
Mathematical Evaluation of Population Changes of Lactobacillus acidophilus and Bifidobacterium animalis ssp. lactis as Free and Encapsulated Cells in Butter

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Preparation of Encapsulated Cells of Probiotics
Activation of Lactobacillus acidophilus (LA5) and Bifidobacterium animal ssp. lactis (BB12)
Whey Protein Hydrolysate–Maltodextrin Solution for Spray-Drying
Drying of Whey Protein Hydrolysate–Maltodextrin-Conjugated Probiotic Solution
2.2.2. Preparation of Butter with Free Cells and Encapsulated Cells of Probiotics
Manufacturing of Butter Samples
Incorporation of Free Cells and Encapsulated Cells of Probiotics in Butter
2.3. Analysis
2.3.1. Viability of Probiotics
2.3.2. Modelling of Population Change of LA5 and BB12
2.3.3. Acid Value
2.3.4. Confocal Laser Scanning Microscopy
3. Results and Discussion
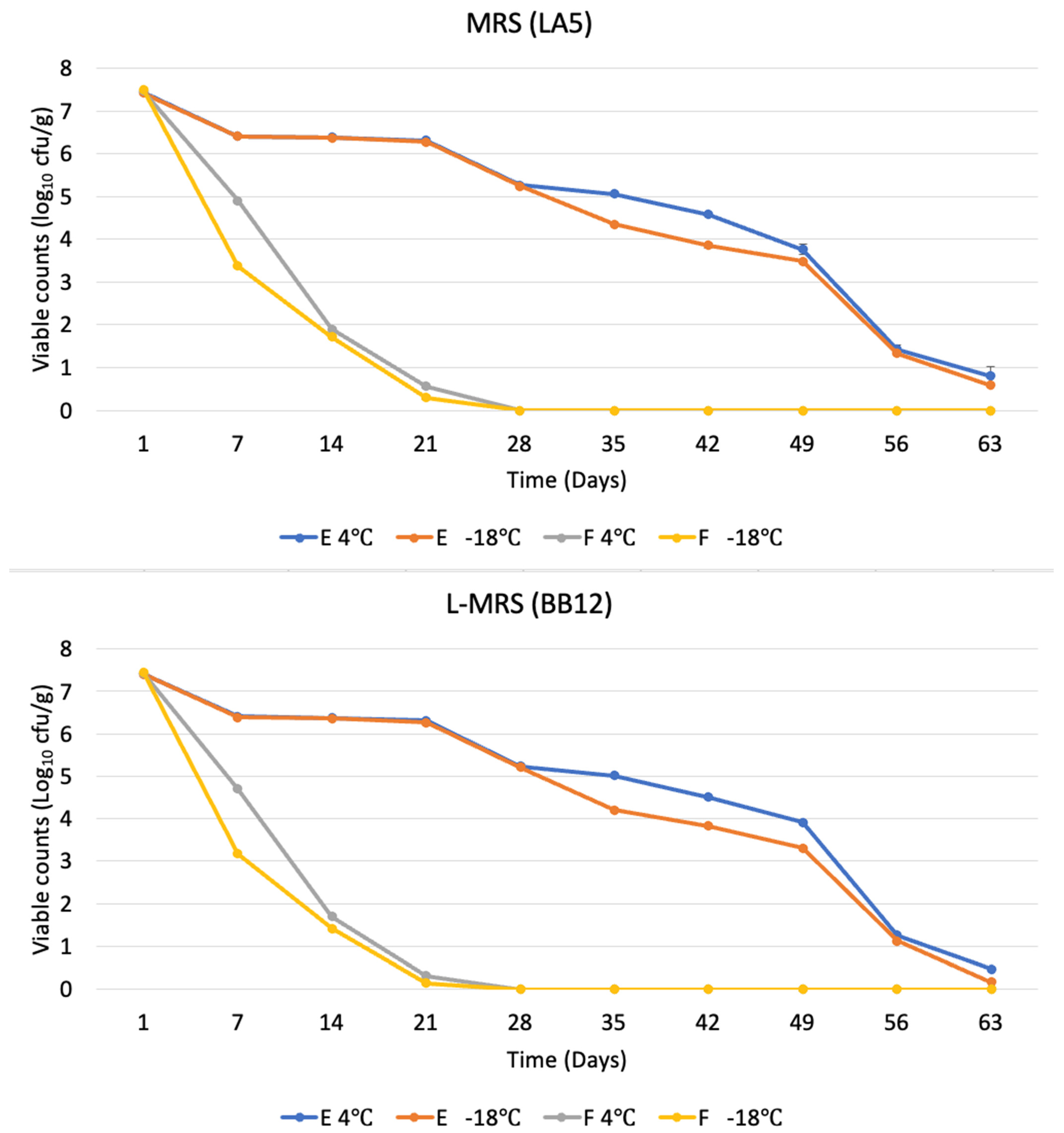
3.1. Viable Probiotic Counts of Butter
3.2. Modelling of LA5 and BB12
3.3. Acid Value of Butter
3.4. Confocal Laser Scanning Microscopy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO/WHO. Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation. Available online: https://www.fao.org/documents/card/en?details=7c102d95-2fd5-5b22-8faf-f0b2e68dfbb6/ (accessed on 4 October 2001).
- da Silva, M.N.; Tagliapietra, B.L.; dos Santos Richards, N.S.P. Encapsulation, storage viability, and consumer acceptance of probiotic butter. LWT 2021, 139, 110536. [Google Scholar] [CrossRef]
- Bellinazo, P.L.; Vitola, H.R.S.; Cruxen, C.E.d.S.; Braun, C.L.K.; Hackbart, H.C.d.S.; da Silva, W.P.; Fiorentini, Â.M. Probiotic butter: Viability of Lactobacillus casei strains and bixin antioxidant effect (Bixa orellana L.). J. Food Process. Preserv. 2019, 43, e14088. [Google Scholar] [CrossRef]
- Zacarchenco, P.B.; Spadoti, L.M.; Silva e Alves, A.T.; Zanetti, V.C.; Verruck, S. Probiotic Butter. In Probiotic Foods and Beverages: Technologies and Protocols; Springer: Berlin/Heidelberg, Germany, 2023; pp. 67–80. [Google Scholar]
- Terpou, A.; Papadaki, A.; Lappa, I.K.; Kachrimanidou, V.; Bosnea, L.A.; Kopsahelis, N. Probiotics in food systems: Significance and emerging strategies towards improved viability and delivery of enhanced beneficial value. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef] [PubMed]
- Frakolaki, G.; Giannou, V.; Kekos, D.; Tzia, C. A review of the microencapsulation techniques for the incorporation of probiotic bacteria in functional foods. Crit. Rev. Food Sci. Nutr. 2021, 61, 1515–1536. [Google Scholar] [CrossRef]
- Gaba, K.; Anand, S. Potential of Incorporating a Functional Probiotic Encapsulant in Whipped Cream. Fermentation 2023, 9, 928. [Google Scholar] [CrossRef]
- Gaba, K.; Anand, S.; Syamala, A. Development of Value-Added Butter by Incorporating Whey Protein Hydrolysate-Encapsulated Probiotics. Microorganisms 2023, 11, 1139. [Google Scholar] [CrossRef] [PubMed]
- Minj, S.; Anand, S. Development of a spray-dried conjugated whey protein hydrolysate powder with entrapped probiotics. J. Dairy Sci. 2022, 105, 2038–2048. [Google Scholar] [CrossRef]
- Ewe, J.-A.; Loo, S.-Y. Effect of cream fermentation on microbiological, physicochemical and rheological properties of L. helveticus-butter. Food Chem. 2016, 201, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Rønholt, S.; Kirkensgaard, J.J.K.; Pedersen, T.B.; Mortensen, K.; Knudsen, J.C. Polymorphism, microstructure and rheology of butter. Effects of cream heat treatment. Food Chem. 2012, 135, 1730–1739. [Google Scholar] [CrossRef]
- Yilmaz-Ersan, L.; Ozcan, T.; Akpinar-Bayizit, A.; Turan, M.A.; Taskin, M.B. Probiotic cream: Viability of probiotic bacteria and chemical characterization. J. Food Process. Preser. 2017, 41, e12797. [Google Scholar] [CrossRef]
- Olszewska, M.; Staniewski, B.; Łaniewska-Trokenheim, Ł. Cell viability of Bifidobacterium lactis strain in long-term storage butter assessed with the plate count and fluorescence techniques. Czech J. Food Sci. 2012, 30, 421–428. [Google Scholar] [CrossRef]
- Dianawati, D.; Mishra, V.; Shah, N.P. Survival of Bifidobacterium longum 1941 microencapsulated with proteins and sugars after freezing and freeze drying. Food Res. Int. 2013, 51, 503–509. [Google Scholar] [CrossRef]
- Zanjani, M.A.K.; Ehsani, M.R.; Tarzi, B.G.; Sharifan, A. Promoting probiotics survival by microencapsualtion with Hylon starch and genipin cross-linked coatings in simulated gastro-intestinal condition and heat treatment. Iran. J. Pharm. Res. IJPR 2018, 17, 753. [Google Scholar]
- Ming, L.C.; Rahim, R.A.; Wan, H.Y.; Ariff, A.B. Formulation of protective agents for improvement of Lactobacillus salivarius I 24 survival rate subjected to freeze drying for production of live cells in powderized form. Food Bioprocess Techno. 2009, 2, 431–436. [Google Scholar] [CrossRef]
- Ugwu, S.O.; Apte, S.P. The effect of buffers on protein conformational stability. Pharm. Technol. 2004, 28, 86–109. [Google Scholar]
- Menezes, M.F.D.S.C.D.; Silva, T.M.D.; Etchepare, M.D.A.; Fonseca, B.D.S.; Sonza, V.P.; Codevilla, C.F.; Barin, J.S.; Silva, C.D.B.D.; Menezes, C.R.d. Improvement of the viability of probiotics (Lactobacillus acidophilus) by multilayer encapsulation. Ciênc. Rural 2019, 49, e20181020. [Google Scholar] [CrossRef]
- Feng, K.; Huang, R.-M.; Wu, R.-Q.; Wei, Y.-S.; Zong, M.-H.; Linhardt, R.J.; Wu, H. A novel route for double-layered encapsulation of probiotics with improved viability under adverse conditions. Food Chem. 2020, 310, 125977. [Google Scholar] [CrossRef]
- Yao, M.; Li, B.; Ye, H.; Huang, W.; Luo, Q.; Xiao, H.; McClements, D.J.; Li, L. Enhanced viability of probiotics (Pediococcus pentosaceus Li05) by encapsulation in microgels doped with inorganic nanoparticles. Food Hydrocoll. 2018, 83, 246–252. [Google Scholar] [CrossRef]
- Martín, M.J.; Lara-Villoslada, F.; Ruiz, M.A.; Morales, M.E. Microencapsulation of bacteria: A review of different technologies and their impact on the probiotic effects. Innov. Food Sci. Emerg. Technol. 2015, 27, 15–25. [Google Scholar] [CrossRef]
- Huang, X.; Gänzle, M.; Zhang, H.; Zhao, M.; Fang, Y.; Nishinari, K. Microencapsulation of probiotic lactobacilli with shellac as moisture barrier and to allow controlled release. J. Sci. Food Agric. 2021, 101, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Özer, B.; Kirmaci, H.A.; Şenel, E.; Atamer, M.; Hayaloğlu, A. Improving the viability of Bifidobacterium bifidum BB-12 and Lactobacillus acidophilus LA-5 in white-brined cheese by microencapsulation. Int. Dairy J. 2009, 19, 22–29. [Google Scholar] [CrossRef]
- Erkaya, T.; Ürkek, B.; Doğru, Ü.; Çetin, B.; Şengül, M. Probiotic butter: Stability, free fatty acid composition and some quality parameters during refrigerated storage. Int. Dairy J. 2015, 49, 102–110. [Google Scholar] [CrossRef]
- Nunes, G.L.; Motta, M.H.; Cichoski, A.J.; Wagner, R.; Muller, É.I.; Codevilla, C.F.; Silva, C.d.B.d.; Menezes, C.R.d. Encapsulation of Lactobacillus acidophilus La-5 and Bifidobacterium Bb-12 by spray drying and evaluation of its resistance in simulated gastrointestinal conditions, thermal treatments and storage conditions. Ciênc. Rural 2018, 48, e20180035. [Google Scholar] [CrossRef]
- Holkem, A.T.; Robichaud, V.; Favaro-Trindade, C.S.; Lacroix, M. Chemopreventive properties of extracts obtained from blueberry (Vaccinium myrtillus L.) and jabuticaba (Myrciaria cauliflora Berg.) in combination with probiotics. Nutr. Cancer 2021, 73, 671–685. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Giri, S.K. Probiotic functional foods: Survival of probiotics during processing and storage. J. Funct. Foods 2014, 9, 225–241. [Google Scholar] [CrossRef]
- Boylston, T.D.; Vinderola, C.G.; Ghoddusi, H.B.; Reinheimer, J.A. Incorporation of bifidobacteria into cheeses: Challenges and rewards. Int. Dairy J. 2004, 14, 375–387. [Google Scholar] [CrossRef]
- Chapman, C.; Gibson, G.R.; Rowland, I. Health benefits of probiotics: Are mixtures more effective than single strains? Eur. J. Nutr. 2011, 50, 1–17. [Google Scholar] [CrossRef]
- Alur, M.a.; Grecz, N. Mechanism of injury of Escherichiacoli by freezing and thawing. Biochem. Biophys. Res. Commun. 1975, 62, 308–312. [Google Scholar] [CrossRef]
- El-Kest, S.E.; Marth, E.H. Freezing of Listeria monocytogenes and other microorganisms: A review. J. Food Prot. 1992, 55, 639–648. [Google Scholar] [CrossRef]
- Djira, G.; Hasler, M.; Gerhard, D.; Segbehoe, L.; Schaarschmidt, F. Mratios: Ratios of coefficients in the general linear model. R Package Vers. 2020, 1, 1–46. [Google Scholar]
- Koczoń, P.; Gruczyńska, E.; Kowalski, B. Changes in the acid value of butter during storage at different temperatures as assessed by standard methods or by FT-IR spectroscopy. Am. J. Food Technol. 2008, 3, 154–163. [Google Scholar] [CrossRef]
- Dinçer, E.; Kıvanç, M. Lipolytic activity of lactic acid bacteria isolated from Turkish pastırma. Anadolu Univ. J. Sci. Technol. C-Life Sci. Biotechnol. 2018, 7, 12–19. [Google Scholar] [CrossRef]
- Buldo, P.; Wiking, L. The role of mixing temperature on microstructure and rheological properties of butter blends. J. Am. Oil Chem. Soc. 2012, 89, 787–795. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Acid Value | ||||||
|---|---|---|---|---|---|---|
| Control | Free Cell | Encapsulated Cells | ||||
| Days | 4 °C | −18 °C | 4 °C | −18 °C | 4 °C | −18 °C |
| 1 | 1.246 ± 0.010 Aa | 1.253 ± 0.009 Aa | 1.262 ± 0.008 Aa | 1.287 ± 0.003 Aa | 1.264 ± 0.014 Aa | 1.271 ± 0.007 Aa |
| 28 | 1.256 ± 0.010 Aa | 1.256 ± 0.010 Aa | 1.279 ± 0.009 Aa | 1.294 ± 0.007 Aa | 1.267 ± 0.011 Aa | 1.277 ± 0.005 Aa |
| 56 | 1.294 ± 0.015 Aa | 1.254 ± 0.015 Aa | 1.304 ± 0.012 Aa | 1.275 ± 0.012 Aa | 1.273 ± 0.012 Aa | 1.275 ± 0.013 Aa |
| 84 | 1.306 ± 0.014 Aa | 1.263 ± 0.014 Aa | 1.313 ± 0.012 Aa | 1.284 ± 0.010 Aa | 1.282 ± 0.014 Aa | 1.286 ± 0.013 Aa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaushik, R.; Gaba, K.; Anand, S.; Djira, G. Mathematical Evaluation of Population Changes of Lactobacillus acidophilus and Bifidobacterium animalis ssp. lactis as Free and Encapsulated Cells in Butter. Fermentation 2024, 10, 245. https://doi.org/10.3390/fermentation10050245
Kaushik R, Gaba K, Anand S, Djira G. Mathematical Evaluation of Population Changes of Lactobacillus acidophilus and Bifidobacterium animalis ssp. lactis as Free and Encapsulated Cells in Butter. Fermentation. 2024; 10(5):245. https://doi.org/10.3390/fermentation10050245
Chicago/Turabian StyleKaushik, Rakesh, Kritika Gaba, Sanjeev Anand, and Gemechis Djira. 2024. "Mathematical Evaluation of Population Changes of Lactobacillus acidophilus and Bifidobacterium animalis ssp. lactis as Free and Encapsulated Cells in Butter" Fermentation 10, no. 5: 245. https://doi.org/10.3390/fermentation10050245