
Study of the Impact of the Parasitic Microalgae Coccomyxa parasitica on the Health of Bivalve Modiolus kurilensis


Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Animals
2.2. Examination of M. kurilensis
2.3. Hemolymph Sampling
2.4. Analysis of Cellular Parameters of Hemolymph
2.4.1. Haemocyte Count and Their Viability
2.4.2. May–Gruenwald Typing of Cell Populations
2.4.3. Phagocytosis Assay
2.4.4. Estimation of Reactive Oxygen Species of Hemocytes and Content of Lysosomes
2.5. Analysis of Humoral Parameters of Hemolymph
2.5.1. Hemagglutination Reaction
2.5.2. Degree of Hemolysis
2.5.3. Biochemical Analysis
2.6. Determination of the Degree of Infection
2.7. Histological Examination
2.8. Data Analysis
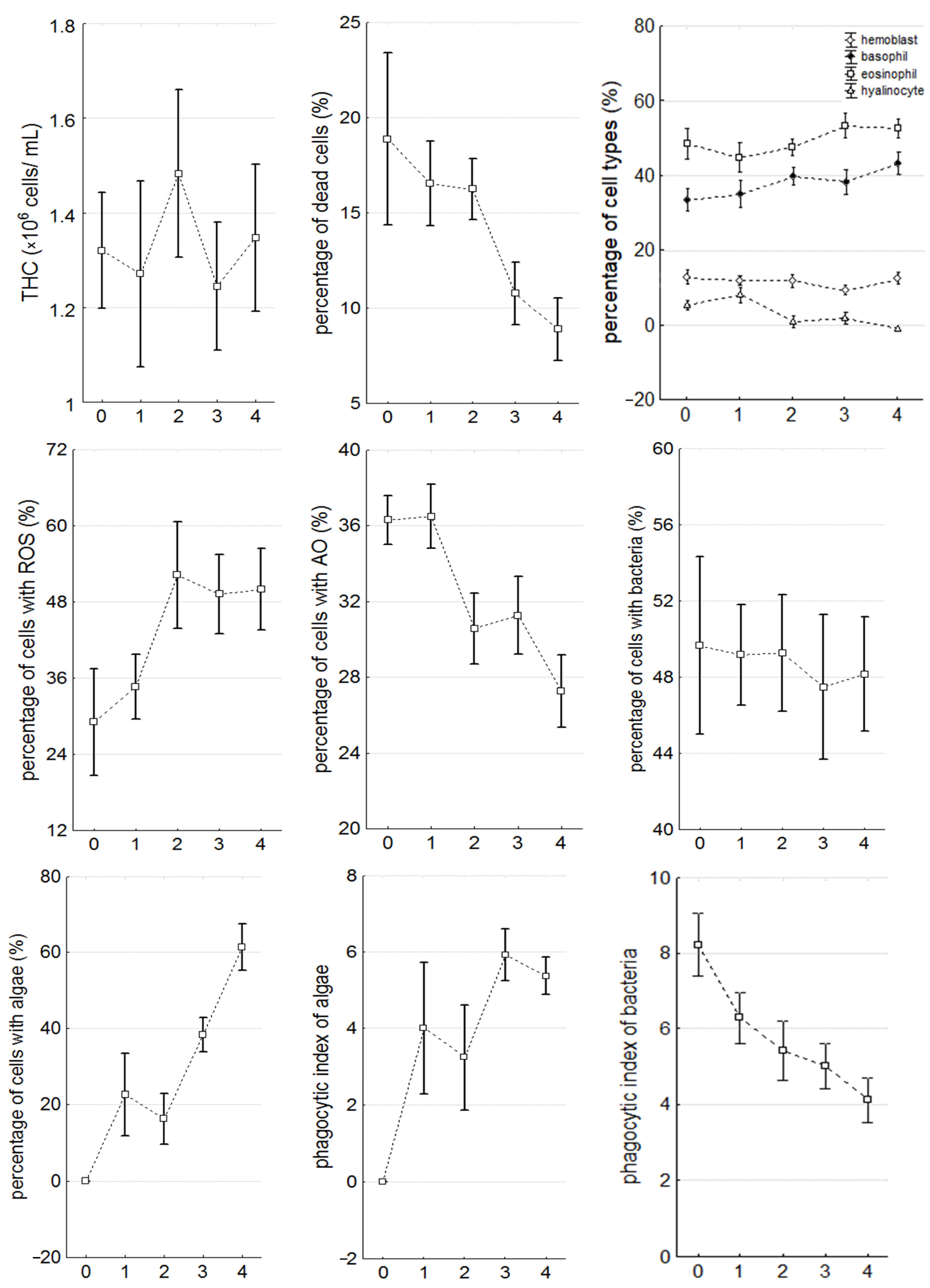
3. Results
3.1. Stages of Infection Caused by C. parasitica
3.2. Morphometric Characteristics
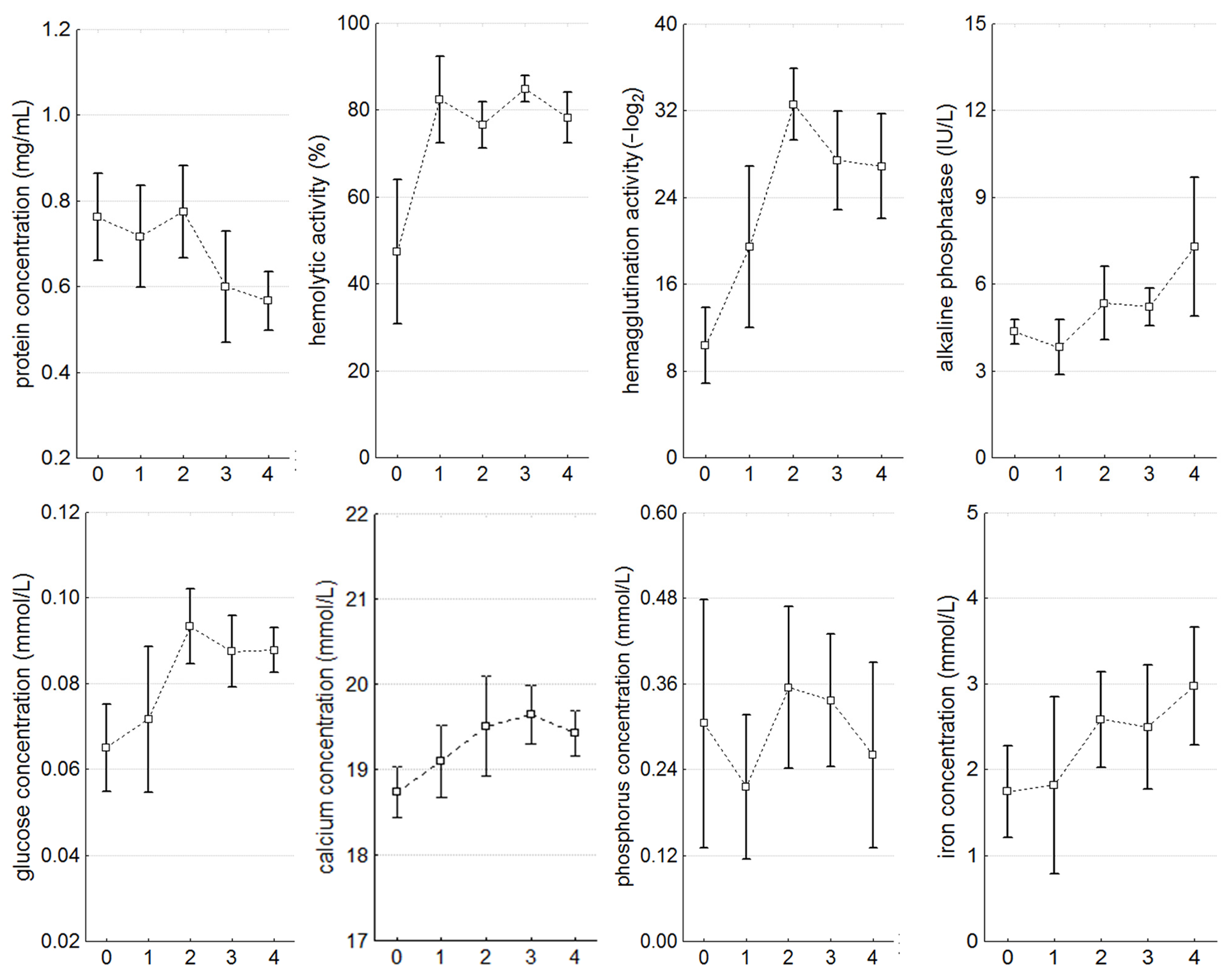
3.3. Hemolymph Parameters
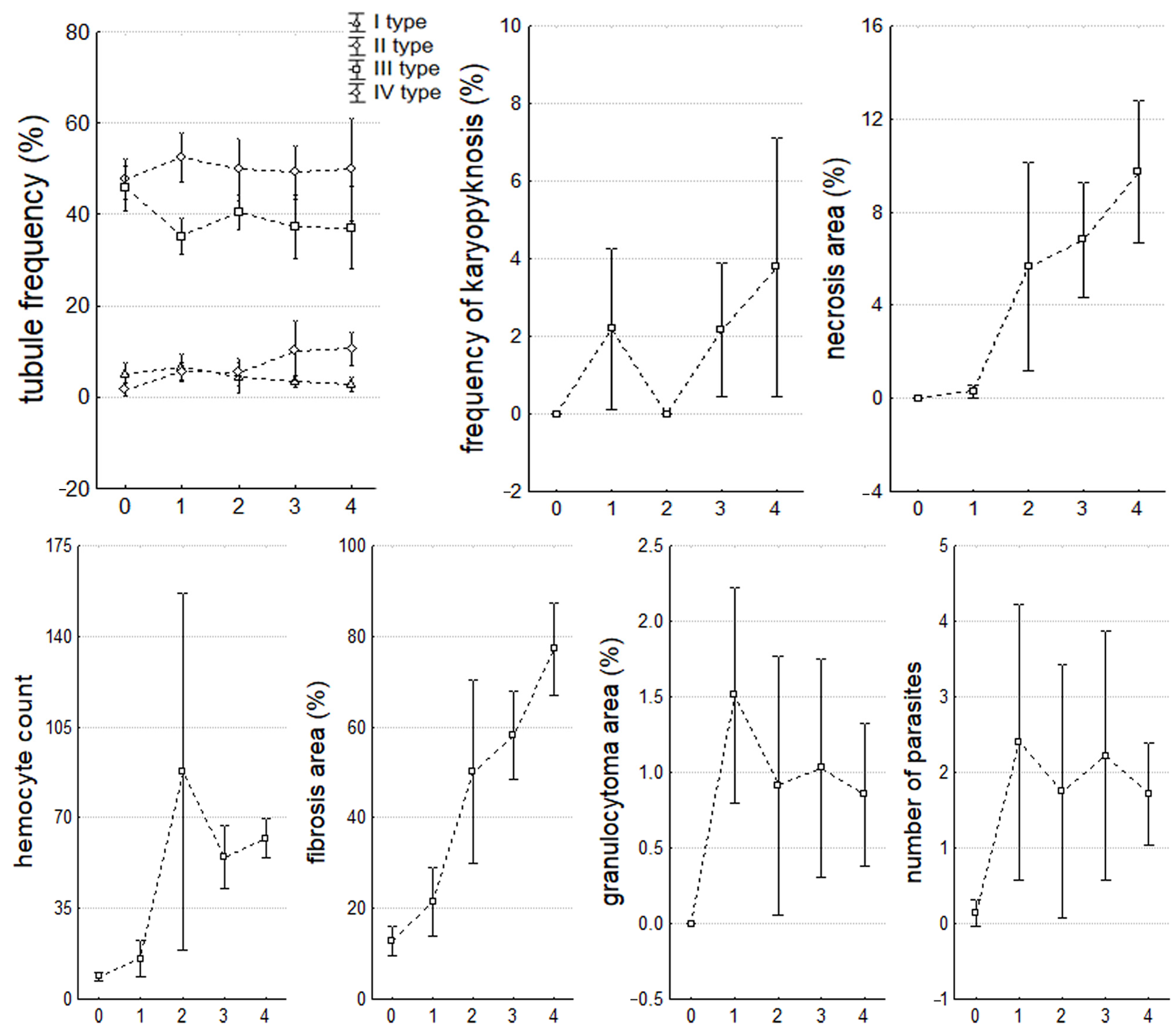
3.4. Histopathological State of the Digestive Gland and Nephridia
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hart, J.; Mooney, L.; Arthur, I.; Inglis, T.J.J.; Murray, R. First Case of Chlorella Wound Infection in a Human in Australia. New Microbes New Infect. 2014, 2, 132–133. [Google Scholar] [CrossRef]
- Jagielski, T.; Bakuła, Z.; Gawor, J.; Maciszewski, K.; Kusber, W.H.; Dyląg, M.; Nowakowska, J.; Gromadka, R.; Karnkowska, A. The Genus Prototheca (Trebouxiophyceae, Chlorophyta) Revisited: Implications from Molecular Taxonomic Studies. Algal Res. 2019, 43, 101639. [Google Scholar] [CrossRef]
- Riet-Correa, F.; Carmo, P.M.S.d.; Uzal, F.A. Protothecosis and Chlorellosis in Sheep and Goats: A Review. J. Vet. Diagn. Investig. 2021, 33, 283–287. [Google Scholar] [CrossRef]
- Fujimoto, M.; Inaba, Y.; Takahashi, T.; Nakanishi, G.; Muraosa, Y.; Yahiro, M.; Kamei, K.; Murata, S.I. Image Gallery: Granulomatous Dermatitis Due to Infection with the Chlorophyllic Green Alga Desmodesmus. Br. J. Dermatol. 2018, 179, e167. [Google Scholar] [CrossRef]
- Tartar, A. The Non-Photosynthetic Algae Helicosporidium spp.: Emergence of a Novel Group of Insect Pathogens. Insects 2013, 4, 375–391. [Google Scholar] [CrossRef] [PubMed]
- Sokolnikova, Y.; Magarlamov, T.; Stenkova, A.; Kumeiko, V. Permanent Culture and Parasitic Impact of the Microalga Coccomyxa Parasitica, Isolated from Horse Mussel Modiolus kurilensis. J. Invertebr. Pathol. 2016, 140, 25–34. [Google Scholar] [CrossRef]
- Sokolnikova, Y.; Tumas, A.; Stenkova, A.; Slatvinskaya, V.; Magarlamov, T.; Smagina, E. Novel Species of Parasitic Green Microalgae Coccomyxa veronica Sp. Nov. Infects Anadara broughtonii from the Sea of Japan. Symbiosis 2022, 87, 293–305. [Google Scholar] [CrossRef]
- Vaschenko, M.A.; Kovaleva, A.L.; Syasina, I.G.; Kukhlevsky, A.D. Reproduction-Related Effects of Green Alga Coccomyxa sp. Infestation in the Horse Mussel Modiolus modiolus. J. Invertebr. Pathol. 2013, 113, 86–95. [Google Scholar] [CrossRef]
- Sokolnikova, Y. Endobiotic Microalgae in Molluscan Life. Fish. Aquatic Sci. 2022, 25, 499–516. [Google Scholar] [CrossRef]
- Zuykov, M.; Anderson, J.; Pelletier, E. Does Photosynthesis Provoke Formation of Shell Deformity in Wild Mytilid Mussels Infested with Green Microalgae Coccomyxa?—A Conceptual Model and Research Agenda. J. Exp. Mar. Biol. Ecol. 2018, 505, 9–11. [Google Scholar] [CrossRef]
- Villalba, A.; Reece, K.S.; Ordás, M.C.; Casas, S.M.; Figueras, A. Perkinsosis in Molluscs: A Review. Aquat. Living Resour. 2004, 17, 411–432. [Google Scholar] [CrossRef]
- Itoh, N.; Komatsu, Y.; Maeda, K.; Hirase, S.; Yoshinaga, T. First Discovery of Perkinsus beihaiensis in Mediterranean Mussels (Mytilus galloprovincialis) in Tokyo Bay, Japan. J. Invertebr. Pathol. 2019, 166, 107226. [Google Scholar] [CrossRef]
- Muznebin, F.; Alfaro, A.C.; Webb, S.C. Perkinsus olseni and Other Parasites and Abnormal Tissue Structures in New Zealand Greenshell Mussels (Perna canaliculus) across Different Seasons. Aquac. Int. 2023, 31, 547–582. [Google Scholar] [CrossRef]
- Syasina, I.G.; Kukhlevsky, A.D.; Kovaleva, A.L.; Vaschenko, M.A. Phylogenetic and Morphological Characterization of the Green Alga Infesting the Horse Mussel Modiolus modiolus from Vityaz Bay (Peter the Great Bay, Sea of Japan). J. Invertebr. Pathol. 2012, 111, 175–181. [Google Scholar] [CrossRef]
- Mortensen, S.; Harkestad, L.S.; Stene, R.; Renault, T. Picoeucaryot Alga Infecting Blue Mussel Mytilus edulis in Southern Norway. Dis. Aquat. Organ. 2005, 63, 25–32. [Google Scholar] [CrossRef]
- Gray, A.P.; Lucas, I.A.N.; Seed, R.; Richardson, C.A. Mytilus edulis chilensis Infested with Coccomyxa parasitica (Chlorococcales, Coccomyxaceae). J. Molluscan Stud. 1999, 65, 289–294. [Google Scholar] [CrossRef]
- Vázquez, N.; Rodríguez, F.; Ituarte, C.; Klaich, J.; Cremonte, F. Host-Parasite Relationship of the Geoduck Panopea abbreviata and the Green Alga Coccomyxa parasitica in the Argentinean Patagonian Coast. J. Invertebr. Pathol. 2010, 105, 254–260. [Google Scholar] [CrossRef]
- Zuykov, M.; Allam, B.; Gosselin, M.; Archambault, P.; Spiers, G.; Schindler, M. First Report of Signs of Infection by Coccomyxa-like Algae in Wild Blue Mussels, Mytilus spp., in the Gulf of Maine (USA, Maine). J. Fish. Dis. 2020, 43, 775–778. [Google Scholar] [CrossRef]
- Zuykov, M.; Anderson, J.; Archambault, P.; Dufresne, F.; Pelletier, E. Mytilus trossulus and Hybrid (M. edulis-M. trossulus)-New Hosts Organisms for Pathogenic Microalgae Coccomyxa sp. from the Estuary and Northwestern Gulf of St. Lawrence, Canada. J. Invertebr. Pathol. 2018, 153, 145–146. [Google Scholar] [CrossRef]
- Lysenko, L.; Sukhovskaya, I.; Kantserova, N.; Bakhmet, I. Integrative Biomarker Indices in a Benthic Indicator Species Modiolus modiolus (L.) under a Simulated Oil Spill. KnE Soc. Sci. 2022, 2022, 83–92. [Google Scholar] [CrossRef]
- Bakhmet, I.N.; Sazhin, A.; Maximovich, N.; Ekimov, D. In Situ Long-Term Monitoring of Cardiac Activity of Two Bivalve Species from the White Sea, the Blue Mussel Mytilus edulis and Horse Mussel Modiolus Modiolus. J. Mar. Biol. Assoc. U. K. 2019, 99, 833–840. [Google Scholar] [CrossRef]
- Veldhoen, N.; Lowe, C.J.; Davis, C.; Mazumder, A.; Helbing, C.C. Gene Expression Profiling in the Deep Water Horse Mussel Modiolus modiolus (L.) Located near a Marine Municipal Wastewater Outfall. Aquat. Toxicol. 2009, 93, 116–124. [Google Scholar] [CrossRef]
- Podgurskaya, O.V.; Kavun, V.Y. Assessment of the Adaptation Capabilities of the Bivalves Modiolus modiolus (Linnaeus, 1758) and Crenomytilus grayanus (Dunker, 1853) under Increased Levels of Heavy Metals in the Environment. Russ. J. Mar. Biol. 2012, 38, 188–196. [Google Scholar] [CrossRef]
- Bourdelin, F. Physiological Responses of the Tropical Mussel, Modiolus aurieulatus—A Possible Biological Monitor in French Polynesia. Mar. Pollut. Bull. 1996, 32, 480–485. [Google Scholar] [CrossRef]
- Sedova, L.G.; Sokolenko, D.A. Resources and Structure of Horsemussel Modiolus kurilensis Settlements in Peter the Great Bay (The Sea of Japan). Mar. Biol. J. 2021, 6, 83–94. [Google Scholar] [CrossRef]
- Khristoforova, N.K.; Gnetetskiy, A.V. The Contents of Heavy Metals in Long-Lived Mytilids from Ussuriisky Bay. Russ. J. Mar. Biol. 2022, 48, 26–32. [Google Scholar] [CrossRef]
- Rodríguez, F.; Feist, S.W.; Guillou, L.; Harkestad, L.S.; Bateman, K.; Renault, T.; Mortensen, S. Phylogenetic and Morphological Characterisation of the Green Algae Infesting Blue Mussel Mytilus edulis in the North and South Atlantic Oceans. Dis. Aquat. Org. 2008, 81, 231–240. [Google Scholar] [CrossRef]
- Zuykov, M.; Belzile, C.; Lemaire, N.; Gosselin, M.; Dufresne, F.; Pelletier, E. First Record of the Green Microalgae Coccomyxa sp. in Blue Mussel Mytilus edulis (L.) from the Lower St. Lawrence Estuary (Québec, Canada). J. Invertebr. Pathol. 2014, 120, 23–32. [Google Scholar] [CrossRef]
- Crespo, C.; Rodríguez, H.; Segade, P.; Iglesias, R.; García-Estévez, J.M. Coccomyxa sp. (Chlorophyta: Chlorococcales), a New Pathogen in Mussels (Mytilus galloprovincialis) of Vigo Estuary (Galicia, NW Spain). J. Invertebr. Pathol. 2009, 102, 214–219. [Google Scholar] [CrossRef]
- Sokolnikova, Y.; Mokrina, M.; Magarlamov, T.; Grinchenko, A.; Kumeiko, V. Specification of Hemocyte Subpopulations Based on Immune-Related Activities and the Production of the Agglutinin MkC1qDC in the Bivalve Modiolus Kurilensis. Heliyon 2023, 9, e15577. [Google Scholar] [CrossRef] [PubMed]
- Grinchenko, A.; Sokolnikova, Y.; Korneiko, D.; Kumeiko, V. Dynamics of the Immune Response of the Horse Mussel Modiolus Kurilensis (Bernard, 1983) Following Challenge with Heat-Inactivated Bacteria. J. Shellfish Res. 2015, 34, 909–917. [Google Scholar] [CrossRef]
- Kumeiko, V.V.; Sokolnikova, Y.N.; Grinchenko, A.V.; Mokrina, M.S.; Kniazkina, M.I. Immune State Correlates with Histopathological Level and Reveals Molluscan Health in Populations of Modiolus kurilensis by Integral Health Index (IHI). J. Invertebr. Pathol. 2018, 154, 42–57. [Google Scholar] [CrossRef]
- Shinn, A.P.; Pratoomyot, J.; Bron, J.E.; Paladini, G.; Brooker, E.E.; Brooker, A.J. Economic Costs of Protistan and Metazoan Parasites to Global Mariculture. Parasitology 2015, 142, 196–270. [Google Scholar] [CrossRef]
- Carella, F.; Feist, S.W.; Bignell, J.P.; De Vico, G. Comparative Pathology in Bivalves: Aetiological Agents and Disease Processes. J. Invertebr. Pathol. 2015, 131, 107–120. [Google Scholar] [CrossRef]
- Paillard, C.; Gueguen, Y.; Wegner, M.; Bass, D.; Pallavicini, A.; Vezzulli, L.; Arzul, I. Recent Advances in Bivalve-Microbiota Interactions for Disease Prevention in Aquaculture. Curr. Opin. Biotechnol. 2022, 73, 225–232. [Google Scholar] [CrossRef]
- Zannella, C.; Mosca, F.; Mariani, F.; Franci, G.; Folliero, V.; Galdiero, M.; Tiscar, P.G.; Galdiero, M. Microbial Diseases of Bivalve Mollusks: Infections, Immunology and Antimicrobial Defense. Mar. Drugs 2017, 15, 182. [Google Scholar] [CrossRef]
- Catanese, G.; Grau, A.; Valencia, J.M.; Garcia-March, J.R.; Vázquez-Luis, M.; Alvarez, E.; Deudero, S.; Darriba, S.; Carballal, M.J.; Villalba, A. Haplosporidium pinnae sp. Nov., a Haplosporidan Parasite Associated with Mass Mortalities of the Fan Mussel, Pinna nobilis, in the Western Mediterranean Sea. J. Invertebr. Pathol. 2018, 157, 9–24. [Google Scholar] [CrossRef]
- Carnegie, R.B.; Barber, B.J.; Distel, D.L. Detection of the Oyster Parasite Bonamia ostreae by Fluorescent in Situ Hybridization. Dis. Aquat. Organ. 2003, 55, 247–252. [Google Scholar] [CrossRef]
- Gestal, C.; Roch, P.; Renault, T.; Pallavicini, A.; Paillard, C.; Novoa, B.; Oubella, R.; Venier, P.; Figueras, A. Study of Diseases and the Immune System of Bivalves Using Molecular Biology and Genomics. Rev. Fish. Sci. 2008, 16, 131–154. [Google Scholar] [CrossRef]
- Allam, B.; Espinosa, P. Mucosal Immunity in Mollusks. In Mucosal Health in Aquaculture; Beck Benjamin, H., Peatman, E., Eds.; Academic Press: London, UK, 2015; pp. 325–370. [Google Scholar]
- Villalba, A.; Mourelle, S.G.; Carballal, M.J.; López, C. Symbionts and Diseases of Farmed Mussels Mytilus galloprovincialis throughout the Culture Process in the Rias of Galicia (NW Spain). Dis. Aquat. Organ. 1997, 31, 127–139. [Google Scholar] [CrossRef]
- Auffret, M. Bivalve Hemocyte Morphology. In Disease Processes in Marine Bivalve Molluscs; Fisher, W.S., Ed.; American Fisheries Society: New York, NY, USA, 1988; pp. 169–177. [Google Scholar]
- Da Silva, P.M.; Hégaret, H.; Lambert, C.; Wikfors, G.H.; Le Goïc, N.; Shumway, S.E.; Soudant, P. Immunological Responses of the Manila Clam (Ruditapes philippinarum) with Varying Parasite (Perkinsus olseni) Burden, during a Long-Term Exposure to the Harmful Alga, Karenia selliformis, and Possible Interactions. Toxicon 2008, 51, 563–573. [Google Scholar] [CrossRef]
- Burreson, E.M.; Reece, K.S.; Dungan, C.F. Molecular, Morphological, and Experimental Evidence Support the Synonymy of Perkinsus chesapeaki and Perkinsus andrewsi. J. Eukaryot. Microbiol. 2005, 52, 258–270. [Google Scholar] [CrossRef]
- Arzul, I.; Chollet, B.; Michel, J.; Robert, M.; Garcia, C.; Joly, J.P.; François, C.; Miossec, L. One Perkinsus Species May Hide Another: Characterization of Perkinsus Species Present in Clam Production Areas of France. Parasitology 2012, 139, 1757–1771. [Google Scholar] [CrossRef]
- Sheppard, B.J.; Phillips, A.C. Perkinsus Olseni Detected in Vietnamese Aquacultured Reef Clams Tridacna crocea Imported to the USA, Following a Mortality Event. Dis. Aquat. Organ. 2008, 79, 229–235. [Google Scholar] [CrossRef]
- Fernández-Boo, S.; Gervais, O.; Prado-Alvarez, M.; Chollet, B.; Claverol, S.; Lecadet, C.; Dubreuil, C.; Arzul, I. Is Pallial Mucus Involved in Ostrea edulis Defenses against the Parasite Bonamia ostreae? J. Invertebr. Pathol. 2020, 169, 107259. [Google Scholar] [CrossRef]
- Lane, H.S.; Webb, S.C.; Duncan, J. Bonamia ostreae in the New Zealand Oyster Ostrea chilensis: A New Host and Geographic Record for This Haplosporidian Parasite. Dis. Aquat. Organ. 2016, 118, 55–63. [Google Scholar] [CrossRef]
- Kleeman, S.N.; Adlard, R.D. Molecular Detection of Marteilia sydneyi, Pathogen of Sydney Rock Oysters. Dis. Aquat. Organ. 2000, 40, 137–146. [Google Scholar] [CrossRef]
- Lynch, S.A.; Lepée-Rivero, S.; Kelly, R.; Quinn, E.; Coghlan, A.; Bookelaar, B.; Morgan, E.; Finarelli, J.A.; Carlsson, J.; Culloty, S.C. Detection of Haplosporidian Protistan Parasites Supports an Increase to Their Known Diversity, Geographic Range and Bivalve Host Specificity. Parasitology 2020, 147, 584–592. [Google Scholar] [CrossRef]
- Zuykov, M.; Kolyuchkina, G.; Archambault, P.; Gosselin, M.; Anderson, J.; McKindsey, C.W.; Spiers, G.; Schindler, M. Shell Deformity as a Marker for Retrospective Detection of a Pathogenic Unicellular Alga, Coccomyxa sp., in Mytilid Mussels: A First Case Study and Research Agenda. J. Invertebr. Pathol. 2020, 169, 107311. [Google Scholar] [CrossRef]
- Cáceres-Martínez, J.; Vásquez-Yeomans, R.; Padilla-Lardizábal, G.; del Río Portilla, M.A. Perkinsus marinus in Pleasure Oyster Crassostrea corteziensis from Nayarit, Pacific Coast of México. J. Invertebr. Pathol. 2008, 99, 66–73. [Google Scholar] [CrossRef]
- McLaughlin, S.M.; Faisal, M. Histopathological Alterations Associated with Perkinsus spp. Infection in the Softshell Clam Mya arenaria. Parasite 1998, 5, 263–271. [Google Scholar] [CrossRef]
- Dang, C.; Dungan, C.F.; Scott, G.P.; Reece, K.S. Perkinsus sp. Infections and in Vitro Isolates from Anadara trapezia (Mud Arks) of Queensland, Australia. Dis. Aquat. Organ. 2015, 113, 51–58. [Google Scholar] [CrossRef]
- Karnisa, Y.; Desrina, D.; Widowati, I. Parasites Identification and Histopathology Changes on Blood Cookle (Anadara granosa Linnaeus, 1758). Ilmu Kelaut. 2019, 24, 171–178. [Google Scholar] [CrossRef]
- Casas, S.M. Estudio de La Perkinsosis En La Almeja Fina, Tapes decussatus (Linnaeus, 1758), de Galicia. Ph.D. Thesis, University of Santiago de Compostela, Santiago de Compostela, Spain, 2002. [Google Scholar]
- Kolyuchkina, G.A.; Ismailov, A.D. Morpho-Functional Characteristics of Bivalve Mollusks under the Experimental Environmental Pollution by Heavy Metals. Oceanology 2011, 51, 804–813. [Google Scholar] [CrossRef]
- Aarab, N.; Godal, B.F.; Bechmann, R.K. Seasonal Variation of Histopathological and Histochemical Markers of PAH Exposure in Blue Mussel (Mytilus edulis L.). Mar. Environ. Res. 2011, 71, 213–217. [Google Scholar] [CrossRef]
- Barmo, C.; Ciacci, C.; Canonico, B.; Fabbri, R.; Cortese, K.; Balbi, T.; Marcomini, A.; Pojana, G.; Gallo, G.; Canesi, L. In Vivo Effects of N-TiO2 on Digestive Gland and Immune Function of the Marine Bivalve Mytilus galloprovincialis. Aquat. Toxicol. 2013, 132–133, 9–18. [Google Scholar] [CrossRef]
- Boehs, G.; Villalba, A.; Ceuta, L.O.; Luz, J.R. Parasites of Three Commercially Exploited Bivalve Mollusc Species of the Estuarine Region of the Cachoeira River (Ilhéus, Bahia, Brazil). J. Invertebr. Pathol. 2010, 103, 43–47. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, R.K.; Das, S.; Das, V.K. Pathological Changes in Hepatopancreas of Freshwater Mussel (Lamellidens marginalis Lamarck) Exposed to Sub-Lethal Concentration of Dimethoate. GERF Bull. Biosci. 2011, 2, 18–23. [Google Scholar]
- Larguinho, M.; Cordeiro, A.; Diniz, M.S.; Costa, P.M.; Baptista, P.V. Metabolic and Histopathological Alterations in the Marine Bivalve Mytilus galloprovincialis Induced by Chronic Exposure to Acrylamide. Environ. Res. 2014, 135, 55–62. [Google Scholar] [CrossRef]
- Mantecca, P.; Vailati, G.; Bacchetta, R. Histological Changes and Micronucleus Induction in the Zebra Mussel Dreissena polymorpha after Paraquat Exposure. Histol. Histopathol. 2006, 21, 829–840. [Google Scholar] [CrossRef]
- Ward, M.E.; Shields, J.D.; Van Dover, C.L. Parasitism in Species of Bathymodiolus (Bivalvia: Mytilidae) Mussels from Deep-Sea Seep and Hydrothermal Vents. Dis. Aquat. Organ. 2004, 62, 1–16. [Google Scholar] [CrossRef]
- Moëzzi, F.; Javanshir, A.; Eagderi, S.; Poorbagher, H. Histopathological Effects of Zinc (Zn) on Mantle, Digestive Gland and Foot in Freshwater Mussel, Anodonta cygnea (Linea, 1876). Int. J. Aquat. Biol. 2013, 1, 61–67. [Google Scholar] [CrossRef]
- Wang, K.; Pales Espinosa, E.; Tanguy, A.; Allam, B. Alterations of the Immune Transcriptome in Resistant and Susceptible Hard Clams (Mercenaria mercenaria) in Response to Quahog Parasite Unknown (QPX) and Temperature. Fish Shellfish Immunol. 2016, 49, 163–176. [Google Scholar] [CrossRef]
- Sheir, S.K.; Handy, R.D. Tissue Injury and Cellular Immune Responses to Cadmium Chloride Exposure in the Common Mussel Mytilus Edulis: Modulation by Lipopolysaccharide. Arch. Environ. Contam. Toxicol. 2010, 59, 602–613. [Google Scholar] [CrossRef]
- Smolowitz, R. A Review of Current State of Knowledge Concerning Perkinsus marinus Effects on Crassostrea virginica (Gmelin) (the Eastern Oyster). Vet. Pathol. 2013, 50, 404–411. [Google Scholar] [CrossRef]
- Galimany, E.; Sunila, I.; Hégaret, H.; Ramón, M.; Wikfors, G.H. Pathology and Immune Response of the Blue Mussel (Mytilus edulis L.) after an Exposure to the Harmful Dinoflagellate Prorocentrum minimum. Harmful Algae 2008, 7, 630–638. [Google Scholar] [CrossRef]
- Au, D.W.T. The Application of Histo-Cytopathological Biomarkers in Marine Pollution Monitoring: A Review. Mar. Pollut. Bull. 2004, 48, 817–834. [Google Scholar] [CrossRef] [PubMed]
- Meyhöfer, E.; Morse, M.P.; Robinson, W.E. Podocytes in Bivalve Molluscs: Morphological Evidence for Ultrafiltration. J. Comp. Physiol. B 1985, 156, 151–161. [Google Scholar] [CrossRef]
- George, S.G. Heavy Metal Detoxication in Mytilus Kidney—An In Vitro Study of Cd- and Zn-Binding to Isolated Tertiary Lysosomes. Comp. Biochem. Physiol. C Comp. Pharmacol. Toxicol. 1983, 76, 59–65. [Google Scholar] [CrossRef]
- Da Silva, P.M.; Magalhães, A.R.M.; Barracco, M.A. Pathologies in Commercial Bivalve Species from Santa Catarina State, Southern Brazil. J. Mar. Biol. Assoc. U. K. 2012, 92, 571–579. [Google Scholar] [CrossRef]
- Chong, R.S.-M. Kidney Coccidiosis (Scallop, Abalone, Mussels, Oysters, and Clams). In Aquaculture Pathophysiology; Kibenge Frederick, S.B., Bernardo, B., Sie-Maen, C.R., Eds.; Academic Press: London, UK, 2022; Volume 2, pp. 551–553. ISBN 9780323954341. [Google Scholar]
- Pales Espinosa, E.; Bouallegui, Y.; Grouzdev, D.; Brianik, C.; Czaja, R.; Geraci-Yee, S.; Kristmundsson, A.; Muehl, M.; Schwaner, C.; Tettelbach, S.T.; et al. An Apicomplexan Parasite Drives the Collapse of the Bay Scallop Population in New York. Sci. Rep. 2023, 13, 6655. [Google Scholar] [CrossRef] [PubMed]
- Maeno, Y.; Suzuki, K.; Yurimoto, T.; Kiyomoto, S.; Fuseya, R.; Fujisaki, H.; Yoshida, M.; Nasu, H. Laboratory and Field Studies on Gill and Kidney Associated Virus in the Pen Shell Atrina lischkeana (Mollusca: Bivalvia). Bull. Eur. Assoc. Fish. Pathol. 2012, 32, 78–86. [Google Scholar]
- Belzile, C.; Gosselin, M. Free-Living Stage of the Unicellular Algae Coccomyxa sp. Parasite of the Blue Mussel (Mytilus edulis): Low-Light Adaptation, Capacity for Growth at a Very Wide Salinity Range and Tolerance to Low PH. J. Invertebr. Pathol. 2015, 132, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Ronza, P.; Cao, A.; Robledo, D.; Gómez-Tato, A.; Álvarez-Dios, J.A.; Hasanuzzaman, A.F.M.; Quiroga, M.I.; Villalba, A.; Pardo, B.G.; Martínez, P. Long-Term Affected Flat Oyster (Ostrea edulis) Haemocytes Show Differential Gene Expression Profiles from Naïve Oysters in Response to Bonamia Ostreae. Genomics 2018, 110, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Garbayo, I.; Torronteras, R.; Forján, E.; Cuaresma, M.; Casal, C.; Mogedas, B.; Ruiz-Domínguez, M.C.; Márquez, C.; Vaquero, I.; Fuentes-Cordero, J.L.; et al. Identification and Physiological Aspects of a Novel Carotenoid-Enriched, Metal-Resistant Microalga Isolated from an Acidic River in Huelva (Spain). J. Phycol. 2012, 48, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Vaquero, I.; Mogedas, B.; Ruiz-Domínguez, M.C.; Vega, J.M.; Vílchez, C. Light-Mediated Lutein Enrichment of an Acid Environment Microalga. Algal Res. 2014, 6, 70–77. [Google Scholar] [CrossRef]
- Sun, L.; Jin, Y.; Dong, L.; Sumi, R.; Jahan, R.; Li, Z. The Neuroprotective Effects of Coccomyxa gloeobotrydiformis on the Ischemic Stroke in a Rat Model. Int. J. Biol. Sci. 2013, 9, 811–817. [Google Scholar] [CrossRef]
- Ruiz-Domínguez, M.C.; Vaquero, I.; Obregón, V.; de la Morena, B.; Vílchez, C.; Vega, J.M. Lipid Accumulation and Antioxidant Activity in the Eukaryotic Acidophilic Microalga Coccomyxa sp. (Strain Onubensis) under Nutrient Starvation. J. Appl. Phycol. 2015, 27, 1099–1108. [Google Scholar] [CrossRef]
- Soudant, P.; Chu, F.L.E.; Volety, A. Host–Parasite Interactions: Marine Bivalve Molluscs and Protozoan Parasites, Perkinsus Species. J. Invertebr. Pathol. 2013, 114, 196–216. [Google Scholar] [CrossRef]
- Lau, Y.T.; Santos, B.; Barbosa, M.; Pales Espinosa, E.; Allam, B. Regulation of Apoptosis-Related Genes during Interactions between Oyster Hemocytes and the Alveolate Parasite Perkinsus marinus. Fish Shellfish Immunol. 2018, 83, 180–189. [Google Scholar] [CrossRef]
- Zhang, H.; Song, L.; Li, C.; Zhao, J.; Wang, H.; Qiu, L.; Ni, D.; Zhang, Y. A Novel C1q-Domain-Containing Protein from Zhikong Scallop Chlamys farreri with Lipopolysaccharide Binding Activity. Fish Shellfish Immunol. 2008, 25, 281–289. [Google Scholar] [CrossRef] [PubMed]
- La Peyre, J.F.; Chu, F.-L.E.; Meyers, J.M. Haemocytic and Humoral Activities of Eastern and Pacific Oysters Following Challenge by the Protozoan Perkinsus marinus. Fish Shellfish Immunol. 1995, 5, 179–190. [Google Scholar] [CrossRef]
- Ordas, M.C.; Ordas, A.; Beloso, C.; Figueras, A. Immune Parameters in Carpet Shell Clams Naturally Infected with Perkinsus Atlanticus. Fish Shellfish Immunol. 2000, 10, 597–609. [Google Scholar] [CrossRef]
- Olafsen, J.A.; Fletcher, T.C.; Grant, P.T. Agglutinin Activity in Pacific Oyster (Crassostrea gigas) Hemolymph Following in Vivo Vibrio anguillarum Challenge. Dev. Comp. Immunol. 1992, 16, 123–138. [Google Scholar] [CrossRef]
- Garcia, A.; Estêvão, J.; Costas, B.; Cruz, A.; Fernández-Boo, S. Evaluation of the Ruditapes decussatus Immune Response after Differential Injected Doses of Perkinsus olseni. J. Invertebr. Pathol. 2022, 195, 107849. [Google Scholar] [CrossRef] [PubMed]
- Hughes, F.M.; Foster, B.; Grewal, S.; Sokolova, I.M. Apoptosis as a Host Defense Mechanism in Crassostrea virginica and Its Modulation by Perkinsus marinus. Fish Shellfish Immunol. 2010, 29, 247–257. [Google Scholar] [CrossRef]
- Witkop, E.M.; Wikfors, G.H.; Proestou, D.A.; Lundgren, K.M.; Sullivan, M.; Gomez-Chiarri, M. Perkinsus marinus Suppresses in Vitro Eastern Oyster Apoptosis via IAP-Dependent and Caspase-Independent Pathways Involving TNFR, NF-KB, and Oxidative Pathway Crosstalk. Dev. Comp. Immunol. 2022, 129, 104339. [Google Scholar] [CrossRef]
- Gervais, O.; Renault, T.; Arzul, I. Molecular and Cellular Characterization of Apoptosis in Flat Oyster a Key Mechanisms at the Heart of Host-Parasite Interactions. Sci. Rep. 2018, 8, 12494. [Google Scholar] [CrossRef]
- Leggat, W.; Buck, B.H.; Grice, A.; Yellowlees, D. The Impact of Bleaching on the Metabolic Contribution of Dinoflagellate Symbionts to Their Giant Clam Host. Plant Cell Environ. 2003, 26, 1951–1961. [Google Scholar] [CrossRef]
- Ishikura, M.; Adachi, K.; Maruyama, T. Zooxanthellae Release Glucose in the Tissue of a Giant Clam, Tridacna crocea. Mar. Biol. 1999, 133, 665–673. [Google Scholar] [CrossRef]
- Nash, I.H. Lichen Biology, 2nd ed.; Nash, I.H., Ed.; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Richardson, D.H.S.; Jackson, D.; Smith, D.C. Lichen Physiology. XI. The Role of the Alga in Determining the Pattern of Carbohydrate Movement between Lichen Symbionts. New Phytol. 1968, 67, 469–486. [Google Scholar] [CrossRef]
- Grigoryeva, N.I.; Zhuravel, E.V.; Mazur, A.A. Seasonal Variations of Water Quality in the Vostok Bay, Peter the Great Gulf, the Sea of Japan. Water Resour. 2020, 47, 249–256. [Google Scholar] [CrossRef]
- Khristoforova, N.K.; Boychenko, T.V.; Kobzar, A.D. Hydrochemical and Microbiological Assessment of the Current State of the Vostok Bay. Bull. Far East. Branch Russ. Acad. Sci. 2020, 2, 64–72. [Google Scholar] [CrossRef]
- Galysheva, Y.A.; Khristoforova, N.K. Environments and Macrobenthos in the Vostok Bay (Japan Sea) in Conditions of Anthropogenic Impact. Izvestiya TINRO 2007, 149, 270–309. [Google Scholar]
- Vernon, E.L.; Smith, J.T.; Jha, A.N. Relative Comparison of Tissue Specific Bioaccumulation and Radiation Dose Estimation in Marine and Freshwater Bivalve Molluscs Following Exposure to Phosphorus-32. J. Environ. Radioact. 2018, 192, 312–320. [Google Scholar] [CrossRef]
- Lyubas, A.A.; Tomilova, A.A.; Chupakov, A.V.; Vikhrev, I.V.; Travina, O.V.; Orlov, A.S.; Zubrii, N.A.; Kondakov, A.V.; Bolotov, I.N.; Pokrovsky, O.S. Iron, Phosphorus and Trace Elements in Mussels’ Shells, Water, and Bottom Sediments from the Severnaya Dvina and the Onega River Basins (Northwestern Russia). Water 2021, 13, 3227. [Google Scholar] [CrossRef]
- Shen, C.; Liao, Q.; Bootsma, H.A. Modelling the Influence of Invasive Mussels on Phosphorus Cycling in Lake Michigan. Ecol. Model. 2020, 416, 108920. [Google Scholar] [CrossRef]
- Wang, X.; Li, C.; Lv, Z.; Zhang, Z.; Qiu, L. A Calcification-Related Calmodulin-like Protein in the Oyster Crassostrea Gigas Mediates the Enhanced Calcium Deposition Induced by CO2 Exposure. Sci. Total Environ. 2022, 833, 155114. [Google Scholar] [CrossRef]
- Marchant, H.K.; Calosi, P.; Spicer, J.I. Short-Term Exposure to Hypercapnia Does Not Compromise Feeding, Acid-Base Balance or Respiration of Patella vulgata but Surprisingly Is Accompanied by Radula Damage. J. Mar. Biol. Assoc. U. K. 2010, 90, 1379–1384. [Google Scholar] [CrossRef]
- Parker, L.M.; Ross, P.M.; O’Connor, W.A.; Pörtner, H.O.; Scanes, E.; Wright, J.M. Predicting the Response of Molluscs to the Impact of Ocean Acidification. Biology 2013, 2, 651–692. [Google Scholar] [CrossRef]
- Wang, X.; Wang, M.; Wang, W.; Liu, Z.; Xu, J.; Jia, Z.; Chen, H.; Qiu, L.; Lv, Z.; Wang, L.; et al. Transcriptional Changes of Pacific Oyster Crassostrea gigas Reveal Essential Role of Calcium Signal Pathway in Response to CO2-Driven Acidification. Sci. Total Environ. 2020, 741, 140177. [Google Scholar] [CrossRef]
- Thomsen, J.; Ramesh, K.; Sanders, T.; Bleich, M.; Melzner, F. Calcification in a Marginal Sea-Influence of Seawater [Ca2+] and Carbonate Chemistry on Bivalve Shell Formation. Biogeosciences 2018, 15, 1469–1482. [Google Scholar] [CrossRef]
- Burreson, E.M.; Ford, S.E. A Review of Recent Information on the Haplosporidia, with Special Reference to Haplosporidium nelsoni (MSX Disease). Aquat. Living Resour. 2004, 17, 499–517. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Significance Level, p | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Healthy–Infested | 0–1 | 0–2 | 0–3 | 0–4 | 1–2 | 1–3 | 1–4 | 2–3 | 2–4 | 3–4 | |
| percentage of dead cells (%) | NS | NS | NS | 0.016 * | 0.009 * | NS | 0.001 * | 0.001 * | 0.000 * | 0.000 * | NS |
| percentage of basophils (%) | 0.032 * | NS | 0.050 * | NS | 0.008 * | NS | NS | NS | NS | NS | NS |
| percentage of eosinophils (%) | 0.050 * | NS | NS | 0.042 * | NS | NS | 0.008 * | 0.012 * | 0.028 * | 0.035 * | NS |
| percentage of hyalinocytes (%) | 0.000 * | NS | 0.000 * | 0.001 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | NS | NS | 0.016 * |
| percentage of cells with ROS (%) | 0.000 * | NS | 0.002 * | 0.000 * | 0.000 * | NS | NS | 0.011 * | NS | NS | NS |
| percentage of cells with AO (%) | 0.001 * | NS | 0.002 * | 0.007 * | 0.000 * | 0.004 * | 0.011 * | 0.000 * | NS | NS | 0.050 * |
| phagocytic index of bacteria | 0.000 * | NS | 0.000 * | 0.002 * | 0.000 * | 0.023 * | 0.020 * | 0.018 * | NS | NS | NS |
| phagocytic index of algae | 0.000 * | 0.000 * | 0.001 * | 0.000 * | 0.000 * | NS | 0.029 * | 0.046 * | 0.009 * | 0.044 * | NS |
| percentage of cells with algae (%) | 0.000 * | 0.000 * | 0.001 * | 0.000 * | 0.000 * | NS | 0.029 * | 0.001 * | 0.000 * | 0.000 * | 0.000 * |
| hemolytic activity (%) | 0.000 * | 0.006 * | 0.008 * | 0.000 * | 0.021 * | NS | NS | NS | 0.032 * | NS | 0.029 * |
| hemagglutination activity (−log2) | 0.000 * | NS | 0.000 * | 0.000 * | 0.000 * | 0.011 * | NS | NS | NS | NS | NS |
| glucose concentration (mmol/L) | 0.001 * | NS | 0.001 | 0.002 * | 0.000 * | 0.010 * | 0.021 * | 0.004 * | NS | NS | NS |
| calcium concentration (mmol/L) | 0.004 * | NS | NS | 0.002 * | 0.003 * | NS | NS | NS | NS | NS | NS |
| Digestive Gland Parameters | Significance Level, p | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Healthy–Infested | 0–1 | 0–2 | 0–3 | 0–4 | 1–2 | 1–3 | 1–4 | 2–3 | 2–4 | 3–4 | |
| percentage of IV type tubules (%) | 0.00 * | 0.00 * | 0.05 * | 0.02 * | 0.00 * | NS | NS | 0.03 * | NS | 0.05 * | NS |
| fibrosis area in the IS (%) | 0.00 * | NS | 0.02 * | 0.00 * | 0.00 * | NS | 0.00 * | 0.00 * | NS | NS | 0.03 * |
| hemocyte count in the IS | 0.00 * | NS | 0.01 * | 0.00 * | 0.00 * | 0.03 * | 0.00 * | 0.00 * | NS | NS | NS |
| parasite number | 0.01 * | NS | NS | NS | 0.00 * | NS | NS | NS | NS | NS | NS |
| necrosis area (%) | 0.00 * | NS | 0.00 * | 0.00 * | 0.00 * | 0.01 * | 0.00 * | 0.00 * | NS | 0.05 * | NS |
| frequency of kariopiknosis (%) | 0.05 * | NS | - | 0.04 * | 0.01 * | NS | NS | NS | 0.05 * | 0.01 * | NS |
| granulocytoma area (%) | 0.00 * | 0.00 * | 0.01 * | 0.00 * | 0.00 * | NS | NS | NS | NS | NS | NS |
| Nephridium Parameters | |||||||||||
| max. membrane thickness (µm) | NS | NS | NS | 0.02 * | 0.01 * | NS | NS | NS | 0.00 * | NS | 0.00 * |
| membrane thickness deviation (µm) | 0.00 * | 0.00 * | 0.00 * | 0.00 * | 0.00 * | NS | NS | NS | NS | NS | NS |
| epithelial roundness | 0.01 * | NS | 0.05 * | NS | 0.00 * | NS | NS | 0.03 * | NS | 0.04 * | 0.01 * |
| concretions number | 0.00 * | NS | 0.05 * | 0.01 * | 0.00 * | NS | NS | NS | NS | NS | 0.04 * |
| concretions roundness | 0.00 * | NS | 0.00 * | 0.01 * | 0.04 * | NS | NS | NS | NS | NS | NS |
| concretions density | 0.05 * | NS | 0.09 | 0.05 * | 0.00 * | 0.02 * | 0.01 * | 0.00 * | NS | NS | NS |
| parasite number | NS | NS | NS | NS | 0.02 * | - | NS | 0.01 * | NS | 0.02 * | NS |
| necrosis area (%) | 0.01 * | 0.02 * | NS | NS | NS | NS | NS | 0.01 * | NS | NS | NS |
| fibrosis area in the IS (%) | 0.00 * | 0.00 * | 0.00 * | 0.00 * | 0.00 * | NS | 0.01 * | 0.00 * | NS | 0.03 * | NS |
| hemocyte count in the IS | 0.00 * | NS | 0.03 * | 0.04 * | 0.00 * | NS | NS | 0.01 * | NS | NS | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tumas, A.V.; Slatvinskaya, V.A.; Kumeiko, V.V.; Sokolnikova, Y.N. Study of the Impact of the Parasitic Microalgae Coccomyxa parasitica on the Health of Bivalve Modiolus kurilensis. Microorganisms 2024, 12, 997. https://doi.org/10.3390/microorganisms12050997
Tumas AV, Slatvinskaya VA, Kumeiko VV, Sokolnikova YN. Study of the Impact of the Parasitic Microalgae Coccomyxa parasitica on the Health of Bivalve Modiolus kurilensis. Microorganisms. 2024; 12(5):997. https://doi.org/10.3390/microorganisms12050997
Chicago/Turabian StyleTumas, Ayna V., Veronika A. Slatvinskaya, Vadim V. Kumeiko, and Yulia N. Sokolnikova. 2024. "Study of the Impact of the Parasitic Microalgae Coccomyxa parasitica on the Health of Bivalve Modiolus kurilensis" Microorganisms 12, no. 5: 997. https://doi.org/10.3390/microorganisms12050997
APA StyleTumas, A. V., Slatvinskaya, V. A., Kumeiko, V. V., & Sokolnikova, Y. N. (2024). Study of the Impact of the Parasitic Microalgae Coccomyxa parasitica on the Health of Bivalve Modiolus kurilensis. Microorganisms, 12(5), 997. https://doi.org/10.3390/microorganisms12050997