
Mammoths, Deer, and a Dog: Fossil and (Sub) Recent Allochthonous Remains from the Northeastern Croatia (Podravina Region), with the First Radiocarbon Dating of the Croatian Woolly Mammoths (Mammuthus primigenius)

Abstract
:1. Introduction
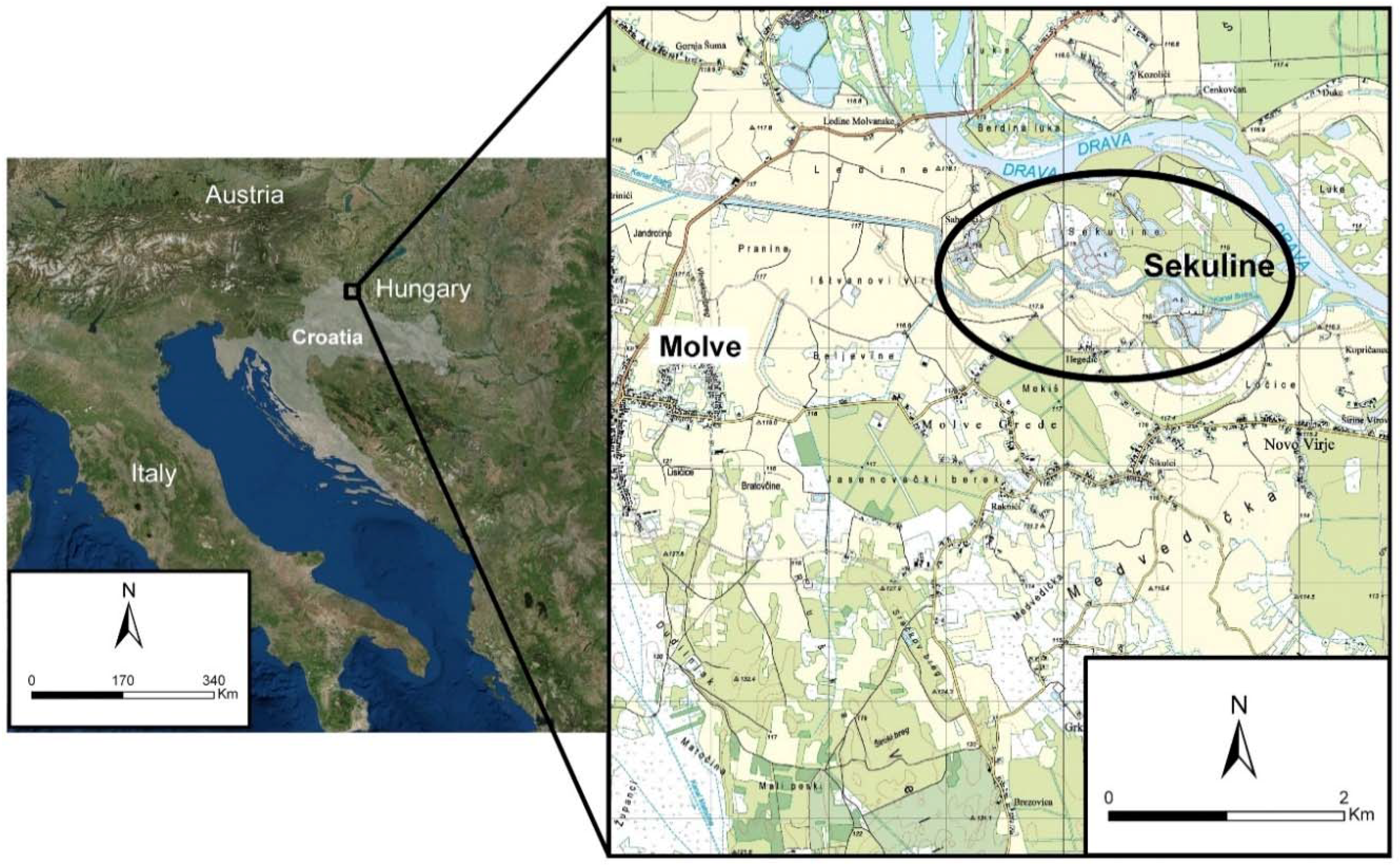
2. Material and Methods
3. Results
3.1. Order Proboscidea Illiger, 1811; Family Elephantidae Gray, 1821; Genus Mammuthus Brookes, 1828
Mammuthus primigenius (Blumenbach, 1799)
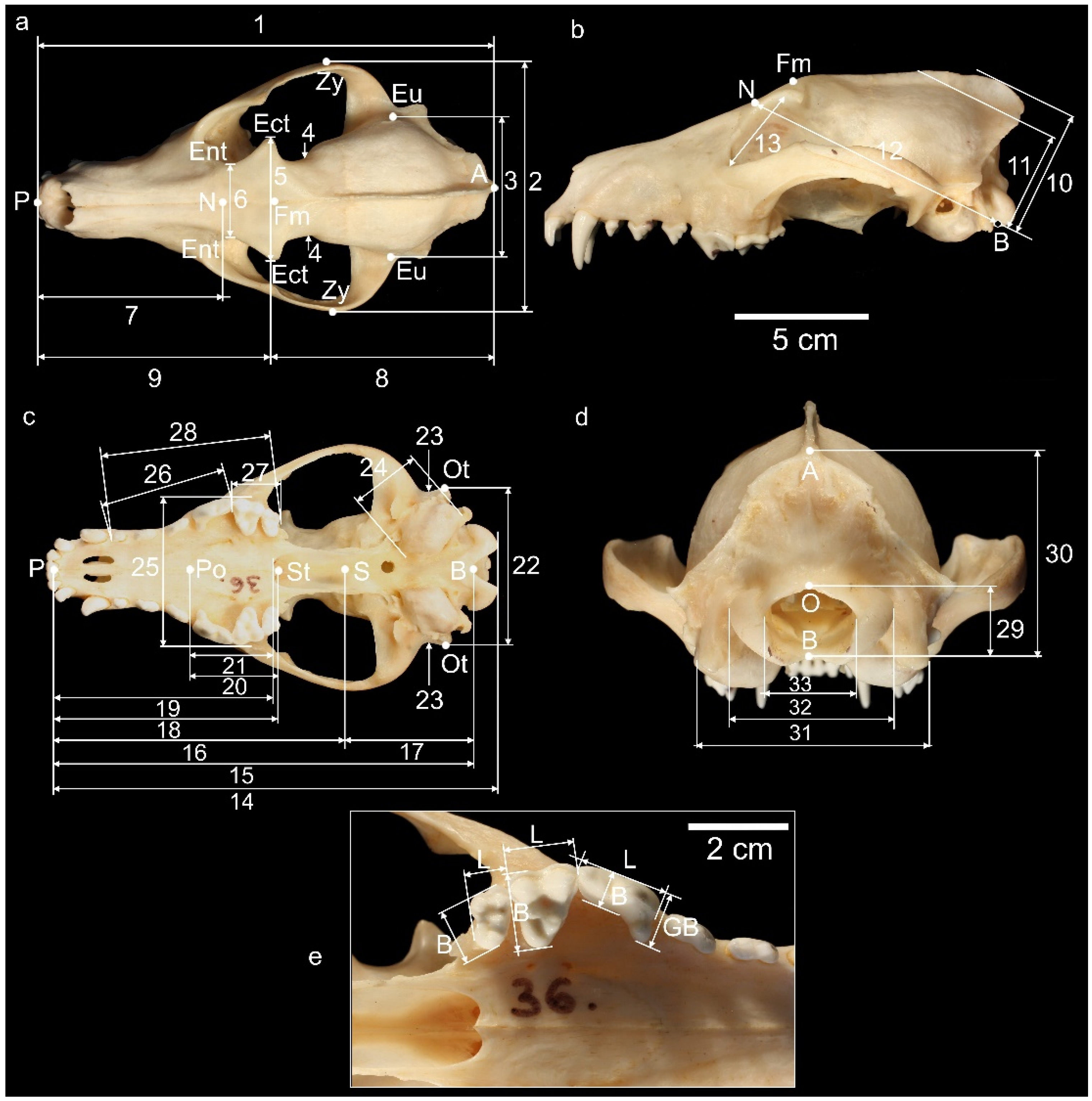
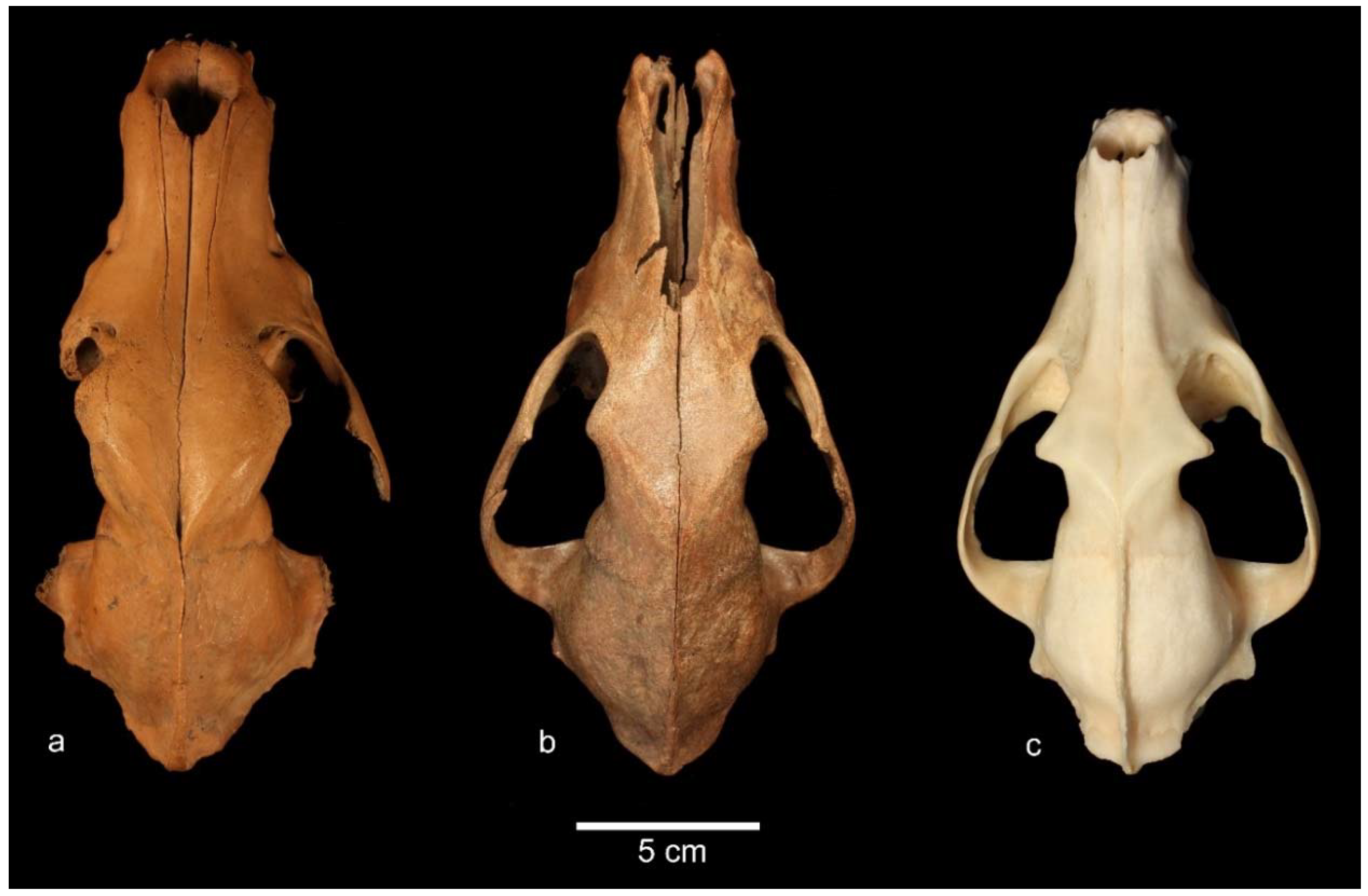
3.2. Order Carnivora Bowdich (1821); Family Canidae Fischer von Waldheim, 1817; Genus Canis Linnaeus, 1758
Canis familiaris Linnaeus, 1758
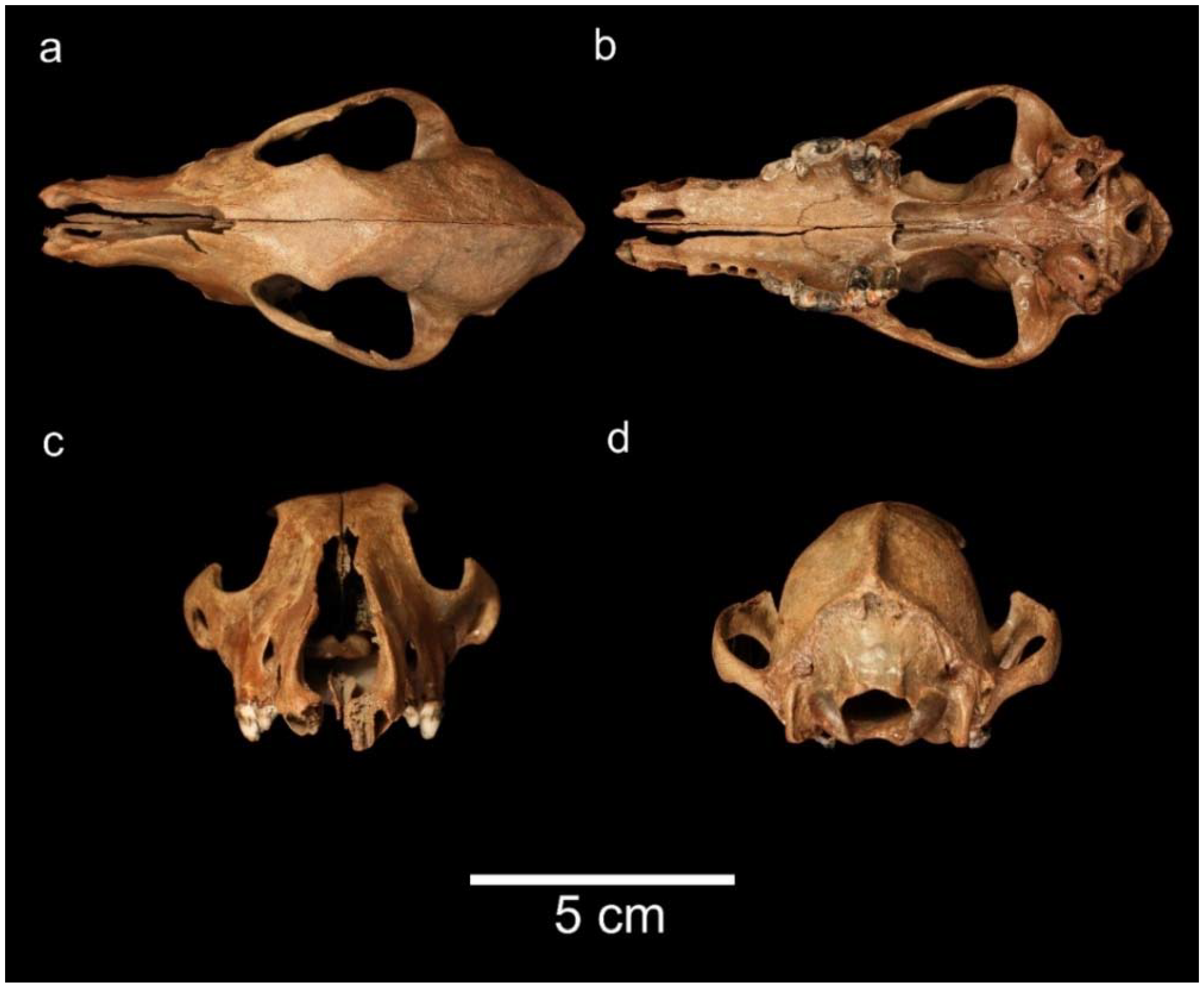
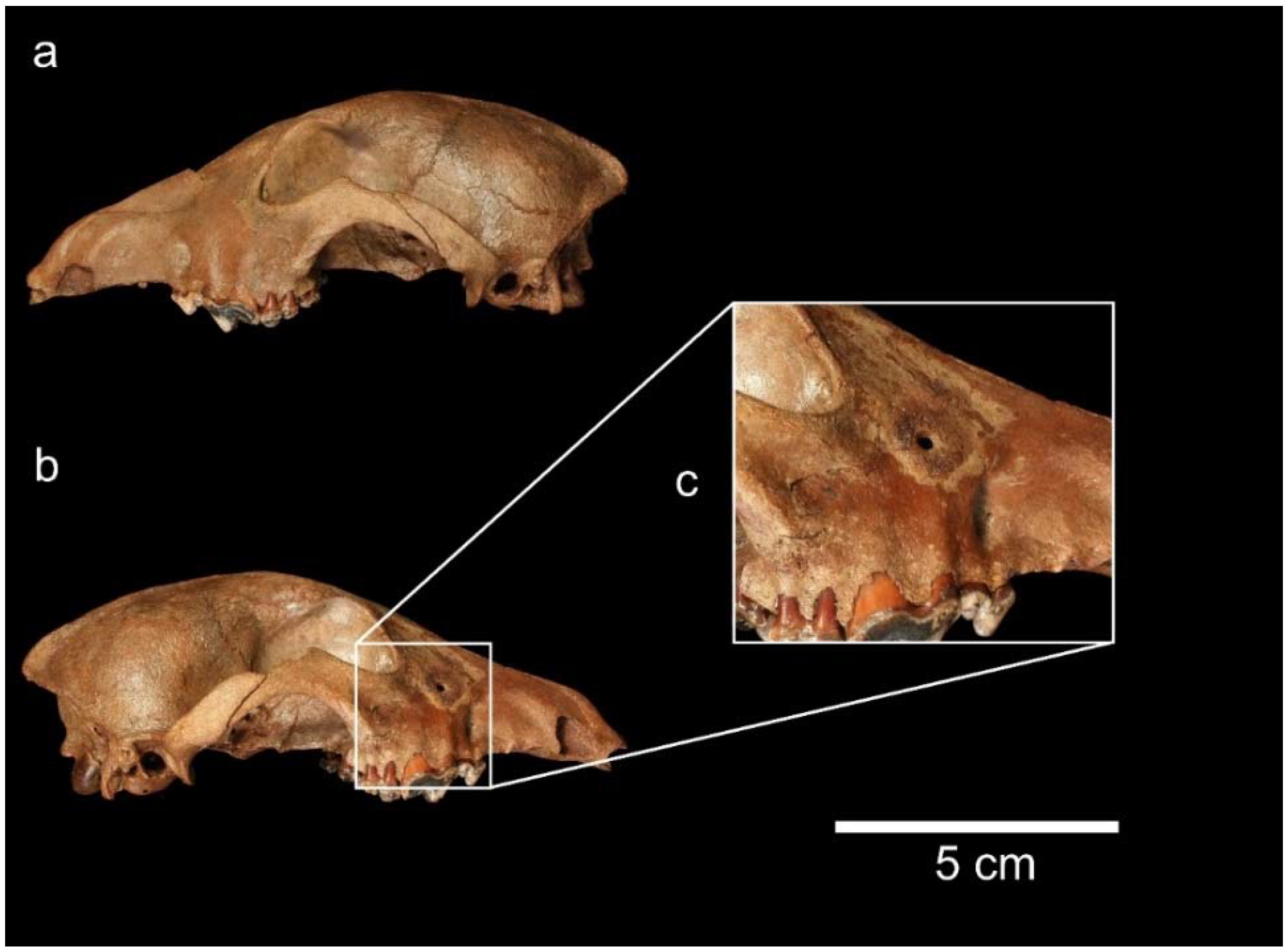
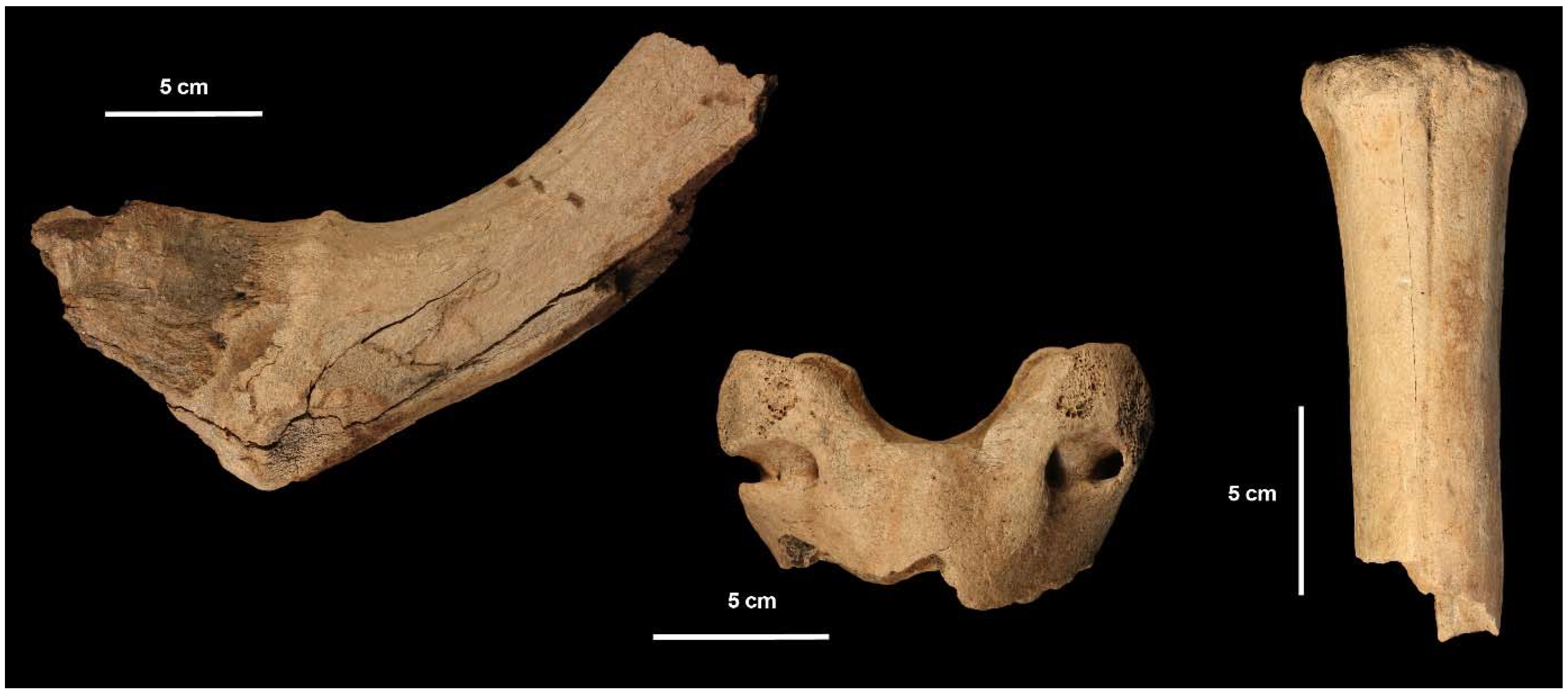
3.3. Order Artiodactyla Owen, 1848; Family Cervidae Goldfuss, 1820
3.3.1. Genus Megaloceros Brookes, 1828; Megaloceros giganteus (Blumenbach, 1799)
3.3.2. Genus Cervus Linnaeus, 1758; Cervus elaphus Linnaeus, 1758
4. Discussion
4.1. Mammuthus primigenius
4.2. Canis familiaris
4.3. Megaloceros giganteus
4.4. Cervus elaphus
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Panonska nizina. Hrvatska Enciklopedija; Leksikografski zavod Miroslav Krleža. Available online: http://www.enciklopedija.hr/natuknica.aspx?id=46451 (accessed on 20 July 2018).
- Malez, M.; Brajković, D. Gornjopleistocenski sisavci u naplavinama rijeke Drave (Hrvatska) [Upper-Pleistocene mammals in alluvial deposits of the river Drava (Croatia)]. Geol. Vjesn. 1991, 44, 31–37, (In Croatian with English Summary). [Google Scholar]
- Winkler, E.M.; Paunović, M. Ein menschlicher Unterkiefer aus den jungpleistozänen Ablagerungen der Drau bei Šašnato polje, Kroatien. Mitt. Anthropol. Ges. Wien 1992, 122, 109–153. [Google Scholar]
- Mauch Lenardić, J. Miocene to Late Pleistocene proboscideans of Croatia. Quat. Int. 2012, 276–277, 120–128. [Google Scholar] [CrossRef]
- Mauch Lenardić, J.; Oros Sršen, A.; Radović, S. Quaternary fauna of the Eastern Adriatic (Croatia) with the special review on the Late Pleistocene sites. Quat. Int. 2017, in press. [Google Scholar] [CrossRef]
- Von den Driesch, A. A guide to the measurement of animal bones from archaeological sites. Bull. Peabody Mus. Nat. Hist. 1976, 1, 1–137. [Google Scholar]
- Zinoviev, A.V. Study of the Medieval Dogs from Novgorod, Russia (X–XIV Century). Int. J. Osteoarchaeol. 2012, 22, 145–157. [Google Scholar] [CrossRef]
- Mihelić, D.; Pavić, M.; Hincak, Z. A Craniometrical Analysis of the Early Bronze Age Dogs from Vučedol Site (East Slavonia, Croatia). Coll. Antropol. 2013, 37, 239–244. [Google Scholar] [PubMed]
- Horvatinčić, N.; Barešić, J.; Krajcar Bronić, I.; Obelić, B. Measurement of low 14C activitie in a liquid scintillation counter in the Zagreb Radiocarbon Laboratory. Radiocarbon 2004, 46, 105–116. [Google Scholar] [CrossRef]
- Naeini, A.T.; Jahromi, A.R.; Mehrshad, S. Bilateral abscesses of the maxillary carnassial teeth in a female Pekinese. Turk. J. Vet. Anim. Sci. 2010, 34, 461–464. [Google Scholar] [CrossRef]
- Reiter, A.M. Endodontic Disease in Small Animals. Available online: https://www.msdvetmanual.com/digestive-system/dentistry/endodontic-disease-in-small-animals (accessed on 9 April 2018).
- The Human Society of the United States. How to Determine a Cat’s or Dog’s Age. 1996. Available online: http://www.ruralareavet.org/PDF/Physical_Exam-How_to_Determine_Age.pdf (accessed on 23 May 2018).
- Lyman, R.L. Vertebrate Taphonomy. Cambridge Manuals in Archaeology; Cambridge University Press: Cambridge, UK, 1999; pp. 1–524. ISBN 0-521-45840-4. [Google Scholar]
- Schmid, E. Atlas of Animal Bones. For Prehistorians, Archaeologists and Quaternary Geologists; Elsevier Publishing Company: Amsterdam, The Netherlands, 1972; pp. 1–159. ISBN 0-444-40831-2. [Google Scholar]
- Brown, W.A.B.; Chapman, N.G. The dentition of red deer (Cervus elaphus): A scoring scheme to assess age from wear of the permanent molariform teeth. J. Zool. 1991, 224, 519–536. [Google Scholar] [CrossRef]
- Reitz, E.J.; Wing, E.S. Zooarchaeology. Cambridge Manuals in Archaeology; Cambridge University Press: Cambridge, UK, 1999; pp. 1–455. ISBN 0-521-48529-0. [Google Scholar]
- Markova, A.K.; Puzachenko, A.Y.; van Kolfschoten, T.; van der Plicht, J.; Ponomarev, D.V. New data on changes in the European distribution of the mammoth and the woolly rhinoceros during the second half of the Late Pleistocene and the early Holocene. Quat. Int. 2013, 292, 4–14. [Google Scholar] [CrossRef]
- Stuart, A.J.; Sulerzhitsky, L.D.; Orlova, L.A.; Kuzmin, Y.V.; Lister, A.M. The latest woolly mammoths (Mammuthus primigenius Blumenbach) in Europe and Asia: a review of the current evidence. Quat. Sci. Rev. 2002, 21, 1559–1569. [Google Scholar] [CrossRef]
- Konrád, G.; Kovács, J.; Halász, A.; Sebe, K.; Pálffy, H. Late Quaternary woolly mammoth (Mammuthus primigenius Blum) remains from southern Transdanubia, Hungary. C. R. Palevol. 2010, 9, 47–54. [Google Scholar] [CrossRef]
- Kovács, J. Radiocarbon chronology of Late Pleistocene large mammal faunas from the Pannonian basin (Hungary). Bull. Geosci. 2011, 87, 13–19. [Google Scholar] [CrossRef]
- Thalmann, O.; Shapiro, B.; Cui, P.; Schuenemann, V.J.; Sawyer, S.K.; Greenfield, D.L.; Germonpré, M.B.; Sablin, M.V.; López-Giráldez, F.; Domingo-Roura, X.; et al. Complete Mitochondrial Genomes of Ancient Canids Suggest a European Origin of Domestic Dogs. Science 2013, 342, 871–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germonpré, M.; Fedorov, S.; Danilov, P.; Galeta, P.; Jimenez, E.-L.; Sablin, M.; Losey, R.J. Palaeolithic and prehistoric dogs and Pleistocene wolves from Yakutia: Identification of isolated skulls. J. Archaeol. Sci. 2017, 78, 1–19. [Google Scholar] [CrossRef]
- Germonpré, M.; Sablin, M.V.; Stevens, R.E.; Hedges, R.E.M.; Hofreiter, M.; Stiller, M.; Després, V.R. Fossil dogs and wolves from Palaeolithic sites in Belgium, the Ukraine and Russia: Osteometry, ancient DNA and stable isotopes. J. Archaeol. Sci. 2009, 36, 473–490. [Google Scholar] [CrossRef]
- Kurtén, B. Pleistocene Mammals of Europe; Library of Congress Catalog Card Number 68-29953; Aldine Publishing Company: Chicago, IL, USA; G. Weidenfeld & Nicolson Ltd.: London, UK, 1968; pp. 1–317. [Google Scholar]
- Valli, A.M.F. Late Pleistocene Deer in the Region of the National Park “Serra da Capivara” (Piauí, Brazil). Quaternary 2018, 1, 4. [Google Scholar] [CrossRef]
- Kahlke, R.-D. The History of the Origins, Evolution and Dispersal of the Late Pleistocene Mammuthus-Coelodonta Faunal Complex in Eurasia (Large Mammals); Fenske Companies: Rapid City, SD, USA, 1999; pp. 1–219. ISBN 0-913062-04-9. [Google Scholar]
- Lister, A.M.; Edwards, C.J.; Nock, D.A.W.; Bunce, M.; van Pijlen, I.A.; Bradley, D.G.; Thomas, M.G.; Barnes, I. The phylogenetic position of the “giant deer” Megaloceros giganteus. Nature 2005, 438, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Malez, M.; Rukavina, D. Položaj naslaga spilje Vindije u sustavu članjenja kvartara šireg područja Alpa (Lage der Höhlenablagerungen von Vindija im System der quartären Vollgliederung des Alpengebietes). Rad JAZU 1979, 383, 187–218, (In Croatian with German summary). [Google Scholar]
- Miracle, P.T.; Mauch Lenardić, J.; Brajković, D. Last glacial climates, “Refugia”, and faunal change in Southeastern Europe: Mammalian assemblages from Veternica, Velika pećina, and Vindija caves (Croatia). Quat. Int. 2010, 212, 137–148. [Google Scholar] [CrossRef]
- Malez, M. Hijenske pećine u doba Pleistocena na području Hrvatske (Hyänenhöhlen aus dem Pleistozän in Kroatien). Rad JAZU 1975, 371, 307–316, (In Croatian with German summary). [Google Scholar]
- Aulagnier, S.; Haffner, P.; Mitchell-Jones, A.J.; Moutou, F.; Zima, J. Mammals of Europe, North Africa and the Middle East; A&C Black Publishers Ltd.: London, UK, 2008; pp. 1–272. ISBN 978-1-4081-1399-8. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Locality | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| Canis familiaris | Sekuline | 194.6 | 98.5 | 54.0 | 34.8 | 45.2 | 35.5 | 99.0 | 94.0 | 117.4 | 55.0 |
| Canis familiaris | Vučedol | 151–191 | 94–104 | 52–54 | - | 49–55 | 35–38 | 75–99 | 73–91 | 83–110 | - |
| Canis aureus | Slavonia | 177.3 | 95.9 | 55.4 | 28.5 | 45.0 | 27.3 | 85.4 | 83.5 | 100.0 | 52.5 |
| Canis lupus (♀) | Lika | 213.3 | 113.5 | 66.5 | 40.5 | 53.1 | 40.0 | 106 | 98.0 | 123.0 | 61.8 |
| Taxa | Locality | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 |
| Canis familiaris | Sekuline | 48.0 | 101.8 | 28.3 | 177.7 | 170.5 | 125.4 | 47.1 | 94.9 | 93.0 | 33.2 |
| Canis familiaris | Vučedol | - | 80–101 | 26–30 | 148–186 | 139–174 | - | - | - | - | - |
| Canis aureus | Slavonia | 48.0 | 92.5 | 30.0 | 166.0 | 155.0 | 111.0 | 46.2 | 83.2 | 81.8 | 33.0 |
| Canis lupus (♀) | Lika | 56.5 | 110.0 | 33.0 | 204.5 | 194.0 | 141.0 | 53.3 | 104.5 | 103.8 | 37.1 |
| Taxa | Locality | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 |
| Canis familiaris | Sekuline | 31.3 | 60.8 | 61.0 | 20.0 | 57.9 | 50.5 | 20.0 | 67.5 | 15.1 | 43.0 |
| Canis familiaris | Vučedol | - | 56–69 | - | - | - | - | - | - | 14–16 | 37–40 |
| Canis aureus | Slavonia | 31.6 | 60.6 | 58.5 | 25.9 | 55.2 | 46.8 | 19.6 | 62.5 | 14.5 | 40.0 |
| Canis lupus (♀) | Lika | 36.4 | 73.5 | 71.0 | 28.0 | 71.6 | 56.0 | 21.5 | 73.0 | 17.0 | 51.0 |
| Taxa | Locality | 31 | 32 | 33 | |||||||
| Canis familiaris | Sekuline | 50.2 | 35.5 | 18.5 | |||||||
| Canis familiaris | Vučedol | 45–53 | 34–37 | 18–19 | |||||||
| Canis aureus | Slavonia | 47.5 | 33.2 | 18.4 | |||||||
| Canis lupus (♀) | Lika | 60.0 | 44.0 | 22.2 | |||||||
| Taxa | Locality | GB (P4) | B (P4) | B (M1) | B (M2) | L (P4) | L (M1) | L (M2) | |||
| Canis familiaris | Sekuline | 11.5 | 8.7 | 14.6 | 10.3 | 19.5 | 12.5 | 8.0 | |||
| Canis aureus | Slavonia | 9.4 | 6.4 | 13.5 | 9.6 | 17.0 | 12.2 | 7.5 | |||
| Canis lupus (♀) | Lika | 12.5 | 9.8 | 17.5 | 10.8 | 22.5 | 15.0 | 8.5 |
| Locality | Skull Index 2/1 × 100 | Cranial Index 3/NA × 100 | Facial Index 2/7 × 100 | Foramen Magnum Index 29/33 × 100 | Length–Length Index 2 NA/7 | Ratio of Cranium to Facial Bones NA/7 × 100 |
|---|---|---|---|---|---|---|
| Sekuline | 50.62 | 49.09 | 99.49 | 81.62 | 1.11 | 111.11 |
| Vučedol | 54.25–62.25 | 51.49–67.50 | 105.05–125.33 | 73.68–88.89 | 1.02–1.08 | 102.02–108.23 |
| Measurements (occlusal)/Teeth | P3 | P4 | M1 | M2 | M3 |
|---|---|---|---|---|---|
| L | 18.4 | 17.0 | 21.8 | 26.4 | 27.7 |
| B | 15.0 | 17.2 | 19.4 | 22.4 | 21.7 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mauch Lenardić, J.; Radović, S.; Oros Sršen, A.; Horvatinčić, N.; Kostešić, P.; Bermanec, V. Mammoths, Deer, and a Dog: Fossil and (Sub) Recent Allochthonous Remains from the Northeastern Croatia (Podravina Region), with the First Radiocarbon Dating of the Croatian Woolly Mammoths (Mammuthus primigenius). Quaternary 2018, 1, 11. https://doi.org/10.3390/quat1020011
Mauch Lenardić J, Radović S, Oros Sršen A, Horvatinčić N, Kostešić P, Bermanec V. Mammoths, Deer, and a Dog: Fossil and (Sub) Recent Allochthonous Remains from the Northeastern Croatia (Podravina Region), with the First Radiocarbon Dating of the Croatian Woolly Mammoths (Mammuthus primigenius). Quaternary. 2018; 1(2):11. https://doi.org/10.3390/quat1020011
Chicago/Turabian StyleMauch Lenardić, Jadranka, Siniša Radović, Ankica Oros Sršen, Nada Horvatinčić, Petar Kostešić, and Vladimir Bermanec. 2018. "Mammoths, Deer, and a Dog: Fossil and (Sub) Recent Allochthonous Remains from the Northeastern Croatia (Podravina Region), with the First Radiocarbon Dating of the Croatian Woolly Mammoths (Mammuthus primigenius)" Quaternary 1, no. 2: 11. https://doi.org/10.3390/quat1020011