
Epidemiological, Virulence, and Antibiotic Resistance Analysis of Klebsiella pneumoniae, a Major Source of Threat to Livestock and Poultry in Some Regions of Xinjiang, China
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation and Identification of K. pneumoniae
2.3. Capsular Polysaccharide Characterization and Multilocus Sequence Typing
2.4. Virulence Gene Detection
2.5. Antibiotic Resistance Gene Detection
2.6. Antibiotic Susceptibility Test
2.7. Statistical Analysis
3. Results
3.1. Prevalence of K. pneumoniae in Different Animal Farms
3.2. Capsular Serotyping and Multilocus Sequence Typing
3.3. Virulence Gene Detection
3.4. Antibiotic Susceptibility Testing
3.5. Antibiotic Resistance Genes
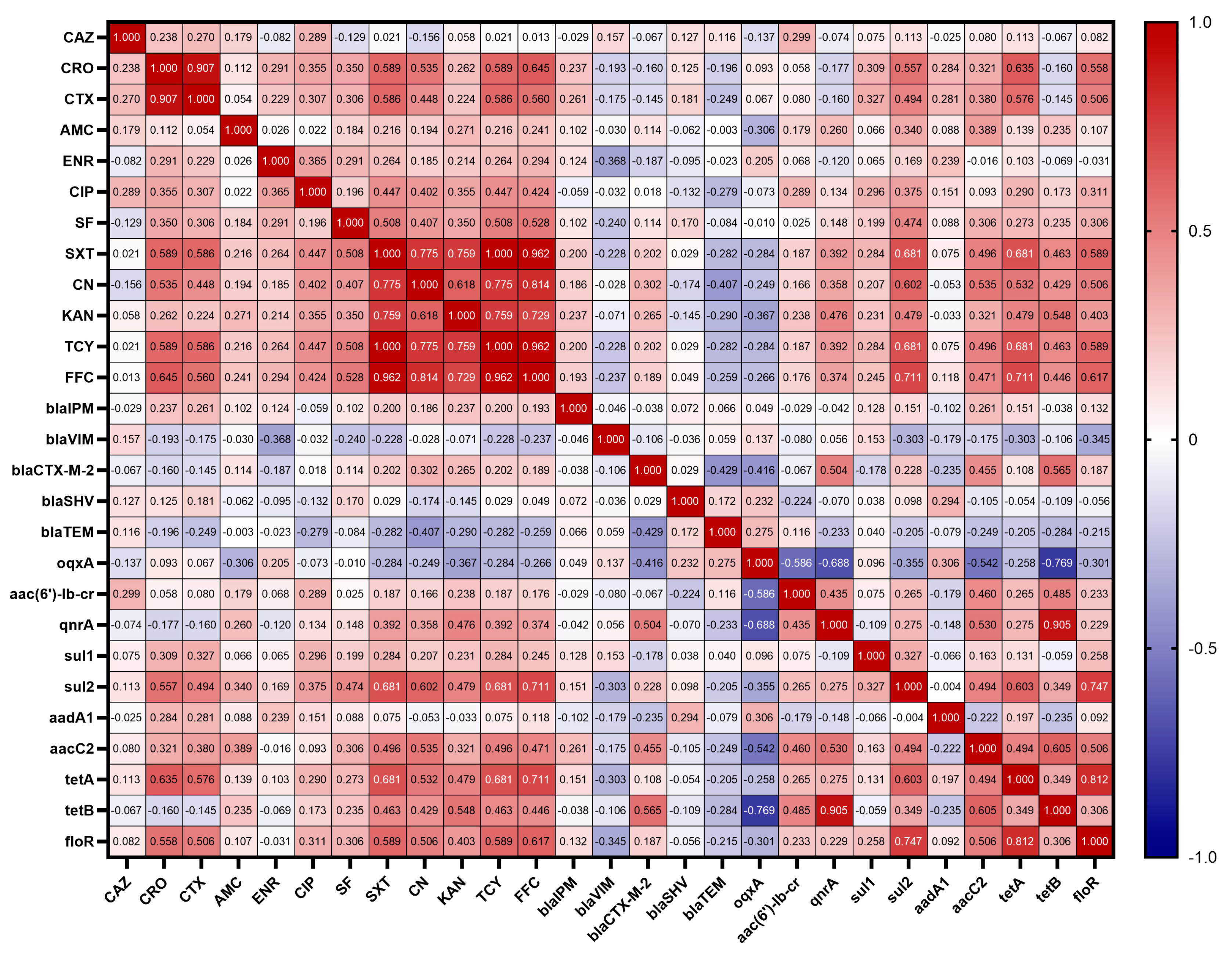
3.6. Correlations between Antibiotic Resistant Phenotypes and Resistant Genes
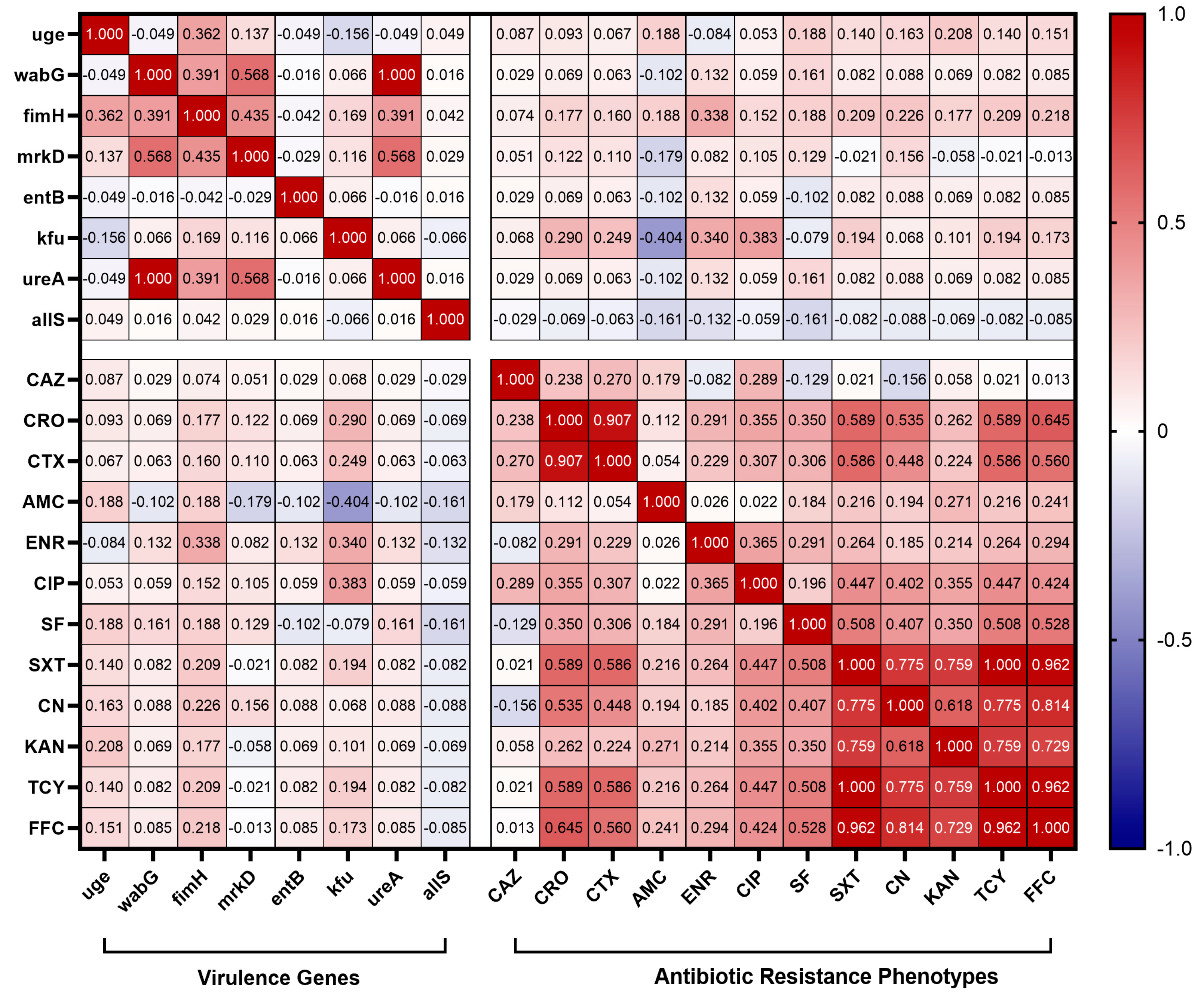
3.7. Correlation Analysis of Virulence and Antibiotic Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef] [PubMed]
- Michael, S.A.; Hayman, D.; Gray, R.; Roe, W.D. Clinical parameters of hypervirulent Klebsiella pneumoniae disease and ivermectin treatment in New Zealand sea lion (Phocarctos hookeri) pups. PLoS ONE 2022, 17, e0264582. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Zhang, J.; Han, B.; Barkema, H.W.; Cobo, E.R.; Kastelic, J.P.; Zhou, M.; Shi, Y.; Wang, J.; Yang, R.; et al. Klebsiella pneumoniae isolated from bovine mastitis is cytopathogenic for bovine mammary epithelial cells. J. Dairy. Sci. 2020, 103, 3493–3504. [Google Scholar] [CrossRef] [PubMed]
- Mahrous, S.H.; El-Balkemy, F.A.; Abo-Zeid, N.Z.; El-Mekkawy, M.F.; El Damaty, H.M.; Elsohaby, I. Antibacterial and Anti-Biofilm Activities of Cinnamon Oil against Multidrug-Resistant Klebsiella pneumoniae Isolated from Pneumonic Sheep and Goats. Pathogens 2023, 12, 1138. [Google Scholar] [CrossRef]
- Mobasseri, G.; The, C.; Ooi, P.T.; Thong, K.L. The emergence of colistin-resistant Klebsiella pneumoniae strains from swine in Malaysia. J. Glob. Antimicrob. Resist. 2019, 17, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xin, L.; Peng, C.; Liu, C.; Wang, P.; Yu, L.; Liu, M.; Wang, F. Prevalence and antimicrobial susceptibility profiles of ESBL-producing Klebsiella Pneumoniae from broiler chicken farms in Shandong Province, China. Poult. Sci. 2022, 101, 102002. [Google Scholar] [CrossRef] [PubMed]
- Shon, A.S.; Bajwa, R.P.; Russo, T.A. Hypervirulent (hypermucoviscous) Klebsiella pneumoniae: A new and dangerous breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the Offense with a Strong Defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.J.; Lin, T.L.; Chen, Y.H.; Hsu, C.R.; Hsieh, P.F.; Wu, M.C.; Wang, J.T. Capsular types of Klebsiella pneumoniae revisited by wzc sequencing. PLoS ONE 2013, 8, e80670. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sun, G.; Yu, Y.; Li, N.; Chen, M.; Jin, R.; Jiao, Y.; Wu, H. Increasing occurrence of antimicrobial-resistant hypervirulent (hypermucoviscous) Klebsiella pneumoniae isolates in China. Clin. Infect. Dis. 2014, 58, 225–232. [Google Scholar] [CrossRef]
- Russo, T.A.; Shon, A.S.; Beanan, J.M.; Olson, R.; MacDonald, U.; Pomakov, A.O.; Visitacion, M.P. Hypervirulent K. pneumoniae secretes more and more active iron-acquisition molecules than “classical” K. pneumoniae thereby enhancing its virulence. PLoS ONE 2011, 6, e26734. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, G.; Chao, X.; Xie, L.; Wang, H. The Characteristic of Virulence, Biofilm and Antibiotic Resistance of Klebsiella pneumoniae. Int. J. Environ. Res. Public Health 2020, 17, 6278. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, Y.; Zhu, Y.; Jia, P.; Li, X.; Jia, X.; Yu, W.; Cui, Y.; Yang, R.; Xia, W.; et al. Emergence of colistin-resistant hypervirulent Klebsiella pneumoniae (CoR-HvKp) in China. Emerg. Microbes Infect. 2022, 11, 648–661. [Google Scholar] [CrossRef] [PubMed]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef]
- Guo, Y.; Zhou, H.; Qin, L.; Pang, Z.; Qin, T.; Ren, H.; Pan, Z.; Zhou, J. Frequency, Antimicrobial Resistance and Genetic Diversity of Klebsiella pneumoniae in Food Samples. PLoS ONE 2016, 11, e0153561. [Google Scholar] [CrossRef]
- Liao, W.; Liu, Y.; Zhang, W. Virulence evolution, molecular mechanisms of resistance and prevalence of ST11 carbapenem-resistant Klebsiella pneumoniae in China: A review over the last 10 years. J. Glob. Antimicrob. Resist. 2020, 23, 174–180. [Google Scholar] [CrossRef]
- Yang, Y.; Peng, Y.; Jiang, J.; Gong, Z.; Zhu, H.; Wang, K.; Zhou, Q.; Tian, Y.; Qin, A.; Yang, Z.; et al. Isolation and characterization of multidrug-resistant Klebsiella pneumoniae from raw cow milk in Jiangsu and Shandong provinces, China. Transbound. Emerg. Dis. 2021, 68, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Xue, J.Z.; Li, S.S.; Elken, E.M.; Haqmal, M.A.; Li, X.S.; Xu, G.Y.; Kong, L.C.; Ma, H.X. Analysis of an IncR plasmid carried by carbapenem-resistant Klebsiella pneumoniae: A survey of swine Klebsiella pneumoniae in Jilin Province. J. Glob. Antimicrob. Resist. 2023, 34, 83–90. [Google Scholar] [CrossRef]
- Wang, X.; Sun, N.; Liu, X.; Li, F.; Sun, J.; Huang, J.; Li, R.; Wang, L. Small clone dissemination of tmexCD1-toprJ1-carrying Klebsiella pneumoniae isolates in a chicken farm. J. Glob. Antimicrob. Resist. 2022, 29, 105–112. [Google Scholar] [CrossRef]
- Wenz, J.R.; Barrington, G.M.; Garry, F.B.; Dinsmore, R.P.; Callan, R.J. Use of systemic disease signs to assess disease severity in dairy cows with acute coliform mastitis. J. Am. Vet. Med. Assoc. 2001, 218, 567–572. [Google Scholar] [CrossRef]
- NY/T 2692-2015; Rapid Diagnostic Techniques for Subclinical Mastitis in Dairy Cow. National Standards of the People’s Republic of China: Beijing, China, 2015.
- Neuberger, A.; Oren, I.; Sprecher, H. Clinical impact of a PCR assay for rapid identification of Klebsiella pneumoniae in blood cultures. J. Clin. Microbiol. 2008, 46, 377–379. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Guo, X.; Xiang, S.; Li, J.; Li, X.; Xiang, H.; He, J.; Chen, D.; Chen, J. Comparative analyses of phenotypic methods and 16S rRNA, khe, rpoB genes sequencing for identification of clinical isolates of Klebsiella pneumoniae. Antonie Van Leeuwenhoek 2016, 109, 1029–1040. [Google Scholar] [CrossRef]
- Brisse, S.; Passet, V.; Haugaard, A.B.; Babosan, A.; Kassis-Chikhani, N.; Struve, C.; Decré, D. wzi Gene sequencing, a rapid method for determination of capsular type for Klebsiella strains. J. Clin. Microbiol. 2013, 51, 4073–4078. [Google Scholar] [CrossRef]
- Alcántar-Curiel, M.D.; Blackburn, D.; Saldaña, Z.; Gayosso-Vázquez, C.; Iovine, N.M.; De la Cruz, M.A.; Girón, J.A. Multi-functional analysis of Klebsiella pneumoniae fimbrial types in adherence and biofilm formation. Virulence 2013, 4, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Russo, T.A.; Olson, R.; Fang, C.T.; Stoesser, N.; Miller, M.; MacDonald, U.; Hutson, A.; Barker, J.H.; La Hoz, R.M.; Johnson, J.R. Identification of Biomarkers for Differentiation of Hypervirulent Klebsiella pneumoniae from Classical K. pneumoniae. J. Clin. Microbiol. 2018, 56, e00776-18. [Google Scholar] [CrossRef]
- Wu, X.; Liu, J.; Feng, J.; Shabbir, M.A.B.; Feng, Y.; Guo, R.; Zhou, M.; Hou, S.; Wang, G.; Hao, H.; et al. Epidemiology, Environmental Risks, Virulence, and Resistance Determinants of Klebsiella pneumoniae From Dairy Cows in Hubei, China. Front. Microbiol. 2022, 13, 858799. [Google Scholar] [CrossRef]
- Candan, E.D.; Aksoz, N. Klebsiella pneumoniae: Characteristics of carbapenem resistance and virulence factors. Acta Biochim. Pol. 2015, 62, 867–874. [Google Scholar] [CrossRef]
- Fatemeh, F.; Abdollah, J.; Babak, K. Phenotypic and genotypic study on antimicrobial resistance patterns of E. coli isolates from bovine mastitis. Microb. Pathog. 2019, 132, 355–361. [Google Scholar]
- M100-S21; Performance Standards for Antimicrobial Susceptibility Testing, 21st ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011.
- M100; Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022.
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Schukken, Y.; Chuff, M.; Moroni, P.; Gurjar, A.; Santisteban, C.; Welcome, F.; Zadoks, R. The “other” gram-negative bacteria in mastitis: Klebsiella, serratia, and more. Vet. Clin. N. Am. Food Anim. Pract. 2012, 28, 239–256. [Google Scholar] [CrossRef]
- Patel, S.; Chauhan, H.; Patel, A.; Shrimali, M.; Patel, K.; Prajapati, B.; Kala, J.; Patel, M.; Rajgor, M.; Patel, M. Isolation and identification of Klebsiella pneumoniae from sheep-case report. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 331–334. [Google Scholar] [CrossRef]
- Fouad, E.A.; Khalaf, D.D.; Farahat, E.; Hakim, A.S. Identification of predominant pathogenic bacteria isolated from respiratory manifested small ruminants in western north Egypt with regard to their susceptibility to antibiotics. Int. J. Health Sci. 2022, 6, 10818–10828. [Google Scholar] [CrossRef]
- Franklin-Alming, F.V.; Kaspersen, H.; Hetland, M.A.K.; Bakksjø, R.J.; Nesse, L.L.; Leangapichart, T.; Löhr, I.H.; Telke, A.A.; Sunde, M. Exploring Klebsiella pneumoniae in Healthy Poultry Reveals High Genetic Diversity, Good Biofilm-Forming Abilities and Higher Prevalence in Turkeys Than Broilers. Front. Microbiol. 2021, 12, 725414. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Zhou, M.; Nobrega, D.B.; Barkema, H.W.; Xu, S.; Li, M.; Kastelic, J.P.; Shi, Y.; Han, B.; Gao, J. Genetic diversity and molecular epidemiology of outbreaks of Klebsiella pneumoniae mastitis on two large Chinese dairy farms. J. Dairy Sci. 2021, 104, 762–775. [Google Scholar] [CrossRef]
- Wareth, G.; Neubauer, H. The Animal-foods-environment interface of Klebsiella pneumoniae in Germany: An observational study on pathogenicity, resistance development and the current situation. Vet. Res. 2021, 52, 16. [Google Scholar] [CrossRef]
- Lepuschitz, S.; Hauser, K.; Schriebl, A.; Schlagenhaufen, C.; Stöger, A.; Chakeri, A.; Vötsch, K.; Pekard-Amenitsch, S.; Springer, B.; Allerberger, F.; et al. Fecal Klebsiella pneumoniae Carriage Is Intermittent and of High Clonal Diversity. Front. Microbiol. 2020, 11, 581081. [Google Scholar] [CrossRef]
- Fu, S.; Wen, C.; Wang, Z.; Qiu, Y.; Zhang, Y.; Zuo, J.; Xu, Y.; Han, X.; Luo, Z.; Chen, W.; et al. Molecular Epidemiology and Antimicrobial Resistance of Outbreaks of Klebsiella pneumoniae Clinical Mastitis in Chinese Dairy Farms. Microbiol. Spectr. 2022, 10, e0299722. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Deng, B.; Liao, W.; Wang, P.; Chen, P.; Wei, J. High rate of multiresistant Klebsiella pneumoniae from human and animal origin. Infect. Drug Resist. 2019, 12, 2729–2737. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Wu, X.; Cao, H.; Pei, T.; Zhou, Y.; Yang, Y.; Yang, Z. The Characteristics of Multilocus Sequence Typing, Virulence Genes and Drug Resistance of Klebsiella pneumoniae Isolated from Cattle in Northern Jiangsu, China. Animals 2022, 12, 2627. [Google Scholar] [CrossRef]
- Turton, J.; Davies, F.; Turton, J.; Perry, C.; Payne, Z.; Pike, R. Hybrid Resistance and Virulence Plasmids in “High-Risk” Clones of Klebsiella pneumoniae, Including Those Carrying blaNDM-5. Microorganisms 2019, 7, 326. [Google Scholar] [CrossRef]
- Yang, X.; Dong, N.; Chan, E.W.; Zhang, R.; Chen, S. Carbapenem Resistance-Encoding and Virulence-Encoding Conjugative Plasmids in Klebsiella pneumoniae. Trends Microbiol. 2021, 29, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Salauddin, M.; Akter, M.R.; Hossain, M.K.; Rahman, M.M. Isolation of multi-drug resistant Klebsiella sp. from bovine mastitis samples in Rangpur, Bangladesh. J. Adv. Vet. Anim. Res. 2019, 6, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Zhao, T.; Liu, Q.; He, J.; He, D.; Wu, G.; Li, Y.; Jiang, C.; Xu, Z. Residual veterinary antibiotics in pig excreta after oral administration of sulfonamides. Environ. Geochem. Health 2016, 38, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.S.; Li, Y.Z.; Ge, J.; Xiao, G.; Mo, Y.; Wen, Y.Q.; Liu, J.P.; Xiong, Y.Q.; Qiu, M.; Huo, S.T.; et al. Comparisons of microbiological characteristics and antibiotic resistance of Klebsiella pneumoniae isolates from urban rodents, shrews, and healthy people. BMC Microbiol. 2020, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Phetburom, N.; Boueroy, P.; Chopjitt, P.; Hatrongjit, R.; Akeda, Y.; Hamada, S.; Nuanualsuwan, S.; Kerdsin, A. Klebsiella pneumoniae Complex Harboring mcr-1, mcr-7, and mcr-8 Isolates from Slaughtered Pigs in Thailand. Microorganisms 2021, 9, 2436. [Google Scholar] [CrossRef] [PubMed]
- Sukmawinata, E.; Uemura, R.; Sato, W.; Thu Htun, M.; Sueyoshi, M. Multidrug-Resistant ESBL/AmpC-Producing Klebsiella pneumoniae Isolated from Healthy Thoroughbred Racehorses in Japan. Animals 2020, 10, 369. [Google Scholar] [CrossRef] [PubMed]
- García-Cobos, S.; Köck, R.; Mellmann, A.; Frenzel, J.; Friedrich, A.W.; Rossen, J.W. Molecular Typing of Enterobacteriaceae from Pig Holdings in North-Western Germany Reveals Extended- Spectrum and AmpC β-Lactamases Producing but no Carbapenem Resistant Ones. PLoS ONE 2015, 10, e0134533. [Google Scholar]
- Lee, C.R.; Lee, J.H.; Park, K.S.; Kim, Y.B.; Jeong, B.C.; Lee, S.H. Global Dissemination of Carbapenemase-Producing Klebsiella pneumoniae: Epidemiology, Genetic Context, Treatment Options, and Detection Methods. Front. Microbiol. 2016, 7, 895. [Google Scholar] [CrossRef] [PubMed]
- Conley, Z.C.; Bodine, T.J.; Chou, A.; Zechiedrich, L. Wicked: The untold story of ciprofloxacin. PLoS Pathog. 2018, 14, e1006805. [Google Scholar] [CrossRef]
- Kareem, S.M.; Al-Kadmy, I.M.S.; Kazaal, S.S.; Mohammed Ali, A.N.; Aziz, S.N.; Makharita, R.R.; Algammal, A.M.; Al-Rejaie, S.; Behl, T.; Batiha, G.E.; et al. Detection of gyrA and parC Mutations and Prevalence of Plasmid-Mediated Quinolone Resistance Genes in Klebsiella pneumoniae. Infect. Drug Resist. 2021, 14, 555–563. [Google Scholar] [CrossRef]
- Martínez-Casanova, J.; Gómez-Zorrilla, S.; Prim, N.; Dal Molin, A.; Echeverría-Esnal, D.; Gracia-Arnillas, M.P.; Sendra, E.; Güerri-Fernández, R.; Durán-Jordà, X.; Padilla, E.; et al. Risk Factors for Amoxicillin-Clavulanate Resistance in Community-Onset Urinary Tract Infections Caused by Escherichia coli or Klebsiella pneumoniae: The Role of Prior Exposure to Fluoroquinolones. Antibiotics 2021, 10, 582. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updat. 2010, 13, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Nobrega, D.B.; Calarga, A.P.; Nascimento, L.C.; Chande Vasconcelos, C.G.; de Lima, E.M.; Langoni, H.; Brocchi, M. Molecular characterization of antimicrobial resistance in Klebsiella pneumoniae isolated from Brazilian dairy herds. J. Dairy Sci. 2021, 104, 7210–7224. [Google Scholar] [CrossRef] [PubMed]
- Beutlich, J.; Jahn, S.; Malorny, B.; Hauser, E.; Hühn, S.; Schroeter, A.; Rodicio, M.R.; Appel, B.; Threlfall, J.; Mevius, D.; et al. Antimicrobial resistance and virulence determinants in European Salmonella genomic island 1-positive Salmonella enterica isolates from different origins. Appl. Environ. Microbiol. 2011, 77, 5655–5664. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Farm Types | Total Number of Samples | Types of Samples | Number of Samples |
|---|---|---|---|
| Dairy farm A | 88 | milk from cows with CM | 10 |
| milk from cows with SCM | 13 | ||
| nasal swab of healthy cows | 31 | ||
| anal swab of healthy cows | 5 | ||
| anal swab of calves | 10 | ||
| nasal swab of calves | 10 | ||
| feed | 3 | ||
| bedding | 6 | ||
| Dairy farm B | 41 | nasal swab of healthy cows | 13 |
| anal swab of healthy cows | 10 | ||
| anal swab of calves | 8 | ||
| nasal swab of calves | 8 | ||
| feed | 1 | ||
| bedding | 1 | ||
| Chicken farm | 76 | anal swab of laying hens | 69 |
| environmental samples | 7 | ||
| Sheep farm | 40 | nasal swabs of sheep with respiratory disease | 34 |
| environmental samples | 6 | ||
| Swine farm | 66 | nasal swabs of pig with respiratory disease | 53 |
| environmental samples | 13 |
| Types | Virulence Gene | Positive Rates (%) | No. of Positive Samples/Total (%) from Farm | |||
|---|---|---|---|---|---|---|
| Dairy | Chicken | Sheep | Pig | |||
| regulation | rmpA | 0.00 | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) |
| magA | 0.00 | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) | |
| lipopolysaccharides | uge | 88.71 | 37/43(86.07) | 10/11(90.91) | 5/5(100.00) | 3/3(100.00) |
| wabG | 98.39 | 43/43(100.00) | 11/11(100.00) | 4/5(80.00) | 3/3(100.00) | |
| iron uptake | entB | 98.39 | 42/43(97.67) | 11/11(100.00) | 5/5(100.00) | 3/3(100.00) |
| kfu | 20.97 | 8/43(18.60) | 5/11(45.45) | 0/5(0.00) | 0/3(0.00) | |
| iroN | 0.00 | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) | |
| icuA | 0.00 | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) | |
| fimbriae | fimH | 90.32 | 40/43(93.02) | 10/11(90.91) | 4/5(80.00) | 2/3(66.67) |
| mrkD | 95.16 | 42/43(97.67) | 10/11(90.91) | 4/5(80.00) | 3/3(100.00) | |
| urease | ureA | 98.39 | 43/43(100.00) | 11/11(100.00) | 4/5(80.00) | 3/3(100.00) |
| allantoin | allS | 1.61 | 1/43(2.33) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) |
| Antibiotics | No. of Resistant Samples/Total (%) from Farm | |||
|---|---|---|---|---|
| Dairy | Chicken | Sheep | Pig | |
| Florfenicol | 8/43(18.60) | 8/11(72.73) | 1/5(20.00) | 2/3(66.67) |
| Enrofloxacin | 24/43(55.81) | 7/11(63.64) | 0/5(0.00) | 1/3(33.33) |
| Ciprofloxacin | 5/43(11.63) | 6/11(54.55) | 0/5(0.00) | 0/3(0.00) |
| Ceftazidime | 2/43(4.65) | 0/11(0.00) | 1/5(20.00) | 0/3(0.00) |
| Ceftriaxone | 7/43(16.28) | 5/11(45.45) | 1/5(20.00) | 1/3(33.33) |
| Cefotaxime | 6/43(13.95) | 4/11(36.36) | 1/5(20.00) | 1/3(33.33) |
| Ampicillin | 43/43(100.00) | 11/11(100.00) | 5/5(100.00) | 3/3(100.00) |
| Amoxicillin/clavulanic acid | 26/43(60.47) | 5/11(45.45) | 5/5(100.00) | 2/3(66.67) |
| Sulfisoxazole | 24/43(55.81) | 10/11(90.91) | 1/5(20.00) | 3/3(100.00) |
| Sulfamethoxazole/trimethoprim | 7/43(16.28) | 8/11(72.73) | 1/5(20.00) | 2/3(66.67) |
| Gentamicin | 8/43(18.60) | 8/11(72.73) | 2/5(40.00) | 2/3(66.67) |
| kanamycin | 4/43(9.30) | 6/11(54.55) | 2/5(40.00) | 2/3(66.67) |
| Tetracycline | 7/43(16.28) | 8/11(72.73) | 1/5(20.00) | 2/3(66.67) |
| Imipenem | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 1/3(33.33) |
| Meropenem | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) |
| Types | Resistance Gene | Positive Rates (%) | No. of Positive Samples/Total (%) from Farm | |||
|---|---|---|---|---|---|---|
| Dairy | Chicken | Sheep | Pig | |||
| β-Lactams | blaIMP | 0.00 | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) |
| blaVIM | 11.29 | 4/43(9.30) | 0/11(0.00) | 2/5(40.00) | 1/3(33.33) | |
| blaOXA-48 | 0.00 | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) | |
| blaNDM | 0.00 | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) | |
| blaKPC | 0.00 | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) | |
| blaDHA | 0.00 | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) | |
| blaFOX | 0.00 | 0/43(0.00) | 0/11(0.00) | 0/5(0.00) | 0/3(0.00) | |
| blaCTX-M-2 | 8.06 | 2/43(4.65) | 1/11(9.09) | 1/5(20.00) | 1/3(33.33) | |
| blaSHV | 75.81 | 34/43(79.07) | 7/11(63.63) | 3/5(60.00) | 3/3(100.00) | |
| blaTEM | 79.03 | 39/43(90.70) | 5/11(45.45) | 3/5(60.00) | 2/3(66.67) | |
| Quinolones | oqxA | 87.10 | 42/43(97.67) | 8/11(72.73) | 2/5(40.00) | 2/3(66.67) |
| aac(6′)-Ib-cr | 4.84 | 1/43(2.33) | 1/11(9.09) | 1/5(20.00) | 0/3(0.00) | |
| qnrA | 9.68 | 1/43(2.33) | 2/11(18.18) | 2/5(40.00) | 1/3(33.33) | |
| gyrA | 100.00 | 43/43(100.00) | 11/11(100.00) | 5/5(100.00) | 3/3(100.00) | |
| Amphenicols | floR | 48.39 | 18/43(41.86) | 7/11(63.63) | 3/5(60.00) | 2/3(66.67) |
| Sulfonamides | sul1 | 50.00 | 21/43(48.84) | 6/11(54.55) | 2/5(40.00) | 2/3(66.67) |
| sul2 | 41.94 | 14/43(32.56) | 7/11(63.63) | 3/5(60.00) | 2/3(66.67) | |
| Tetracyclines | tetA | 41.94 | 15/43(34.88) | 7/11(63.63) | 2/5(40.00) | 2/3(66.67) |
| tetB | 8.06 | 1/43(2.33) | 2/11(18.18) | 1/5(20.00) | 1/3(33.33) | |
| Aminoglycosides | aadA1 | 38.71 | 19/43(44.19) | 4/11(36.37) | 1/5(20.00) | 0/3(0.00) |
| aacC2 | 19.35 | 6/43(13.95) | 2/11(18.18) | 2/5(40.00) | 2/3(66.67) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, G.; Ahmad, S.; Li, Y.; Yan, D.; Yang, S.; Chen, S.; Qiu, Z.; Yu, X.; Li, N.; Li, Y.; et al. Epidemiological, Virulence, and Antibiotic Resistance Analysis of Klebsiella pneumoniae, a Major Source of Threat to Livestock and Poultry in Some Regions of Xinjiang, China. Animals 2024, 14, 1433. https://doi.org/10.3390/ani14101433
Hou G, Ahmad S, Li Y, Yan D, Yang S, Chen S, Qiu Z, Yu X, Li N, Li Y, et al. Epidemiological, Virulence, and Antibiotic Resistance Analysis of Klebsiella pneumoniae, a Major Source of Threat to Livestock and Poultry in Some Regions of Xinjiang, China. Animals. 2024; 14(10):1433. https://doi.org/10.3390/ani14101433
Chicago/Turabian StyleHou, Gongmingzhu, Sajjad Ahmad, Yanfang Li, Duo Yan, Shuhan Yang, Siqi Chen, Zhengqing Qiu, Xingyu Yu, Nana Li, Yang Li, and et al. 2024. "Epidemiological, Virulence, and Antibiotic Resistance Analysis of Klebsiella pneumoniae, a Major Source of Threat to Livestock and Poultry in Some Regions of Xinjiang, China" Animals 14, no. 10: 1433. https://doi.org/10.3390/ani14101433