends Parasitol. 2005, 21, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Controlling worms in pigs. Available online: https://www.daf.qld.gov.au/animal-industries/pigs/pig-health-and-diseases/disease-prevention/controlling-worms-in-pigs (accessed on 11 October 2017).
- Graves, S. Human infection with ‘Ascaris suum’ in Tasmania? Ann. ACTM 2005, 6, 16. [Google Scholar]
- Steinmann, P.; Yap, P.; Utzinger, J.; Du, Z.W.; Jiang, J.Y.; Chen, R.; Wu, F.W.; Chen, J.X.; Zhou, H.; Zhou, X.N. Control of soil-transmitted helminthiasis in Yunnan province, People’s Republic of China: Experiences and lessons from a 5-year multi-intervention trial. Acta Trop. 2015, 141, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Luo, N.J. A cross-sectional study of intestinal parasitic infections in a rural district of west China. Can. J. Infect. Dis. 2003, 14, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.-J.; Liu, L.; Zhu, H.-R.; Griffiths, S.M.; Tanner, M.; Bergquist, R.; Utzinger, J.; Zhou, X.-N. China’s sustained drive to eliminate neglected tropical diseases. Lancet Infect. Dis. 2014, 14, 881–892. [Google Scholar] [CrossRef]
- Dunn, J.J.; Columbus, S.T.; Aldeen, W.E.; Davis, M.; Carroll, K.C. Trichuris vulpis recovered from a patient with chronic diarrhea and five dogs. J. Clin. Microbiol. 2002, 40, 2703–2704. [Google Scholar] [CrossRef] [PubMed]
- Ravasi, D.F.; O’Riain, M.J.; Davids, F.; Illing, N. Phylogenetic evidence that two distinct Trichuris genotypes infect both humans and non-human primates. PLoS ONE 2012, 7, e44187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawash, M.B.F.; Andersen, L.O.; Gasser, R.B.; Stensvold, C.R.; Nejsum, P. Mitochondrial genome analyses suggest multiple Trichuris species in humans, baboons, and pigs from different geographical regions. PLoS Negl. Trop. Dis. 2015, 9, e0004059. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.H.; Zhou, W.; Nisbet, A.J.; Xu, M.J.; Zhou, D.H.; Zhao, G.H.; Wang, S.K.; Song, H.Q.; Lin, R.Q.; Zhu, X.Q. Characterization of Trichuris trichiura from humans and T. suis from pigs in China using internal transcribed spacers of nuclear ribosomal DNA. J. Helminthol. 2014, 88, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.-H.; Gasser, R.B.; Su, A.; Nejsum, P.; Peng, L.; Lin, R.-Q.; Li, M.-W.; Xu, M.-J.; Zhu, X.-Q. Clear genetic distinctiveness between human- and pig-derived Trichuris based on analyses of mitochondrial datasets. PLoS Negl. Trop. Dis. 2012, 6, e1539. [Google Scholar] [CrossRef] [PubMed]
- Bager, P.; Kapel, C.; Roepstorff, A.; Thamsborg, S.; Arnved, J.; Rønborg, S.; Kristensen, B.; Poulsen, L.K.; Wohlfahrt, J.; Melbye, M. Symptoms after ingestion of pig whipworm Trichuris suis eggs in a randomized placebo-controlled double-blind clinical trial. PLoS ONE 2011, 6, e22346. [Google Scholar] [CrossRef] [PubMed]
- Helmby, H. Human helminth therapy to treat inflammatory disorders-where do we stand? BMC Immunol. 2015, 16, 12. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.A.; Friberg, I.M.; Little, S.; Bradley, J.E. Review series on helminths, immune modulation and the hygiene hypothesis: Immunity against helminths and immunological phenomena in modern human populations: Coevolutionary legacies? Immunology 2009, 126, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Hewitson, J.P.; Grainger, J.R.; Maizels, R.M. Helminth immunoregulation: The role of parasite secreted proteins in modulating host immunity. Mol. Biochem. Parasitol. 2009, 167, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Feary, J.R.; Venn, A.J.; Mortimer, K.; Brown, A.P.; Hooi, D.; Falcone, F.H.; Pritchard, D.I.; Britton, J.R. Experimental hookworm infection: A randomized placebo-controlled trial in asthma. Clin. Exp. Allergy 2010, 40, 299–306. [Google Scholar] [CrossRef] [PubMed]
- TOS. Available online: http://wormswell.com/science-research (accessed on 17 July 2017).
- The helminthic therapy. Available online: https://tanawisa.com/ (accessed on 17 July 2017).
- Turner, K.J.; Quinn, E.H.; Anderson, H.R. Regulation of asthma by intestinal parasites. Investigation of possible mechanisms. Immunology 1978, 35, 281–288. [Google Scholar] [PubMed]
- Mpairwe, H.; Ndibazza, J.; Webb, E.L.; Nampijja, M.; Muhangi, L.; Apule, B.; Lule, S.; Akurut, H.; Kizito, D.; Kakande, M.; et al. Maternal hookworm modifies risk factors for childhood eczema: Results from a birth cohort in Uganda. Pediatr. Allergy Immunol. 2014, 25, 481–488. [Google Scholar] [CrossRef] [PubMed]
- McSorley, H.J.; Gaze, S.; Daveson, J.; Jones, D.; Anderson, R.P.; Clouston, A.; Ruyssers, N.E.; Speare, R.; McCarthy, J.S.; Engwerda, C.R.; et al. Suppression of inflammatory immune responses in celiac disease by experimental hookworm infection. PLoS ONE 2011, 6, e24092. [Google Scholar] [CrossRef] [PubMed]
- Daveson, A.J.; Jones, D.M.; Gaze, S.; McSorley, H.; Clouston, A.; Pascoe, A.; Cooke, S.; Speare, R.; Macdonald, G.A.; Anderson, R.; et al. Effect of hookworm infection on wheat challenge in celiac disease—A randomised double-blinded placebo controlled trial. PLoS ONE 2011, 6, e17366. [Google Scholar] [CrossRef] [PubMed]
- Giacomin, P.; Zakrzewski, M.; Jenkins, T.P.; Su, X.; Al-Hallaf, R.; Croese, J.; de Vries, S.; Grant, A.; Mitreva, M.; Loukas, A.; et al. Changes in duodenal tissue-associated microbiota following hookworm infection and consecutive gluten challenges in humans with coeliac disease. Sci. Rep. 2016, 6, 36797. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.; Walker, A.W.; Reyes, J.; Chico, M.; Salter, S.J.; Vaca, M.; Parkhill, J. Patent human infections with the whipworm, Trichuris trichiura, are not associated with alterations in the faecal microbiota. PLoS ONE 2013, 8, e76573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.R.; Dige, A.; Rasmussen, T.K.; Hvas, C.L.; Dahlerup, J.F.; Iversen, L.; Stensvold, C.R.; Agnholt, J.; Nejsum, P. Immune responses and parasitological observations induced during probiotic treatment with medicinal Trichuris suis ova in a healthy volunteer. Immunol. Lett. 2017, 188, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Ruyssers, N.E.; De Winter, B.Y.; De Man, J.G.; Loukas, A.; Herman, A.G.; Pelckmans, P.A.; Moreels, T.G. Worms and the treatment of inflammatory bowel disease: Are molecules the answer? Clin. Dev. Immunol. 2008, 2008, 567314. [Google Scholar] [CrossRef] [PubMed]
- Giacomin, P.; Croese, J.; Krause, L.; Loukas, A.; Cantacessi, C. Suppression of inflammation by helminths: A role for the gut microbiota? Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [PubMed]
- Hays, R.; Esterman, A.; McDermott, R. Control of chronic Strongyloides stercoralis infection in an endemic community may be possible by pharmacological means alone: Results of a three-year cohort study. PLoS Negl. Trop. Dis. 2017, 11, e0005825. [Google Scholar] [CrossRef] [PubMed]
- Flannery, G.; White, N. Immunological Parameters in Northeast Arnhem Land Aboriginies: Consequences of Changing Settlement and Lifestyles; Cambridge University Press: New York, NY, USA, 1993; pp. 202–220. [Google Scholar]
Figure 1.
Lifecycles of soil-transmitted helminths (STH), S. sterocoralis, and Toxocara. 1. Adult worms reside in the gastrointestinal tract (GIT). Hookworm, A. lumbricoides, and S. stercoralis adults reside in the small intestine while T. trichiura adults reside in the cecum and ascending colon. Female worms produce eggs which are passed in the stool of an infected person. 2. T. trichiura, Toxocara, and A. lumbricoides eggs mature in soil but do not hatch. Hookworm eggs hatch in soil and mature into L3 hookworm larvae. S. stercoralis eggs hatch into rhabditiform larvae in the gut, which are then excreted via the faeces. Rhabditiform larvae then mature into infective filariform larvae or free-living adults. 3. Infectious L3 filariform larvae of hookworm and S. stercoralis penetrate the skin directly, enter the circulation and migrate to the GIT after passing into the lumen of the lungs. 4. Mature eggs of Toxocara, A. lumbricoides, and T. trichiura are swallowed by the host. The eggs hatch, releasing larvae in the GIT. T. trichiura larvae hatch in the small intestine and mature into adults in the colon while Toxocara and A. lumbricoides larvae penetrate the gut. Toxocara larvae are carried by the circulation to a variety of tissue types while A. lumbricoides larvae are carried to the lungs. 5. Hookworm and A. lumbricoides larvae penetrate the alveolar walls and ascend the bronchial tree to the throat and are swallowed. Once they reach the small intestine the larvae mature into adults. S. stercoralis can also follow bronchial migration, or they can penetrate straight to the GIT. 6. Toxocara larvae can be carried to any tissue type. As humans are dead-end hosts the larvae do not undergo further development once they reach these sites, they can cause local reactions, known as the disease toxocariasis. Ocular toxocariasis, where the larvae penetrate the eye, can result in blindness. 7. S. stercoralis can also undergo autoinfection, where the rhabditiform larvae become infective filarial form larvae in the small intestine and penetrate the gut or perianal region. The filariform larvae can then disseminate to throughout the body. 8. Strongyloides rhabditiform larvae develop into free-living adults that produce eggs from which rhabditidorm larvae hatch. Rhabditiform larvae then develop into infectious filariform larvae and penetrate a human host. The free-living cycle exists for one generation cycle only.
Figure 1.
Lifecycles of soil-transmitted helminths (STH), S. sterocoralis, and Toxocara. 1. Adult worms reside in the gastrointestinal tract (GIT). Hookworm, A. lumbricoides, and S. stercoralis adults reside in the small intestine while T. trichiura adults reside in the cecum and ascending colon. Female worms produce eggs which are passed in the stool of an infected person. 2. T. trichiura, Toxocara, and A. lumbricoides eggs mature in soil but do not hatch. Hookworm eggs hatch in soil and mature into L3 hookworm larvae. S. stercoralis eggs hatch into rhabditiform larvae in the gut, which are then excreted via the faeces. Rhabditiform larvae then mature into infective filariform larvae or free-living adults. 3. Infectious L3 filariform larvae of hookworm and S. stercoralis penetrate the skin directly, enter the circulation and migrate to the GIT after passing into the lumen of the lungs. 4. Mature eggs of Toxocara, A. lumbricoides, and T. trichiura are swallowed by the host. The eggs hatch, releasing larvae in the GIT. T. trichiura larvae hatch in the small intestine and mature into adults in the colon while Toxocara and A. lumbricoides larvae penetrate the gut. Toxocara larvae are carried by the circulation to a variety of tissue types while A. lumbricoides larvae are carried to the lungs. 5. Hookworm and A. lumbricoides larvae penetrate the alveolar walls and ascend the bronchial tree to the throat and are swallowed. Once they reach the small intestine the larvae mature into adults. S. stercoralis can also follow bronchial migration, or they can penetrate straight to the GIT. 6. Toxocara larvae can be carried to any tissue type. As humans are dead-end hosts the larvae do not undergo further development once they reach these sites, they can cause local reactions, known as the disease toxocariasis. Ocular toxocariasis, where the larvae penetrate the eye, can result in blindness. 7. S. stercoralis can also undergo autoinfection, where the rhabditiform larvae become infective filarial form larvae in the small intestine and penetrate the gut or perianal region. The filariform larvae can then disseminate to throughout the body. 8. Strongyloides rhabditiform larvae develop into free-living adults that produce eggs from which rhabditidorm larvae hatch. Rhabditiform larvae then develop into infectious filariform larvae and penetrate a human host. The free-living cycle exists for one generation cycle only.
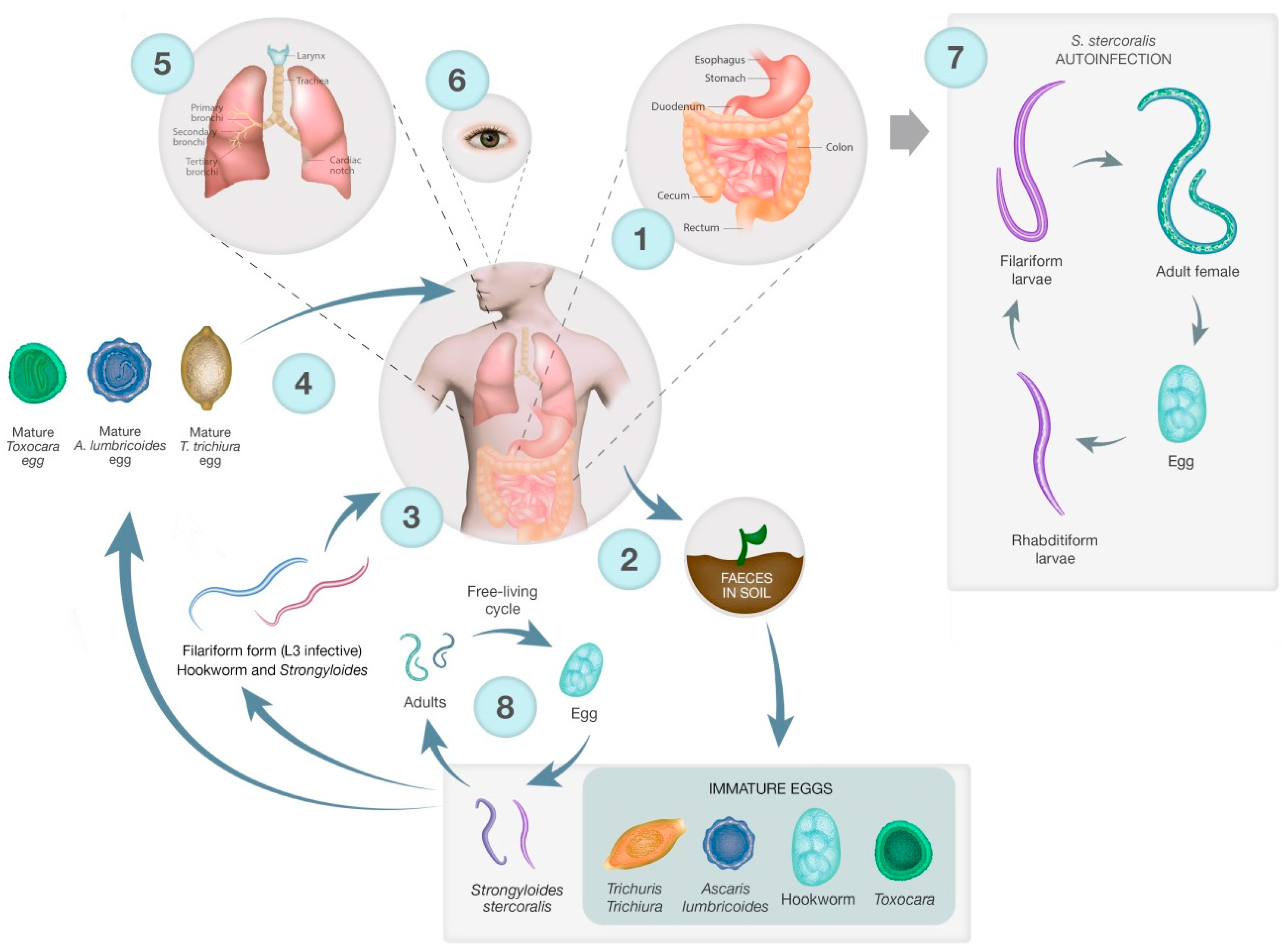
Figure 2.
Distribution of STH in South-East Asia and Australia, modified from Brooker et al., [64].
Figure 2.
Distribution of STH in South-East Asia and Australia, modified from Brooker et al., [64].
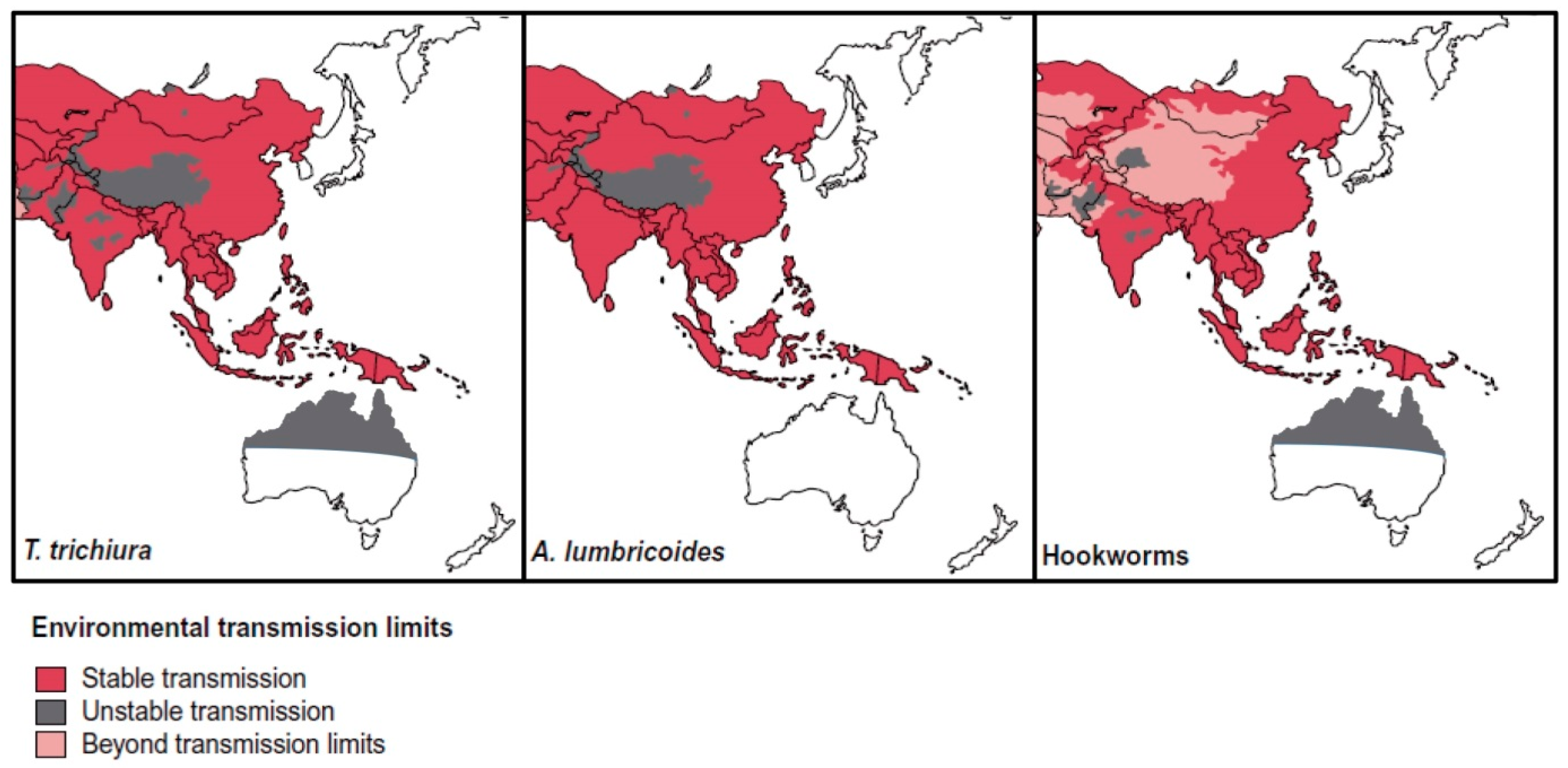
Figure 3.
Prevalence of S. stercoralis in Asia and Australia based on community-based studies, hospital-based studies, and prevalence in immigrants and refugees. Modified from Schäret al., [84].
Figure 3.
Prevalence of S. stercoralis in Asia and Australia based on community-based studies, hospital-based studies, and prevalence in immigrants and refugees. Modified from Schäret al., [84].
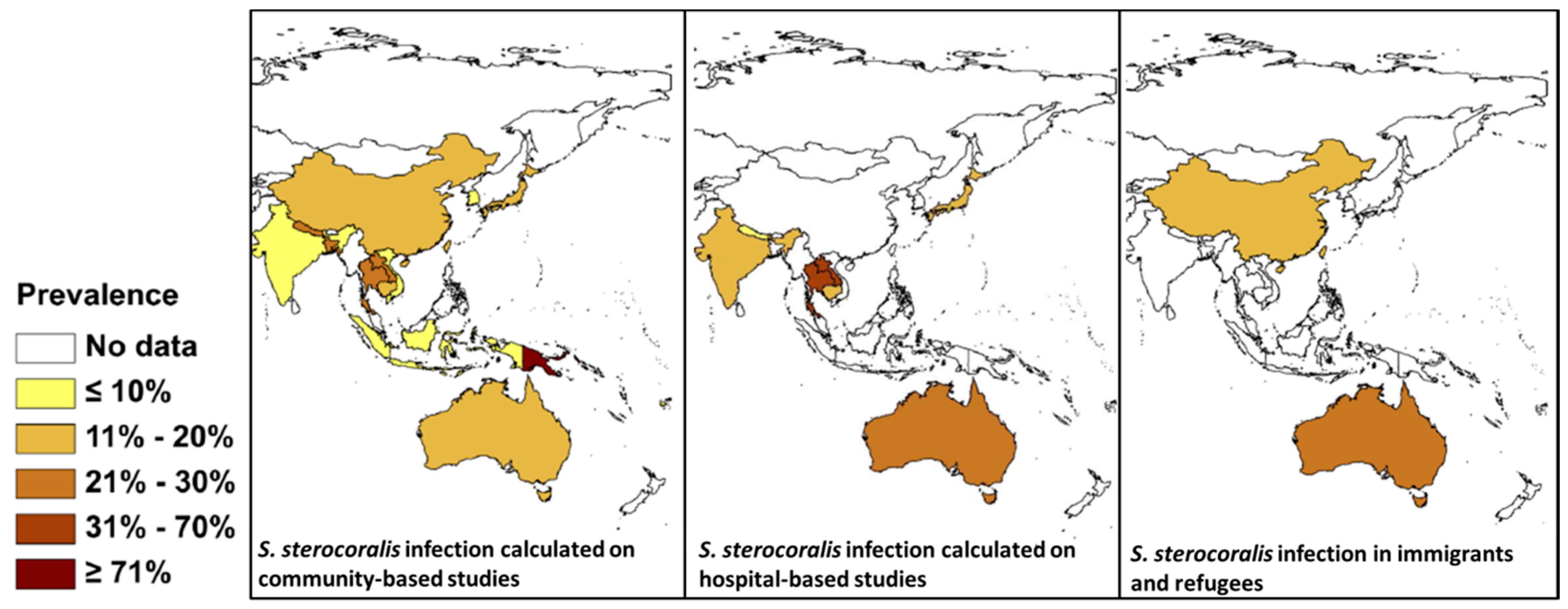

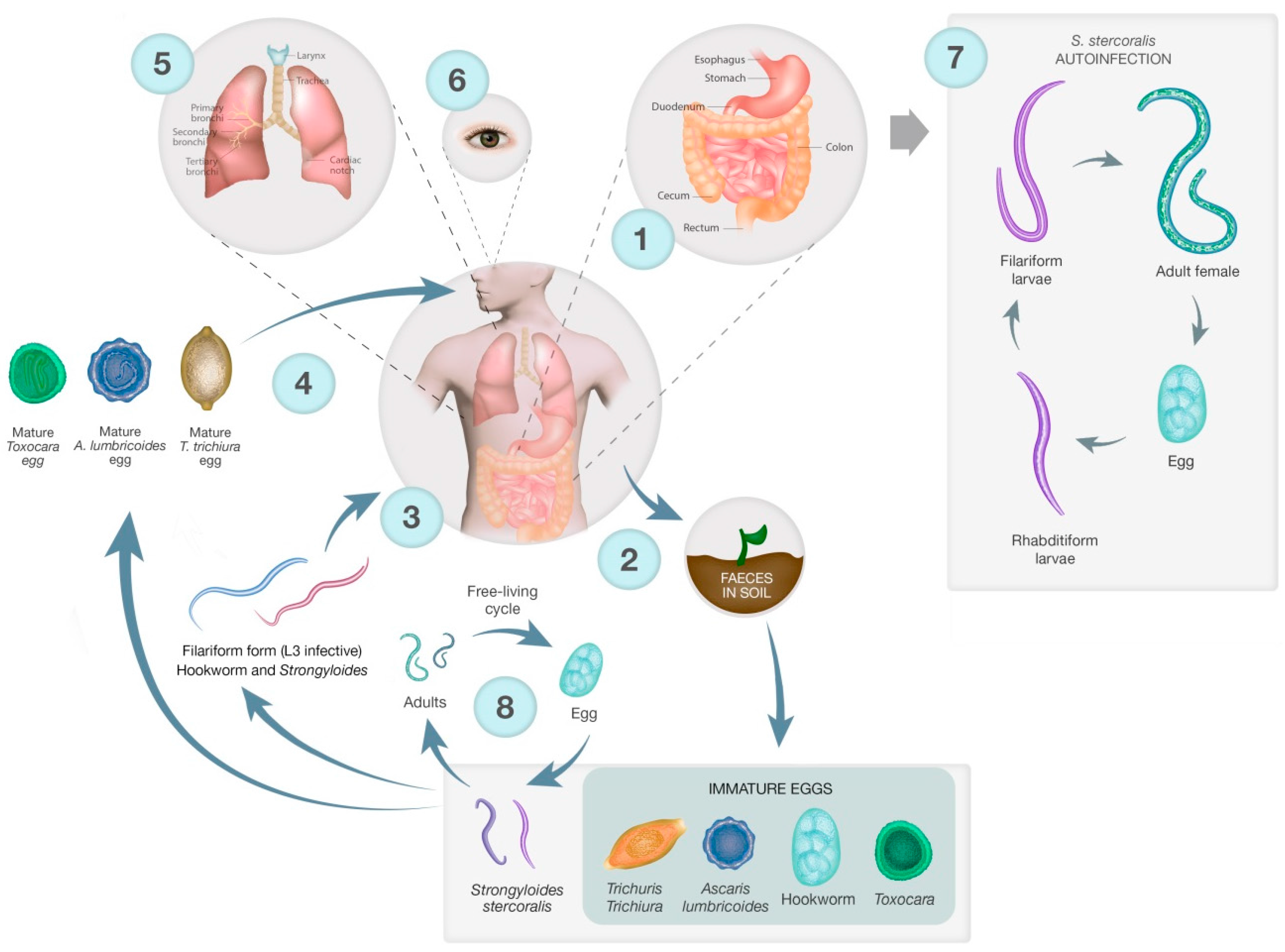{kind=link}
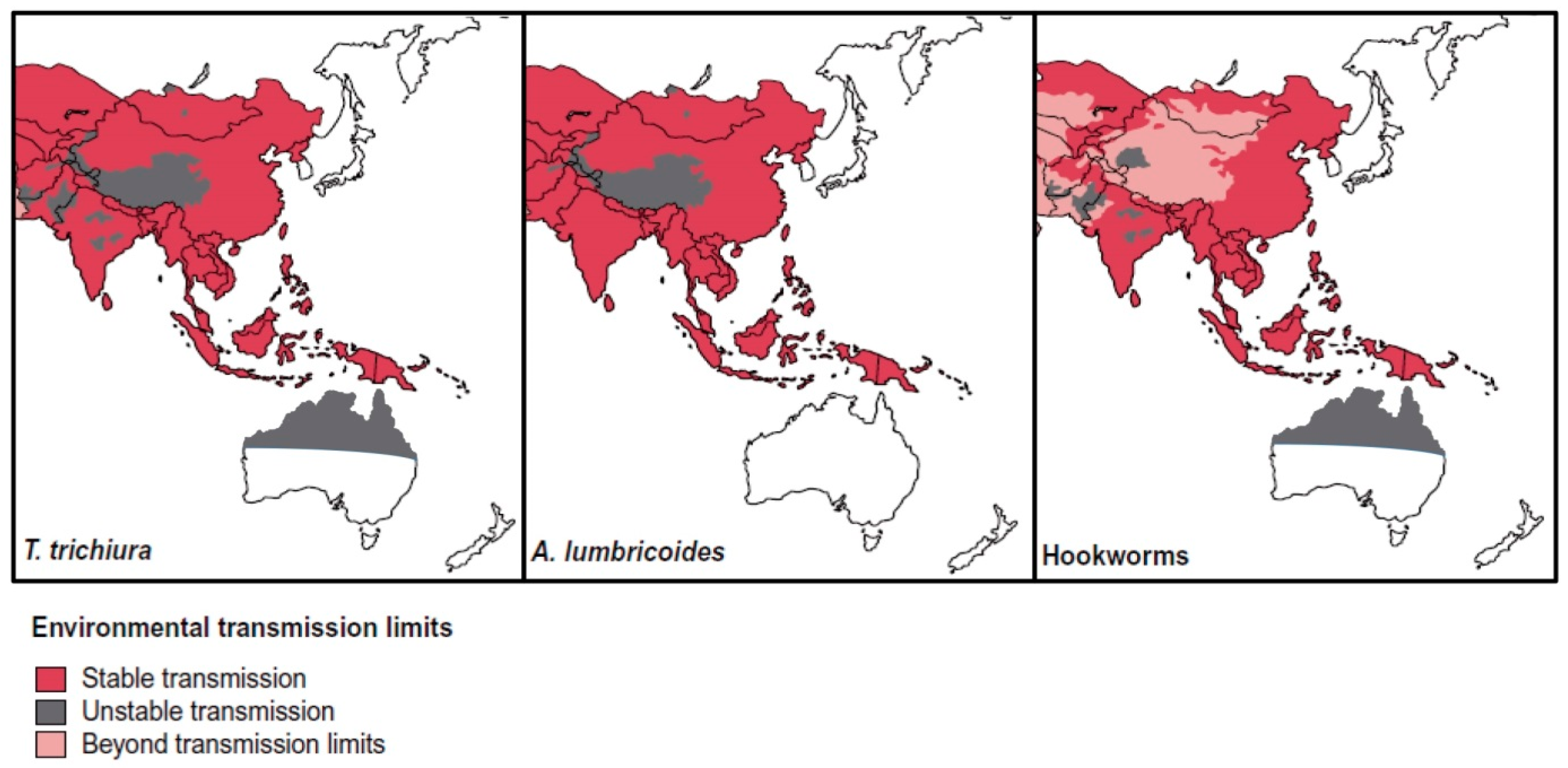{kind=link}
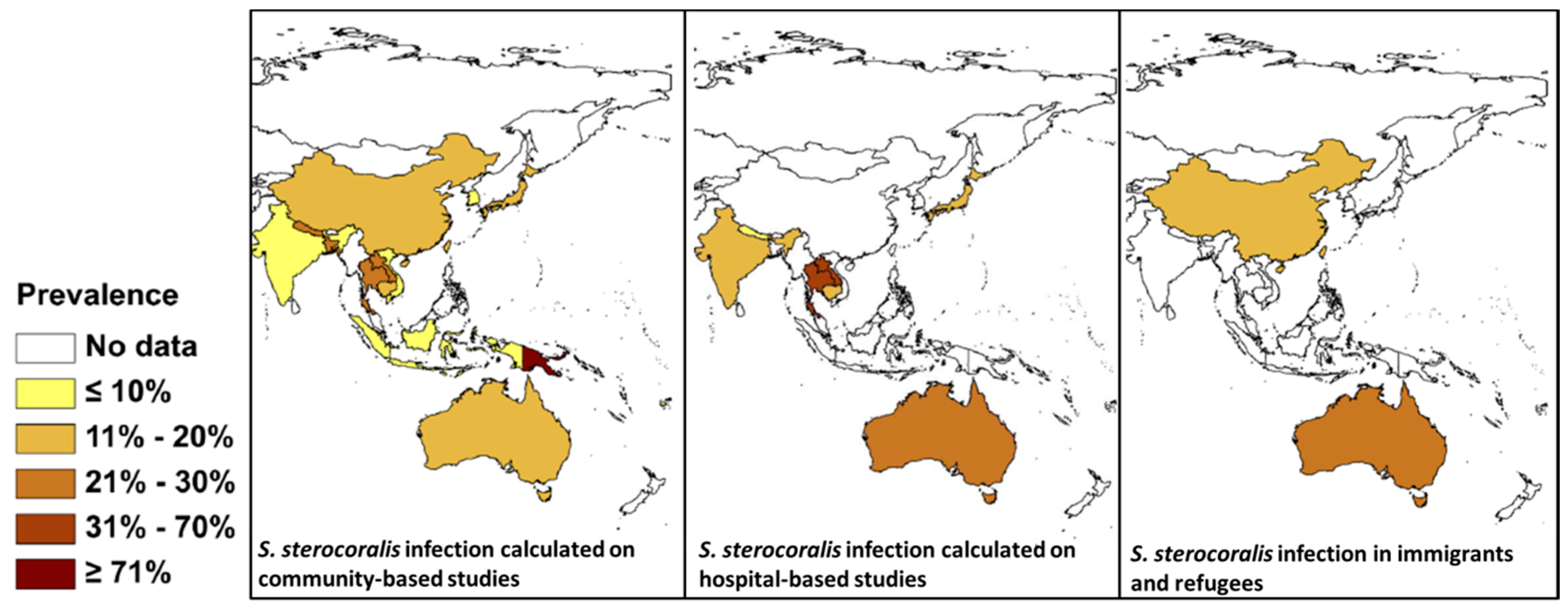{kind=link}
Table 1.
Prevalence of soil-transmitted helminths diagnosed in immigrants, refugees, ADF# personnel, and returned travellers in Australia since 2000.
Table 1.
Prevalence of soil-transmitted helminths diagnosed in immigrants, refugees, ADF# personnel, and returned travellers in Australia since 2000.
| Years Sampled | Reference | Status | Country of Origin | Parasite Species | Prevalence | Diagnostics |
|---|---|---|---|---|---|---|
| 2000, 2002 | [36] | Immigrant | East Africa Cambodia | S. stercoralis T. trichiura S. stercoralis Hookworm spp. | 11% (n = 124) 4% (n = 124) 42% (n = 230) 1.96% (n = 230) | Faecal samples (method unclear) Serology (method unclear) |
| 7–20 years after resettlement | [37] | Immigrant | Laos | S. stercoralis | 24.21% (n = 95) | Faecal microscopy Strongyloides serology |
| 2–52 years after resettlement 1998–2005 | [38] * | Immigrant | Fiji (1), SEA (5), China (1), Sri Lanka (1), India (2), Seychelles (2), Ethiopia (2), Russia (1), Italy (1), Greece (1) | S. stercoralis | 100% (n = 17) * | Faecal microscopy Strongyloides serology |
| 1998–2005 | [38]* | Returned travellers | Papua New Guinea (1), Vanuatu (1), SEA (7), Africa (2) | S. stercoralis | 100% (n = 11) * | Faecal microscopy Strongyloides serology |
| 2004 | [39] | Returned ADF # member | Solomon Islands | A. ceylanicum | 100% (n = 1) | Harada-Mori culture, direct faecal smear |
| Served 1962–1975 2010 | [40] | ADF veterans | Vietnam | S. stercoralis | 11.6% (n = 249) | Faecal microscopy ELISA |
| 2006–2007 | [41] | RAMSI personnel *** | Solomon Islands | S. stercoralis | 100% (n = 14) * | Faecal microscopy, Serology (ELISA) |
| 2002–2012 | [42] | Residents Northern Territory | Australia | T. trichiura | 0.65% (n = 63,668) ** | Wet mount microscopy, Concentration method |
| 2002–2011 | [43] | Residents Northern Territory | Australia | Hookworm | 0.17% (n = 64,691) ** | Wet mount microscopy, Concentration method |
| 2004–2008 | [44] | Immigrants | Burma | S. stercoralis | 26% (n = 156) | Serology |
| 2002 | [45] | Immigrants | Cambodia | S. stercoralis | 36% (n = 234) * | ELISA, faecal microscopy |
| 2010–2011 | [46] | Residents Northern Territory | Australia | S. stercoralis | 16.5% (n = 124) pre-treatment 12% (n = 30) post-treatment | Serology (NIE ELISA, NIE-DBS-ELISA) |
| 2000–2006 | [29] | Residents | Australia | S. stercoralis | 100% (n = 18) * | Faecal microscopy, serology |
| 1994–1996 | [47] | Residents | Australia | T. canis S. stercoralis S. stercoralis | 21% (n = 29) 28% (n = 29) 19% (n = 314) | Serology Serology Formol-ether |
| [48] | Immigrant | Laos | S. stercoralis | Single patient | Larvae in sputum | |
| 2010–2011 | [49] | Immigrants Residents Residents | Australia | N. americanus A. ceylanicum A. duodenale | (n = 5/227) **** (n = 2/227) (n = 4/227) | PCR Sequencing |
# Australian Defence Force (ADF); * retrospective review of positive cases; ** faecal samples; *** Regional Assistance Mission to Solomon Islands; **** faecal samples from individuals with a documented history of gastrointestinal disorders.
Table 2.
Reported prevalences of A. ceylanicum human and animal infections in studies conducted in Australia and Asia since 2000.
Table 2.
Reported prevalences of A. ceylanicum human and animal infections in studies conducted in Australia and Asia since 2000.
| Ref | Year | Country * | Human/Animal | Prevalence % (Total no.) | Species | Diagnostic |
|---|---|---|---|---|---|---|
| [95] | - | Taiwan | Human | Single patient | A. ceylanicum | Morphology |
| [69] | 2009 | Laos | Human | 17.6% (n = 17) 82.4% (n = 17) | A. ceylanicum N. americanus | Nested PCR |
| [96] | 2012 | Cambodia | Human | 51.6% (n = 124) 51.6% (n = 124) 3.2% (n = 124) | A. ceylanicum N. americanus A. duodenale | Microscopy, PCR |
| Dog | 94.4% (n = 90) 8.9% (n = 90) 1.1% (n = 90) | A. ceylanicum A. caninum N. americanus | Microscopy, PCR | |||
| [49] # | 2010–2011 | Australia | Human | 0.88% (n = 227) 1.76% (n = 227) 1.76% (n = 227) | A. ceylanicum A. duodenale N. americanus a | PCR |
| [97] | - | China | Dog Cat Human | 3 (n = 254) 5 (n = 102) 14 (n = 14) | A. ceylanicum | PCR sequencing |
| [98] | - | Malaysia | Dog | 52% (n = 224) 48% (n = 224) | A. ceylanicum A. caninum | FECT, PCR |
| [99] | 2007–2010 | Malaysia | Cat | 29.5% (n = 543) | A. ceylanicum | Microscopy |
| [100] | 2009–2011 | Malaysia | Human | 87.2 (n = 47) 23.4 (n = 47) | N. americanus A. ceylanicum | Microscopy, PCR |
| [101] | 2013 | Malaysia (Chinese) | Human | Single patient | A. ceylanicum | Microscopy |
| [11] | 2009–2011 | Malaysia | Human | 12.8% (n = 634) 76.6% (n = 634) 10.6% (n = 634) | A. ceylanicum N. americanus Both species | Microscopy, PCR |
| Cats and dogs | 52% (n = 105) 46% (n = 105) | A. caninum A. ceylanicum | Microscopy, PCR | |||
| [102] | - | Myanmar | Human | 72.72% (n = 11) 27.27% (n = 11) | N. americanus A. ceylanicum | PCR sequencing |
| [103] | 2004–2005 | Australia | Dog | 6.5% (n = 92) 70.7% (n = 92) 4.3% (n = 92) 2.2% (n = 92) | A. ceylanicum A. caninum A. caninum + A. ceylanicum A. caninum + U. stenocephala | Microscopy, PCR-RFLP |
| Cat | 30% (n = 10) | A. caninum | ||||
| [12] | 2011–2013 | Thailand | Human | 60% (n = 10) 30% (n = 10) 10% (n = 10) | N. americanus A. ceylanicum A. duodenale | PCR sequencing |
| [86] | >2007 | Australia | Wild dog | 100% (n = 26) 11.5% (n = 26) | A. caninum A. ceylanicum + A. caninum | Microscopy, PCR |
| Dog scat | 65.31% (n = 89) 71.43% (n = 89) 38.78% (n = 89) | A. ceylanicum A. caninum A. caninum + A. ceylanicum | ||||
| [39] | 2004 | Australia (Solomon Islands) | Human | Single patient | A. ceylanicum | Microscopy |
| [51] | 2004–2005 | Thailand | Dog | 77% (n = 229) 9% (n = 229) 14% (n = 229) | A. ceylanicum A. caninum Both species | PCR |
| Human | 71.43% (n = 204) 28.57% (n = 204) | N. americanus A. ceylanicum | ||||
| [104] | 2008 | India | Dog | 50.46% (n = 325) 51.92% (n = 104) 33.65% (n = 104) 15.38% (n = 104) | Hookworm spp. A. caninum A. ceylanicum A. caninum + A. ceylanicum | Microscopy, PCR-RFLP |
| [105] | 2000 | India | Dog | 36% (n = 101) 38% (n = 101) | A. caninum A. caninum + A. braziliense | Microscopy, PCR-RFLP + sequencing |
| [85] | 2011 | Australia | Dog | 96.4% (n = 84) 16.67%(n = 84) 14.0% (n = 84) | A. caninum A. ceylanicum A. caninum + A. ceylanicum | Microscopy, PCR |
| [106] | 2013–2014 | Malaysia | Dog | 29.6% (n = 227) 6.6% (n = 227) | A. ceylanicum A. caninum | FECT, PCR |
| Soil samples | 14.3% (n = 126) 2.4% (n = 126) | A. ceylanicum A. caninum | ||||
| Cat | 29.6% (n = 152) 6.6% (n = 152) | A. ceylanicum A. caninum | ||||
| [107] | 2015 | Japan (Lao) | Human | Single patient | A. ceylanicum | Microscopy, PCR |
| [108] | 2013–2015 | India | Human | 100% (n = 143) 16.8% (n = 143) 8.4% (n = 143) | N. americanus A. caninum A. duodenale | |
| Dog | 27.9% 76.4% | A. ceylanicum A. caninum | PCR-RFLP | |||
| Soil samples | 60.2% (n = 78) 29.4% (n = 78) 16.6% (n = 78) 1.4% (n = 78) | A. ceylanicum A. caninum A. duodenale N. americanus | ||||
| [109] | 2014 | Thailand | Dog Cat | 33.0% (n = 197) 58.46% (n = 180) | A. ceylanicum | Microscopy, PCR |
| [110] | 2014 | France (Myanmar) | Human | Single patient | A. ceylanicum | Microscopy, PCR |
| [111] | 2014 | Vietnam | Dog | 54.3% (n = 94) 33% (n = 94) 12.7% (n = 94) | A. ceylanicum A. caninum Both species | PCR-RFLP, PCR (cox1) |
| [112] | 2014 | China | Cat | 40.8% (n = 112) 59.2% (n = 112) 20.4% (n = 112) | A. ceylanicum A. caninum Both species | Microscopy, PCR |
| [113] | - | India | Human | 95% 15% 5% | N. americanus A. duodenale A. ceylanicum | PCR-RFLP |
| [114] | 2005 | Thailand | Human | 92% 4% 2% 2% | N. americanus A. ceylanicum A. duodenale N. americanus + A. ceylanicum | KK, PCR |
| [115] | 2008 | Lao | Human | 5.91% 2.46% 1.48% 0.49% | N. americanus A. duodenale A. caninum A. ceylanicum | KK, PCR |
| [99] | 2007–2010 | Malaysia | Feral cats | 29.5% (n = 251) | A. ceylanicum | Microscopy of adults (staining paracarmine) |
| [116] | - | Lao | Feral cats | 69% (n = 55) | A. ceylanicum | Microscopy of adults (staining Mayers carmine) |
| [117] | - | Taiwan | Human | Single patient | A. ceylanicum | Method unclear. Adult identification. |
* Origin of infection in brackets if not the same as the country of detection; # faecal samples from individuals with a documented history of gastrointestinal disorders; a individuals infected with N. americanus were refugees from Sierra Leone and Sudan, likely to be acquired in those countries.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gordon, C.A.; Kurscheid, J.; Jones, M.K.; Gray, D.J.; McManus, D.P. Soil-Transmitted Helminths in Tropical Australia and Asia. Trop. Med. Infect. Dis. 2017, 2, 56. https://doi.org/10.3390/tropicalmed2040056
AMA Style
Gordon CA, Kurscheid J, Jones MK, Gray DJ, McManus DP. Soil-Transmitted Helminths in Tropical Australia and Asia. Tropical Medicine and Infectious Disease. 2017; 2(4):56. https://doi.org/10.3390/tropicalmed2040056
Chicago/Turabian StyleGordon, Catherine A., Johanna Kurscheid, Malcolm K. Jones, Darren J. Gray, and Donald P. McManus. 2017. "Soil-Transmitted Helminths in Tropical Australia and Asia" Tropical Medicine and Infectious Disease 2, no. 4: 56. https://doi.org/10.3390/tropicalmed2040056