Treading the Path towards Genetic Control of Snail Resistance to Schistosome Infection


Department of Medical Microbiology and Parasitology, College of Medicine, University of Lagos, Idi-Araba, Surulere, Lagos 100254, Nigeria
Trop. Med. Infect. Dis. 2018, 3(3), 86; https://doi.org/10.3390/tropicalmed3030086
Submission received: 18 July 2018
/
Revised: 8 August 2018
/
Accepted: 13 August 2018
/
Published: 15 August 2018
(This article belongs to the Special Issue Prospects for Schistosomiasis Elimination)
Abstract
:Schistosomiasis remains the most important tropical snail-borne trematodiasis that threatens many millions of human lives. In achieving schistosomiasis elimination targets, sustainable control of the snail vectors represents a logical approach. Nonetheless, the ineffectiveness of the present snail control interventions emphasizes the need to develop new complementary strategies to ensure more effective control outcomes. Accordingly, the use of genetic techniques aimed at driving resistance traits into natural vector populations has been put forward as a promising tool for integrated snail control. Leveraging the Biomphalaria-Schistosoma model system, studies unraveling the complexities of the vector biology and those exploring the molecular basis of snail resistance to schistosome infection have been expanding in various breadths, generating many significant discoveries, and raising the hope for future breakthroughs. This review provides a compendium of relevant findings, and without neglecting the current existing gaps and potential future challenges, discusses how a transgenic snail approach may be adapted and harnessed to control human schistosomiasis.
1. Introduction
It is presently more than a century since malacological discoveries established that various genera of freshwater snails (Mollusca: Gastropoda) serve as biological vectors of human diseases caused by parasitic trematodes. These findings have made the study of snail biology an important aspect of infectious disease research, particularly in tropical helminthology. Among these human snail-borne trematodiases, schistosomiasis (bilharziasis) ranks as the most important disease, afflicting more than 206 million humans [1], and being responsible for over 3.51 million disability-adjusted life years (DALYs) [2], most prominently in the tropics. Most cases of human schistosomiasis are caused by three parasitic schistosomes (blood flukes): Schistosoma haematobium, S. mansoni, and S. japonicum [3]. Both S. haematobium and S. mansoni are found in Africa and the Middle East; only S. mansoni occurs in the Americas, and S. japonicum is a major disease-causing species in China, Indonesia, and the Philippines [3]. However, the geographical distribution of these flukes is synchronous to, and importantly determined by, the local distribution of their snail vectors. S. haematobium is transmitted by Bulinus snails, S. mansoni by Biomphalaria snails, and S. japonicum by Oncomelania snails [4].
The human-to-snail-to-human transmission of Schistsosoma occurs when adult male and female living in copula within the human host mate and produce fertilized eggs. Some of the eggs are voided with urine (in S. haematobium) or feces (in S. mansoni and S. japonicum) into the environment. The eggs that reach the vectors’ freshwater habitats hatch and release the enclosed miracidia larvae, which swim actively to locate and infect their snail vectors. A miracidium that successfully infects a susceptible vector undergoes intramolluscan polyembryonic development to produce thousands of actively-swimming tailed cercariae larvae that emerge continuously from the snail host for the rest of the its lifetime (spanning months) [3,4,5,6]. Human infection with schistosomes is acquired through skin contact with, and subsequent penetration by, the cercariae during recreational, domestic, or occupational activities with contaminated water [5]. Following penetration, the worms transform into immature schistosomes (schistosomulae) and are carried in body circulation, from where they enter the portal veins and mature in about 5–7 weeks [3,5]. Mature worm pairs migrate to their preferred host sites—S. mansoni and S. japonicum to the mesenteric venules of the bowel or rectum, and S. haematobium to the venous plexus of the bladder, where they mate and the females lay eggs to repeat the cycle [4,5]. Adult schistosomes have an average lifespan of 3–10 years, but they may also live as long as 30–40 years in their human hosts [3,4,5]. The eggs are highly immunogenic and are majorly responsible for disease outcomes by triggering localized pathologic reactions within the human host [4,7,8]. Although human infection with Schistosoma species may cause non-specific but incapacitating systemic morbidities such as malnutrition, anemia, and impaired physical and cognitive development in children, poor birth outcomes in infected pregnant women, and neurological aberrations, S. haematobium is specifically responsible for urogenital pathologies, while other Schistosoma species majorly cause gastrointestinal complications, but also hepatosplenic enlargement, ascites, and portal hypertension in advanced cases [3,7,9,10]. Again, there is growing evidence that female urogenital schistosomiasis poses an increased risk of HIV transmission and/or progression [11,12,13].
Taking a leap towards the beginning of the end human schistosomiasis requires an integrated control approach that cuts across both the vector and the human cycles. Current strategy in the fight against the disease co-implements ongoing preventive chemotherapy through mass drug administration (MDA), with complementary public-health interventions. This approach, as defined by WHO/AFRO, is known as PHASE—preventive chemotherapy, health education, access to clean water, sanitation improvement, and environmental snail control and focal mollusciciding [14]. Recent efforts made to evaluate the degree of importance of snail control in schistosomiasis elimination [15,16,17,18] clearly showed that sustainable snail control is pivotal in achieving targeted disease elimination. This is especially true in the present era of highly challenging anti-schistosome vaccine development, as well as the monochemotherapeutic availability of praziquantel and its feared resistance by schistosomes [19,20,21]. Strategies currently in use for controlling schistosomiasis snail vectors are: biocontrol using competitors or predators, modification of snail habitats, and application of molluscicides. These approaches, used either singly or in combination, have evidently contributed to many successful schistosomiasis control efforts in different localities and countries [15,22,23,24,25,26,27]; however, each approach is not without limitations [24]. The application of chemical molluscicides has been mostly exploited. Among other chemical molluscicidal agents, niclosamide has a long track record of being successful against snail hosts, and is often regarded as the molluscicide of choice. Nevertheless, apart from its expensiveness, toxicity of niclosamide to a variety of non-target aquatic life forms (plants, invertebrates and vertebrates including amphibians) has led to its decreased acceptability. Again, the inability of niclosamide to prevent snail recolonization, especially in large permanent water bodies, necessitates repeated applications that result in high cost [24,28,29,30].
In view of the present challenges facing schistosomiasis control efforts, coupled with the endorsement by the World Health Assembly Resolution 65.21 to take full advantage of non-drug-based interventions to prevent schistosomiasis transmission [31], it will be timely to adapt new strategies in order to interrupt snail-mediated schistosome transmission, and thus, forestall human infection. The use of genetic techniques to manipulate snail vectors of schistosomiasis has long been stressed as a novel biocontrol strategy with the potential to constitute an important complementary tool for transmission reduction or breaking. Embracing all the means to actualize this potential, studies unraveling the complexities of the vector biology and those exploring the molecular underpinnings of snail resistance/susceptibility to schistosome infection have been expanding in various breadths, generating many significant discoveries and raising the hope for future breakthroughs. The aim of this review is to provide a compendium of relevant findings, and discuss how transgenic snail approach may be adapted and harnessed to control human schistosomiasis.
2. Biology of Snail Resistance/Susceptibility to Schistosoma Infections–Major Exploits so Far
The first groundbreaking discovery on the identification of intermediate snail hosts of schistosomes was made by Miyairi and Suzuki, who observed stages of S. japonicum in Oncomelania snails in Japan in 1913 [32,33]. This was followed by the achievements of Robert Leiper, who also demonstrated the complete life cycles of S. haematobium and S. mansoni in their respective snail hosts in Egypt [34,35]. Subsequent to these watershed moments in the long history of schistosomiasis, investigations on the interactions between schistosomes and their snail vectors became kinetic. The genetic study of snail-schistosome compatibility was pioneered by Newton [36,37], who demonstrated that susceptibility of snail vectors to Schistosoma infections is fundamentally genetic and a heritable character. This was later underscored by other investigators who revealed that resistance character, which is acquired at the maturity phase in the adults of resistant snail stocks, is monogenic, dominant, and heritable by a simple Mendelian pattern of inheritance [38,39,40,41]. This genetic dominance of the resistance trait has been confirmed by various crossbreeding experiments in Biomphalaria species [42,43,44,45,46]. Be that as it may, Rosa et al. [45] showed that resistance in B. tenagophila is determined by two dominant genes. In contrast, in juvenile B. glabrata, resistance is a complex trait governed by a minimum of four genes, each having multiple alleles (alternative forms of the same gene) [40,47]. From these various lines of evidence, it could be understood that genetic determinism of resistance is governed by a single major locus (position of a particular gene or allele on a chromosome) to a potentially high number of loci, and snails with significantly increased resistance could be artificially selected in the laboratory; meanwhile, molecular markers mapped to resistance could be identified in genetic crosses.
Thus far, work has been done most extensively using the Biomphalaria-Schistosoma model, and has led to the nomenclature of some stocks known for resistance (e.g., pigmented BS-90 [48], black-eye 10-R2 [49], and 13-16-IR [50]) or susceptibility (e.g., the albino M-line and NMRI [51], and BB02 [52]) to S. mansoni infection, which are now maintained in the laboratory for research purposes. In contrast to the 10-R2 and 13-16-IR strains, however, BS-90 demonstrates unflinching resistance stability, irrespective of age (juvenile or adult), under laboratory conditions [40,53].
A major physiological determinant of snail resistance/susceptibility to infections, which is also under genetic influence, is the snail internal defense system (IDS). The IDS comprises the cellular elements (hemocytes) and the humoral (plasma) factors of the hemolymph that work independently or in concert to recognize, encapsulate, kill, and clear intruding trematodes [6,54,55,56]. Establishment of the B. glabrata embryonic (Bge) cell line in 1976 [57] provided an enabling avenue for investigators to delve into the molecular and cellular aspects of the complex snail immune functions against schistosomes by using an in vitro culture model, rather than using the whole intact animal, which could have resulted in a rudimentary understanding of the complex biological events. Moreover, major advances in Biomphalaria omic studies, such as the recent availability of the whole genome sequence of B. glabrata [58], provide a useful resource in deciphering complex functions of the snail biology that were previously obscure. Using various strain and species combinations of the Biomphalaria-Schistosoma model system, robust molecular studies have been carried out, leveraging various techniques to identify and characterize endogenous effector protein/gene candidates that are functional in the snail internal defense machinery against schistosomes. Table 1 below presents a synopsis of various endogenous factors that have been implicated in Biomphalaria resistance to schistosomes.
3. Transgenic Snail Methods for Schistosomiasis Control
The use of genetically engineered vectors to either suppress (reduce) or modify (replace) the natural populations of the biological vectors of some globally important infectious diseases has been a convincing concept that is now on the verge of deployment to control disease transmission. This rapidly emerging genetic control approach is distinguished from other biocontrol strategies (such as the use of natural parasitoids, predators, competitors, and infectious microbial agents), as it is mating-based, highly species-specific, and capable of being transmitted or inherited vertically [83]. In this context, suppression or elimination of natural local vector populations could be achieved by releasing transgenic vectors carrying sterile or detrimental characters into the local populations, while the population modification approach requires the release of transgenic pathogen-resistant vector strains or species. Depending on their characteristics or specific configurations, genetic methods for transgenic vector control may be self-limiting with transient persistence among subsequent vector generations unless replenished by repeated release of engineered vectors, or self-sustaining with indefinite persistence from the initial target population to the subsequent ones [83]. With revolutions in biotechnology, the use of gene drive systems (self-sustaining, selfish genetic elements that are inherited by progenies at frequencies largely exceeding those expected in Mendelian inheritance) has become an attractive method for vector control applications, as gene drivers are invasive wherever present, and so may overcome some evolutionary constraints [83,84].
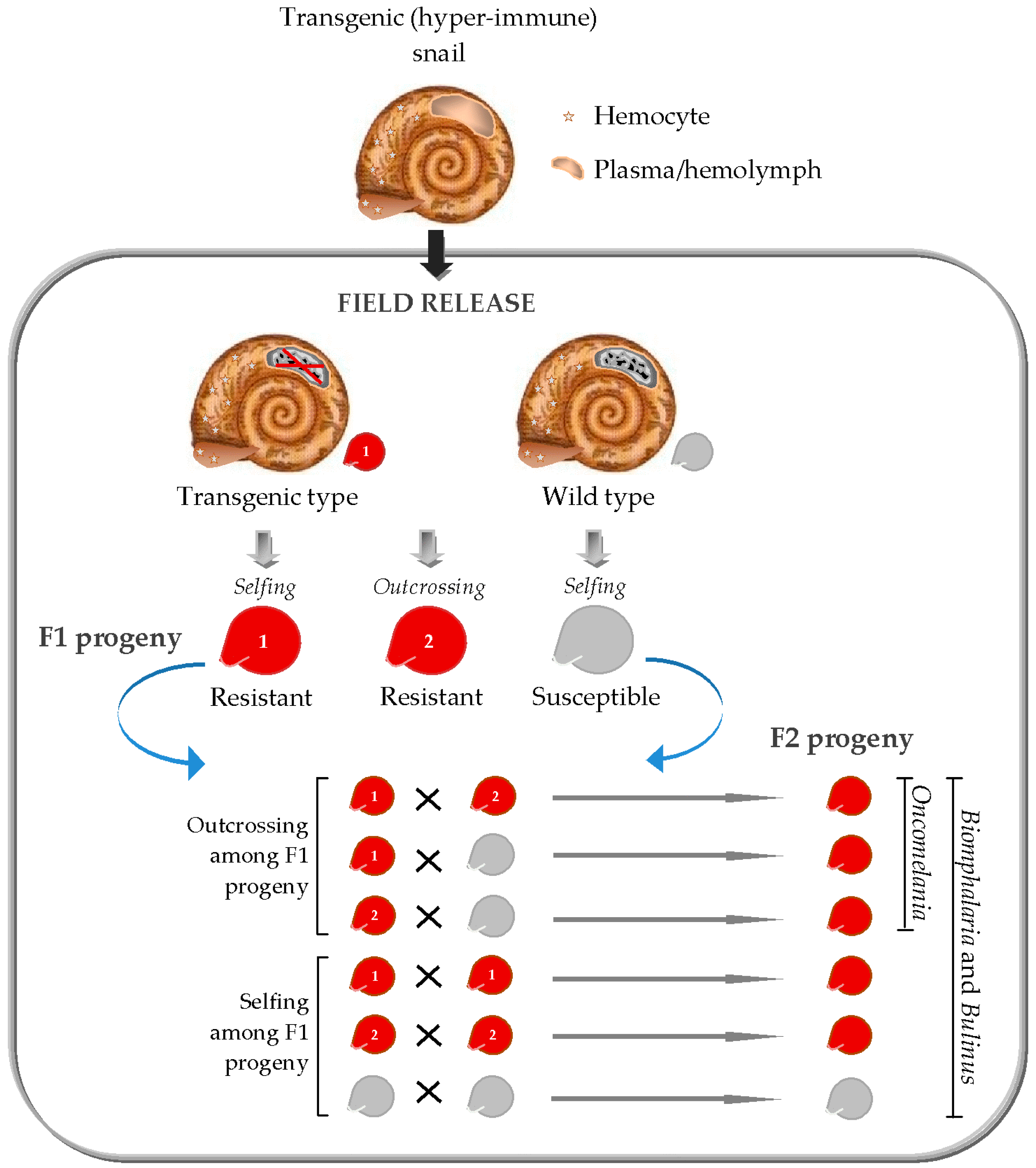
As first proposed in 1958 [85], much emphasis has been placed on the use of genetic biocontrol for schistosomiasis vector control. To select a suitable transgenic vector method for schistosomiasis control however, the basics of the biology of the snail vectors must be taken into account. Although a wealth of genetic studies has been centered on the production of transgenic mosquito vectors of human diseases, biological differences between these dipterans and schistosomiasis molluscan vectors create the need for selective transgenic vector strategies for schistosomiasis control application. For example, unlike in mosquitoes where only females are capable of transmitting pathogens, Biomphalaria and Bulinus are hermaphrodites capable of self- or cross-fertilization [86], and all mating types serve as hosts for schistosomes. Oncomelania has separate sexes [86] but nevertheless, both sexes can also carry S. japonicum, only perhaps at varying degree of susceptibility [87,88]. These biological peculiarities render population reduction strategies unsuitable for genetic control of the snail hosts. This is because the newly released transgenic snails carrying harmful characters would remain susceptible to Schistosoma infections, therefore intensifying parasite transmission. Consequently, present focus in the genetic control of schistosomiasis vectors is set on strategies to modify the natural snail populations. According to Hubendick [85], population of the resistant strains can displace that of the susceptible ones in the field by natural selection. Although this scenario is plausible, it can be realized only through the application of self-sustaining transgenic vector systems (Figure 1).
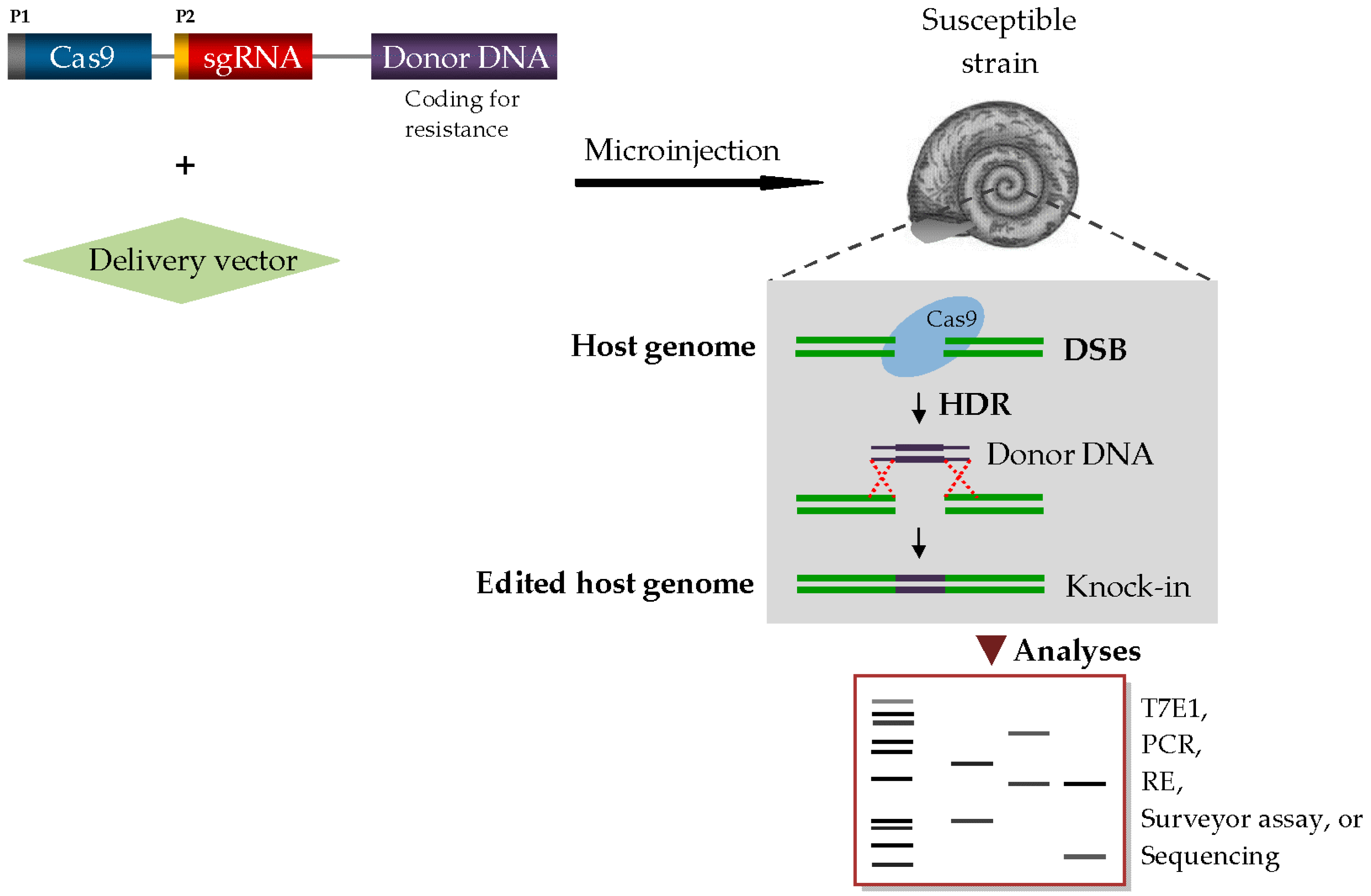
Promoted by its advantages over other gene drive systems—such as transcription activator-like effector nucleases (TALENs) and zinc finger nucleases (ZFNs)—used in genome editing technology, and over other genetic techniques such as RNA interference (RNAi) [84,89,90], the recently-discovered CRISPR/Cas9 (clustered regularly-interspaced short palindromic repeats/CRISPR-associated protein 9) system has flowered, and is being widely used in current research trials and applications to modify genome sequences in diverse species spanning microbes, plants, animals, to even humans [89,90,91,92,93]. In parallel, the use of CRISPR/Cas9 to drive anti-Schistosoma effector genes into the genome of naturally susceptible snail strains is being envisioned, and has become an important subject in current discussions [17,75,94,95]. Fascinatingly, a proof-of-concept study [96] has demonstrated the possibility of CRISPR/Cas9-mediated gene editing in molluscs; indeed, more insights into the possible ways by which this may be achieved for schistosomiasis vector control, as well as the current and potential future challenges, will constitute a key guide for the scientific community in the appropriate fostering of this tantalizing approach in snail molecular research.
The three basic requirements for a CRISPR-based precise gene knock-in editing are Cas9 endonuclease, single-guide RNA (sgRNA), and repair template DNA (donor). The Cas9 enzyme combs through the genome of the host organism, acting as the ‘molecular scissors’ that cuts a specific DNA sequence at a genomic locus. The sgRNA (~20 nucleotides) is designed to match and target the desired DNA sequence to be deleted, while the donor DNA provides a template for genomic repair of the cleaved locus [92,97]. In the case of schistosomiasis snail vectors, Cas9-mediated introgression of refractoriness into susceptible strains will require an engineered donor DNA encoding a locus known to confer resistance. The anti-Schistosoma donor DNA can be tightly anchored to the Cas9/sgRNA complex, and the entire cassette is co-injected with a viral vector, such as lentivirus [98], into the early single-cell stage embryos of the snail vectors (Figure 2). In essence, the viral vector ensures safe and effective delivery of these components into the nuclei of the target cells. Suitable sites for the driver–cargo system injection may include the ovotestis of Biomphalaria/Bulinus snails and the ovary of female Oncomelania snails. In Biomphalaria, the ovotestis is located at the tip of the shell spire [99] and the driver–cargo system insertion into the ovotestis will be more appealing in the albino strain, as the transparency of the snail shell allows easy visibility of internal organs. Further analyses to assess targeting efficiency or screen for transgenic mutants among progenies may be done by T7 endonuclease I (T7E1) assay, restriction enzyme assay, next generation sequencing or direct PCR assay as applicable (Figure 2).
To date, the main genetic loci that have been identified in association with B. glabrata resistance to schistosome are Sod1 and RADres (a restricted-site associated DNA-determined resistance locus) [50,68,69], and a GRC (Guadeloupe Resistance Complex) genomic region (<1 Mb) [69]. In combination with other known and yet unknown resistance genes, Sod1 and RADres occupy haplotype blocks of >2 Mb genomic region [69,100]. Although putative functional gene candidates have been identified in the GRC region [75,95], the Sod1 and RADres regions appear to demonstrate a wider spectrum of snail resistance [69]. Nevertheless, there is still a need to further narrow down these regions to the embedding causative genes, and to understand their immune stability and functions under different genetic backgrounds and environmental conditions.
4. Further Considerations
An early investigator [101] stated that the genetic factors controlling snail insusceptibility to schistosomes must first be clarified, and snail strains ferrying only refractory traits must be developed before we can gainfully engage genetic control methods. The first criterion has largely been met through relentless research unveiling resistance-determining proteins and genes. Despite these advances, current stumbling blocks involve developing snail strains that are reliably recalcitrant to schistosome infection. One major bottleneck is the highly variable strain-by-strain interaction—compatibility polymorphism—that is well-documented to occur in snail-schistosome systems [102,103]. As a consequence, developing a transgenic target for individual strain-to-strain combinations becomes cumbersome, but can be circumvented only if genetic loci with wide-spectrum resistance activities conserved across various strain-to-strain combinations could be identified and characterized. The BS-90 strain of B. glabrata (isolated in Salvadore, Brazil) has been bred in the laboratory for many years and has been shown to be steadily resistant; however, its relative performance in the field remains unpredictable. A tenable reason for this is that generations of the laboratory-bred strains are poor representatives of the genetic variations that actually occur in the original wild populations [103]. Another caveat in the future use of either the resistant BS-90 or transgenic snail strains is global warming, characterized by an increasing earth’s average surface temperature. In sharp contrast to what was earlier known, Knight et al. [78] showed that snail resistance to schistosomes is also temperature-dependent, and even the naturally resistant BS-90 strain could be rendered susceptible at 32 °C. Other local environmental factors such as altitude, water level, soil, and vegetation may also cause differential gene expression and regulation among snails of the same species as a result of local adaptation mechanisms [104].
Organism biodiversity and signatures of interactions between other organisms and the snail vectors living in the same habitat may also impact the outcome of transgenic snail application. In an ecological milieu where natural predators [e.g., Macrobrachium vollenhovenii (a freshwater prawn), Procambarus clarkia (a freshwater crayfish), Marisa cornuarietis (an ampullarid snail), and cichlid fishes such as Trematocranus placodon and Geophagus brasiliensis] or competitors [e.g., thiarid snails such as Melanoides tuberculata and Tarebia granifera] of the snail vectors of schistosomiasis [4,24] exist in meaningful abundance, there is a possibility that the population of the released transgenic snails becomes reduced below levels required to displace that of the naturally susceptible vectors as a result of a more biased killing/eating of the transgenic snails (and eating of their egg masses) or deprivation of resources. When such a scenario operates, the resistance effect tapers off. Given this contingency, the release of transgenic snails may be chosen only in lieu of introducing predators or competitors of snail vectors; co-implementation of both methods in the same freshwater focus may not always complement the transgenic snail approach. In foci where populations of predators or competitors already occur in significant abundance, one-off niclosamide application prior to the release of transgenic snails may offer a more palatable approach in reducing the probability of diluted effect of the transgenic snail release. These phenomena highlight the importance of sampling water habitats for species diversity prior to, and periodically after, releasing transgenic snails.
The merits of using schistosome-resistant transgenic snails beat the limitations of other biological and environmental interventions. For instance, populations of molluscivorous fishes and prawns large enough to eat the snail vectors may rapidly diminish due to indiscriminate fishing by residents of communities where schistosomiasis is endemic, since these molluscivores are also a major source of food for humans. Moreover, introduction of competitor species of snails could greatly endanger agriculture and the ecosystem. On the other hand, environmental modifications (such as removal of vegetation on which the snail vectors feed, lining canals with cement, or draining water habitats) are very expensive and impractical for resource-constrained areas. Meanwhile, vegetation removal poses an increased risk of infection to workers who may not have protective tools [24]. Generally, however, certain issues concerning the use of gene drive systems have come into view. The most important of all include potential off-target mutations that may result in unpredictable effects, development of drive resistance in populations, fitness and competitiveness of released strains compared to wild populations, and possible difficulty in the containment, reversal, or adjustment of gene drive spread [83,84,105]. Nevertheless, it is somewhat relieving that a good number of these limitations can feasibly be overcome through the meticulous design of more specific sgRNAs, and development of reversal drive systems [84,89,91,92,97,105]. Moreover, the majority of the current issues regarding the application of gene drives for the control of disease vectors arose from studies focusing on mosquitoes, implying that some of the risk issues, such as vector dispersal beyond intended political boundaries [84], may be of lesser concern in other non-insect vector control systems. Conversely however, the significant body of research on mosquitoes may have also overcome some series of technical challenges that may remain unresolved for other disease vectors.
Should a breakthrough on the use of CRISPR-based vector control occur, the fine line between mating/reproductive biology of Oncomelania and that of Bulinus or Biomphalaria, as well as the varying degree of selfing among species of the hermaphroditic (Bulinus and Biomphalaria) snail vectors, will also have important implications in schistosomiasis snail control application. As shown in Figure 1, CRISPR/Cas9-driven resistance traits may spread more rapidly among successive progeny of Oncomelania (being a dioecious outcrossing vector) than in Bulinus and Biomphalaria snail vectors. More precisely, in the two latter snail vectors, gene drive approach may not be effective in predominantly selfing species, such as Bulinus truncatus, Bulinus forskalii, and Biomphalaria pfeifferi.
5. Conclusions
The prospective use of genetically manipulated vectors to stop the spread of vector-borne diseases maintains its impressiveness and is awaited by the scientific community. In fast-tracking sustainable schistosomiasis elimination, the use of CRISPR-based vector modification strategy appears fascinating and potentially effective. However, this approach is currently still underdeveloped in snail molecular research. Finding the pertinent missing pieces in our jigsaw of knowledge of schistosome/snail biology, and identifying ways to bypass potential future challenges, are requisites for achieving this promising snail control strategy. Finally, the use of schistosome-resistant transgenic snails may have the propensity to singly interrupt schistosomiasis transmission when only outcrossing vector species are present, but in foci where both predominantly selfing species and outcrossing species of Bulinus or Biomphalaria snails coexist, the integration of additional suitable snail control methods will provide a way of complementing this genetic control method for more effective outcomes.
Funding
This research received no external funding.
Acknowledgments
The author would like to thank Officer Adewale Adeniyi of the Nigerian Customs Service (NCS) for his financial assistance.
Conflicts of Interest
The author declares no conflict of interest.
References
- World Health Organization. Schistosomiasis and soil-transmitted helminthiasis: Number of people treated in 2016. Wkly. Epidemiol. Rec. 2017, 92, 49–60. [Google Scholar]
- World Health Organization. Global Health Estimates Summary Tables. DALYs by Cause, Age and Sex, by WHO Region, 2000–2015; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Colley, D.G.; Bustinduy, A.L.; Secor, W.E.; King, C.H. Human schistosomiasis. Lancet 2014, 383, 2253–2264. [Google Scholar] [CrossRef] [Green Version]
- Muller, R.; Wakelin, D. Worms and Human Disease, 2nd ed.; CABI Publishing: Oxon, UK; New York, NY, USA, 2002; pp. 1–300. [Google Scholar]
- Adenowo, A.B.; Oyinloye, B.E.; Ogunyinka, B.I.; Kappo, A.B. Impact of human schistosomiasis in sub-Saharan Africa. Braz. J. Infect. Dis. 2015, 19, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Famakinde, D.O. Molecular context of Schistosoma mansoni transmission in the molluscan environments: A mini-review. Acta Trop. 2017, 176, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Olveda, D.U.; Li, Y.; Olveda, R.M.; Lam, A.K.; Chau, T.N.P.; Harn, D.A.; Williams, G.M.; Gray, D.J.; Ross, A.G.P. Bilharzia: Pathology, diagnosis, management and control. Trop. Med. Surg. 2013, 1, 135. [Google Scholar] [CrossRef] [PubMed]
- Colley, D.G.; Secor, W.E. Immunology of human schistosomiasis. Parasite Immunol. 2014, 36, 347–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, J.F.; Mital, P.; Kanzaria, H.K.; Olds, G.R.; Kurtis, J.D. Schistosomiasis and pregnancy. Trends Parasitol. 2007, 23, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Salawu, O.T.; Odaibo, A.B. Maternal schistosomiasis: A growing concern in sub-Saharan Africa. Pathog. Glob. Health 2014, 108, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Kjetland, E.F.; Ndhlovu, P.D.; Gomo, E.; Mduluza, T.; Midzi, N.; Gwanzura, L.; Mason, P.R.; Sandvik, L.; Friis, H.; Gunderseen, S.G. Association between genital schistosomiasis and HIV in rural Zimbabwean women. AIDS 2006, 20, 593–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mbabazi, P.S.; Andan, O.; Fitzgerald, D.W.; Chitsulo, L.; Engels, D.; Downs, J.A. Examining the relationship between urogenital schistosomiasis and HIV infection. PLoS Negl. Trop. Dis. 2011, 5, e1396. [Google Scholar] [CrossRef] [PubMed]
- Bustinduy, A.; King, C.; Scott, J.; Appleton, S.; Sousa-Figueiredo, J.C.; Betson, M.; Stothard, J.R. HIV and schistosomiasis co-infection in African children. Lancet Infect. Dis. 2014, 14, 640–649. [Google Scholar] [CrossRef] [Green Version]
- Tchuenté, L.-A.T.; Rollinson, D.; Stothard, J.R.; Molyneux, D. Moving from control to elimination of schistosomiasis in sub-Saharan Africa: Time to change and adapt strategies. Infect. Dis. Poverty 2017, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- King, C.H.; Sutherland, L.J.; Bertsch, D. Systematic review and meta-analysis of the impact of chemical-based mollusciciding for control of Schistosoma mansoni and S. haematobium transmission. PLoS Negl. Trop. Dis. 2015, 9, e0004290. [Google Scholar] [CrossRef] [PubMed]
- Lo, N.C.; Gurarie, D.; Toon, N.; Coulibaly, J.T.; Bendavid, E.; Andrews, J.R.; King, C.H. Impact and cost-effectiveness of snail control to achieve disease control targets for schistosomiasis. Proc. Natl. Acad. Sci. USA 2018, 115, E584–E591. [Google Scholar] [CrossRef] [PubMed]
- Sokolow, S.H.; Wood, C.L.; Jones, I.J.; Lafferty, K.D.; Kuris, A.M.; Hseih, M.H.; De Loe, G.A. To reduce the global burden of human schistosomiasis, use ‘old fashioned’ snail control. Trends Parasitol. 2018, 34, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Sokolow, S.H.; Wood, C.L.; Jones, I.J.; Swartz, S.J.; Lopez, M.; Hseih, M.H.; Lafferty, K.D.; Kuris, A.M.; Rickards, C.; De Leo, G.A. Global assessment of schistosomiasis control over the past century shows targeting the snail intermediate host works best. PLoS. Negl. Trop. Dis. 2016, 10, e0004794. [Google Scholar] [CrossRef] [PubMed]
- Alsaqabi, S.M.; Lofty, W.M. Praziquantel: A review. J. Vet. Sci. Technol. 2014, 5, 1000200. [Google Scholar] [CrossRef]
- Cioli, D.; Pica-Mattoccia, L.; Basso, A.; Guidi, A. Schistosomiasis control: Praziquantel forever? Mol. Biochem. Parasitol. 2014, 195, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Tebeje, B.M.; Harvie, M.; You, H.; Loukas, A.; McManus, D.P. Schistosomiasis vaccines: Where do we stand? Parasit. Vectors 2016, 9, 528. [Google Scholar] [CrossRef] [PubMed]
- Ebisawa, I. Epidemiology and eradication of schistosomiasis japonica in Japan. J. Travel Med. 1998, 5, 33–35. [Google Scholar] [CrossRef] [PubMed]
- Barakat, R.M.R. Epidemiology of schistosomiasis in Egypt: Travel through time: Review. J. Adv. Res. 2013, 4, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Secor, W.E. Water-based interventions for schistosomiasis control. Pathog. Glob. Health 2014, 108, 246–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tlamçani, Z.; Er-Rami, M. Schistosomiasis control: Moroccan experience compared to other endemic countries. Asian Pac. J. Trop. Dis. 2014, 4, 329–332. [Google Scholar] [CrossRef]
- King, C.H.; Bertsch, D. Historical perspective: Snail control to prevent schistosomiasis. PLoS Negl. Trop. Dis. 2015, 9, e0003657. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.P.; Wang, W.; Hong, Q.B.; Li, S.Z.; Liang, Y.S.; Yang, H.T.; Zhou, X.N. Approaches being used in the national schistosomiasis elimination programme in China: A review. Infect. Dis. Poverty 2017, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Lardans, V.; Dissous, C. Snail control strategies for reduction of schistosomiasis transmission. Parasitol. Today 1998, 14, 413–417. [Google Scholar] [CrossRef]
- Huang, D.; Zhen, J.; Quan, S.; Liu, M.; Liu, L. Risk assessment for niclosamide residues in water and sediments from Nan Ji Shan Island within Poyang Lake region, China. Adv. Mat. Res. 2013, 721, 608–612. [Google Scholar] [CrossRef]
- World Health Organization. Field Use of Molluscicides in Schistosomiasis Control Programmes: An Operational Manual for Programme Managers; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- World Health Assembly. Elimination of Schistosomiasis. In Sixty-fifth World Health Assembly: Resolutions and Decisions; World Health Organization: Geneva, Switzerland, 2012; pp. 36–37. [Google Scholar]
- Miyairi, K.; Suzuki, M. On the development of Schistosoma japonicum. Tokyo Med. J. 1913, 1836, 1961–1965. [Google Scholar]
- Tanaka, H.; Tsuji, M. From discovery to eradication of schistosomiasis in Japan: 1847–1996. Int. J. Parasitol. 1997, 27, 1465–1480. [Google Scholar] [CrossRef]
- Leiper, R.T. On the relation between the terminal-spined and lateral-spined eggs of bilharzia. Br. Med. J. 1916, 1, 411. [Google Scholar] [CrossRef] [PubMed]
- Leiper, R.T. Report on the results of the bilharzias mission in Egypt, 1915. J. R. Army Med. Corps 1918, 30, 235–260. [Google Scholar]
- Newton, W.L. The comparative tissue reaction of two strains of Australorbis glabratus to infection with Schistosoma mansoni. J. Parasitol. 1952, 38, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Newton, W.L. The inheritance of susceptibility to infection with Schistosoma mansoni in Australorbis glabratus. Exp. Parasitol. 1953, 2, 242–257. [Google Scholar] [CrossRef]
- Richards, C.S. Genetic studies of a molluscan vector of schistosomiasis. Nature 1970, 227, 231–241. [Google Scholar] [CrossRef]
- Richards, C.S. Genetic factors in susceptibility of Biomphalaria glabrata for different strains of Schistosoma mansoni. Parasitology 1975, 70, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Richards, C.S. Schistosoma mansoni: susceptibility reversal with age in the snail host. Exp. Parasitol. 1977, 42, 165–168. [Google Scholar] [CrossRef]
- Richards, C.S.; Knight, M.; Lewis, F.A. Genetics of Biomphalaria glabrata and its effects on the outcome of Schistosoma mansoni infection. Parasitol. Today 1992, 8, 171–174. [Google Scholar] [CrossRef]
- Abou-El-Naga, I.F.; Eissa, M.M.; Mossallam, S.F.; El-Halim, S.I.A. Inheritance of Schistosoma mansoni infection incompatibility in Biomphalaria alexandrina snails. Mem. do Inst. Oswaldo Cruz 2010, 105, 149–154. [Google Scholar] [CrossRef]
- Dos Santos, M.B.L.; Freitas, J.R.; Correia, M.C.R.; Coelho, P.M.Z. Susceptibility of Biomphalaria tenagophila hybrids to Schistosoma mansoni: Crossing between strains from Taim (RS), Cabo Frio (RJ), and Belo Horizonte (MG), Brasil. Rev. Inst. de Med. Trop. São Paulo 1979, 21, 281–286. [Google Scholar]
- Knight, M.; Miller, A.N.; Patterson, C.N.; Rowe, C.G.; Michaels, G.; Carr, D.; Richards, C.S.; Lewis, F.A. The identification of markers segregating with resistance to Schistosoma mansoni infection in the snail Biomphalaria glabrata. Proc. Natl. Acad. Sci. USA 1999, 96, 1510–1515. [Google Scholar] [CrossRef] [PubMed]
- Rosa, F.M.; Godard, A.L.B.; Azevedo, V.; Coelho, P.M.Z. Biomphalaria tenagophila: dominant character of the resistance to Schistosoma mansoni in descendants of crossbreedings between resistant (Taim, RS) and susceptible (Joinville, SC) strains. Mem. do Inst. Oswaldo Cruz 2005, 100, 19–23. [Google Scholar] [CrossRef]
- Haggag, S.H.; El-Sherbiny, M. Molecular markers associated with resistance to Schistosoma mansoni infection in the Biomphalaria glabrata snails. Biotechnology 2006, 5, 404–412. [Google Scholar]
- Ittiprasert, W.; Miller, A.; Myers, J.; Nene, V.; El-Sayed, N.M.; Knight, M. Identification of immediate response genes dominantly expressed in juvenile resistant and susceptible Biomphalaria glabrata snails upon exposure to Schistosoma mansoni. Mol. Biochem. Parasitol. 2010, 169, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Paraense, W.L.; Correa, L.R. Variation in susceptibility of populations of Australorbis glabratus to a strain of Schistosoma mansoni. Rev. Inst. Med. Trop. São Paulo 1963, 5, 15–22. [Google Scholar] [PubMed]
- Richards, C.S. Susceptibility of adult Biomphalaria glabrata to Schistosoma mansoni infection. Am. J. Trop. Med. Hyg. 1973, 22, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Goodall, C.P.; Bender, R.C.; Brooks, J.K.; Bayne, C.J. Biomphalaria glabrata cytosolic copper/zinc superoxide dismutase (SOD1) gene: Association of SOD1 alleles with resistance/susceptibility to Schistosoma mansoni. Mol. Biochem. Parasitol. 2006, 147, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Newton, W.L. The establishment of a strain of Australorbis glabratus which combines albinism and high susceptibility to infection with Schistosoma mansoni. J. Parasitol. 1955, 41, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Adema, C.M.; Luo, M.Z.; Hanelt, B.; Hertel, L.A.; Marshall, J.J.; Zhang, S.M.; DeJong, R.J.; Kim, H.R.; Kudrna, D.; Wing, R.A.; et al. A bacterial artificial chromosome library for Biomphalaria glabrata, intermediate snail host of Schistosoma mansoni. Mem. Inst. Oswaldo Cruz 2006, 101, S167–S177. [Google Scholar] [CrossRef]
- Ittiprasert, W.; Myers, J.; Odoemelam, E.C.; Raghavan, N.; Lewis, F.; Bridger, J.M.; Knight, M. Advances in the genomics and proteomics of the freshwater intermediate snail host of Schistosoma mansoni, Biomphalaria glabrata. In Biomphalaria Snails and Larval Trematodes; Toledo, R., Fried, B., Eds.; Springer: New York, NY, USA, 2011; pp. 191–213. [Google Scholar]
- Pinaud, S.; Portela, J.; Duval, D.; Nowacki, F.C.; Olive, M.A.; Allienne, J.F.; Galinier, R.; Dheilly, N.M.; Kieffer-Jaquinod, S.; Mitta, G.; et al. A shift from cellular to humoral responses contributes to innate immune memory in the vector snail Biomphalaria glabrata. PLoS Pathog. 2016, 12, e1005361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshino, T.P.; Coustau, C. Immunobiology of Biomphalaria–trematode interactions. In Biomphalaria Snails and Larval Trematodes; Toledo, R., Fried, B., Eds.; Springer: New York, NY, USA, 2011; pp. 159–189. [Google Scholar]
- Pila, E.A.; Li, H.; Hambrook, J.R.; Wu, X.; Hanington, P. Schistosomiasis from a snail’s perspective: Advances in snail immunity. Trends Parasitol. 2017, 33, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Hansen, E.L. A cell line from embryos of Biomphalaria glabrata (Pulmonata): Establishment and characteristics. In Invertebrate Tissue Culture: Research Applications; Maramorosch, K., Ed.; Academic Press: New York, NY, USA, 1976; pp. 75–97. [Google Scholar]
- Adema, C.M.; Hillier, L.D.W.; Jones, C.S.; Loker, E.S.; Knight, M.; Minx, P.; Oliveira, G.; Raghavan, N.; Shedlock, A.; do Amaral, L.R.; et al. Whole genome analysis of a schistosomiasis-transmitting freshwater snail. Nat. Comm. 2017, 8, 15451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahoor, Z.; Lockyer, A.E.; Davies, A.J.; Kirk, R.S.; Emery, A.M.; Rollinson, D.; Jones, C.S.; Noble, L.R.; Walker, A.J. Differences in the gene expression profiles of haemocytes from schistosome-susceptible and -resistant Biomphalaria glabrata exposed to Schistosoma mansoni excretory secretory products. PLoS ONE 2014, 9, e93215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, M.K.; Bender, R.C.; Bayne, C.J. Resistance of Biomphalaria glabrata 13–16-R1 snails to Schistosoma mansoni PR1 is a function of haemocyte abundance and constitutive levels of specific transcripts in haemocytes. Int. J. Parasitol. 2014, 44, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Pila, E.A.; Gordy, M.A.; Phillips, V.K.; Kabore, A.L.; Rudkom, S.P.; Hanington, P.C. Endogenous growth factor stimulation of hemocyte proliferation induces resistance to Schistosoma mansoni challenge in the snail host. Proc. Natl. Acad. Sci. USA 2016, 113, 5305–5310. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.B.; Pierce, R.J.; Gourbal, B.; Werkmeister, E.; Colinet, D.; Reichhart, J.; Dissous, C.; Coustau, C. Involvement of the cytokine MIF in the snail host immune response to the parasite Schistosoma mansoni. PLoS Pathog. 2010, 6, e1001115. [Google Scholar]
- Pila, E.A.; Tarrabain, M.; Kabore, A.L.; Hanington, P.C. A novel toll-like receptor (TLR) influences compatibility between the gastropod Biomphalaria glabrata, and the digenean trematode Schistosoma mansoni. PLoS Pathog. 2016, 12, e1005513. [Google Scholar] [CrossRef] [PubMed]
- Galinier, R.; Portela, J.; Mone, Y.; Allienne, J.F.; Henri, H.; Delbecq, S.; Mitta, G.; Gourbal, B.; Duval, D. Biomphalysin, a new β pore-forming toxin involved in Biomphalaria glabrata immune defense against Schistosoma mansoni. PLoS Pathog. 2013, 9, e1003216. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.; Ittiprasert, N.; Raghavan, N.; Miller, A.; Knight, M. Differences in cysteine protease activity in Schistosoma mansoni-resistant and -susceptible Biomphalaria glabrata and characterization of the hepatopancreas cathepsin B fulllength cDNA. J. Parasitol. 2008, 94, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, A.E.; Spinks, J.; Kane, R.A.; Hoffmann, K.F.; Fitzpatrick, J.M.; Rollinson, D.; Noble, L.R.; Jones, C.S. Biomphalaria glabrata transcriptome: cDNA microarray profiling identifies resistant- and susceptible-specific gene expression in haemocytes from snail strains exposed to Schistosoma mansoni. BMC Genomics 2008, 9, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Dinguirard, N.; Sabat, G.; Lui, H.; Gonzalez, L.; Gehring, M.; Bickham-Wright, U.; Yoshino, T.P. Proteomic analysis of Biomphalaria glabrata plasma proteins with binding affinity to those expressed by early developing larval Schistosoma mansoni. PLoS Pathog. 2017, 13, e1006081. [Google Scholar] [CrossRef] [PubMed]
- Bonner, K.M.; Bayne, C.J.; Larson, M.K.; Blouin, M.S. Effects of Cu/Zn superoxide dismutase (sod1) genotype and genetic background on growth, reproduction and defense in Biomphalaria glabrata. PLoS Negl. Trop. Dis. 2012, 6, e1701. [Google Scholar] [CrossRef] [PubMed]
- Tennessen, J.A.; Bonner, K.M.; Bollmann, S.R.; Johnstun, J.A.; Yeh, J.Y.; Marine, M.; Tavalire, H.F.; Bayne, C.J.; Blouin, M.S. Genome-wide scan and test of candidate genes in the snail Biomphalaria glabrata reveal new locus influencing resistance to Schistosoma mansoni. PLoSNegl. Trop. Dis. 2015, 9, e0004077. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, A.E.; Emery, A.M.; Kane, R.A.; Walker, A.J.; Mayer, C.D.; Mitta, G.; Coustau, C.; Adema, C.M.; Hanelt, B.; Rollinson, D.; et al. Early differential gene expression in haemocytes from resistant and susceptible Biomphalaria glabrata strains in response to Schistosoma mansoni. PLoS ONE 2012, 7, e51102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, T.S.; Woodards, A.C.; Jung, Y.; Adema, C.M.; Loker, E.S. Identification of transcripts generated during the response of resistant Biomphalaria glabrata to Schistosoma mansoni infection using suppression subtractive hybridization. J. Parasitol. 2004, 90, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Hanington, P.C.; Forys, M.A.; Dragoo, J.W.; Zhang, S.; Adema, C.M.; Loker, E.S. Role for a somatically diversified lectin in resistance of an invertebrate to parasite infection. Proc. Natl. Acad. Sci. USA 2010, 107, 21087–21092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanington, P.C.; Forys, M.A.; Loker, E.S. A somatically diversified defense factor, FREP3, is a determinant of snail resistance to schistosome infection. PLoS Negl. Trop. Dis. 2012, 6, e1591. [Google Scholar] [CrossRef] [PubMed]
- Martins-Souza, R.L.; Pereira, C.A.J.; Rodrigues, L.; Araújo, E.S.; Coelho, P.M.Z.; Corrêa, A., Jr.; Negrão-Corrêa, D. Participation of N-acetyl-D-glucosamine carbohydrate moieties in the recognition of Schistosoma mansoni sporocysts by haemocytes of Biomphalaria tenagophila. Mem. Inst. Oswaldo Cruz 2011, 106, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Allan, E.R.O.; Tennessen, J.A.; Bollmann, S.R.; Hanington, P.C.; Bayne, C.J.; Blouin, M.S. Schistosome infectivity in the snail, Biomphalaria glabrata, is partially dependent on the expression of Grctm6, a Guadeloupe Resistance Complex protein. PLoS Negl. Trop. Dis. 2017, 11, e0005362. [Google Scholar] [CrossRef] [PubMed]
- Ittiprasert, W.; Knight, M. Reversing the resistance phenotype of the Biomphalaria glabrata snail host Schistosoma mansoni infection by temperature modulation. PLoS Pathog. 2012, 8, e1002677. [Google Scholar] [CrossRef] [PubMed]
- Ittiprasert, W.; Nene, R.; Miller, A.; Raghavan, N.; Lewis, F.; Hodgson, J.; Knight, M. Schistosoma mansoni infection of juvenile Biomphalaria glabrata induces a differential stress response between resistant and susceptible snails. Exp. Parasitol. 2009, 123, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.; Elhelu, O.; Smith, M.; Haugen, B.; Miller, A.; Raghavan, N.; Wellman, C.; Cousin, C.; Dixon, F.; Mann, V.; et al. Susceptibility of snails to infection with schistosomes is influenced by temperature and expression of heat shock proteins. Epidemiology 2015, 5, 1–18. [Google Scholar]
- Granath, W.O., Jr.; Connors, V.A.; Tarleton, R.L. Interleukin 1 activity haemolymph from strains of the snail Biomphalaria glabrata varying in susceptibility to the human blood fluke, Schistosoma mansoni: Presence, differential expression, and biological function. Cytokine 1994, 6, 21–27. [Google Scholar] [CrossRef]
- Zhang, S.M.; Coultas, K.A. Identification and characterization of five transcription factors that are associated with evolutionarily conserved immune signaling pathways in the schistosome-transmitting snail Biomphalaria glabrata. Mol. Immunol. 2011, 48, 1868–1881. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.; Raghavan, N.; Goodal, C.; Cousin, C.; Ittiprasert, W.; Sayed, A.; Miller, A.; Williams, D.L.; Bayne, C. Biomphalaria glabrata peroxiredoxin: effect of Schistosoma mansoni infection on differential gene regulation. Mol. Biochem. Parasitol. 2009, 167, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Ouwe-Missi-Oukem-Boyer, O.; Porchet, E.; Capron, A.; Dissous, C. Characterization of immunoreactive TNF-α molecules in the gastropod Biomphalaria glabrata. Dev. Comp. Immunol. 1994, 18, 211–218. [Google Scholar] [CrossRef]
- Alphey, L. Genetic control of mosquitoes. Annu. Rev. Emtomol. 2014, 59, 205–224. [Google Scholar] [CrossRef] [PubMed]
- Macias, V.M.; Ohm, J.R.; Rasgon, J.L. Gene drive for mosquito control: Where did it come from and where are we headed? Int. J. Environ. Res. Public Health 2017, 14, 1006. [Google Scholar] [CrossRef] [PubMed]
- Hubendick, B. A possible method of schistosome-vector control by competition between resistant and susceptible strains. Bull. World Health Organ. 1958, 18, 1113–1116. [Google Scholar] [PubMed]
- Rozendaal, J.A.; World Health Organization. Freshwater snails. In Vector Control: Methods for Use by Individuals and Communities; World Health Organization: Geneva, Switzerland, 1997; pp. 337–356. [Google Scholar]
- Claveria, F.G.; Etges, F.J. Differential susceptibility of male and female Oncomelania hupensis quadrasi infected with Schistosoma japonicum. Int. J. Parasitol. 1987, 17, 1273–1277. [Google Scholar] [CrossRef]
- Moose, J.W.; Williams, J.E. Infection rates of Schistosoma japonicum in experimentally exposed female and male oncomelanid snails. Jan. J. Med. Sci. Biol. 1964, 17, 333–334. [Google Scholar] [CrossRef]
- Sun, D.; Gou, Z.; Liu, Y.; Zhang, Y. Progress and prospects of CRISPR/Cas systems in insects and other arthropods. Front. Physiol. 2017, 8, 608. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Y.; Gao, F.; Han, S.; Cheah, K.S.; Tse, H.F.; Lian, Q. CRISPR/Cas9 genome-editing system in human stem cells: Current status and future prospects. Mol. Ther. Nucleic Acids 2017, 9, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Braddick, D.; Dhar, P.K. Exploring the potential of genome editing CRISPR-Cas9 technology. Gene 2017, 599, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, S.; Xu, J.; Suin, C.; Wei, J. Application of CRISPR/Cas9 in plant biology. Acta Pharm. Sin. B 2017, 7, 292–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Fisher, A.L.; Huang, H.; Xie, Z. CRISR-mediated genome editing and human diseases. Genes Dis. 2016, 3, 244–251. [Google Scholar] [CrossRef]
- Mouahid, G.; Rognon, A.; de Carvalho-Augusto, R.; Driguez, P.; Geyer, K.; Karinshak, S.; Luviano, N.; Mann, V.; Quack, T.; Rawlinson, K.; et al. Transplantation of schistosome sporocysts between host snails: A video guide. Wellcome Open Res. 2018, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tennessen, J.A.; Théron, A.; Marine, M.; Yeh, J.Y.; Rognon, A.; Blouin, M.S. Hyperdiverse gene cluster in snail host conveys resistance to human schistosome parasites. PLoS Genet. 2015, 11, e1005067. [Google Scholar] [CrossRef] [PubMed]
- Perry, K.J.; Henry, J.Q. CRISPR/Cas9-mediated genome modification in the mollusc, Crepidula fornicata. Genesis 2015, 53, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Bortesi, L.; Zhu, C.; Zischewski, J.; Perez, L.; Bassié, L.; Nadi, R.; Forni, G.; Lade, S.B.; Soto, E.; Jin, X.; et al. Patterns of CRISPR/Cas9 activity in plants, animals and microbes. Plant Biotechnol. J. 2016, 14, 2203–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Li, F.; Dang, L.; Liang, D.; Wang, C.; He, B.; Liu, J.; Li, D.; Wu, X.; Xu, X.; et al. In vivo delivery systems for therapeutic genome editing. Int. J. Mol. Sci. 2016, 17, 626. [Google Scholar] [CrossRef] [PubMed]
- Jarne, P.; Pointier, J.-P.; David, P. Biosystematics of Biomphalaria spp. with an emphasis on Biomphalaria glabrata. In Biomphalaria Snails and Larval Trematodes; Toledo, R., Fried, B., Eds.; Springer: New York, NY, USA, 2011; pp. 1–32. [Google Scholar]
- Blouin, M.S.; Bonner, K.M.; Cooper, B.; Amarasinghe, V.; O’Donnell, R.P.; Bayne, C.J. Three genes involved in the oxidative burst are closely linked in the genome of the snail, Biomphalaria glabrata. Int. J. Parasitol. 2013, 43, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Berg, C.O. Biological control of snail-borne diseases: A review. Exp. Parasitol. 1973, 33, 318–330. [Google Scholar] [CrossRef]
- Galinier, R.; Roger, E.; Moné, Y.; Duval, D.; Portet, A.; Pinaud, S.; Chaparro, C.; Grunau, C.; Genthon, C.; Dubois, E.; et al. A multistrain approach to studying the mechanisms underlying compatibility in the interaction between Biomphalaria glabrata and Schistosoma mansoni. PLoS Negl. Trop. Dis. 2017, 11, e0005398. [Google Scholar] [CrossRef] [PubMed]
- Mitta, G.; Adema, C.M.; Gourbal, B.; Loker, E.S.; Theron, A. Compatibility polymorphism in snail/schistosome interactions: From field to theory to molecular mechanisms. Dev. Comp. Immunol. 2012, 37, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.S.; Wang, A.Y.; Zhao, H.B.; Chen, Y.H. Transcriptome sequencing and differential gene expression analysis of the schistosome-transmitting snail Oncomelania hupensis inhabiting hilly and marshland regions. Sci. Rep. 2017, 7, 15809. [Google Scholar] [CrossRef] [PubMed]
- Esvelt, K.M.; Smidler, A.L.; Catteruccia, F.; Church, G.M. Concerning RNA-guided gene drives for the alteration of wild population. eLife 2014, 3, e03401. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Transgenic snail system for field control of schistosomiasis transmission.

Figure 2.
A schematic representation of CRISPR/Cas9 genome editing system in a snail vector of schistosome. Abbreviations: DSB, double-strand break; HDR, homology-directed repair; P1 & P2, promoters; PCR, polymerase chain reaction; RE, restriction enzyme; T7E1, T7 endonuclease I.
Figure 2.
A schematic representation of CRISPR/Cas9 genome editing system in a snail vector of schistosome. Abbreviations: DSB, double-strand break; HDR, homology-directed repair; P1 & P2, promoters; PCR, polymerase chain reaction; RE, restriction enzyme; T7E1, T7 endonuclease I.


{kind=link}
{kind=link}
Table 1.
Putative genes and proteins conferring Biomphalaria resistance to Schistosoma infection.
| Resistance Factor | Snail spp. | Schistosoma spp. | Function | Reference(s) |
|---|---|---|---|---|
| 40S ribosomal protein S9 | B. glabrata | S. mansoni | Protein translation in hemocytes. | [59] |
| BgAIF | B. glabrata | S. mansoni | Modulates hemocyte activation. | [60] |
| BgGRN | B. glabrata | S. mansoni | Production of adherent hemocytes. | [61] |
| BgMIF | B. glabrata | S. mansoni | Induces hemocyte proliferation. | [62] |
| BgTLR | B. glabrata | S. mansoni | Parasite recognition and activation of effector functions. | [63] |
| Biomphalysin | B. glabrata | S. mansoni | Binds to the sporocyst surface and lyses it. | [54,64] |
| Cathepsin B | B. glabrata | S. mansoni | Lysis of encapsulated sporocyst. | [65] |
| Cathepsin L | B. glabrata | S. mansoni | Lysis of encapsulated sporocyst. | [66] |
| Copine 1 | B. glabrata | S. mansoni | Involves in signaling processes. | [66] |
| CREPs | B. glabrata | S. mansoni | Pattern recognition receptors/adhesion proteins. | [67] |
| Cu/Zn SOD (SOD1) | B. glabrata | S. mansoni | Catalyzes the production of H2O2 which is cytotoxic to sporocyst. | [50,68,69] |
| Cystatin 2 | B. glabrata | S. mansoni | Protease inhibitor. | [70,71] |
| Cytidine deaminase | B. glabrata | S. mansoni | Nucleobase, nucleoside, nucleotide, and nucleic acid metabolism. | [47] |
| Cytochrome b | B. glabrata | S. mansoni | Mitochondrial respiration. | [70] |
| Cytochrome C oxidase subunits | B. glabrata | S. mansoni | Mitochondrial respiration. | [70,71] |
| Dermatopontin2 | B. glabrata | S. mansoni | Participates in hemocyte adhesion and encapsulation responses. | [59,67,70] |
| Elastase2 | B. glabrata | S. mansoni | Lysis of encapsulated sporocyst. | [66,70] |
| Elongation factors 1α & 2 | B. glabrata | S. mansoni | Transcription enzymes (bind t-RNA to ribosomes). | [59,67] |
| Endo-1,4-β-glucanase | B. glabrata | S. mansoni | Carbohydrate metabolism. | [70] |
| Ferritin | B. glabrata | S. mansoni | Stores and transport iron in non-toxic form. | [70,71] |
| FREP1, 2, 3 & 12 | B. glabrata | S. mansoni | Pattern recognition receptors/adhesion proteins. | [67,70,72,73] |
| Fribillin | B. glabrata | S. mansoni | Participates in hemocyte adhesion and encapsulation responses. | [70] |
| GlcNAc ↓ | B. tenagophila | S. mansoni | Increases hemocyte binding to sporocyst. | [74] |
| GPCR kinase 2 | B. glabrata | S. mansoni | Signal transduction. | [70] |
| Grctm6 | B. glabrata | S. mansoni | Modulates cercarial shedding. | [75] |
| GREPs | B. glabrata | S. mansoni | Pattern recognition receptors/adhesion proteins. | [67] |
| GSTs | B. glabrata | S. mansoni | Prevent cellular damage to the hemocytes. | [70] |
| Hsp40, 60 & 70 # | B. glabrata | S. mansoni | Housekeeping cell repair activities. | [66,67,70,76,77,78] |
| Importin 7 | B. glabrata | S. mansoni | Involves in signaling processes. | [66] |
| Inferred phagocyte oxidase | B. glabrata | S. mansoni | Production of superoxide anions. | [60] |
| Interleukin 1 | B. glabrata | S. mansoni | Stimulates hemocyte defense response. | [79] |
| LPS-binding protein | B. glabrata | S. mansoni | Adhesion protein. | [67] |
| Matrilin | B. glabrata | S. mansoni | Participates in hemocyte adhesion and encapsulation responses. | [59,70] |
| Metalloproteases | B. glabrata | S. mansoni | Tissue morphogenesis/remodeling. | [67] |
| MPEG 1 | B. glabrata | S. mansoni | Participates in hemocyte defense responses. | [47] |
| Neo-calmodulin | B. glabrata | S. mansoni | Cacium signaling and homeostasis. | [67] |
| NF-kB | B. glabrata | S. mansoni | Downstream transcription in the TLR pathway. | [59,63,70,80] |
| NADH dehydrogenase subunis | B. glabrata | S. mansoni | Mitochondrial respiration. | [70] |
| Peroxiredoxines 1 & 4 | B. glabrata | S. mansoni | Neutralize ROS and RNS that can damage cellular functions. | [60,81] |
| PGRP 1 | B. glabrata | S. mansoni | Pattern recognition receptor. | [70] |
| PKC receptor | B. glabrata | S. mansoni | Signal transduction. | [47] |
| TEPs | B. glabrata | S. mansoni | Pattern recognition receptors/adhesion proteins. | [67] |
| TNF-α | B. glabrata | S. mansoni | Stimulates hemocyte defense response. | [82] |
Symbols: ↓ in lower concentrations; # contrasting reports (see [67,78] for some details). Abbreviations: BgAIF, B. glabrata allograft inflammatory factor; BgGRN, B. glabrata granulin; BgMIF, B. glabrata macrophage migration-inhibitory factor; BgTLR, B. glabrata Toll-like receptor; CREP, C-type lectin-related protein; Cu/Zn SOD, copper/zinc superoxide dismutase; FREP, fibrinogen-related protein; GlcNac, N-acetyl-D-glucosamine; GPCR, G-protein coupled receptor; Grctm, Guadeloupe resistance complex transmembrane; GREP, galectin-related proteins; GSTs, glutathione-S-transferases; H2O2, hydrogen peroxide; Hsp, heat shock protein; LPS, lipopolysaccharide; MPEG, macrophage expressed gene; NADH, reduced nicotinamide adenine dinucleotide; NF-kB, nuclear factor kappa B; PKC, protein kinase C; PGRP, peptidoglycan recognition protein; RNS, reactive nitrogen species; ROS, reactive oxygen species; t-RNA, transfer ribonucleic acid; TEP, thioester-containing protein; TNF-α, tumor necrosis factor-alpha.
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Famakinde, D.O. Treading the Path towards Genetic Control of Snail Resistance to Schistosome Infection. Trop. Med. Infect. Dis. 2018, 3, 86. https://doi.org/10.3390/tropicalmed3030086
AMA Style
Famakinde DO. Treading the Path towards Genetic Control of Snail Resistance to Schistosome Infection. Tropical Medicine and Infectious Disease. 2018; 3(3):86. https://doi.org/10.3390/tropicalmed3030086
Chicago/Turabian StyleFamakinde, Damilare O. 2018. "Treading the Path towards Genetic Control of Snail Resistance to Schistosome Infection" Tropical Medicine and Infectious Disease 3, no. 3: 86. https://doi.org/10.3390/tropicalmed3030086