

TRPV1-Dependent Antiproliferative Activity of Dioecious Maclura pomifera Extracts in Estrogen Receptor-Positive Breast Cancer Cell Lines Involves Multiple Apoptotic Pathways

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Antiproliferative Activities of Male and Female M. pomifera in MCF-7 and T47D Cells
2.2. Antiproliferative Activities of Osajin and Pomiferin in MCF-7 and T47D Cells
2.3. Male and Female M. pomifera Extracts Induce Apoptosis in MCF-7 and T47D Cells
2.4. Blocking TRPV1 Activity Increased MCF-7 and T47D Cell Viability
2.5. Calcium Chelation Increases MCF-7 and T47D Cell Viability
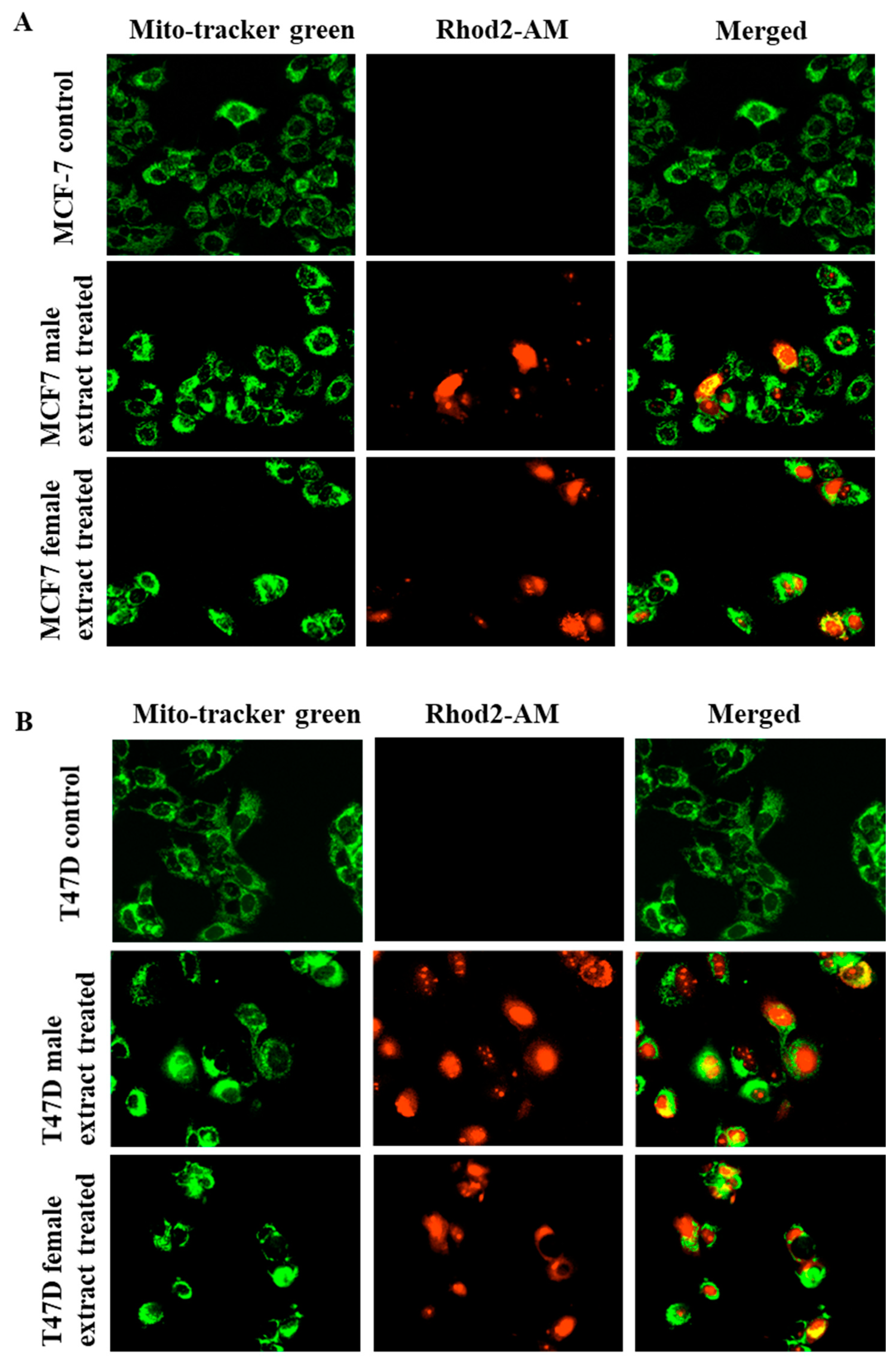
2.6. Male and Female M. pomifera Extracts Induce Mitochondrial Calcium Overload in MCF-7 and T47D Cells
2.7. Male and Female M. pomifera Extracts Induce Mitochondrial Intrinsic and Extrinsic Apoptotic Signaling Pathways in MCF-7 and T47D Cells
2.8. Male and Female M. pomifera Extracts Induce ERK-Dependent Apoptosis in MCF-7 and T47D Cells
2.9. Male and Female M. pomifera Extracts Induce Endoplasmic Reticulum-Dependent Apoptosis in MCF-7 and T47D Cells
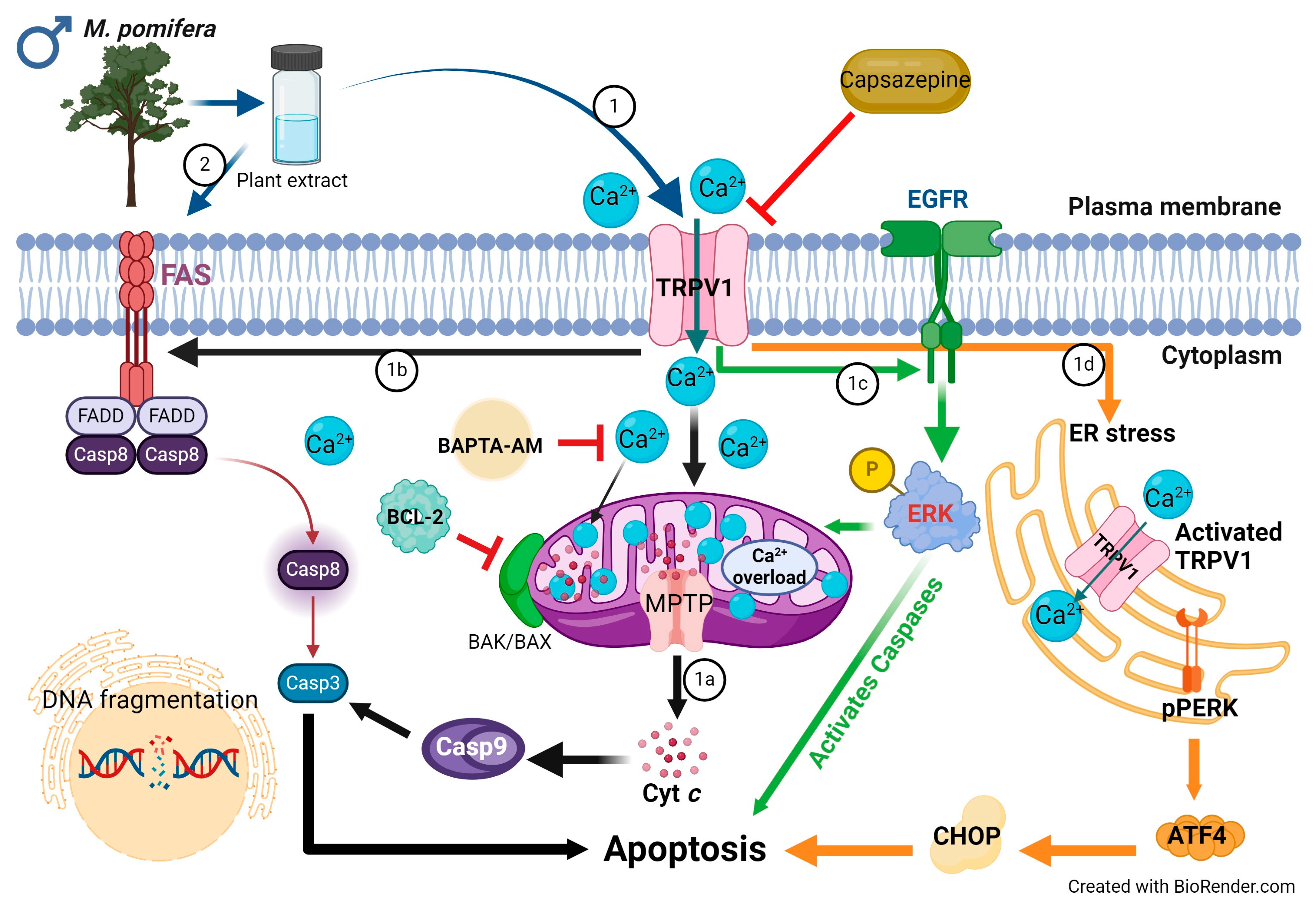
3. Discussion
4. Materials and Methods
4.1. Plant Collection and Preparation of Extracts
4.2. Cell Lines and Cell Culture Conditions
4.3. Cell Culture Treatments
4.4. Antiproliferative Assays
4.5. IC50 Estimation
4.6. Visualization of Cytotoxic Effects
4.7. Detection of Apoptosis
4.8. Capsazepine Treatments to Block TRPV1
4.9. Calcium Chelation
4.10. Visualization of Mitochondrial Calcium
4.11. Caspase 3 Activation Assay
4.12. Western Blotting
4.13. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wilkinson, L.; Gathani, T. Understanding breast cancer as a global health concern. Brit. J. Radiol. 2022, 95, 20211033. [Google Scholar] [CrossRef] [PubMed]
- Giaquinto, A.N.; Sung, H.; Miller, K.D.; Kramer, J.L.; Newman, L.A.; Minihan, A.; Jemal, A.; Siegel, R.L. Breast cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 524–541. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Chikara, S.; Nagaprashantha, L.D.; Singhal, J.; Horne, D.; Awasthi, S.; Singhal, S.S. Oxidative stress and dietary phytochemicals: Role in cancer chemoprevention and treatment. Cancer Lett. 2018, 413, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Pucci, C.; Martinelli, C.; Ciofani, G. Innovative approaches for cancer treatment: Current perspectives and new challenges. Ecancermedicalscience 2019, 13, 961. [Google Scholar] [CrossRef]
- Liu, Y.-Q.; Wang, X.-L.; He, D.-H.; Cheng, Y.-X. Protection against chemotherapy-and radiotherapy-induced side effects: A review based on the mechanisms and therapeutic opportunities of phytochemicals. Phytomedicine 2021, 80, 153402. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, A.P.A.; Conte-Junior, C.A. Health benefits of phytochemicals from Brazilian native foods and plants: Antioxidant, antimicrobial, anti-cancer, and risk factors of metabolic/endocrine disorders control. Trends Food Sci. Technol. 2021, 111, 534–548. [Google Scholar] [CrossRef]
- Basu, P.; Maier, C. Phytoestrogens and breast cancer: In vitro anti-cancer activities of isoflavones, lignans, coumestans, stilbenes and their analogs and derivatives. Biomed. Pharmacother. 2018, 107, 1648–1666. [Google Scholar] [CrossRef] [PubMed]
- Alibakhshi, A.; Malekzadeh, R.; Hosseini, S.A.; Yaghoobi, H. Investigation of the therapeutic role of native plant compounds against colorectal cancer based on system biology and virtual screening. Sci. Rep. 2023, 13, 11451. [Google Scholar] [CrossRef]
- Khan, M.I.; Bouyahya, A.; Hachlafi, N.E.; Menyiy, N.E.; Akram, M.; Sultana, S.; Zengin, G.; Ponomareva, L.; Shariati, A.; Ojo, A.; et al. Anti-cancer properties of medicinal plants and their bioactive compounds against breast cancer: A review on recent investigations. Environ. Sci. Pollut. Res. 2022, 29, 24411–24444. [Google Scholar] [CrossRef]
- Shrihastini, V.; Muthuramalingam, P.; Adarshan, S.; Sujitha, M.; Chen, J.T.; Shin, H.; Ramesh, M. Plant derived bioactive compounds, their anti-cancer effects and in silico approaches as an alternative target treatment strategy for breast cancer: An updated overview. Cancers 2021, 13, 6222. [Google Scholar] [CrossRef] [PubMed]
- Ferro, M.L. A cultural and entomological review of the Osage orange (Maclura pomifera (Raf.) Schneid.) (Moraceae) and the origin and early spread of “hedge apple” folklore. Southeast. Nat. 2014, 13, 1–34. [Google Scholar] [CrossRef]
- Gajić, I.; Dinić, A.; Stanojević, L.; Zvezdanović, J.; Nikolić, V.; Urošević, M.; Savić, V. Osage orange (Maclura pomifera (Raf.) Schneid) fruit extracts: UHPLC-DAD-ESI-MS/MS analysis, antioxidant activity and in vivo skin tests. Nat. Prod. Res. 2023, 1–6. [Google Scholar]
- Kupeli, E.; Orhan, I.; Toker, G.; Yesilada, E. Anti-inflammatory and antinociceptive potential of Maclura pomifera (Rafin.) Schneider fruit extracts and its major isoflavonoids, scandenone and auriculasin. J. Ethnopharmacol. 2006, 107, 169–174. [Google Scholar] [CrossRef]
- Yang, R.; Hanwell, H.; Zhang, J.; Tsao, R.; Meckling, K.A. Antiproliferative activity of pomiferin in normal (MCF-10A) and transformed (MCF-7) breast epithelial cells. J. Agric. Food Chem. 2011, 59, 13328–13336. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Yao, C.; Chen, X.; Xia, H.; Zhang, L.; Liu, H.; Jiang, X.; Dai, Y.; Liu, J. The fruits of Maclura pomifera extracts inhibits glioma stem-like cell growth and invasion. Neurochem. Res. 2013, 38, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.; Orazbekov, Y.; Srivedavyasasri, R.; Mombekov, S.; Datkhayev, U.; Makhatov, B. Isolation and biological evaluation of prenylated flavonoids from Maclura pomifera. Planta Med. 2017, 4, Mo-PO-61. [Google Scholar]
- Xu, S.; Cheng, X.; Wu, L.; Zheng, J.; Wang, X.; Wu, J.; Yu, H.; Bao, J.; Zhang, L. Capsaicin induces mitochondrial dysfunction and apoptosis in anaplastic thyroid carcinoma cells via TRPV1-mediated mitochondrial calcium overload. Cell. Signal. 2020, 75, 109733. [Google Scholar] [CrossRef] [PubMed]
- Tajada, S.; Villalobos, C. Calcium permeable channels in cancer hallmarks. Front. Pharmacol. 2020, 11, 968. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Ma, Y.; Ye, X.; Zhang, N.; Pan, L.; Wang, B. TRP (transient receptor potential) ion channel family: Structures, biological functions and therapeutic interventions for diseases. Signal Transduct Target Ther. 2023, 8, 261. [Google Scholar]
- Calixto, J.B.; Kassuya, C.A.; André, E.; Ferreira, J. Contribution of natural products to the discovery of the transient receptor potential (TRP) channels family and their functions. Pharmacol. Ther. 2005, 106, 179–208. [Google Scholar] [CrossRef]
- Rosenberger, D.C.; Binzen, U.; Treede, R.-D.; Greffrath, W. The capsaicin receptor TRPV1 is the first line defense protecting from acute non damaging heat: A translational approach. J. Transl. Med. 2020, 18, 28. [Google Scholar] [CrossRef] [PubMed]
- Bao, Z.; Dai, X.; Wang, P.; Tao, Y.; Chai, D. Capsaicin induces cytotoxicity in human osteosarcoma MG63 cells through TRPV1-dependent and-independent pathways. Cell Cycle 2019, 18, 1379–1392. [Google Scholar] [CrossRef] [PubMed]
- Asadi-Samani, M.; Rafieian-Kopaei, M.; Lorigooini, Z.; Shirzad, H. The effect of Euphorbia szovitsii Fisch. & CA Mey extract on the viability and the proliferation of MDA-MB-231 cell line. Biosci. Rep. 2019, 39, BSR20181538. [Google Scholar]
- Zhai, K.; Liskova, A.; Kubatka, P.; Büsselberg, D. Calcium entry through TRPV1: A potential target for the regulation of proliferation and apoptosis in cancerous and healthy cells. Int. J. Mol. Sci. 2020, 21, 4177. [Google Scholar] [CrossRef] [PubMed]
- Bujak, J.K.; Kosmala, D.; Szopa, I.M.; Majchrzak, K.; Bednarczyk, P. Inflammation, cancer and immunity—Implication of TRPV1 channel. Front Oncol. 2019, 9, 1087. [Google Scholar] [CrossRef]
- Arnold, M.; Morgan, E.; Rumgay, H.; Mafra, A.; Singh, D.; Laversanne, M.; Vignat, J.; Garlow, J.; Cardoso, F.; Siesling, S.; et al. Current and future burden of breast cancer: Global statistics for 2020 and 2040. Breast J. 2022, 66, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Khan, M.I.A.; Ayesha, S.; Zahid, Z.; Younis, M.; Kashan, M.; Khan, G.; Tahir, M.; Khan, S.; Kamal, M.; et al. Screening of phytochemicals and anti-cancer potential of Maclura pomifera leaves extract. Pak. J. Med. Sci. 2023, 17, 468. [Google Scholar]
- Maier, C.G.-A.; Chapman, K.D.; Smith, D.W. Differential estrogenic activities of male and female plant extracts from two dioecious species. Plant Sci. 1995, 109, 31–43. [Google Scholar] [CrossRef]
- Nguyen, M.; Osipo, C. Targeting breast cancer stem cells using naturally occurring phytoestrogens. Int. J. Mol. Sci. 2022, 23, 6813. [Google Scholar] [CrossRef]
- Maggiolini, M.; Bonofiglio, D.; Marsico, S.; Panno, M.L.; Cenni, B.; Picard, D.; Andò, S. Estrogen receptor α mediates the proliferative but not the cytotoxic dose-dependent effects of two major phytoestrogens on human breast cancer cells. Mol. Pharmacol. 2001, 60, 595–602. [Google Scholar]
- Seo, H.-S.; DeNardo, D.G.; Jacquot, Y.; Laïos, I.; Vidal, D.S.; Zambrana, C.R.; Leclercq, G.; Brown, P. Stimulatory effect of genistein and apigenin on the growth of breast cancer cells correlates with their ability to activate ER alpha. Breast Cancer Res. Treat. 2006, 99, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Yu, X.; Chen, X.; Zhong, H.; Liang, C.; Xu, X.; Xu, W.; Cheng, Y.; Wang, W.; Yu, L.; et al. Individual factors define the overall effects of dietary genistein exposure on breast cancer patients. Nutr. Res. 2019, 67, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.H.; Fultz, J.; Allred, K.F.; Doerge, D.R.; Helferich, W.G. Effects of dietary daidzein and its metabolite, equol, at physiological concentrations on the growth of estrogen-dependent human breast cancer (MCF-7) tumors implanted in ovariectomized athymic mice. Carcinogenesis 2006, 27, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Lecomte, S.; Demay, F.; Ferrière, F.; Pakdel, F. Phytochemicals targeting estrogen receptors: Beneficial rather than adverse effects? Int. J. Mol. Sci. 2017, 18, 1381. [Google Scholar] [CrossRef] [PubMed]
- Merlin, J.P.; Rupasinghe, H.P.; Dellaire, G.; Murphy, K. Role of dietary antioxidants in p53-mediated cancer chemoprevention and tumor suppression. Oxidative Med. Cell. Longev. 2021, 2021, 9924328. [Google Scholar] [CrossRef] [PubMed]
- Erden, Y. Sour black mulberry (Morus nigra L.) causes cell death by decreasing mutant p53 expression in HT-29 human colon cancer cells. Food Biosci. 2021, 42, 101113. [Google Scholar] [CrossRef]
- Bolanz, K.A.; Hediger, M.A.; Landowski, C.P. The role of TRPV6 in breast carcinogenesis. Mol Cancer Ther. 2008, 7, 271–279. [Google Scholar] [CrossRef]
- Schultz, J.R.; Petz, L.N.; Nardulli, A.M. Estrogen receptor alpha and Sp1 regulate progesterone receptor gene expression. Mol. Cell. Endocrinol. 2003, 201, 165–175. [Google Scholar] [CrossRef]
- Monteith, G.R.; McAndrew, D.; Faddy, H.M.; Roberts-Thomson, S.J. Calcium and cancer: Targeting Ca2+ transport. Nat. Rev. Cancer 2007, 7, 519–530. [Google Scholar] [CrossRef]
- Varghese, E.; Samuel, S.M.; Sadiq, Z.; Kubatka, P.; Liskova, A.; Benacka, J.; Pazinka, P.; Kruzliak, P.; Büsselberg, D. Anti-cancer agents in proliferation and cell death: The calcium connection. Int. J. Mol. Sci. 2019, 20, 3017. [Google Scholar] [CrossRef]
- Lau, J.K.; Brown, K.C.; Dom, A.M.; Witte, T.R.; Thornhill, B.A.; Crabtree, C.M.; Perry, H.E.; Brown, M.; Ball, J.G.; Creel, R.G.; et al. Capsaicin induces apoptosis in human small cell lung cancer via the TRPV6 receptor and the calpain pathway. Apoptosis 2014, 19, 1190–1201. [Google Scholar] [CrossRef]
- Stueber, T.; Eberhardt, M.J.; Caspi, Y.; Lev, S.; Binshtok, A.; Leffler, A. Differential cytotoxicity and intracellular calcium-signalling following activation of the calcium-permeable ion channels TRPV1 and TRPA1. Cell Calcium 2017, 68, 34–44. [Google Scholar] [CrossRef]
- Tojkander, S.S. TRPV family ion channels in the mammary epithelium: Role in normal tissue homeostasis and along breast cancer progression. In Ion Transporters-From Basic Properties to Medical Treatment; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Weber, L.V.; Al-Refae, K.; Wolk, G.; Bonatz, G.; Altmuller, J.; Becker, C.; Gisselman, G.; Hatt, H. Expression and functionality of TRPV1 in breast cancer cells. Breast Cancer 2016, 8, 243–252. [Google Scholar] [CrossRef]
- Wu, T.T.; Peters, A.A.; Tan, P.T.; Roberts-Thomson, S.J.; Monteith, G.R. Consequences of activating the calcium permeable ion channel TRPV1 in breast cancer cells with regulated TRPV1 expression. Cell Calcium 2014, 56, 59–67. [Google Scholar] [CrossRef]
- Nazıroğlu, M.; Çiğ, B.; Blum, W.; Vizler, C.; Buhala, A.; Marton, A.; Katona, R.; Josvay, K.; Schwaller, B.; Olah, Z.; et al. Targeting breast cancer cells by MRS1477, a positive allosteric modulator of TRPV1 channels. PLoS ONE 2017, 12, e0179950. [Google Scholar] [CrossRef]
- Marchi, S.; Patergnani, S.; Missiroli, S.; Morciano, G.; Rimessi, A.; Wieckowski, M.R.; Giorgi, C.; Pinton, P. Mitochondrial and endoplasmic reticulum calcium homeostasis and cell death. Cell Calcium 2018, 69, 62–72. [Google Scholar] [CrossRef]
- Basu, A. The interplay between apoptosis and cellular senescence: Bcl-2 family proteins as targets for cancer therapy. Pharmacol. Ther. 2022, 230, 107943. [Google Scholar] [CrossRef]
- Alkhalaf, M.; El-Mowafy, A.; Renno, W.; Rachid, O.; Ali, A.; Al-Attyiah, R. Resveratrol-induced apoptosis in human breast cancer cells is mediated primarily through the caspase-3-dependent pathway. Arch. Med. Res. 2008, 39, 162–168. [Google Scholar] [CrossRef]
- Antonsson, B.; Montessuit, S.; Sanchez, B.; Martinou, J.-C. Bax is present as a high molecular weight oligomer/complex in the mitochondrial membrane of apoptotic cells. J. Biol. Chem. 2001, 276, 11615–11623. [Google Scholar] [CrossRef]
- Kim, S.; Kang, C.; Shin, C.Y.; Hwang, S.W.; Yang, Y.D.; Shim, W.S.; Park, M.; Kim, E.; Kim, M.; Kim, B.; et al. TRPV1 recapitulates native capsaicin receptor in sensory neurons in association with Fas-associated factor 1. J. Neurosci. 2006, 26, 2403–2412. [Google Scholar] [CrossRef]
- Abd El-Hafeez, A.A.; Marzouk, H.M.M.; Abdelhamid, M.A.; Khalifa, H.O.; Hasanin, T.H.; Habib, A.G.; Pack, S.P. Anti-cancer effect of Hyoscyamus muticus extract via its activation of Fas/FasL-ASK1-p38 pathway. Biotechnol. Bioprocess Eng. 2022, 27, 833–845. [Google Scholar] [CrossRef]
- Wozniak, A.L.; Wang, X.; Stieren, E.S.; Scarbrough, S.G.; Elferink, C.J.; Boehning, D. Requirement of biphasic calcium release from the endoplasmic reticulum for Fas-mediated apoptosis. J. Cell Biol. 2006, 175, 709–714. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Z.; Capo-Aponte, J.; Zhang, F.; Pan, Z.; Reinach, P. Epidermal growth factor receptor transactivation by the cannabinoid receptor (CB1) and transient receptor potential vanilloid 1 (TRPV1) induces differential responses in corneal epithelial cells. Exp. Eye Res. 2010, 91, 462–471. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, Y.; Dong, L.; Qu, X.Y.; Tao, L.N.; Zhang, Y.M.; Zhai, J.-H.; Song, Y.-Q. Triptolide induces autophagy and apoptosis through ERK activation in human breast cancer MCF 7 cells. Exp. Ther. Med. 2018, 15, 3413–3419. [Google Scholar] [CrossRef]
- Zhao, R.; Tsang, S.Y. Versatile roles of intracellularly located TRPV1 channel. J. Cell. Physiol. 2017, 232, 1957–1965. [Google Scholar] [CrossRef]
- Thomas, K.C.; Sabnis, A.S.; Johansen, M.E.; Lanza, D.L.; Moos, P.J.; Yost, G.S.; Reilly, C.A. Transient receptor potential vanilloid 1 agonists cause endoplasmic reticulum stress and cell death in human lung cells. J. Pharmacol. Exp. Ther. 2007, 321, 830–838. [Google Scholar] [CrossRef]
- Tsai, T.-C.; Lai, K.-H.; Su, J.-H.; Wu, Y.-J.; Sheu, J.-H. 7-Acetylsinumaximol B induces apoptosis and autophagy in human gastric carcinoma cells through mitochondria dysfunction and activation of the PERK/eIF2α/ATF4/CHOP signaling pathway. Mar. Drugs. 2018, 16, 104. [Google Scholar] [CrossRef]
- He, Y.; Sun, M.M.; Zhang, G.G.; Yang, J.; Chen, K.S.; Xu, W.W.; Li, B. Targeting PI3K/Akt signal transduction for cancer therapy. Signal Transduct. Target. Ther. 2021, 6, 425. [Google Scholar] [CrossRef]
- Yang, J.; Nie, J.; Ma, X.; Wei, Y.; Peng, Y.; Wei, X. Targeting PI3K in cancer: Mechanisms and advances in clinical trials. Mol. Cancer 2019, 18, 26. [Google Scholar] [CrossRef]
- Almilaibary, A. Phyto-therapeutics as anti-cancer agents in breast cancer: Pathway targeting and mechanistic elucidation. Saudi J. Biol. Sci. 2024, 31, 103935. [Google Scholar] [CrossRef]
- Vercelli, C.; Barbero, R.; Cuniberti, B.; Racca, S.; Abbadessa, G. Transient receptor potential vanilloid 1 expression and functionality in mcf-7 cells: A preliminary investigation. J. Breast Cancer 2014, 17, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Basu, P.; Tongkhuya, S.A.; Harris, T.L.; Riley, A.R.; Maier, C.; Granger, J.; Wojtaszek, J.; Averitt, D.L. Euphorbia bicolor (Euphorbiaceae) latex phytochemicals induce long-lasting non-opioid peripheral analgesia in a rat model of inflammatory pain. Front. Pharmacol. 2019, 10, 958. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rumpa, M.M.; Maier, C. TRPV1-Dependent Antiproliferative Activity of Dioecious Maclura pomifera Extracts in Estrogen Receptor-Positive Breast Cancer Cell Lines Involves Multiple Apoptotic Pathways. Int. J. Mol. Sci. 2024, 25, 5258. https://doi.org/10.3390/ijms25105258
Rumpa MM, Maier C. TRPV1-Dependent Antiproliferative Activity of Dioecious Maclura pomifera Extracts in Estrogen Receptor-Positive Breast Cancer Cell Lines Involves Multiple Apoptotic Pathways. International Journal of Molecular Sciences. 2024; 25(10):5258. https://doi.org/10.3390/ijms25105258
Chicago/Turabian StyleRumpa, Mafia Mahabub, and Camelia Maier. 2024. "TRPV1-Dependent Antiproliferative Activity of Dioecious Maclura pomifera Extracts in Estrogen Receptor-Positive Breast Cancer Cell Lines Involves Multiple Apoptotic Pathways" International Journal of Molecular Sciences 25, no. 10: 5258. https://doi.org/10.3390/ijms25105258