
The Coupling Effects of PGPR Inoculation and Foliar Spraying of Strigolactone in Mitigating the Negative Effect of Salt Stress in Wheat Plants: Insights from Phytochemical, Growth, and Yield Attributes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Inoculation Procedure of PGPR
2.2. Plant Material and Growing Conditions
2.3. Induction of Salt Stress
2.4. Exogenous Application of GR24
2.5. Determination of RWC and EL
2.6. Growth and Yield Parameters
2.7. Estimation of Photosynthetic Pigments
2.8. Estimation of Na+ and K+ Ions in Roots and Leaves
2.9. Determination of MDA and H2O2
2.10. Leaf Proline and GB Content
2.11. Soluble Sugars and Starch Content
2.12. Total Protein Content and Antioxidative Enzymes Activities
2.13. Statistical Analysis
3. Results
3.1. Chlorophyll Pigment Content, RWC and EL
3.2. Ionic and Phytochemical Analysis
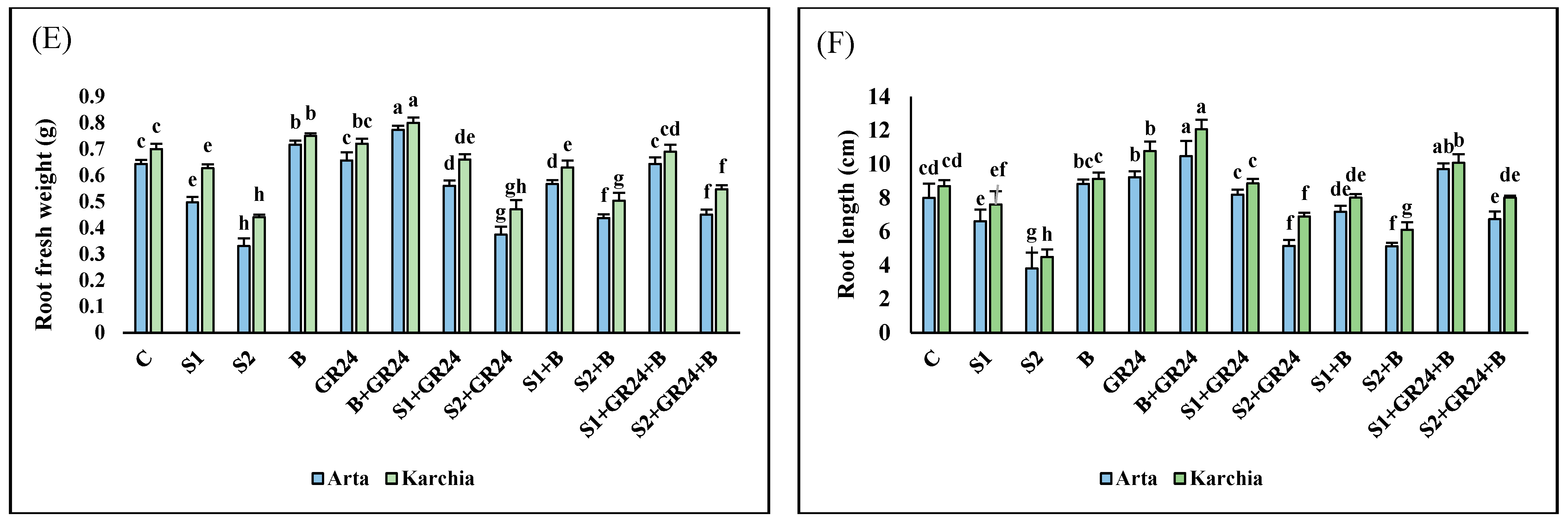
3.3. Yield Traits and Root Growth
3.4. Antioxidant Enzymes Activity and Protein Content
3.5. MDA, H2O2, GB and Proline Contents
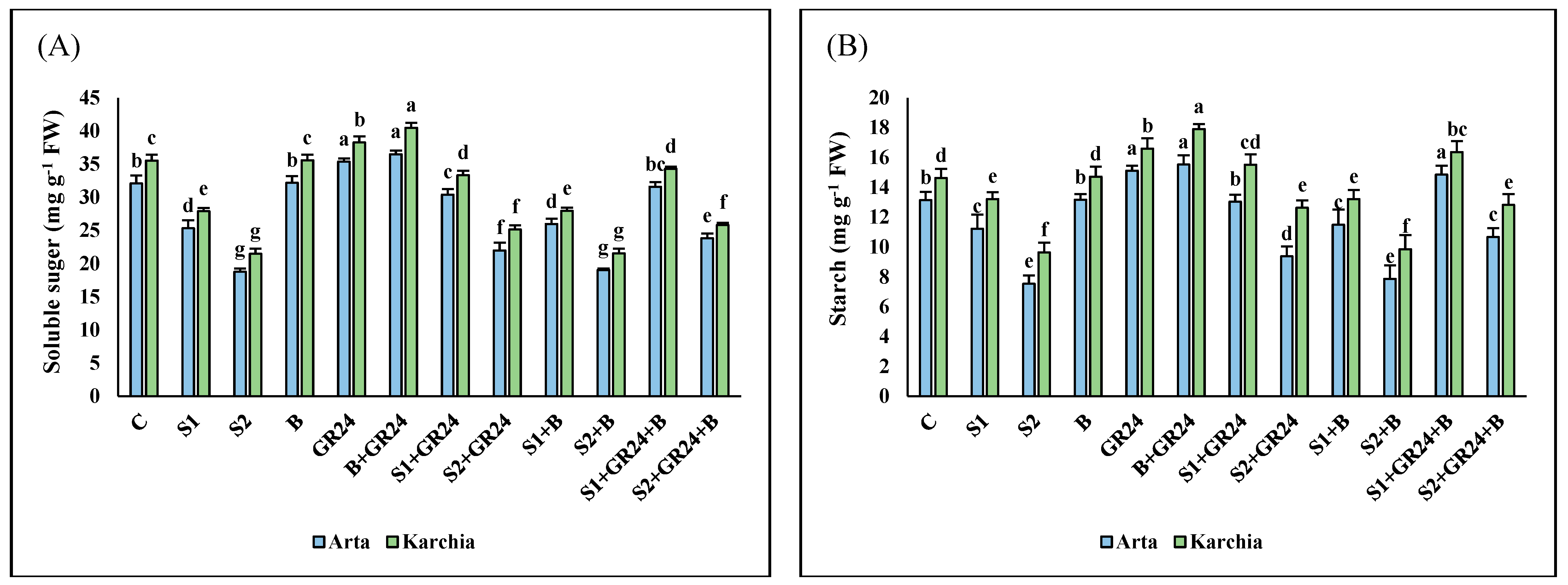
3.6. Soluble Sugars and Starch Contents


3.7. Pearson Correlation Analysis
3.8. Hierarchical Clustering Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Soothar, M.K.; Mounkaila Hamani, A.K.; Kumar Sootahar, M.; Sun, J.; Yang, G.; Bhatti, S.M.; Traore, A. Assessment of acidic biochar on the growth, physiology, and nutrients uptake of maize (Zea mays L.) seedlings under salinity stress. Sustainability 2021, 13, 3150. [Google Scholar] [CrossRef]
- Bibi, S.; Ullah, S.; Hafeez, A.; Khan, M.N.; Javed, M.A.; Ali, B.; Din, I.U.; Bangash, S.A.K.; Wahab, S.; Wahid, N.; et al. Exogenous Ca/Mg quotient reduces the inhibitory effects of PEG induced osmotic stress on Avena sativa L. Braz. J. Biol. 2022, 84, e264642. [Google Scholar] [CrossRef] [PubMed]
- Yasmeen, S.; Wahab, A.; Saleem, M.H.; Ali, B.; Qureshi, K.A.; Jaremko, M. Melatonin as a foliar application and adaptation in lentil (Lens culinaris Medik.) crops under drought stress. Sustainability 2022, 14, 16345. [Google Scholar] [CrossRef]
- Salam, A.; Afridi, M.S.; Javed, M.A.; Saleem, A.; Hafeez, A.; Khan, A.R.; Zeeshan, M.; Ali, B.; Azhar, W.; Sumaira and Ulhassan, Z. Nano-priming against abiotic stress: A way forward towards sustainable agriculture. Sustainability 2022, 14, 14880. [Google Scholar] [CrossRef]
- Gul, S.; Javed, S.; Azeem, M.; Aftab, A.; Anwaar, N.; Mehmood, T.; Zeshan, B. Application of Bacillus subtilis for the Alleviation of Salinity Stress in Different Cultivars of Wheat (Tritium aestivum L.). Agronomy 2023, 13, 437. [Google Scholar] [CrossRef]
- Azeem, M.; Pirjan, K.; Qasim, M.; Mahmood, A.; Javed, T.; Muhammad, H.; Yang, S.; Dong, R.; Ali, B.; Rahimi, M. Salinity stress improves antioxidant potential by modulating physio-biochemical responses in Moringa oleifera Lam. Sci. Rep. 2023, 13, 2895. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Aslam, M.T.; Alhammad, B.A.; Hassan, M.U.; Maqbool, R.; Chattha, M.U.; Khan, I.; Gitari, H.I.; Uslu, O.S.; Roy, R.; et al. Salinity stress in wheat: Effects, mechanisms and management strategies. Phyton Int. J. Exp. Bot. 2022, 91, 667–694. [Google Scholar] [CrossRef]
- Nafees, M.; Rizwan, M.; Ahmad, S.; Ali, S.; Obaid, W.A.; Alsubeie, M.S.; Darwish, D.B.E.; Abeed, A.H. Combined effect of Zinc lysine and biochar on growth and physiology of wheat (Triticum aestivum L.) to alleviate salinity stress. Front. Plant Sci. 2023, 13, 1017282. [Google Scholar]
- Zeeshan, M.; Lu, M.; Sehar, S.; Holford, P.; Wu, F. Comparison of biochemical, anatomical, morphological, and physiological responses to salinity stress in wheat and barley genotypes deferring in salinity tolerance. Agronomy 2020, 10, 127. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A. Abiotic stress, generation of reactive oxygen species, and their consequences: An overview. In Reactive Oxygen Species in Plants: Boon or Bane-Revisiting the Role of ROS; Singh, V.P., Singh, S., Tripathi, D.K., Prasad, S.M., Chauhan, D.K., Eds.; Wiley: Hoboken, NJ, USA, 2017; pp. 23–50. [Google Scholar]
- Faizan, M.; Faraz, A.; Hayat, S. Dose-dependent response of epibrassinolide on the growth, photosynthesis, and antioxidant system of tomato plants. Ind. Hort. J. 2018, 8, 68–76. [Google Scholar]
- Cook, C.E.; Whichard, L.P.; Wall, M.; Egley, G.H.; Coggon, P.; Luhan, P.A.; McPhail, A.T. Germination stimulants. II. Structure of strigol, a potent seed germination stimulant for witchweed (Striga lutea). J. Am. Chem. Soc. 1972, 94, 6198–6199. [Google Scholar] [CrossRef]
- Seto, Y.; Yasui, R.; Kameoka, H.; Tamiru, M.; Cao, M.; Terauchi, R.; Sakurada, A.; Hirano, R.; Kisugi, T.; Hanada, A.; et al. Strigolactone perception and deactivation by a hydrolase receptor DWARF14. Nat. Commun. 2019, 10, 191. [Google Scholar] [CrossRef]
- Alvi, A.F.; Sehar, Z.; Fatma, M.; Masood, A.; Khan, N.A. Strigolactone: An Emerging Growth Regulator for developing resilience in plants. Plants 2022, 11, 2604. [Google Scholar] [CrossRef]
- Begum, N.; Zhao, T.; Antwi-Boasiako, A.; Zhang, L.; Ashraf, M. Role of strigolactones in rhizobiology: Plant-microbe interactions. In Phytohormones and Stress-Responsive Secondary Metabolites; Ozturk, M., Bhat, R.A., Ashraf, M., Tonelli, F.M.P., Turkyilmaz Unal, B., Dar, G.H., Eds.; Academic Press: New York, NY, USA, 2023; pp. 13–25. [Google Scholar]
- Koltai, H.; Prandi, C. Strigolactones: Biosynthesis, Synthesis, and Functions in Plant Growth and Stress Responses. In Phytohormones: A Window to Metabolism, Signaling and Biotechnological Applications; Tran, L.S., Pal, S., Eds.; Springer: New York, NY, USA, 2014; pp. 265–288. [Google Scholar]
- Li, C.; Lu, X.; Liu, Y.; Xu, J.; Yu, W. Strigolactone Alleviates the Adverse Effects of Salt Stress on Seed Germination in Cucumber by Enhancing Antioxidant Capacity. Antioxidants 2023, 12, 1043. [Google Scholar] [CrossRef]
- Ma, N.; Hu, C.; Wan, L.; Hu, Q.; Xiong, J.; Zhang, C. Strigolactones improve plant growth and photosynthesis and alleviate oxidative stress under salinity in rapeseed (Brassica napus L.) by regulating gene expression. Front. Plant Sci. 2017, 8, 1671. [Google Scholar] [CrossRef] [PubMed]
- Kausar, F.; Shahbaz, M. Influence of Strigolactone (GR24) as a Seed Treatment on Growth, Gas Exchange and Chlorophyll Fluorescence of Wheat under Saline Conditions. Int. J. Agric. Biol. 2017, 19, 321–327. [Google Scholar] [CrossRef]
- Ling, F.; Su, Q.; Jiang, H.; Cui, J.; He, X.; Wu, Z.; Zhang, Z.; Liu, J.; Zhao, Y. Effects of strigolactone on photosynthetic and physiological characteristics in salt-stressed rice seedlings. Sci. Rep. 2020, 10, 6183. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, M.; Zulfiqar, H.; Farooq, M.A.; Ali, S.; Tufail, A.; Kanwal, S.; Shaheen, M.R.; Sajid, M.; Gul, H.; Jamal, A.; et al. Strigolactone (GR24) Application positively regulates photosynthetic attributes, stress-related metabolites and antioxidant enzymatic activities of ornamental sunflower (Helianthus annuus cv. Vincent’s Choice) under salinity stress. Agriculture 2022, 13, 50. [Google Scholar] [CrossRef]
- Song, Y.; Lv, D.; Jiang, M.; Zhiying, E.; Han, Y.; Sun, Y.; Zhu, S.; Chen, J.; Zhao, T. Exogenous Strigolactones Enhance Salinity Tolerance in Cotton (Gossypium hirsutum L.) Seedlings. Plant Stress 2023, 10, 100235. [Google Scholar] [CrossRef]
- Faisal, M.; Faizan, M.; Tonny, S.H.; Rajput, V.D.; Minkina, T.; Alatar, A.A.; Pathirana, R. Strigolactone-Mediated Mitigation of Negative Effects of Salinity Stress in Solanum lycopersicum through Reducing the Oxidative Damage. Sustainability 2023, 15, 5805. [Google Scholar] [CrossRef]
- Etesami, H.; Glick, B.R. Halotolerant plant growth–promoting bacteria: Prospects for alleviating salinity stress in plants. Environ. Exp. Bot. 2020, 178, 104124. [Google Scholar] [CrossRef]
- Etesami, H. Potential advantage of rhizosheath microbiome, in contrast to rhizosphere microbiome, to improve drought tolerance in crops. Rhizosphere 2021, 20, 100439. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant growth-promoting bacteria: Biological tools for the mitigation of salinity stress in plants. Front. Microbiol. 2020, 11, 1216. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Jeong, B.R.; Glick, B.R. Potential use of Bacillus spp. as an effective biostimulant against abiotic stresses in crops—A review. Curr. Res. Biotechnol. 2023, 5, 100128. [Google Scholar] [CrossRef]
- Bagheri, N.; Ahmadzadeh, M.; Ghasemi, S.; Vahidinasab, M.; Ghoreshi, S.S. Bacillus amyloliquefaciens UTB96, an effective biocontrol and aflatoxin-degrading bacterium. Biocontrol Plant Protect. 2018, 6, 1–17. [Google Scholar]
- Bagheri, N.; Ahmadzadeh, M.; Mariotte, P.; Jouzani, G.S. Behavior and interactions of the plant growth-promoting bacteria Azospirillum oryzae NBT506 and Bacillus velezensis UTB96 in a co-culture system. World J. Microbiol. Biotechnol. 2022, 38, 101. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, N.; Ahmadzadeh, M.; Salehi Jouzani, G. Interaction of Bacillus amyloliquefaciens and Azospirillum oryzae on wheat growth promotion and Fusarium graminearum disease inhibition. Crop Biotechnol. 2019, 9, 19–33. [Google Scholar]
- Bai, Y.; Zhou, Y.; Yue, T.; Huang, Y.; He, C.; Jiang, W.; Liu, H.; Zeng, H.; Wang, J. Plant growth-promoting rhizobacteria Bacillus velezensis JB0319 promotes lettuce growth under salt stress by modulating plant physiology and changing the rhizosphere bacterial community. Environ. Exp. Bot. 2023, 213, 105451. [Google Scholar] [CrossRef]
- Çam, S.; Küçük, Ç.; Almaca, A. Bacillus strains exhibit various plant growth-promoting traits, and their biofilm-forming capability correlates to their salt stress alleviation effect on maize seedlings. J. Biotech. 2023, 369, 35–42. [Google Scholar] [CrossRef]
- Zahra, S.T.; Tariq, M.; Abdullah, M.; Azeem, F.; Ashraf, M.A. Dominance of Bacillus species in the wheat (Triticum aestivum L.) rhizosphere and their plant growth promoting potential under salt stress conditions. PeerJ 2023, 11, 14621. [Google Scholar] [CrossRef]
- Ghazala, I.; Chiab, N.; Saidi, M.N.; Gargouri-Bouzid, R. The Plant Growth-Promoting Bacteria Strain Bacillus mojavensis I4 Enhanced Salt Stress Tolerance in Durum Wheat. Curr. Microbiol. 2023, 80, 178. [Google Scholar] [CrossRef]
- Haroon, U.; Munis, M.F.H.; Liaquat, F.; Khizar, M.; Elahi, M.; Chaudhary, H.J. Biofilm formation and flocculation potential analysis of halotolerant Bacillus tequilensis and its inoculation in soil to mitigate salinity stress of chickpea. Physiol. Mol. Biol. Plants 2023, 292, 277–288. [Google Scholar] [CrossRef]
- Mahdi, I.; Allaoui, A.; Fahsi, N.; Biskri, L. Bacillus velezensis QA2 potentially induced salt stress tolerance and enhanced phosphate uptake in quinoa plants. Microorganisms 2022, 10, 1836. [Google Scholar] [CrossRef]
- Bashir, T.; Naz, S.; Bano, A. Plant growth-promoting rhizobacteria, in combination with plant growth regulators, attenuates the effect of drought stress. Pak. J. Bot. 2020, 52, 783–792. [Google Scholar] [CrossRef]
- Ali, Q.; Ahmad, M.; Kamran, M.; Ashraf, S.; Shabaan, M.; Babar, B.H.; Zulfiqar, U.; Haider, F.U.; Ali, M.A.; Elshikh, M.S. Synergistic effects of rhizobacteria and salicylic acid on maize salt-stress tolerance. Plants 2023, 12, 2519. [Google Scholar] [CrossRef]
- Porcel, R.; Zamarreño, Á.M.; García-Mina, J.M.; Aroca, R. Involvement of plant endogenous ABA in Bacillus megaterium PGPR activity in tomato plants. BMC Plant Biol. 2014, 14, 36. [Google Scholar] [CrossRef]
- Mohamed, I.; Eid, K.E.; Abbas, M.H.; Salem, A.A.; Ahmed, N.; Ali, M.; Shah, G.M.; Fang, C. Use of plant growth promoting Rhizobacteria (PGPR) and mycorrhizae to improve the growth and nutrient utilization of common bean in a soil infected with white rot fungi. Ecotoxicol. Environ. Saf. 2019, 171, 539–548. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A. Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant Soil. 2003, 253, 201–218. [Google Scholar] [CrossRef]
- Schonfeld, M.A.; Johnson, R.C.; Carver, B.F.; Mornhinweg, D.W. Water relations in winter wheat as drought resistance indicators. Crop Sci. 1988, 28, 526–531. [Google Scholar] [CrossRef]
- Valentovič, P.; Luxová, M.; Kolarovič, L.; Gašparíková, O. Effect of osmotic stress on compatible solutes content, membrane stability, and water relations in two maize cultivars. Plant Soil Environ. 2006, 52, 184. [Google Scholar] [CrossRef]
- Arnon, A.N. Method of extraction of chlorophyll in the plants. Agron. J. 1967, 23, 112–121. [Google Scholar]
- Allen, D.G.; Eisner, D.A.; Wray, S.C. Sodium pump: Birthday present for digitalis. Nature 1985, 316, 674–675. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A.J.P.S. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.A.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water-soluble quaternary ammonium compounds. Plant Soil. 1983, 70, 303–307. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Sheligl, H.Q. Die verwertung orgngischer souren durch chlorella lincht. Planta J. 1986, 47, 51. [Google Scholar]
- Pagariya, M.C.; Devarumath, R.M.; Kawar, P.G. Biochemical characterization, and identification of differentially expressed candidate genes in salt-stressed sugarcane. Plant Sci. 2012, 184, 1–13. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for quantitating microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- MacAdam, J.W.; Nelson, C.J.; Sharp, R.E. Peroxidase activity in the leaf elongation zone of tall fescue: I. Spatial distribution of ionically bound peroxidase activity in genotypes differing in length of the elongation zone. Plant Physiol. 1992, 99, 872–878. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.A.M.E.L.A.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, M.R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of reactive oxygen species and antioxidant defense in plants under salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef] [PubMed]
- Kleman, J.; Matusova, R. Strigolactones: Current research progress in the response of plants to abiotic stress. Biologia 2023, 78, 307–318. [Google Scholar] [CrossRef]
- Bhat, M.A.; Kumar, V.; Bhat, M.A.; Wani, I.A.; Dar, F.L.; Farooq, I.; Bhatti, F.; Koser, R.; Rahman, S.; Jan, A.T. Mechanistic insights of the interaction of plant growth-promoting rhizobacteria (PGPR) with plant roots toward enhancing plant productivity by alleviating salinity stress. Front. Microbiol. 2020, 11, 1952. [Google Scholar] [CrossRef]
- Sen, A.; Islam, M.M.; Zaman, E.; Ghosh, U.K.; Momtaz, M.B.; Islam, M.A.; Urmi, T.A.; Mamun, M.A.A.; Rahman, M.M.; Kamal, M.Z.U.; et al. Agro-Morphological, Yield and Biochemical Responses of Selected Wheat (Triticum aestivum L.) Genotypes to Salt Stress. Agronomy 2022, 12, 3027. [Google Scholar] [CrossRef]
- Fu, H.; Yang, Y. How plants tolerate salt stress. Curr. Issues Mol. Biol. 2023, 45, 5914–5934. [Google Scholar] [CrossRef] [PubMed]
- Mazhar, Z.; Akhtar, J.; Alhodaib, A.; Naz, T.; Zafar, M.I.; Iqbal, M.M.; Fatima, H.; Naz, I. Efficacy of ZnO nanoparticles in Zn fortification and partitioning of wheat and rice grains under salt stress. Sci. Rep. 2023, 13, 2022. [Google Scholar] [CrossRef]
- El-Taher, A.M.; Abd El-Raouf, H.S.; Osman, N.A.; Azoz, S.N.; Omar, M.A.; Elkelish, A.; Abd El-Hady, M.A. Effect of salt stress and foliar application of salicylic acid on morphological, biochemical, anatomical, and productivity characteristics of cowpea (Vigna unguiculata L.) plants. Plants 2021, 11, 115. [Google Scholar] [CrossRef]
- Huang, S.; Gill, S.; Ramzan, M.; Ahmad, M.Z.; Danish, S.; Huang, P.; Al Obaid, S.; Alharbi, S.A. Uncovering the impact of AM fungi on wheat nutrient uptake, ion homeostasis, oxidative stress, and antioxidant defense under salinity stress. Sci. Rep. 2023, 13, 8249. [Google Scholar] [CrossRef] [PubMed]
- Shelden, M.C.; Munns, R. Crop root system plasticity for improved yields in saline soils. Front. Plant Sci. 2023, 14, 1120583. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, I.; Shahbaz, M.; Wahid, M.A. Potential role of foliage applied strigolactone (GR24) on photosynthetic pigments, gas exchange attributes, mineral nutrients and yield components of Zea mays (L.) under saline regimes. Gesunde Pflanz. 2023, 75, 577–591. [Google Scholar] [CrossRef]
- Sharifi, P.; Bidabadi, S.S. Strigolactone could enhance gas exchange through augmented antioxidant defense system in Salvia nemorosa L. plants subjected to saline conditions stress. Ind. Crops Prod. 2020, 151, 112460. [Google Scholar] [CrossRef]
- Liu, R.; Hou, J.; Li, H.; Xu, P.; Zhang, Z.; Zhang, X. Association of TaD14-4D, a gene involved in strigolactone signaling, with yield contributing traits in wheat. Int. J. Mol. Sci. 2021, 22, 3748. [Google Scholar] [CrossRef] [PubMed]
- Ruyter-Spira, C.; Kohlen, W.; Charnikhova, T.; van Zeijl, A.; van Bezouwen, L.; De Ruijter, N.; Cardoso, C.; Lopez-Raez, J.A.; Matusova, R.; Bours, R.; et al. Physiological effects of the synthetic strigolactone analog GR24 on root system architecture in Arabidopsis: Another belowground role for strigolactones? Plant Physiol. 2011, 155, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Kusajima, M.; Fujita, M.; Soudthedlath, K.; Nakamura, H.; Yoneyama, K.; Nomura, T.; Akiyama, K.; Maruyama-Nakashita, A.; Asami, T.; Nakashita, H. Strigolactones modulate salicylic acid-mediated disease resistance in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 5246. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, A.; Turgay, O.C.; Farooq, M.; Hayat, R. Seed biopriming with plant growth promoting rhizobacteria: A review. FEMS Microbiol. Ecol. 2016, 92, 112. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Bano, A. Role of PGPR in the reclamation and revegetation of saline land. Pak. J. Bot. 2019, 51, 27–35. [Google Scholar] [CrossRef]
- Panwar, M.; Tewari, R.; Nayyar, H. Native halo-tolerant plant growth promoting rhizobacteria Enterococcus and Pantoea sp. improve seed yield of Mungbean (Vigna radiata L.) under soil salinity by reducing sodium uptake and stress injury. Physiol. Mol. Biol. Plants 2016, 22, 445–459. [Google Scholar] [CrossRef]
- Nawaz, A.; Shahbaz, M.; Asadullah; Imran, A.; Marghoob, M.U.; Imtiaz, M.; Mubeen, F. Potential of salt tolerant PGPR in growth and yield augmentation of wheat (Triticum aestivum L.) under saline conditions. Front. Microbiol. 2020, 11, 2019. [Google Scholar] [CrossRef] [PubMed]
- Tena, W.; Wolde-Meskel, E.; Walley, F. Symbiotic efficiency of native and exotic Rhizobium strains nodulating lentil (Lens culinaris Medik.) in soils of Southern Ethiopia. Agronomy 2016, 6, 11. [Google Scholar] [CrossRef]
- Ren, J.; Ye, J.; Yin, L.; Li, G.; Deng, X.; Wang, S. Exogenous melatonin improves salt tolerance by mitigating osmotic, ion, and oxidative stresses in maize seedlings. Agronomy 2020, 10, 663. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, L.; Li, H.; Zhang, S.; Fu, X.; Zhai, X.; Yang, N.; Shen, J.; Li, R.; Li, D. Exogenous melatonin promotes salt tolerance by removing active oxygen and maintaining ion balance in wheat (Triticum aestivum L.). Front. Plant Sci. 2022, 12, 787062. [Google Scholar] [CrossRef]
- Peña Calzada, K.; Calero Hurtado, A.; Olivera Viciedo, D.; Habermann, E.; de Mello Prado, R.; de Oliveira, R.; Ajila, G.; Tenesaca, L.F.L.; Rodríguez, J.C.; Gratão, P.L. Regulatory role of silicon on growth, potassium uptake, ionic homeostasis, proline accumulation, and antioxidant capacity of soybean plants under salt stress. J. Plant Growth Regul. 2023, 42, 4528–4540. [Google Scholar] [CrossRef]
- Mehrabi, S.S.; Sabokdast, M.; Bihamta, M.R.; Soorni, J.; Mirmazloum, I. Strigolactone-mediated amelioration of salinity stress in bread wheat: Insights from phytochemical and ion channels related genes expression analyses. Plant Stress 2024, 11, 100324. [Google Scholar] [CrossRef]
- Zulfiqar, H.; Shahbaz, M.; Ahsan, M.; Nafees, M.; Nadeem, H.; Akram, M.; Maqsood, A.; Ahmar, S.; Kamran, M.; Alamri, S.; et al. Strigolactone (GR24) induced salinity tolerance in sunflower (Helianthus annuus L.) by ameliorating morpho-physiological and biochemical attributes under in vitro conditions. J. Plant Growth Regul. 2021, 40, 2079–2091. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, F.; Mickan, B.; Wang, D.; Wang, W. Physiological, proteomic, and metabolomic analysis provide insights into Bacillus sp.-mediated salt tolerance in wheat. Plant Cell Rep. 2022, 41, 95–118. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Wu, G.; Veronican Njeri, K.; Shen, Q.; Zhang, N.; Zhang, R. Induced maize salt tolerance by rhizosphere inoculation of Bacillus amyloliquefaciens SQR9. Physiol. Plant. 2016, 158, 34–44. [Google Scholar] [CrossRef]
- Asif, S.; Jan, R.; Kim, N.; Asaf, S.; Lubna; Khan, M.A.; Kim, E.G.; Jang, Y.H.; Bhatta, D.; Lee, I.J.; et al. Halotolerant endophytic bacteria alleviate salinity stress in rice (Oryza sativa L.) by modulating ion content, endogenous hormones, the antioxidant system and gene expression. BMC Plant Biol. 2023, 23, 494. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- Jesmin, A.; Anh, L.H.; Mai, N.P.; Khanh, T.D.; Xuan, T.D. Fulvic Acid Improves Salinity Tolerance of Rice Seedlings: Evidence from Phenotypic Performance, Relevant Phenolic Acids, and Momilactones. Plants 2023, 12, 2359. [Google Scholar] [CrossRef]
- Akter, T.; Mimma, A.A.; Haque, M.A.; Hossain, M.M.; Ghosh, T.K.; Zinan, N.; Chowdhury, M.Z.H.; Islam, S.M.N. Seed priming with Beauveria bassiana improves growth and salt stress response in rice. Environ. Exp. Bot. 2023, 213, 105427. [Google Scholar] [CrossRef]
- Lu, T.; Yu, H.; Li, Q.; Chai, L.; Jiang, W. Improving plant growth and alleviating photosynthetic inhibition and oxidative stress from low-light stress with exogenous GR24 in tomato (Solanum lycopersicum L.) seedlings. Front. Plant Sci. 2019, 10, 490. [Google Scholar] [CrossRef]
- Ilyas, N.; Mazhar, R.; Yasmin, H.; Khan, W.; Iqbal, S.; Enshasy, H.E.; Dailin, D.J. Rhizobacteria isolated from saline soil induce systemic tolerance in wheat (Triticum aestivum L.) against salinity stress. Agronomy 2020, 10, 989. [Google Scholar] [CrossRef]
- Patani, A.; Prajapati, D.; Ali, D.; Kalasariya, H.; Yadav, V.K.; Tank, J.; Bagatharia, S.; Joshi, M.; Patel, A. Evaluation of the growth-inducing efficacy of various Bacillus species on the salt-stressed tomato (Lycopersicon esculentum Mill.). Front. Plant Sci. 2023, 14, 1168155. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Curá, J.A. The Role of beneficial microorganisms and salicylic acid in improving rainfed agriculture and future food safety. Microorganisms 2020, 8, 1018. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Ul-Allah, S.; Ijaz, M.; Sher, A.; Butt, M.; Abbas, T.; Irfan, M.; Fatima, T.; Alfarraj, S.; Alharbi, S.A. Exogenous application of strigolactone alleviates drought stress in maize seedlings by regulating the physiological and antioxidants defense mechanisms. Cereal Res. Commun. 2022, 50, 263–272. [Google Scholar] [CrossRef]
- Liu, J.; He, H.; Vitali, M.; Visentin, I.; Charnikhova, T.; Haider, I.; Schubert, A.; Ruyter-Spira, C.; Bouwmeester, H.J.; Lovisolo, C.; et al. Osmotic stress represses strigolactone biosynthesis in Lotus japonicus roots: Exploring the interaction between strigolactones and ABA under abiotic stress. Planta 2015, 241, 1435–1451. [Google Scholar] [CrossRef] [PubMed]
- ALKahtani, M.D.; Attia, K.A.; Hafez, Y.M.; Khan, N.; Eid, A.M.; Ali, M.A.; Abdelaal, K.A. Chlorophyll fluorescence parameters and antioxidant defense system can display salt tolerance of salt acclimated sweet pepper plants treated with chitosan and plant growth promoting rhizobacteria. Agronomy 2020, 10, 1180. [Google Scholar] [CrossRef]
- Vurro, M.; Prandi, C.; Baroccio, F. Strigolactones: How far is their commercial use for agricultural purposes? Pest Manag. Sci. 2016, 72, 2026–2034. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Rahman, M.M.; Nguyen, K.H.; Li, W.; Watanabe, Y.; Tran, C.D.; Zhang, M.; Itouga, M.; Fujita, M.; Tran, L.S.P. Strigolactones regulate arsenate uptake, vacuolar-sequestration and antioxidant defense responses to resist arsenic toxicity in rice roots. J. Hazard. Mater. 2021, 415, 125589. [Google Scholar] [CrossRef]
- Guo, S.; Wei, X.; Ma, B.; Ma, Y.; Yu, Z.; Li, P. Foliar application of strigolactones improves the desiccation tolerance, grain yield and water use efficiency in dryland wheat through modulation of non-hydraulic root signals and antioxidant defense. Stress Biology 2023, 3, 54. [Google Scholar] [CrossRef]
- Gururani, M.A.; Upadhyaya, C.P.; Baskar, V.; Venkatesh, J.; Nookaraju, A.; Park, S.W. Plant growth-promoting rhizobacteria enhance abiotic stress tolerance in Solanum tuberosum through inducing changes in the expression of ROS-scavenging enzymes and improved photosynthetic performance. J. Plant Growth Regul. 2013, 32, 245–258. [Google Scholar] [CrossRef]
- Ren, L.; Zhang, Y.; Zhou, J.L.; Wang, G.; Mo, Y.; Ling, Y.; Huang, Y.; Zhang, Y.; Hu, H.; Wang, Y. RL-WG26 mediated salt stress tolerance in rice seedlings: A new insight into molecular mechanisms. Plant Stress 2024, 11, 100306. [Google Scholar] [CrossRef]
- Azeem, M.A.; Ali, F.; Ullah, A.; Iqbal, M.; Ali, K.; Al Farraj, D.A.; Elshikh, M.S.; Naz, Q.; Munis, M.F.H.; Chaudhary, H.J. Exploration of plant growth promoting traits and regulatory mechanisms of Bacillus anthracis PM21 in enhancing salt stress tolerance in maize. Environ. Sci. Pollut. Res. 2023, 30, 77499–77516. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Asma, H.F.; Iqbal, M. Exogenous menadione sodium bisulfite mitigates specific ion toxicity and oxidative damage in salinity-stressed okra (Abelmoschus esculentus Moench). Acta Physiol. Plant 2019, 41, 187. [Google Scholar] [CrossRef]
- Rasheed, R.; Ashraf, M.A.; Parveen, S.; Iqbal, M.; Hussain, I. Effect of salt stress on different growth and biochemical attributes in two canola (Brassica napus L.) cultivars. Commun. Soil Sci. Plant Anal. 2014, 45, 669–679. [Google Scholar] [CrossRef]
- Aouz, A.; Khan, I.; Chattha, M.B.; Ahmad, S.; Ali, M.; Ali, I.; Ali, A.; Alqahtani, F.M.; Hashem, M.; Albishi, T.S.; et al. Silicon Induces Heat and Salinity Tolerance in Wheat by Increasing Antioxidant Activities, Photosynthetic Activity, Nutrient Homeostasis, and Osmo-Protectant Synthesis. Plants 2023, 12, 2606. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Hafeez, A.; Rasheed, R.; Hussain, I.; Farooq, U.; Rizwan, M.; Ali, S. Evaluation of Physio-Morphological and Biochemical Responses for Salt Tolerance in Wheat (Triticum aestivum L.) Cultivars. J. Plant Growth Regul. 2023, 42, 4402–4422. [Google Scholar] [CrossRef]
- Tariq, A.; Ullah, I.; Sardans, J.; Graciano, C.; Mussarat, S.; Ullah, A.; Zeng, F.; Wang, W.; Al-Bakre, D.A.; Ahmed, Z.; et al. Strigolactones can be a potential tool to fight environmental stresses in arid lands. Environ. Res. 2023, 229, 115966. [Google Scholar] [CrossRef]
- Sharma, A.; Chakdar, H.; Vaishnav, A.; Srivastava, A.K.; Khan, N.; Bansal, Y.K.; Kaushik, R. Multifarious Plant Growth-Promoting Rhizobacterium Enterobacter sp. CM94-Mediated Systemic Tolerance and Growth Promotion of Chickpea (Cicer arietinum L.) under Salinity Stress. Front. Biosci. (Landmark Ed.) 2023, 28, 241. [Google Scholar]
- YILDIZ, M.T.; Okan, A.C.A.R.; ÖZTÜRK, F.; HACIOĞLU, N. Some Physiological and Biochemical Effects of Bacillus thuringiensis LU3 Biopriming in Common Wheat (Triticum aestivum L.) under Salt Stress. KSÜ Tarım Ve Dŏga Derg. 2023, 26, 1086–1098. [Google Scholar]
- Khan, V.; Umar, S.; Iqbal, N. Palliating salt stress in mustard through plant-growth-promoting rhizobacteria: Regulation of secondary metabolites, osmolytes, antioxidative enzymes, and stress ethylene. Plants 2023, 12, 705. [Google Scholar] [CrossRef]
- Ali, B.; Wang, X.; Saleem, M.H.; Sumaira; Hafeez, A.; Afridi, M.S.; Khan, S.; Ullah, I.; Amaral Júnior, A.T.D.; Alatawi, A.; et al. PGPR-mediated salt tolerance in maize by modulating plant physiology, antioxidant defense, compatible solutes accumulation and bio-surfactant producing genes. Plants 2022, 11, 345. [Google Scholar] [CrossRef]
- El Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savouré, A. How does proline treatment promote salt stress tolerance during crop plant development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef]
- Al-Amri, A.A.; Alsubaie, Q.D.; Alamri, S.A.; Siddiqui, M.H. Strigolactone Analog GR24 Induces Seed Germination and Improves Growth Performance of Different Genotypes of Tomato. J. Plant Growth Regul. 2023, 42, 5653–5666. [Google Scholar] [CrossRef]
- Karimzadeh, J.; Alikhani, H.A.; Etesami, H.; Pourbabaei, A.A. Improved phosphorus uptake by the wheat plant (Triticum aestivum L.) with rhizosphere fluorescent Pseudomonads strains under water-deficit stress. J. Plant Growth Regul. 2020, 40, 162–178. [Google Scholar] [CrossRef]
- Shirmohammadi, E.; Alikhani, H.A.; Pourbabaei, A.A.; Etesami, H. Improved phosphorus (P) uptake and yield of rainfed wheat fed with P fertilizer by drought-tolerant phosphate-solubilizing fluorescent pseudomonads strains: A field study in drylands. J. Soil Sci. Plant Nutr. 2020, 20, 2195–2211. [Google Scholar] [CrossRef]
- Mahmoud, O.M.B.; Hidri, R.; Talbi-Zribi, O.; Taamalli, W.; Abdelly, C.; Djébali, N. Auxin and proline producing rhizobacteria mitigate salt-induced growth inhibition of barley plants by enhancing water and nutrient status. S. Afr. J. Bot. 2020, 128, 209–217. [Google Scholar] [CrossRef]
- Misra, S.; Chauhan, P.S. ACC deaminase-producing rhizosphere competent Bacillus spp. Mitigate salt stress and promote Zea mays growth by modulating ethylene metabolism. 3 Biotech 2020, 10, 119. [Google Scholar] [CrossRef]
- Chen, T.H.; Murata, N. Glycinebetaine protects plants against abiotic stress: Mechanisms and biotechnological applications. Plant Cell Environ. 2011, 34, 1–20. [Google Scholar] [CrossRef]
- Mäkelä, P.S.A.; Jokinen, K.; Himanen, K. Roles of Endogenous Glycinebetaine in Plant Abiotic Stress Responses. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants; Hossain, M., Kumar, V., Burritt, D., Fujita, M., Mäkelä, P., Eds.; Springer: Cham, Switzerland; Geneva, Switzerland, 2019; pp. 153–173. [Google Scholar]
- Mujahid, N.; Shahbaz, M.; Kiran, A.; Wahid, M.A. Modulations Induced by seed priming of strigolactone (GR24) in morpho-physiological and biochemical attributes of ajwain (Trachyspermum ammi L.) under salt stress. J. Plant Growth Regul. 2023, 42, 2893–2906. [Google Scholar] [CrossRef]
- Luqman, M.; Shahbaz, M.; Maqsood, M.F.; Farhat, F.; Zulfiqar, U.; Siddiqui, M.H.; Masood, A.; Aqeel, M.; Haider, F.U. Effect of strigolactone on growth, photosynthetic efficiency, antioxidant activity, and osmolytes accumulation in different maize (Zea mays L.) hybrids grown under drought stress. Plant Signal. Behav. 2023, 18, 2262795. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alamri, S.A.; Ali, H.M.; Alayafi, A.A. Bacillus firmus (SW5) augments salt tolerance in soybean (Glycine max L.) by modulating root system architecture, antioxidant defense systems, and stress-responsive genes expression. Plant Physiol. Biochem. 2018, 132, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Van Ha, C.; Leyva-González, M.A.; Osakabe, Y.; Tran, U.T.; Nishiyama, R.; Watanabe, Y.; Tanaka, M.; Seki, M.; Yamaguchi, S.; Van Dong, N.; et al. Positive regulatory role of strigolactone in plant responses to drought and salt stress. Proc. Natl. Acad. Sci. USA 2014, 111, 851–856. [Google Scholar]
- Kong, C.C.; Ren, C.G.; Li, R.Z.; Xie, Z.H.; Wang, J.P. Hydrogen peroxide and strigolactones signaling are involved in the alleviation of salt stress induced by arbuscular mycorrhizal fungus in Sesbania cannabina seedlings. J. Plant Growth Regul. 2017, 36, 734–742. [Google Scholar] [CrossRef]
- Gupta, A.; Rai, S.; Bano, A.; Sharma, S.; Kumar, M.; Binsuwaidan, R.; Suhail Khan, M.; Upadhyay, T.K.; Alshammari, N.; Saeed, M.; et al. ACC deaminase produced by PGPR mitigates the adverse effect of osmotic and salinity stresses in Pisum sativum through modulating the antioxidants activities. Plants 2022, 11, 3419. [Google Scholar] [CrossRef]
- Ali, F.; Bano, A.; Hassan, T.U.; Nazir, M.; Khan, R.T. Plant growth promoting rhizobacteria induced modulation of physiological responses in rice under salt and drought stresses. Pak. J. Bot. 2023, 55, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Gu, M.; Lu, J.; Li, X.E.; Liu, D.; Wang, L. Metabolomic Analysis of Key Metabolites and Their Pathways Revealed the Response of Alfalfa (Medicago sativa L.) Root Exudates to rac-GR24 under Drought Stress. Plants 2023, 12, 1163. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Tan, Z.; Zhou, Y.; Guo, S.; Sang, T.; Wang, Y.; Shu, S. Physiological mechanism of strigolactone enhancing tolerance to low light stress in cucumber seedlings. BMC Plant Biol. 2022, 22, 30. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Wang, C.; Feng, Q.; Liou, R.M.; Lin, Y.F.; Qiao, J.; Lu, Y.; Chang, Y. The mechanisms of sodium chloride stress mitigation by salt-tolerant plant growth promoting rhizobacteria in wheat. Agronomy 2022, 12, 543. [Google Scholar] [CrossRef]
- Khan, N.; Zandi, P.; Ali, S.; Mehmood, A.; Adnan Shahid, M.; Yang, J. Impact of salicylic acid and PGPR on the drought tolerance and phytoremediation potential of Helianthus annus. Front. Microbiol. 2018, 9, 2507. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Cha (mg g−1 FW) | Chb (mg g−1 FW) | Car (mg g–1 FW) | RWC (%) | EL (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Arta | Karchia | Arta | Karchia | Arta | Karchia | Arta | Karchia | Arta | Karchia | |
| C | 4.41 ± 0.15 c | 3.89 ± 0.11 bc | 3.19 ± 0.07 bcd | 3.20 ± 0.14 cd | 0.79 ± 0.045 bc | 0.81 ± 0.033 bc | 74.13 ± 1.93 d | 83.45 ± 0.92 bc | 30.40 ± 1.28 f | 22.32 ± 0.99 g |
| S1 | 2.62 ± 0.21 g | 3.17 ± 0.07 e | 2.03 ± 0.07 g | 2.70 ± 0.26 e | 0.67 ± 0.027 e | 0.70 ± 0.014 e | 69.10 ± 1.07 g | 77.64 ± 0.70 f | 42.25 ± 1.05 d | 34.41 ± 1.16 c |
| S2 | 2.08 ± 0.21 h | 2.56 ± 0.12 f | 1.14 ± 0.22 h | 1.62 ± 0.10 g | 0.42 ± 0.054 i | 0.50 ± 0.027 h | 62.39 ± 1.62 h | 71.01 ± 1.25 g | 76.14 ± 0.96 a | 59.34 ± 1.00 a |
| B | 4.44 ± 0.22 c | 4.07 ± 0.16 ab | 3.24 ± 0.12 bc | 3.32 ± 0.12 bc | 0.83 ± 0.026 ab | 0.84 ± 0.024 ab | 76.35 ± 1.07 c | 83.55 ± 0.88 bc | 27.49 ± 0.70 g | 19.63 ± 1.20 h |
| GR24 | 4.85 ± 0.07 b | 4.24 ± 0.11 a | 3.30 ± 0.24 b | 3.54 ± 0.13 b | 0.85 ± 0.013 a | 0.84 ± 0.011 ab | 80.45 ± 1.14 b | 84.27 ± 0.71 b | 16.84 ± 1.74 h | 17.54 ± 1.50 i |
| B + GR24 | 5.23 ± 0.06 a | 4.11 ± 0.11 a | 3.82 ± 0.16 a | 3.99 ± 0.12 a | 0.86 ± 0.005 a | 0.87 ± 0.017 a | 84.03 ± 0.90 a | 88.20 ± 0.79 a | 15.55 ± 0.92 h | 16.00 ± 0.33 i |
| S1 + GR24 | 3.89 ± 0.14 d | 3.73 ± 0.12 cd | 2.90 ± 0.21 de | 3.12 ± 0.10 cd | 0.74 ± 0.011 d | 0.79 ± 0.020 cd | 72.82 ± 0.74 de | 82.35 ± 0.75 cd | 32.14 ± 0.71 f | 25.49 ± 1.20 f |
| S2 + GR24 | 3.33 ± 0.10 ef | 3.04 ± 0.12 e | 2.56 ± 0.25 f | 2.27 ± 0.14 f | 0.56 ± 0.025 g | 0.55 ± 0.016 g | 70.13 ± 1.03 fg | 78.00 ± 1.20 f | 45.05 ± 1.18 c | 32.56 ± 0.90 d |
| S1 + B | 3.57 ± 0.17 e | 3.53 ± 0.13 | 2.91 ± 0.12 de | 3.04 ± 0.12 d | 0.73 ± 0.016 d | 0.77 ± 0.029 d | 73.65 ± 0.86 d | 81.77 ± 0.92 de | 35.68 ± 1.16 e | 31.52 ± 1.04 d |
| S2 + B | 3.21 ± 0.11 f | 2.70 ± 0.19 f | 2.03 ± 0.18 g | 2.06 ± 0.17 f | 0.51 ± 0.021 h | 0.53 ± 0.026 gh | 69.50 ± 0.55 g | 76.62 ± 0.91 f | 63.36 ± 1.11 b | 41.58 ± 1.34 b |
| S1 + GR24 + B | 4.17 ± 0.15 cd | 3.77 ± 0.11 c | 3.16 ± 0.07 bcd | 3.19 ± 0.07 cd | 0.75 ± 0.031 cd | 0.81 ± 0.015 bc | 76.45 ± 0.92 c | 82.82 ± 0.70 bcd | 27.80 ± 0.82 g | 21.89 ± 0.95 g |
| S2 + GR24 + B | 3.51 ± 0.39 ef | 3.19 ± 0.10 e | 3.00 ± 0.20 cde | 2.60 ± 0.11 e | 0.61 ± 0.019 f | 0.61 ± 0.014 f | 71.54 ± 1.02 ef | 80.60 ± 1.25 e | 41.68 ± 1.00 d | 27.75 ± 1.03 e |
| LSD 5% | 0.32 | 0.21 | 0.29 | 0.23 | 0.05 | 0.04 | 1.90 | 1.57 | 1.83 | 1.83 |
| Treatments | LNa+ (mg g−1 DW) | LK+ (mg g−1 DW) | LK+/Na+ (Ratio) | RNa+ (mg g−1 DW) | RK+ (mg g−1 DW) | RK+/Na+ (Ratio) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Arta | Karchia | Arta | Karchia | Arta | Karchia | Arta | Karchia | Arta | Karchia | Arta | Karchia | |
| C | 3.84 ± 0.11 ef | 2.62 ± 0.30 f | 13.54 ± 0.40 cd | 13.86 ± 0.33 def | 3.53 ± 0.13 de | 5.34 ± 0.67 c | 18.31 ± 0.07 cd | 17.50 ± 1.17 e | 9.87 ± 0.88 c | 12.18 ± 0.87 c | 0.54 ± 0.050 d | 0.70 ± 0.041 d |
| S1 | 5.60 ± 0.45 b | 4.84 ± 0.18 b | 11.91 ± 0.33 e | 12.28 ± 0.53 h | 2.14 ± 0.12 g | 2.54 ± 0.20 f | 21.33 ± 0.49 b | 20.13 ± 0.37 b | 7.28 ± 0.07 e | 9.09 ± 0.29 ef | 0.34 ± 0.009 h | 0.45 ± 0.009 g |
| S2 | 8.90 ± 0.23 a | 7.70 ± 0.13 a | 9.40 ± 0.38 f | 9.58 ± 0.77 i | 1.06 ± 0.04 h | 1.24 ± 0.09 g | 25.33 ± 1.15 a | 22.79 ± 0.73 a | 4.81 ± 0.40 g | 6.66 ± 0.42 g | 0.19 ± 0.025 i | 0.29 ± 0.013 h |
| B | 1.64 ± 0.12 h | 1.21 ± 0.11 g | 15.31 ± 0.43 b | 14.82 ± 0.17 bc | 9.38 ± 0.54 b | 12.29 ± 0.93 b | 14.94 ± 0.20 f | 16.34 ± 0.42 f | 13.22 ± 0.29 b | 12.60 ± 0.67 c | 0.88 ± 0.010 b | 0.77 ± 0.039 c |
| GR24 | 2.12 ± 0.32 g | 1.36 ± 0.04 g | 15.17 ± 0.55 b | 15.18 ± 0.13 ab | 7.25 ± 1.07 c | 11.14 ± 0.42 b | 16.25 ± 0.56 e | 13.91 ± 0.79 g | 12.61 ± 0.50 b | 13.51 ± 0.53 b | 0.78 ± 0.005 c | 0.97 ± 0.086 b |
| B + GR24 | 1.03 ± 0.06 i | 1.03 ± 0.12 g | 17.11 ± 0.22 a | 15.66 ± 0.21 a | 16.62 ± 0.92 a | 15.34 ± 1.96 a | 14.04 ± 0.10 f | 13.19 ± 0.17 g | 14.15 ± 0.27 a | 14.45 ± 0.45 a | 1.01 ± 0.022 a | 1.10 ± 0.045 a |
| S1 + GR24 | 4.21 ± 0.22 de | 3.65 ± 0.27 d | 13.37 ± 0.24 cd | 14.13 ± 0.30 de | 3.18 ± 0.21 ef | 3.89 ± 0.38 de | 19.18 ± 0.33 c | 17.84 ± 0.53 e | 8.72 ± 0.38 d | 10.67 ± 0.38 d | 0.45 ± 0.018 f | 0.60 ± 0.033 ef |
| S2 + GR24 | 4.89 ± 0.25 c | 4.40 ± 0.41 c | 12.13 ± 0.70 e | 12.64 ± 0.55 gh | 2.48 ± 0.08 fg | 2.88 ± 0.17 ef | 18.59 ± 0.64 cd | 18.90 ± 0.23 cd | 6.49 ± 0.14 f | 8.48 ± 0.48 f | 0.35 ± 0.007 h | 0.45 ± 0.029 g |
| S1 + B | 3.96 ± 0.16 ef | 3.05 ± 0.14 e | 13.74 ± 0.45 c | 13.68 ± 0.43 ef | 3.48 ± 0.18 de | 4.49 ± 0.12 cd | 18.10 ± 0.23 d | 17.97 ± 0.61 de | 9.03 ± 0.29 d | 11.17 ± 0.63 d | 0.50 ± 0.010 e | 0.62 ± 0.014 e |
| S2 + B | 4.58 ± 0.14 cd | 5.08 ± 0.14 b | 12.95 ± 0.07 d | 13.27 ± 0.23 fg | 2.83 ± 0.09 efg | 2.62 ± 0.11 f | 20.47 ± 0.74 b | 19.41 ± 0.57 bc | 7.02 ± 0.10 ef | 8.67 ± 0.41 f | 0.34 ± 0.008 h | 0.45 ± 0.032 g |
| S1 + GR24 + B | 3.68 ± 0.17 f | 2.86 ± 0.16 ef | 14.81 ± 0.34 b | 14.96 ± 0.13 bc | 4.03 ± 0.20 d | 5.23 ± 0.26 c | 17.87 ± 0.53 d | 16.21 ± 0.34 f | 9.28 ± 0.19 cd | 12.31 ± 0.33 c | 0.52 ± 0.009 de | 0.76 ± 0.029 cd |
| S2 + GR24 + B | 4.38 ± 0.15 d | 3.93 ± 0.17 d | 12.98 ± 0.05 d | 14.49 ± 0.18 cd | 2.96 ± 0.09 ef | 3.69 ± 0.11 def | 18.43 ± 0.42 cd | 17.72 ± 0.45 e | 7.09 ± 0.08 ef | 9.76 ± 0.23 e | 0.38 ± 0.006 g | 0.55 ± 0.018 f |
| LSD 5% | 0.37 | 0.37 | 0.66 | 0.64 | 0.76 | 1.15 | 0.91 | 1.00 | 0.62 | 0.85 | 0.03 | 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehrabi, S.S.; Sabokdast, M.; Bihamta, M.R.; Dedičová, B. The Coupling Effects of PGPR Inoculation and Foliar Spraying of Strigolactone in Mitigating the Negative Effect of Salt Stress in Wheat Plants: Insights from Phytochemical, Growth, and Yield Attributes. Agriculture 2024, 14, 732. https://doi.org/10.3390/agriculture14050732
Mehrabi SS, Sabokdast M, Bihamta MR, Dedičová B. The Coupling Effects of PGPR Inoculation and Foliar Spraying of Strigolactone in Mitigating the Negative Effect of Salt Stress in Wheat Plants: Insights from Phytochemical, Growth, and Yield Attributes. Agriculture. 2024; 14(5):732. https://doi.org/10.3390/agriculture14050732
Chicago/Turabian StyleMehrabi, Shadi Sadat, Manijeh Sabokdast, Mohammad Reza Bihamta, and Beáta Dedičová. 2024. "The Coupling Effects of PGPR Inoculation and Foliar Spraying of Strigolactone in Mitigating the Negative Effect of Salt Stress in Wheat Plants: Insights from Phytochemical, Growth, and Yield Attributes" Agriculture 14, no. 5: 732. https://doi.org/10.3390/agriculture14050732